
Abstract
Polysaccharides are structurally diverse biopolymers that interact intimately with the colonic mucus barrier, modulating its physicochemical properties and biological functions. This review explores direct and indirect interactions between polysaccharides and colonic mucus that collectively shape mucus rheology, penetrability, and barrier function. After brief, separate descriptions of colonic mucus (bilayered architecture and goblet cell–driven secretion) and polysaccharide structural classes (e.g., charge, molecular weight, branching, and substitution patterns), we dissect the molecular and biophysical mechanisms governing polysaccharide–mucus interactions—including electrostatic and hydrogen bonding, hydrophobic association, and steric or depletion effects—and integrate evidence from in vitro, ex vivo, in vivo, organoid, and gut-on-a-chip models. Functional consequences for mucin secretion and degradation, mucus viscosity and structure, barrier integrity, and epithelial and immune signalling are critically evaluated, with particular focus on protection against oxidative stress-induced mucosal dysfunction. By linking specific physicochemical features of polysaccharides to their behaviour within the mucus barrier, we outline design principles for improving gastrointestinal health, optimising mucosal drug delivery, and guiding the development of next-generation polysaccharide-based therapeutics.
Similar content being viewed by others

Introduction
Polysaccharides represent a highly heterogeneous group of natural macromolecules, constructed from monosaccharide residues linked through glycosidic bonds. Their architectures can range from linear chains to extensively branched configurations, resulting in diverse physicochemical behaviors that are dictated by factors such as monosaccharide composition, types of glycosidic linkages, molecular weight, and the presence of various functional moieties1,2. Beyond their conventional structural roles in the cell walls of plants, fungi, and microorganisms, polysaccharides have garnered attention for their extensive biological functionalities—including antioxidant, immunoregulatory, prebiotic, and anti-inflammatory activities3,4,5. These bioactivities position them as both valuable dietary components and promising candidates for therapeutic applications, particularly in the context of gastrointestinal health6,7.
The colonic mucus layer functions as a specialized, viscoelastic hydrogel, integral to the maintenance of gut equilibrium. It exhibits a bilayered architecture: an inner, dense layer that is typically devoid of bacteria and adheres closely to the epithelium, and an outer, loosely organized layer that provides a habitat for commensal microorganisms8. Mucins—particularly the gel-forming mucin MUC2, secreted by goblet cells—constitute the principal structural elements of this matrix9. Upon release, MUC2 undergoes extensive hydration and polymerization, leading to the formation of a structured gel that defines the barrier’s rheological properties10. The coordinated secretion and post-translational modification of mucins are essential for maintaining the mechanical and functional characteristics of the mucus layer11. In recent years, growing interest has focused on how exogenous and dietary polysaccharides interact with this mucus system to influence gastrointestinal physiology. These interactions have been implicated in modulating mucus secretion, barrier architecture, immunological communication, and overall gut function12. A deeper understanding of these processes offers significant promise for developing strategies aimed at enhancing mucosal protection, facilitating targeted drug delivery, and ameliorating intestinal disorders such as inflammatory bowel disease, infections, and metabolic syndromes.
The behavior of polysaccharides within the colonic mucus environment is largely governed by their physicochemical characteristics, including net surface charge, hydrophilic–hydrophobic balance, molecular weight, and conformational dynamics. These parameters dictate whether a polysaccharide will penetrate the mucus network, adhere to its components, or become immobilized within its mesh. For example, anionic or cationic polysaccharides may engage in electrostatic interactions with the negatively charged mucin glycans, whereas hydrophobic segments can drive binding through van der Waals attractions or hydrophobic clustering13,14. These modes of interaction carry important ramifications across several domains. In oral drug delivery, most small-molecule drugs are absorbed in the proximal small intestine, where polysaccharides and polysaccharide-based systems are especially relevant for colon-targeted formulation15. As prebiotics, they selectively promote the growth of beneficial microorganisms, thereby shaping mucus-associated microbiota profiles16. In the context of mucosal healing and immune regulation, polysaccharides have been shown to enhance mucin production, fortify epithelial integrity, and modulate inflammatory signaling, contributing to improved barrier resilience17. Additionally, their intrinsic antioxidant and anti-inflammatory capacities enable them to counteract oxidative insults that compromise mucus structure and function, particularly in disease states such as colitis and microbial dysbiosis18,19. Hence, elucidating how distinct physicochemical traits influence polysaccharide–mucus interactions is fundamental for optimizing their application in health-promoting and therapeutic formulations.
The integration of advanced imaging modalities—such as high-resolution microscopy and label-free spectroscopic techniques—with rheological tools will be essential for providing real-time, quantitative insights into how polysaccharides influence mucus structure and function20. Equally important is the enhancement of physiologically relevant in vitro models. Innovations in gut-on-a-chip systems and patient-derived, mucus-secreting organoids will be instrumental in replicating human-like mucus environments and immune responses, thereby bridging the gap between laboratory models and clinical applications21.
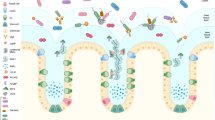
This review provides an in-depth and mechanistically oriented exploration of the interactions between polysaccharides and the colonic mucus barrier, with a particular emphasis on the influence of polysaccharide charge. We examine the underlying biophysical mechanisms—including electrostatic binding, hydrogen bonding, hydrophobic association, and van der Waals forces—that govern the behavior of polysaccharides within the mucus milieu. Furthermore, we critically assess the experimental platforms used to investigate these interactions, such as in vitro models, ex vivo tissue preparations, in vivo systems, organoid cultures, and microfluidic gut-on-a-chip technologies, outlining their respective strengths and methodological constraints. Lastly, we discuss the physiological consequences of these interactions, including effects on mucin dynamics, barrier structure and permeability, immune and epithelial responses, and protection against oxidative mucosal injury. By synthesizing current findings across these dimensions, this review aims to advance the conceptual framework of polysaccharide–mucus interplay and to highlight emerging opportunities for leveraging these interactions in clinical and nutritional interventions targeting mucosal health (as shown in Fig. 1).

Dietary polysaccharide chains entering the colonic lumen encounter a bilayer mucus barrier composed of a microbe-colonized outer layer and a dense, MUC2-rich inner layer adjacent to the epithelium. The schematic highlights four non-mutually exclusive routes by which polysaccharides can influence mucus function: (1) direct mucus physicochemical effects (mucoadhesion, swelling, viscoelasticity, and altered penetrability); (2) epithelial and goblet-cell signalling regulating mucin secretion, glycosylation, and turnover; (3) microbiota-mediated processing and fermentation, generating short-chain fatty acids (SCFAs) and other metabolites that feed back on mucus production and barrier integrity (e.g., via SCFA–GPCR signalling); and (4) downstream immune modulation within the lamina propria (e.g., macrophages, dendritic cells, T cells) that shapes inflammatory tone and mucus homeostasis.
Structural organization of colon mucus
Colon mucus is a bilayer structure and is secreted by goblet cells
The colonic mucus is organized into a specialized bilayer structure comprising a dense, firmly attached inner layer and a more aqueous, loosely structured outer layer22. Each layer plays a distinct and essential role in preserving intestinal homeostasis. The inner mucus layer is closely affixed to the epithelial lining, forming a nearly sterile zone that serves as a protective shield against luminal microbes and harmful substances23. Under physiological conditions, this layer is largely impervious to bacterial infiltration, thereby maintaining a physical and functional separation between the gut microbiota and epithelial cells to avoid aberrant immune stimulation24. In contrast, during pathological conditions such as inflammation, microbial imbalance, or infection, this barrier can become compromised—either thinning or breaking down—which permits microbial encroachment upon epithelial surfaces, potentially provoking immune activation and aggravating disease processes25,26.
The outer mucus layer, in comparison, is less compact and highly hydrated, functioning as a dynamic interface for hosting a broad spectrum of commensal microorganisms27. This layer supports microbial adhesion, nutrient metabolism, and bidirectional host–microbe communication28. Certain health-promoting bacteria preferentially inhabit this layer, utilizing mucin glycans as a nutrient source and contributing to intestinal well-being. Notably, Akkermansia muciniphila is recognized for its ability to degrade mucins in a manner that spares the inner layer, thereby supporting mucus layer maintenance, strengthening epithelial defense, and lowering inflammation—traits that highlight its potential in probiotic applications29. Similarly, species of Bacteroides, including B. thetaiotaomicron and B. fragilis, are adept at metabolizing mucin-derived glycans and are known to support mucus turnover and modulate immune responses30. The shift from the inner to the outer mucus layer involves progressive enzymatic cleavage and expansion of mucin molecules—chiefly MUC2—allowing for structural loosening and microbial accessibility in the outer compartment31. This spatial differentiation is vital for maintaining epithelial defense, mucosal immune function, and stable symbiosis between host and resident microbiota. Perturbation of this architecture is closely linked to various gastrointestinal disorders.
Goblet cells—specialized secretory epithelial cells prevalent throughout the colon and increasingly dense in its distal sections—are the principal producers of mucins, particularly the gel-forming MUC232. MUC2 biosynthesis begins in the endoplasmic reticulum, where it undergoes folding and is subsequently glycosylated in the Golgi apparatus33. It is then polymerized through C-terminal dimerization and N-terminal oligomerization into large, complex mucin networks. These polymers are stored in intracellular granules within goblet cells under conditions of low pH and elevated calcium concentrations34. The secretion of mucins is controlled by a highly orchestrated exocytosis process mediated by the SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor) protein complex35. This mechanism ensures precise vesicle fusion between mucin-containing granules and the apical plasma membrane. The SNARE complex forms through the interaction of vesicle-associated SNAREs, such as VAMP8, with target membrane SNAREs, including syntaxins and SNAPs36. In response to external stimuli—ranging from microbial signals and pro-inflammatory cytokines to mechanical forces—goblet cells activate regulated exocytosis37. Once released, mucins rapidly expand due to hydration and ionic shifts, creating a viscoelastic gel that replenishes the mucus layer and sustains gut barrier integrity.
Classes of polysaccharides relevant to mucus interactions
Dietary and bioactive polysaccharides that reach the colon can be broadly grouped by net surface charge (neutral, anionic, and cationic), which is a primary determinant of how they associate with the MUC2-rich mucus gel and influence barrier function. Neutral polysaccharides lack ionizable carboxyl, sulfate, or amine groups at physiological pH, but present dense arrays of hydroxyls capable of extensive hydrogen bonding. Typical examples include resistant starch from cooked-and-cooled starchy foods, microbial pullulan produced by Aureobasidium pullulans, cereal and fungal β-glucans, inulin-type fructans from chicory, garlic, onion, and asparagus, and galactomannans such as guar gum and locust bean gum38,39,40,41. Because these molecules are often resistant to digestion in the upper gastrointestinal tract, substantial fractions reach the colon intact, where their hydrated chains can interpenetrate the mucin gel and engage PTS domains primarily through hydrogen bonding and chain entanglement rather than long-range electrostatic interactions42.
Anionic polysaccharides are widely used in food, pharmaceutical, and biomedical applications. Alginate and sulfated fucoidans from brown seaweeds, carrageenans from red seaweeds, plant pectins from citrus peel and apple pomace, and hyaluronic acid from vertebrate connective tissues all carry carboxylate and/or sulfate groups that are ionized under intestinal conditions43,44,45. These negatively charged polymers encounter a mucin matrix that is itself decorated with sialylated and sulfated terminal residues, creating zones of electrostatic repulsion but also opportunities for association via counter-ion bridging, hydrogen bonding, and interactions with locally cationic regions of the mucin protein backbone46. Importantly, dextran sulfate sodium (DSS) is a synthetic, highly sulfated anionic polysaccharide that is widely used as an experimental tool (e.g., colitis); because DSS can directly perturb the MUC2 gel and increase mucus permeability, mucus-related endpoints in DSS-based studies should be interpreted with this direct mucus-active confounder in mind47. At the opposite end of the electrostatic spectrum are cationic polysaccharides, most notably chitosan, obtained by partial deacetylation of chitin from crustacean shells, insects, and fungal cell walls48,49. Protonation of its primary amino groups under mildly acidic conditions confers a strong positive charge, which promotes high-affinity binding to the anionic glycans and protein domains of MUC250. This charge-based classification provides a simple navigation aid for later sections discussing electrostatic association, rheology/tribology, and mucus-penetration assays.
Biochemical composition of colon mucus
The colonic mucus layer is predominantly formed by the gel-forming secreted mucin MUC2, which builds a stratified, viscoelastic, and selectively permeable barrier above the epithelium, while additional secreted and transmembrane mucins (e.g., MUC1, MUC3, MUC4, MUC13, and MUC17) are also expressed and contribute to the epithelial glycocalyx and the biochemical properties of the mucus layer51. MUC2, synthesized and secreted by goblet cells, is a large, gel-forming mucin with a complex, modular structure. Its central region is enriched in proline, threonine, and serine—collectively forming the PTS domains—which serve as key sites for extensive O-linked glycosylation52. These carbohydrate side chains account for over 80% of the mucin’s molecular mass and are essential for maintaining hydration, mechanical stability, and resistance to proteolytic attack53.
The functional behavior of mucus is largely determined by the structural diversity of the O-glycans decorating the PTS domains of MUC2. The glycosylation process is initiated in the Golgi apparatus, where N-acetylgalactosamine (GalNAc) is added to serine or threonine residues, forming the Tn antigen—a precursor to mucin-type O-glycans54. Further enzymatic extensions yield various core structures, predominantly Cores 1 to 4, in intestinal mucins55. Core 1 (Galβ1–3GalNAcα1–Ser/Thr) forms the base for further branching and modification56. Core 2 (GlcNAcβ1–6(Galβ1–3)GalNAcα1–Ser/Thr) introduces branching that enhances resistance to enzymatic degradation and modulates microbial recognition57. Core 3 (GlcNAcβ1–3GalNAcα1–Ser/Thr), primarily found in the colon, is instrumental in forming the hydrated, gel-like consistency crucial for effective barrier function58. Core 4 (GlcNAcβ1–6(GlcNAcβ1–3)GalNAcα1–Ser/Thr), although less frequently expressed, plays specialized roles in mucosal defense59. These core structures are further elongated and capped with terminal modifications such as fucose, sialic acid, or sulfate, which modulate mucus charge, hydration, and microbial interactions. Alterations in the expression of O-glycan core structures—such as the reduction of Core 3 expression in inflammatory bowel disease—can significantly impact mucus architecture and its interaction with commensal microbes, pathogens, and dietary components, including polysaccharides and other luminal molecules60.
In the healthy distal colon, MUC2 O-glycans are heavily decorated with sialylated, sulfated, and mono-fucosylated motifs, generating a highly hydrated, strongly anionic brush that both supports commensal colonization and provides abundant electrostatic and hydrogen-bonding sites for cationic and neutral polysaccharides. By contrast, active ulcerative colitis and other inflammatory conditions are associated with a shift toward shorter, less complex O-glycans structures and truncated Tn/sialyl-Tn antigens, and with regionally altered sulfation and sialylation of MUC2. Genetic loss of core 3–derived O-glycans (C3GnT deficiency) or combined core 1/core 3 defects similarly thins the inner mucus layer, increases bacterial proximity to the epithelium, and perturbs the spatial distribution of negative charges within the mucin domain. From the perspective of polysaccharide-mucus interactions, such disease-associated remodeling of MUC2 glycans is expected to weaken classical electrostatic binding sites for cationic polymers like chitosan when sulfated and sialylated termini are lost, while simultaneously exposing more peptide backbone and neutral glycan epitopes that favor hydrogen bonding or hydrophobic contacts with neutral or amphiphilic polysaccharides. Conversely, hyper-sialylation of mucin O-glycans, as reported in certain colitis and colorectal cancer settings or in mice with dysregulated epithelial sialyltransferase activity, increases local negative charge density and may intensify repulsive forces toward strongly anionic polysaccharides (fucoidans) but enhance multivalent binding of cationic or zwitterionic polysaccharide formulations. Changes in terminal fucose and sialic-acid display also rewire recognition by mucin-degrading bacteria that co-metabolize dietary polysaccharides; for example, altered fucosylation and sialylation in inflammatory bowel disease reshape the repertoire of commensals that can adhere to and harvest MUC2 glycans, thereby indirectly influencing how exogenous polysaccharides are fermented and how their microbial breakdown products (such as SCFAs) feed back on goblet-cell function and mucus barrier integrity. Therefore, the compositional and spatial diversity of mucin O-glycans is fundamental not only to the mucus architecture but also to its dynamic interface with microbes and therapeutic agents.
In addition to mucins, the colonic mucus layer is enriched with numerous bioactive substances that reinforce its defensive and regulatory functions. Antimicrobial peptides—such as defensins, cathelicidins, and lysozyme—are secreted by epithelial cells and Paneth cells (particularly in the small intestine), and are embedded within the mucus, where they neutralize microbial threats by disrupting bacterial membranes61,62. Secretory immunoglobulin A, produced by local plasma cells and transported across the epithelium, binds to microbial antigens and toxins to facilitate immune exclusion while minimizing inflammatory responses63. Lipids, including phospholipids, contribute to the hydrophobic characteristics of the mucus and influence mucin structure64. Enzymes such as proteases, glycosidases, and peroxidases participate in mucus turnover, antimicrobial defense, and oxidative homeostasis65. Furthermore, ions like calcium and bicarbonate regulate mucus viscosity, facilitate pH buffering, and support mucin unfolding and cross-linking66. Together, these diverse elements compose a dynamic, multifunctional barrier that preserves epithelial homeostasis and fosters mucosal immune balance.
Biophysical principles governing polysaccharide–mucus interactions
Electrostatic interactions
Electrostatic, or ionic, interactions are a principal mechanism underlying the adhesion of polysaccharides to mucosal surfaces. These interactions occur primarily through the attraction between oppositely charged functional groups on polysaccharide chains and mucin glycoproteins67. Mucins, the predominant structural components of mucus, are extensively glycosylated and enriched with negatively charged moieties—primarily sialic acid and sulfate residues—rendering them highly anionic68. In contrast, cationic polysaccharides, especially those carrying protonatable amine groups, exhibit strong electrostatic affinities for these negatively charged sites69.
Chitosan, one of the most extensively studied cationic polysaccharides, contains primary amino groups that become positively charged (–NH3+) under mildly acidic conditions (typically below pH 6.5)70. These charged groups facilitate ionic bonding with mucin’s anionic side chains, leading to the formation of stable mucoadhesive complexes71. Multiple factors govern the strength and stability of these interactions, including the polysaccharide’s molecular weight, degree of substitution, environmental pH, and ionic strength72,73. Notably, Menchicchi et al. found that chitosan–mucin interactions were markedly stronger in deionized water than in saline (0.1 mol/L NaCl), highlighting the crucial role of low ionic strength in enhancing electrostatic adhesion. Furthermore, additional non-covalent interactions—such as hydrogen bonding and hydrophobic forces—also contribute to complex stabilization, particularly in chitosan samples with higher degrees of acetylation74.
The surrounding ionic environment significantly influences electrostatic binding. In physiological fluids with elevated salt concentrations, mobile ions can shield charged sites, weakening or even preventing electrostatic interactions between chitosan and mucin75,76. pH is another critical determinant. At lower pH values, chitosan’s amino groups remain protonated, maximizing mucoadhesive potential77. However, as the environment becomes neutral or alkaline, these groups lose their charge, diminishing chitosan’s affinity for mucin. This pH responsiveness is physiologically significant: while the colon typically maintains a near-neutral pH (~6.8), inflamed or cancerous tissues often present with more acidic microenvironments (pH < 5.5)78,79. Low-molecular-weight chitosan tends to form dense complexes under acidic conditions and relies on alternative binding modes at higher pH levels. Conversely, strongly anionic high-molecular-weight polysaccharides, like dextran sulfate, can undergo phase separation with mucins, where localized cationic domains mediate binding14.
Due to these properties, chitosan and its chemically modified derivatives have become prominent mucoadhesive agents in both nutraceutical and pharmaceutical formulations80. Their ability to enhance mucosal adhesion extends the retention time of bioactive substances, promotes targeted delivery, and supports sustained release profiles. Advances in polymer chemistry have yielded quaternized chitosan derivatives that possess permanent positive charges, independent of ambient pH81. Likewise, Yang et al. formulated a colon-targeted nanoemulsion using quaternized chitosan loaded with Angelica extract, which preserved mucosal structure and mitigated inflammatory responses in a murine model of ulcerative colitis82.
Hydrogen bonding
Hydrogen bonding represents a critical and synergistic force in the mucoadhesive behavior of polysaccharides, especially those that are neutral or weakly anionic and structurally enriched with hydrophilic functional groups such as hydroxyl (–OH), carboxyl (–COOH), and ether (–O–) linkages. These groups act as both hydrogen bond donors and acceptors, enabling the formation of dynamic non-covalent interactions with the polar moieties on mucins. Mucins, particularly the glycosylated regions of their oligosaccharide side chains, are densely decorated with monosaccharides such as galactose, N-acetylglucosamine (GlcNAc), and sialic acid, which provide a high density of acceptor and donor sites for hydrogen bonding. When polysaccharides bearing complementary hydrophilic structures interface with the mucus gel, they can form an interpenetrating hydrogen-bonded matrix that reinforces adhesion, enhances mucosal hydration, facilitates gelation, and prolongs residence time on epithelial surfaces.
Experimental evidence has consistently underscored the role of hydrogen bonding in polysaccharide–mucin adhesion. For instance, gellan gum, a linear anionic polysaccharide rich in hydroxyl and carboxyl groups, displayed robust mucoadhesive behavior under both acidic (pH 1.2) and near-neutral (pH 6.8) conditions. Scanning electron microscopy (SEM) revealed intimate contact and morphological integration between gellan and mucin, with stronger interaction observed at acidic pH—indicating enhanced hydrogen bonding and possible hydrophobic contributions in such environments83. In contrast, retrograded starch, lacking sufficient polar group accessibility, exhibited minimal binding affinity to mucin, as evidenced by unaltered rheological behavior and weak bioadhesive force relative to mucin alone. Further supporting this mechanism, fucoidan extracted from Thelenota ananas showed considerable mucoadhesive potential in simulated intestinal fluids. The adhesion was driven by multiple non-covalent forces, including hydrogen bonding, hydrophobic interactions, and supramolecular disulfide bridges, with hydrogen bonding recognized as a primary contributor to the observed adhesive strength84. Likewise, polysaccharides from Hohenbuehelia serotina formed stable complexes with mucin, resulting in the self-assembly of nanoparticles. Notable physicochemical changes—including increased particle size and modifications in FT-IR spectral features (e.g., flattening of the broad –OH stretch at 3358 cm⁻¹) and X-ray diffraction peaks (shifts at 2θ = 9.4° and 21.3°)—highlighted the central role of hydrogen bonding and hydrophobic stacking in complex formation85. The involvement of hydrogen bonding has also been elucidated through studies on κ- and κ/β-type carrageenans, which form mucin–polysaccharide networks via hydrogen bridges. The disruption of these networks by denaturing agents such as urea—which specifically breaks hydrogen bonds—further validates the pivotal contribution of hydrogen bonding in carrageenan-mediated mucoadhesion86. Collectively, these findings emphasize that hydrogen bonding, though often cooperative with other non-covalent forces, is a foundational mechanism for the establishment and stability of polysaccharide–mucin interactions in mucosal environments.
Hydrophobic interactions and Van der Waals forces
Hydrophobic interactions, though often considered secondary to electrostatic and hydrogen bonding forces, play an important yet underappreciated role in the mucoadhesion of polysaccharides. These interactions arise between nonpolar segments of mucin glycoproteins and apolar domains on polysaccharides—particularly those that have undergone chemical modifications such as alkylation, acylation, or the grafting of hydrophobic side chains87,88. While mucins are predominantly hydrophilic due to their extensive glycosylation and water-retaining properties, they also possess hydrophobic core regions enriched with nonpolar amino acids like leucine, valine, and phenylalanine, especially within their central protein backbone89,90. These hydrophobic patches provide anchoring sites for polysaccharides that present similar nonpolar moieties, facilitating apolar associations at the mucus interface91.
Native polysaccharides, typically modified by their functionalization with long-chain alkyl or aromatic groups, dramatically enhance their capacity for hydrophobic binding92. Amphiphilic derivatives, such as hydroxypropyl methylcellulose phthalate or thiolated chitosans modified with fatty acid chains, demonstrate significantly improved mucoadhesion through the embedding of their hydrophobic segments into mucin’s nonpolar regions93. Importantly, unlike electrostatic interactions, hydrophobic forces remain largely unaffected by variations in pH and ionic strength, offering stable adhesion across the heterogeneous conditions of the gastrointestinal tract94.
Hydrophobic interactions have further implications in the design of delivery systems for lipophilic bioactives95. In both pharmaceutical and functional food contexts, polysaccharide carriers engineered with hydrophobic domains improve the solubilization and encapsulation efficiency of poorly water-soluble compounds such as curcumin, vitamin D, and omega-3 fatty acids96,97. These carriers not only facilitate the targeted release and enhanced absorption of lipophilic payloads but also benefit from prolonged mucosal adhesion, extending their residence time at the absorptive site.
In addition to hydrophobic interactions, van der Waals forces contribute to the polysaccharide–mucin adhesion profile. These weak, short-range interactions—originating from transient dipole–dipole attractions—become functionally relevant when polysaccharide chains closely approach the mucin network following initial contact mediated by stronger interactions98. While van der Waals forces are individually weak, their cumulative effect across extended molecular surfaces, especially in high-molecular-weight or highly flexible polysaccharides, can reinforce mucoadhesive integrity99. For example, formulations incorporating tamarind seed-derived xyloglucan and sodium chondroitin sulfate demonstrated mucoadhesive enhancement attributed in part to van der Waals contributions, alongside hydrogen bonding and hydrophobic effects100. Molecular docking studies by Periasamy et al. also revealed that tamarind xyloglucan could engage mucin1 through a multipoint interaction scheme involving electrostatic forces, hydrogen bonding, and van der Waals interactions101.
Altogether, hydrophobic and van der Waals forces act in concert with hydrogen bonding and ionic interactions to construct a multifaceted adhesion architecture at the polysaccharide–mucus interface. Understanding and harnessing these composite mechanisms is essential for the rational design of next-generation mucoadhesive systems and gastrointestinal-targeted biotherapeutics.
Experimental models of polysaccharide-mucus interactions
In vitro cell culture models
In vitro cell culture systems provide a highly controlled and reproducible environment for exploring the synthesis, release, and functional characteristics of colonic mucus under both normal and disease-like conditions102. Commonly used intestinal epithelial cell lines for studying polysaccharide–mucus interactions include goblet-like LS174T and HT29-MTX cells, which secrete gel-forming mucins such as MUC2 and MUC5AC and can generate a measurable mucus layer under optimized culture conditions103,104. In contrast, Caco-2 cells differentiate predominantly into absorptive enterocyte-like monolayers and, under standard submerged conditions, exhibit very low production of gel-forming mucins and do not form a physiologically relevant mucus layer. Therefore, Caco-2 cells are more appropriate for investigating epithelial permeability and transporter-mediated uptake than for modeling the native colonic mucus barrier105. To better mimic a mucus-covered epithelium, including the use of co-culture systems in which mucus-producing HT29-MTX or LS174T cells are combined with Caco-2 cells, rather than relying on Caco-2 monocultures as a mucus source106.
Despite their accessibility, standard submerged epithelial monolayers are inherently static and reductionist systems, which can materially impact polysaccharide–mucus readouts. In particular, the absence of luminal flow and shear means that mucus renewal dynamics are not reproduced. These models also typically lack key external factors that shape mucus biology and polysaccharide behavior—such as oxygen and pH gradients, bile acids and digestive components, inflammatory cytokine milieus, vascular/neuronal inputs, and immune cell crosstalk—while microbiota are usually absent107.
In studies examining the stimulatory effects of polysaccharides, laminaran has been found to modulate lipopolysaccharide (LPS)-induced responses in LS174T cells by upregulating proteins involved in mucin secretion. These include sulfotransferases, vesicle-associated membrane protein 8 (VAMP8), and synaptosome-associated protein 23 (SNAP-23), with protein kinase C (PKC) identified as a key component of the signaling cascade108. In another study, the HT29-MTX cell model was employed to explore the influence of chitosan’s degree of polymerization (DP) on mucus binding and crosslinking (Fig. 2). As illustrated in Fig. 2A, binding of chitosan to differentiated, mucus-covered HT29-MTX monolayers was quantified by measuring the reduction in chitosan concentration remaining in the supernatant after incubation, providing an integrated readout of polymer association with the mucus layer. Confocal microscopy images in Fig. 2B–B" show that low-molecular-weight chitosan (DP8) penetrates deeply into the mucus gel and even approaches the epithelial cell layer, producing a more homogeneous green fluorescence signal throughout the mucus, consistent with partial interpenetration and physical crosslinking of mucin polymers. In contrast, higher molecular weight chitosans (DP52 and DP100) form a bright fluorescent band restricted to the apical mucus surface, indicating predominantly surface-level association with limited penetration into the mucus network109.

A Quantification of chitosan binding to mature HT29-MTX monolayers overlaid with mucus. Chitosan association was determined by measuring the reduction in chitosan concentration in the supernatant following incubation with the cell cultures. B–B” Confocal microscopy images of HT29-MTX layers treated with FITC-labeled chitosan of varying degrees of polymerization: DP8 (B), DP52 (B’), and DP100 (B”). Basal focal planes show cells stained with CellTracker™ Deep Red (red), while apical planes highlight chitosan-FITC (green). Cross-sectional images demonstrate that low molecular weight chitosan (DP8) penetrates the mucus and cellular layers more effectively, whereas higher molecular weight chitosans (DP52 and DP100) tend to accumulate on the surface. Scale bar = 100 μm. Adapted with permission from ref. 109, Copyright 2018, American Chemical Society.
The influence of gut microbiota and their fermentation byproducts on mucus secretion can be effectively evaluated using in vitro cell culture systems110. Because many dietary polysaccharides reach the colon intact, they are first utilized by the gut microbiota to generate SCFAs and other organic acids; these fermentation products then act on epithelial/goblet-cell programs to remodel mucin secretion and glycosylation. For instance, the response of O-glycans in HT29-MTX cells was examined following exposure to pectin fragments generated from in vitro fermentation using human fecal inocula. The study demonstrated that these fermentation-derived fragments stimulated the production of fucosylated neutral O-glycans, while simultaneously reducing levels of sulfated and sialylated O-glycans. Specifically, high-molecular-weight fragments (~81.04 kDa) led to a marked increase in the relative abundance of non-fucosylated O-glycans, whereas low-molecular-weight fragments (~16.02 kDa) favored the formation of di-fucosylated O-glycans111. Extending this “polysaccharide, fermentation, host-cell” workflow, the in vitro gut microbiome consortia to ferment structurally distinct dietary-fiber polysaccharides, quantified the resulting SCFAs mixtures, and showed that microbe-derived metabolites promoted morphological differentiation and increased mucin secretion in colon cells112. Moreover, partially hydrolyzed guar gum was shown to elevate SCFAs and succinate concentrations, with succinate promoting MUC2 expression in LS174T cells via phosphorylation of AKT113.
Co-culture and goblet cell–enriched in vitro systems can improve physiological relevance, but changes in mucin transcripts (e.g., MUC2) alone are insufficient evidence of a functional mucus barrier; therefore, studies should prioritise mucus-sensitive readouts (secreted mucin abundance, mucus thickness/continuity, and penetrability) alongside epithelial barrier metrics (e.g., TEER and tracer permeability)114,115.
In addition to cell-based cultures, many in vitro biophysical assays use purified mucins as mucus surrogates; commercially available porcine gastric mucin (PGM; e.g., Sigma type II/III) is convenient for diffusion, rheology, electrochemical, and mucoadhesion screening, but it is derived primarily from gastric mucins (MUC5AC/MUC6) and does not recapitulate the MUC2-rich, bilayered organisation of colonic mucus116. Therefore, PGM-based findings should be interpreted as preliminary and, where feasible, validated using more physiological systems (e.g., freshly isolated colonic mucus, mucus-enriched ex vivo tissues, organoid-derived mucus, or engineered MUC2-containing mimetics).
In vivo models
In vivo models provide a valuable framework for examining the structure, composition, and function of intestinal mucus within its native physiological context117. Importantly, most in vivo mucus endpoints in polysaccharide intervention studies are obtained from endpoint, fixed or otherwise preserved intestinal sections (paraffin-embedded or cryosections) rather than continuous live measurements; thus, they represent spatial snapshots that are sensitive to tissue collection and processing conditions. Mice are the most frequently utilized species due to their genetic accessibility and the availability of various transgenic lines. These models have been instrumental in elucidating the roles of mucins in preserving epithelial barrier integrity, modulating host–microbiota interactions, and influencing immune responses118. The murine colon, which exhibits a two-layered mucus system similar to that found in humans, serves as a robust model for investigating mucus dynamics in conditions such as colitis, colorectal cancer, and microbiota-associated disorders119. Other animals, including pigs and rats, are also employed owing to their closer anatomical and biochemical similarity to the human gastrointestinal tract, particularly in terms of mucin expression patterns120,121.
Integrating in vivo studies with advanced imaging and spectroscopic tools enables a comprehensive assessment of mucus behavior, including its secretion, degradation, and protective functions during inflammation, infection, or dietary polysaccharide interventions. This multidimensional approach enhances our understanding of mucus regulation in biologically relevant conditions and supports the development of therapeutic strategies targeting the mucus barrier122,123. In DSS-induced colitis models, DSS is a common trigger used to generate reproducible mucus injury and inflammation; however, because DSS is itself a highly sulfated anionic polysaccharide, it can directly perturb the MUC2 gel and increase mucus permeability, and thus should be considered when attributing mucus changes to the test polysaccharides47.
A variety of histological and imaging techniques remain indispensable for baseline characterization of the mucus barrier in polysaccharide intervention studies. Commonly used approaches include Alcian blue-periodic acid-Schiff staining to distinguish acidic versus neutral mucins and quantify goblet cells, antibody-based immunohistochemistry or immunofluorescence to localize mucins such as MUC2 and MUC5AC124, fluorescence in situ hybridization to map mucus-associated bacterial populations125, lectin histochemistry to probe mucin glycosylation patterns, and electron microscopy to resolve ultrastructural changes in goblet cells and the mucus gel. For instance, in a DSS-induced colitis model, MUC2 fluorescence was markedly reduced; however, treatment with Tremella fuciformis polysaccharides significantly enhanced MUC2 signal intensity, unlike the conventional anti-inflammatory agent 5-aminosalicylic acid, which had no noticeable impact on MUC2 levels126. In addition, treatment with Abelmoschus manihot polysaccharides reversed this effect by restoring mucus thickness and increasing the microbial–epithelial distance, thereby limiting microbial translocation127.
Lectin histochemistry is a highly informative approach for studying mucin glycosylation patterns in mucus biology128. Lectins are sugar-binding proteins that exhibit high specificity toward distinct carbohydrate residues, such as neutral sugars, sialylated residues, and particular monosaccharides, including GlcNAc, galactose, GalNAc, fucose, N-acetylneuraminic acid, and mannose129,130. Commonly utilized fluorescently labeled lectins include Ulex europaeus agglutinin I (UEA-I, specific for Fuc), concanavalin A (ConA, specific for Man), Dolichos biflorus agglutinin (DBA, specific for GalNAc), peanut agglutinin (PNA, specific for Gal), and wheat germ agglutinin (WGA, GlcNAc)131. This technique proves particularly useful for evaluating glycosylation alterations in various physiological and pathological states—such as inflammation, infection, cancer progression, and dietary modulation—since glycan structure variations can affect mucus properties like viscosity, microbial adhesion, and immune interactions. For example, Wen et al. applied lectin histochemistry to assess the impact of pectin supplementation on the intestinal barrier in LPS-challenged piglets. In the LPS-treated group, GlcNAc expression increased while fucose and galactose levels declined; however, pectin administration normalized these glycan profiles. Mannose levels remained unchanged, and GalNAc expression was not detected in the ileum132. Fucosylation, in particular, is often used as an indicator of MUC2 glycan stability. UEA-I lectin, which targets Fucα1,2 Gal motifs, showed strong staining in the mucus layer of wild-type (WT) mice. Interestingly, treatment with a cocktail of broad-spectrum antibiotics (including neomycin sulfate, metronidazole, vancomycin, and ampicillin) led to a slight enhancement in UEA-I binding, indicating that microbiota-driven degradation may influence the fucosylation status of O- and N-linked mucin glycans133.
Electron microscopy serves as a vital technique in mucus-related research, enabling detailed visualization of the mucus layer, goblet cells, and mucin granules at the ultrastructural level134. The two principal forms of electron microscopy used in this context are transmission electron microscopy (TEM) and SEM. TEM provides high-resolution images of internal cellular components, making it well-suited for examining mucin granule morphology, goblet cell structure, and the processes involved in mucin synthesis and secretion135. For instance, Ma et al. utilized TEM to examine goblet cell ultrastructure in a DSS induced colitis mouse model. Their observations revealed that goblet cells in the colitis group displayed diminished mucin granules and irregularly spaced, sparse microvilli. Treatment with 5-aminosalicylic acid and a modified Gegen Qinlian decoction partially reversed these changes, suggesting an improvement in goblet cell function and mucin release136. In contrast, SEM allows for surface-level imaging that offers a three-dimensional perspective of the epithelial mucus layer, which is crucial for evaluating the physical integrity, texture, and spatial distribution of mucus137. In a study by Taniguchi et al., SEM was used to analyze the colonic mucus surfaces of WT and genetically modified mice lacking the glycosyltransferases β-1,3-galactosyltransferase 5 (B3galt5) and St6 N-acetylgalactosaminide α-2,6-sialyltransferase 6 (St6galnac6)(Fig. 3). The WT mice exhibited a smooth, well-organized mucus layer, while the knockout models showed a rougher, less uniform surface with disrupted mucin polymer networks. These findings indicate that impaired sialylation compromises mucus structure and polymer cohesion138. However, most TEM- and conventional SEM-based studies, primarily capture epithelial and goblet-cell ultrastructure; sample preparation steps such as chemical fixation, dehydration, and coating can collapse or wash away the highly hydrated secreted mucus, limiting insight into its native thickness and three-dimensional organization. To better preserve mucus ultrastructure, cryo-SEM and related cryogenic preparation methods rapidly freeze tissues and maintain the gel-like state during imaging, enabling more faithful visualization of the surface and sub-surface mucin network139.

a SEM images of colonic mucosal surfaces from WT, St6galnac6−/−, and B3galt5−/− mice. Green dashed outlines (top panels) indicate mucus covering the epithelium. The middle panels present higher magnification views of regions marked with white dots. Lower panels show further magnified images of areas outlined by white squares. b Representative SEM images used for mucin fiber orientation analysis. From top to bottom: original images, binarized versions, two-dimensional fast Fourier transform (2D-FFT) representations, and ellipses approximated from Fourier amplitude coefficients, indicating fiber alignment. Scale bar = 100 nm. c Quantification of fiber anisotropy, expressed as the square root of (major axis b / minor axis a) from the ellipses in (b, bottom row). d SEM images showing gut bacteria embedded within the colonic mucus. Red dashed lines denote bacterial orientation, while blue dashed outlines indicate regions of unidirectionally aligned mucin polymers. Blue arrows mark the direction of mucin fiber alignment; red arrows highlight rod-shaped bacteria penetrating between mucin strands. Lower panels provide enlarged views of the boxed regions above. Scale bars: 5 μm (upper) and 1 μm (lower). B3galt5 β-1,3-galactosyltransferase 5, St6galnac6 ST6 N-acetylgalactosaminide α-2,6-sialyltransferase 6, WT wild-type, ns not significant. Adapted with permission from ref. 138, copyright 2025, Elsevier.
A major challenge in in vivo studies is the reliable preservation of the native mucus architecture during tissue collection and processing. The mucus gel is highly hydrated and labile; conventional aqueous fixatives such as neutral buffered formalin often dissolve or collapse the mucus layer, leading to an apparent loss or marked thinning in histological sections. Although in vivo models are indispensable for capturing integrated mucus responses to polysaccharides within an intact host, their susceptibility to fixation-related artefacts (and microbiota-driven confounding) underscores the need to corroborate observations across multiple experimental platforms and to avoid over-attributing observed changes to direct polysaccharide–mucus interactions alone.
Organoids
Recent progress in tissue engineering has facilitated the emergence of advanced in vitro models such as intestinal organoids, which more accurately replicate the physiological characteristics of colonic mucus compared to conventional two-dimensional cell cultures140.
Intestinal organoids, derived from adult stem cells or induced pluripotent stem cells141, self-assemble into three-dimensional structures that can replicate some aspects of the complex cellular composition and architecture of the native intestinal epithelium under specific differentiation and culture conditions, including the generation of mucus-producing goblet cells that secrete MUC2142. These models support prolonged cultivation and enable comprehensive studies of how dietary polysaccharides influence mucus synthesis, degradation, and interactions with both microbial communities and immune signaling pathways143,144. A notable advantage of organoids is their suitability for genetic manipulation, which permits mechanistic exploration of mucin gene regulation and mucus-associated diseases. For instance, ginseng-derived nanofibers (GNFs) were tested for their ability to protect intestinal barrier function in small intestinal organoids derived from BALB/c mice (Fig. 4). Following LPS challenge, immunofluorescence analysis showed a significant downregulation of epithelial markers MUC2 and E-cadherin, which was markedly reversed upon treatment with GNFs, particularly at a concentration of 250 µg/mL145.

A Representative hematoxylin and eosin (H&E)-stained sections of intestinal organoids treated with GNFs at 62.5, 125, and 250 µg/mL, compared to untreated controls and LPS-stimulated groups. Scale bar = 50 µm. B Immunofluorescence staining of organoids for key markers of mucosal integrity and inflammation. MUC2 (red) and E-cadherin (green) indicate mucus production and epithelial cell adhesion, respectively, while ZO-1 (red) and Occludin-1 (green) reflect tight junction protein expression. DAPI (blue) marks nuclei. Scale bar = 50 µm. C Quantification of inflammatory cytokines (TNF-α, IL-6, IL-1β) in organoid supernatants by ELISA following LPS stimulation and treatment with increasing concentrations of GNFs. D Western blot analysis of TLR4, phosphorylated AKT (p-AKT), total AKT, phosphorylated NF-κB p65 (p-p65), and total p65 expression in organoids treated with GNFs, relative to LPS-induced and control groups. Adapted with permission from ref.145, copyright 2025, Elsevier.
It is essential to consider how test compounds access the apical (luminal) surface, where the mucus layer is naturally located. In standard “basolateral-out” 3D organoids embedded in extracellular matrix, the apical surface and associated mucus accumulate within a closed central lumen, whereas culture medium and test compounds primarily contact the basolateral side146. As a result, luminal polysaccharides, microbial products, or SCFAs must be delivered to the organoid interior—typically via microinjection into the lumen—to model true apical exposure. Although microinjection preserves the three-dimensional organization and mucus compartment, it is technically demanding, low-throughput, and usually restricted to relatively small numbers of organoids per experiment147. For mucus-focused polysaccharide studies, greater emphasis should be placed on air–liquid interface organoid-derived monolayers, which provide direct apical access while promoting epithelial maturation and goblet-cell differentiation. The air–liquid interface mucus-monolayer systems have been shown to generate a continuous, MUC2-rich mucus layer and enable quantitative readouts, thereby increasing their relevance for polysaccharide–mucus mechanisms148.
Gut-on-a-chip
Gut-on-a-chip platforms represent a state-of-the-art innovation in in vitro gastrointestinal modeling, employing microfluidic systems to replicate essential physiological parameters such as peristaltic-like motions, fluid shear forces, and controlled oxygen gradients149,150. These biomimetic devices support the co-culture of intestinal epithelial cells alongside live microbial communities in a dynamic environment that closely mirrors in vivo intestinal conditions151. A distinctive feature of gut-on-a-chip technology is its ability to maintain continuous mucus production and accumulation on epithelial surfaces, thereby enabling real-time monitoring of epithelial barrier function, mucus layer thickness, and microbial translocation152,153. Furthermore, these platforms allow integration with advanced imaging modalities and biosensors, facilitating precise, time-resolved assessments of host–microbe–mucus interactions154,155. Although their complexity and maintenance costs remain relatively high, gut-on-a-chip systems offer substantial translational potential for preclinical drug testing, pathophysiological studies, and microbiome research. For example, Jing et al. engineered a gut-on-a-chip model to simulate inflammatory bowel disease, using Escherichia coli 11775 to induce infection-driven inflammation. Their findings indicated that treatment with chitosan oligosaccharides suppressed E. coli adhesion and invasion by downregulating TLR4 expression and inhibiting NF-κB DNA-binding activity within the chip environment156. In another study, McCoy et al. created an electrochemical gut-mucus interface by incorporating microelectrode arrays into the model to examine the impact of mucolytic agents and E. coli biofilms on mucus biophysical properties (Fig. 5). This system utilized poly(3,4-ethylenedioxythiophene): polystyrene sulfonate-coated gold electrodes, including optically transparent versions, enabling simultaneous electrical measurement and optical microscopy. Fig. 5B shows the macroscopic top view of the chip, whereas Fig. 5C presents a confocal image of a single 500 µm working electrode, in which the mucus layer is visualized by WGA–FITC staining as a continuous green coating over the conductive surface. This approach facilitated real-time, label-free evaluation of mucus architecture and its perturbation by microbial colonization or pharmaceutical compounds157. Notably, some chip-integrated electrochemical mucus interfaces use PGM hydrogels as a practical mucus layer116.

A Schematic representation of a microelectrode array consisting of twelve poly(3,4-ethylenedioxythiophene): polystyrene sulfonate (PEDOT:PSS)–coated gold working electrodes assembled with a mucus layer. B Top–down view of the electrode chip. C Confocal microscopy image of a single square working electrode (500 μm width) showing mucus stained green with WGA-FITC lectin. D SEM image of a 5% porcine gastric mucus (PGM) hydrogel used in electrochemical assays. Scale bar = 4 μm. E Representative Z-stack image of the WGA-FITC-stained mucus layer used to assess mucus thickness on transparent electrodes. F Cyclic voltammograms recorded at scan rates of 30, 50, 90, 120, and 150 mV/s in 10 mmol/L [Fe(CN)₆]3−/4− with 0.1 mol/L KCl in phosphate-buffered saline (PBS), measured on electrodes coated with a 1000 μm thick mucus hydrogel composed of 5% PGM and 2% PEG-4SH. Potentials are referenced to Ag/AgCl. Adapted from ref. 157 under the terms of the Creative Commons CC BY license.
Ex vivo
Ex vivo models act as a crucial intermediary between in vitro experiments and in vivo studies, enabling researchers to investigate colonic mucus under conditions that closely mimic those in the body. These systems retain the native tissue architecture and cellular interactions by culturing freshly excised colonic tissues or biopsies from humans or animals over short durations. Because they preserve the mucus-secreting epithelium along with stromal and immune cell components, ex vivo platforms provide a physiologically relevant context for studying mucin release, mucus layer properties such as thickness and permeability, and their interaction with microbes. Tools like Ussing chambers are routinely employed to measure ion transport and assess barrier integrity, while various staining and imaging techniques facilitate real-time observation of the mucus structure. For example, Sharpen et al. utilized an ex vivo perfusion system to monitor mucus barrier function by tracking fluorescent microparticles (Fig. 6)158. A fluorescein-labeled peptide substrate specific for transglutaminase 3 (E51) was added to the luminal buffer, and incorporation of the probe into the secreted mucus was visualized by confocal Z-stacks. In vehicle-treated WT explants, a bright FITC signal was detected within the mucus layer that co-localized with UEA1 lectin staining and overlaid a Calcein violet–labeled viable epithelium, indicating robust extracellular transglutaminase activity at the mucus surface. When the pan-transglutaminase inhibitor Z-DON was added together with the E51 probe, fluorescence in the mucus was markedly reduced, demonstrating that the observed signal reflects enzymatic cross-linking activity within the secreted MUC2 network. Building on this ex vivo set-up, the same study also assessed mucus integrity by tracking the position of fluorescent microparticles before and after protease treatment, revealing that mucus from Tgm3−/− mice deteriorated more rapidly than WT mucus and allowed beads to reach the epithelial surface, consistent with a stabilizing role of TGM3-mediated cross-links in the colonic mucus barrier158.

Tissues were mounted in a perfusion chamber as illustrated in panel (A) and incubated with the glutamine-donor peptide E51 conjugated to FITC (magenta) for 30 min at 37 °C to assess transglutaminase activity. Following the removal of the unbound peptide, tissues were analyzed by confocal microscopy. Mucus and nuclei were counterstained with UEA1 lectin conjugated to rhodamine (green) and Calcein Violet (blue), respectively. Top panels display Z-stack images of explants with (left) and without (right) UEA1 counterstaining. B-D Bottom panels show x/y projections of the highlighted region from the corresponding Z-stacks above. Adapted from ref. 158 under the terms of the Creative Commons CC BY license.
Ex vivo platforms are particularly valuable for evaluating the impact of bioactive substances—such as dietary agents, pharmaceuticals, probiotics, or pathogens—on human-derived tissues, thereby increasing translational relevance. Nonetheless, these models are not without drawbacks, including limited viability of tissue samples, restricted culture times, and inter-donor variability. Despite these constraints, ex vivo approaches provide essential insights into mucus behavior and host–microbe interactions that are often unattainable in purely in vitro systems. Building on this, Sotres et al. contributed significantly to the field by measuring the mechanical properties of intestinal mucus using ex vivo techniques. Their research demonstrated that the mucus layer is both dynamic and responsive to pH changes, capable of engaging with microparticles beyond its surface layer. When applied to ileal mucus, larger silica particles (40 µm) compressed the outer layer, whereas smaller ones (15 µm) were absorbed into it. These observations suggest that the mucus possesses a gradient in pore size ranging between 15 and 40 µm, challenging earlier models that proposed a uniform pore diameter of 0.5–1 µm, and supporting a more complex, stratified architecture designed for selective particle trapping159.
Functional outcomes of polysaccharide–mucus interactions
Modulation of mucus structure and viscosity
Direct physicochemical association of polysaccharides with the mucus gel can rapidly change mucus microstructure and viscoelasticity. The densely packed, viscoelastic nature of the mucin gel network presents a significant challenge for the effective transport of therapeutic compounds—particularly macromolecules and nanoparticles—administered via the oral route160. This glycosylated mucus barrier impedes drug diffusion and limits bioavailability161. Although many conventional immediate-release small-molecule drugs are absorbed predominantly in the proximal small intestine (duodenum and jejunum), the colonic mucus barrier remains highly relevant for oral delivery. Colon-targeted and controlled-release formulations, as well as treatments for local diseases such as inflammatory bowel disease and colorectal cancer, are specifically designed to reach and interact with colonic mucus. To address these challenges, ionic polysaccharides with mucoadhesive characteristics have emerged as promising agents for modulating the intestinal mucus environment to improve drug delivery outcomes75. Cationic polysaccharides such as chitosan can engage in electrostatic interactions with anionic mucins, leading to a temporary loosening of the mucus matrix and improved paracellular permeability162,163. In vivo studies on rats have shown that chitosan treatment notably reduces intestinal mucus viscosity164. Complementary in vitro co-incubation experiments demonstrated a time-dependent decline in viscosity, which was linked to interactions with MUC-2 mucin proteins rather than simple dilution. Chitosan also induced the formation of droplet-like mucus microstructures that promoted enhanced drug transport164. Interestingly, sodium alginate–chitosan nanoparticles carrying a net negative charge penetrated more deeply into the mucus (up to 664 μm), nearly threefold more than their positively charged chitosan-only counterparts. This greater penetration is likely driven by increased electrostatic repulsion from the negatively charged mucin network165. Alginate’s mucoadhesive behavior depends on properties such as molecular weight, chain flexibility, and charge density. High-molecular-weight alginate (~406 kDa) markedly decreased viscosity upon mixing with mucin, possibly due to the formation of tightly packed heterotypic complexes that reduce the overall hydrodynamic volume. Conversely, low-molecular-weight alginate (~4 kDa) caused a slight viscosity increase, suggesting the formation of a more extended network that does not significantly disrupt mucin structure166. In addition to enhancing drug residence time and absorption, mucoadhesive polysaccharides contribute to the repair and stabilization of the mucus barrier, especially in disease states. This dual role—promoting mucosal adhesion and boosting drug uptake—positions ionic polysaccharide-based delivery systems as powerful tools in the design of next-generation oral therapeutics, offering both improved efficacy and protective support for the intestinal mucus layer.
Effects of polysaccharides on mucin secretion and degradation
Beyond these direct biophysical effects, polysaccharides can modulate mucus renewal by influencing mucin secretion, glycosylation, and degradation—either via epithelial/goblet-cell signalling or indirectly via microbiota-derived metabolites. Polysaccharides play a critical role in regulating mucin secretion and maintaining the composition and functionality of the intestinal mucus barrier. Multiple bioactive polysaccharides have demonstrated the capacity to stimulate goblet cells, thereby promoting mucin production. In vitro experiments, for instance, have shown that laminaran enhances the expression of mucin-related proteins in LPS-treated LS174T cells—a widely used goblet cell-like model. This effect involves the upregulation of sulfotransferases and vesicle trafficking proteins such as VAMP8 and SNAP-23, which are integral to mucin secretion, and is largely mediated through the PKC signaling pathway, particularly PKCα and PKCβII108. Similarly, various fructans—including inulin-type and graminan-type with DP 3-60—have been observed to protect LS174T cells from TNFα-induced damage by upregulating MUC2, TFF3, GAL3ST2, and CHST5. These fructans also prevented the suppression of GAL3ST2 by tunicamycin, an inhibitor of N-glycosylation167.
Supporting these in vitro findings, in vivo studies demonstrate that polysaccharides contribute to enhanced mucus production and goblet cell proliferation. For example, Gycyrrhiza polysaccharide significantly elevated goblet cell numbers and mucin levels in the intestinal epithelium of cyclophosphamide-induced immunosuppressed mice168. Likewise, Abelmoschus manihot polysaccharide increased mucus thickness and MUC2 secretion in a dose-dependent manner in DSS-induced colitis models, likely through the activation of endoplasmic reticulum stress-related genes (PERK, ATF4, XBP1s) and O-glycosylation genes (B3GNT6, C1GALT1, and ST3GAL3)127.
Polysaccharides can also exert indirect effects on mucus integrity by modulating the gut microbiota. Table 1 summarizes that neutral, anionic, and cationic polysaccharides modulate the colonic mucus barrier through complementary mechanisms. Dendrobium officinale polysaccharide, for instance, improved the mucus barrier in colitic mice by increasing goblet cell density and enriching Lactobacillales, which produce short-chain fatty acids (SCFAs) that support mucosal health. Further evidence highlights the role of SCFAs—particularly propionate—in enhancing goblet cell differentiation and mucus production169. Wang et al. reported that propionate, generated by Bacteroides through methylmalonyl-CoA mutase, exerted its effects partly via the SCFAs receptor GPR41. In that study, propionate exposure increased MUC2 and other mucus-related gene expression in intestinal organoids, and pharmacological activation of GPR41 recapitulated the barrier-protective effects of propionate, indicating that GPR41-dependent signaling links microbially derived propionate to reinforcement of the mucus barrier170. In addition, dietary polysaccharides such as arabinoxylan and resistant starch shape the gut microbial community by enriching beneficial genera, including Bacteroides, Lactobacillus, Coprococcus, Butyricicoccus, Ruminococcus, and Blautia171. These compositional changes influence bile acid metabolism and further contribute to mucus barrier integrity. Of particular interest are arabinose-rich polysaccharides like pectin and Lycium barbarum arabinogalactan, which have been found to increase the abundance of Akkermansia muciniphila—a mucin-degrading bacterium with paradoxical mucin-stimulatory effects172. A. muciniphila upregulates fucosyltransferase, leading to a shift toward neutral fucosylated O-glycan structures (e.g., core 1 and 2) while reducing sulfated and sialylated glycans. Although this microbe consumes mucin as a carbon source, it also promotes MUC2 expression through indirect interactions with goblet cells while remaining confined to the outer mucus layer. Rather than directly contacting the epithelial surface, A. muciniphila colonizes the outer mucus compartment and releases SCFAs that diffuse through the mucus to activate signaling pathways in goblet cells, thereby upregulating MUC2 transcription and sustaining mucus renewal. Notably, A. muciniphila expresses numerous glycoside hydrolases (GHs), including fucosidases (GH29, GH95) and sialidases (GH33, GHB1), which cleave terminal fucose and sialic acid residues. Interestingly, these liberated sugars are not primarily utilized by A. muciniphila itself but are instead cross-fed to butyrate-producing Clostridia such as Roseburia inulinivorans and Faecalibacterium prausnitzii. This cooperative metabolic interaction supports butyrogenesis, which is associated with anti-inflammatory effects and epithelial homeostasis. Disruption of this syntrophic relationship may explain the reduced abundance of A. muciniphila observed in conditions like inflammatory bowel disease and obesity173.
Potential downstream consequences: immune tone and oxidative stress
Strengthening the mucus barrier—through direct polymer–mucin association and/or indirect increases in mucin output—can secondarily reduce microbial proximity to the epithelium, thereby lowering pattern-recognition receptor stimulation and downstream inflammatory signaling174,175. Conversely, inflammatory and oxidative-stress states can impair goblet-cell function and mucin biosynthesis and accelerate mucin degradation, creating a feed-forward cycle of barrier loss and immune activation. However, in the current literature, many immune or antioxidant polysaccharide studies quantify cytokines and ROS markers without measuring mucus endpoints (e.g., mucus thickness, penetrability, or secreted mucin), and few studies use causal designs (pathway inhibition or metabolite add-back) to demonstrate that immune or oxidative pathways mediate mucus restoration176,177. Therefore, we treat immune and oxidative outcomes as downstream or parallel effects rather than primary mechanisms of polysaccharide–mucus interaction.
Discussion
Looking ahead, progress will depend on harmonizing structural reporting (molecular weight, charge density, and substitution)178, standardizing mucus-sensitive endpoints (rheology, thickness, and penetrability), and validating key findings across complementary platforms (cell-based, organoid/ex vivo, and in vivo) to separate direct mucus biophysics from microbiota-mediated effects179. Such model-bridging designs will accelerate translation toward rationally selected polysaccharides for barrier repair and mucosa-targeted delivery180.
In conclusion, this review provides a comprehensive and mechanistic perspective on the interactions between polysaccharides and the colonic mucus barrier, with a particular focus on the role of polysaccharide charge in determining their behavior and biological function. We have summarized the key biophysical mechanisms—electrostatic attraction, hydrogen bonding, hydrophobic interactions, and van der Waals forces—that govern polysaccharide–mucus binding. We also critically evaluated the advanced experimental platforms used to model these interactions, including in vitro cultures, in vivo animal models, ex vivo tissues, organoids, and gut-on-a-chip technologies. Importantly, we have summarized current evidence on the functional consequences of these interactions, encompassing mucin secretion and degradation, modulation of mucus viscosity and structure, regulation of epithelial and immune responses, and protection against oxidative stress-induced mucosal dysfunction. Together, this integrative review underscores the significance of polysaccharide–mucus interactions and offers a foundational framework to guide future research and the development of next-generation therapeutics targeting the mucosal interface.
Data availability
No datasets were generated or analysed during the current study.
References
Wang, Z. et al. Effect of monosaccharide composition and proportion on the bioactivity of polysaccharides: a review. Int. J. Biol. Macromol. 254, 127955 (2024).
Cheong, K.-L., Chen, W., Wang, M., Zhong, S. & Veeraperumal, S. Therapeutic prospects of Undaria pinnatifida polysaccharides: extraction, purification, and functional activity. Mar. Drugs 23, 163 (2025).
Cheong, K.-L. et al. Trends in polysaccharide-based hydrogels for skin anti-aging and skin antioxidant. Int. J. Biol. Macromol. 319, 145366 (2025).
Ferreira, S. S., Passos, C. P., Madureira, P., Vilanova, M. & Coimbra, M. A. Structure–function relationships of immunostimulatory polysaccharides: a review. Carbohydr. Polym. 132, 378–396 (2015).
Teferra, T. F. Possible actions of inulin as prebiotic polysaccharide: a review. Food Front. 2, 407–416 (2021).
Cheong, K.-L., Sabir, A., Wang, M., Zhong, S. & Tan, K. Advancements in the extraction, characterization, and bioactive potential of laminaran: a review. Foods 14, 1683 (2025).
Ho Do, M., Seul, S. Y. & Park, H.-Y. Polysaccharides: bowel health and gut microbiota. Crit. Rev. Food Sci. Nutr. 61, 1212–1224 (2021).
Johansson, M. E. V. et al. The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc. Natl. Acad. Sci. USA 105, 15064–15069 (2008).
McGuckin, M. A. & Hasnain, S. Z. Goblet cells as mucosal sentinels for immunity. Mucosal Immunol. 10, 1118–1121 (2017).
Stanforth, K. J. et al. The MUC2 gene product: polymerisation and post-secretory organisation—current models. Polymers 16, 1663 (2024).
Liu, Y. et al. Orchestration of MUC2—the key regulatory target of gut barrier and homeostasis: a review. Int. J. Biol. Macromol. 236, 123862 (2023).
Meldrum, O. W., Yakubov, G. E. Journey of dietary fiber along the gastrointestinal tract: role of physical interactions, mucus, and biochemical transformations. Crit. Rev. Food Sci. Nutr. 65, 1–29 (2024).
Albano, K. M., Fazani, C.ÂL. & Nicoletti, V. R. Electrostatic interaction between proteins and polysaccharides: physicochemical aspects and applications in emulsion stabilization. Food Rev. Int. 35, 54–89 (2019).
Ferreira, L. M. B. et al. Understanding mucus modulation behavior of chitosan oligomers and dextran sulfate combining light scattering and calorimetric observations. Carbohydr. Polym. 306, 120613 (2023).
Surendranath, M., M. R, R. & Parameswaran, R. Recent advances in functionally modified polymers for mucoadhesive drug delivery. J. Mater. Chem. B 10, 5913–5924 (2022).
Sauvaitre, T. et al. Tripartite relationship between gut microbiota, intestinal mucus and dietary fibers: towards preventive strategies against enteric infections. FEMS Microbiol. Rev. 45, fuaa052 (2021).
Cai, B. et al. Administration of Gracilariopsis lemaneiformis polysaccharide attenuates cisplatin-induced inflammation and intestinal mucosal damage in colon-26 carcinoma tumor-bearing mice. J. Sci. Food Agric. 104, 3757–3766 (2024).
Rajagopal, H. M., Manjegowda, S. B., Serkad, C. & Dharmesh, S. M. A modified pectic polysaccharide from turmeric (Curcuma longa) with antiulcer effects via anti-secretary, mucoprotective and IL-10 mediated anti-inflammatory mechanisms. Int. J. Biol. Macromol. 118, 864–880 (2018).
Cheong, K.-L., Liu, K., Chen, W., Zhong, S. & Tan, K. Recent progress in Porphyra haitanensis polysaccharides: extraction, purification, structural insights, and their impact on gastrointestinal health and oxidative stress management. Food Chem. X 22, 101414 (2024).
Moulzir, M., Hazart, D., Oheim, M., Ricard, C. Label-free, high-resolution fluorescence imaging of the intestinal wall by image scanning and confocal microscopy. Gastro. Hep. Adv. https://doi.org/10.1016/j.gastha.2025.100735 (2025).
Özkan, A., LoGrande, N. T., Feitor, J. F., Goyal, G. & Ingber, D. E. Intestinal organ chips for disease modelling and personalized medicine. Nat. Rev. Gastro. Hepat. 21, 751–773 (2024).
Gustafsson, J. K. et al. An ex vivo method for studying mucus formation, properties, and thickness in human colonic biopsies and mouse small and large intestinal explants. Am. J. Physiol. Gastrointest. Liver Physiol. 302, G430–G438 (2011).
Johansson, M. E. et al. Bacteria penetrate the normally impenetrable inner colon mucus layer in both murine colitis models and patients with ulcerative colitis. Gut 63, 281–291 (2014).
Schroeder, B. O. Fight them or feed them: how the intestinal mucus layer manages the gut microbiota. Gastroenterol. Rep. 7, 3–12 (2019).
Cai, R. et al. Interactions of commensal and pathogenic microorganisms with the mucus layer in the colon. Gut Microbes 11, 680–690 (2020).
Yu, B. et al. Partially acid-hydrolyzed porphyran improved dextran sulfate sodium-induced acute colitis by modulation of gut microbiota and enhancing the mucosal barrier. J. Agric. Food Chem. 71, 7299–7311 (2023).
Martens, E. C., Neumann, M. & Desai, M. S. Interactions of commensal and pathogenic microorganisms with the intestinal mucosal barrier. Nat. Rev. Microbiol. 16, 457–470 (2018).
Li, H. et al. The outer mucus layer hosts a distinct intestinal microbial niche. Nat. Commun. 6, 8292 (2015).
Aggarwal, V., Sunder, S. & Verma, S. R. Disease-associated dysbiosis and potential therapeutic role of Akkermansia muciniphila, a mucus degrading bacteria of gut microbiome. Folia Microbiol. 67, 811–824 (2022).
Ndeh, D. A. et al. A Bacteroides thetaiotaomicron genetic locus encodes activities consistent with mucin O-glycoprotein processing and N-acetylgalactosamine metabolism. Nat. Commun. 16, 3485 (2025).
Herath, M., Hosie, S., Bornstein, J. C., Franks, A. E. & Hill-Yardin, E. L. The role of the gastrointestinal mucus system in intestinal homeostasis: implications for neurological disorders. Front. Cell Infect. Mi. ume 10, 2020 (2020).
Birchenough, G. M. H., Johansson, M. E., Gustafsson, J. K., Bergström, J. H. & Hansson, G. C. New developments in goblet cell mucus secretion and function. Mucosal Immunol. 8, 712–719 (2015).
Kudelka, M. R., Stowell, S. R., Cummings, R. D. & Neish, A. S. Intestinal epithelial glycosylation in homeostasis and gut microbiota interactions in IBD. Nat. Rev. Gastro. Hepat. 17, 597–617 (2020).
Recktenwald, C. V. et al. The structure of the second CysD domain of MUC2 and role in mucin organization by transglutaminase-based cross-linking. Cell Rep. https://doi.org/10.1016/j.celrep.2024.114207 (2024).
Song, C. et al. Intestinal mucus components and secretion mechanisms: what we do and do not know. Exp. Mol. Med. 55, 681–691 (2023).
Cornick, S., Kumar, M., Moreau, F., Gaisano, H. & Chadee, K. VAMP8-mediated MUC2 mucin exocytosis from colonic goblet cells maintains innate intestinal homeostasis. Nat. Commun. 10, 4306 (2019).
Cornick, S., Moreau, F., Gaisano, H. Y., Chadee, K. Entamoeba histolytica-induced mucin exocytosis is mediated by VAMP8 and is critical in mucosal innate host defense. mBio. https://doi.org/10.1128/mbio.01323-17 (2017).
Kurzyna-Szklarek, M., Cybulska, J. & Zdunek, A. Analysis of the chemical composition of natural carbohydrates—an overview of methods. Food Chem. 394, 133466 (2022).
Ji, X., Yin, M., Nie, H. & Liu, Y. A Review of isolation, chemical properties, and bioactivities of polysaccharides from Bletilla striata. Biomed. Res. Int. 2020, 5391379 (2020).
Wang, M. & Cheong, K.-L. Preparation, structural characterisation, and bioactivities of fructans: a review. Molecules 28, 1613 (2023).
Ji, X. et al. Effect of inulin on pasting, thermal, rheological properties and in vitro digestibility of pea starch gel. Int. J. Biol. Macromol. 193, 1669–1675 (2021).
Yiasmin, M.N. et al. The state-of-the-art on exploring polysaccharide-protein interactions and its mechanisms, stability, and their role in food systems. Food Rev. Int. 41, 1–36 (2025).
Fuchs, A., Romeis, D., Hupfeld, E. & Sieber, V. Biocatalytic conversion of carrageenans for the production of 3,6-anhydro-D-galactose. J. Agric. Food Chem. 72, 5816–5827 (2024).
Hong, T., Zhao, J., Yin, J., Nie, S., Xie, M. Structural characterization of a low molecular weight HG-type pectin from gougunao green tea. Front. Nutr. 9, 2022 (2022).
Liu, K. et al. Pitaya stem polysaccharide promotes wound healing by modulating macrophage polarization via single-cell RNA sequencing evidence. Int. J. Biol. Macromol. 308, 142653 (2025).
An, J. et al. Influence of glycosylation on interfacial properties of recombinant mucins: adsorption, surface forces, and friction. Langmuir 33, 4386–4395 (2017).
Johansson, M. E. V. et al. Bacteria penetrate the inner mucus layer before inflammation in the dextran sulfate colitis model. PLoS One 5, e12238 (2010).
de Almeida, W. S. & da Silva, D. A. Does polysaccharide quaternization improve biological activity?. Int. J. Biol. Macromol. 182, 1419–1436 (2021).
Liu, H. et al. Positively charged chitosan and N-trimethyl chitosan inhibit Aβ40 fibrillogenesis. Biomacromolecules 16, 2363–2373 (2015).
Corfield, A. P. Mucins: a biologically relevant glycan barrier in mucosal protection. Biochim. Biophys. Acta Gen. Subj. 1850, 236–252 (2015).
Johansson, M. E. V. et al. Composition and functional role of the mucus layers in the intestine. Cell. Mol. Life Sci. 68, 3635–3641 (2011).
Shon, D. J., Kuo, A., Ferracane, M. J. & Malaker, S. A. Classification, structural biology, and applications of mucin domain-targeting proteases. Biochem. J. 478, 1585–1603 (2021).
Liu, Y. et al. The role of MUC2 mucin in intestinal homeostasis and the impact of dietary components on MUC2 expression. Int. J. Biol. Macromol. 164, 884–891 (2020).
Sun, L., Zhang, Y., Li, W., Zhang, J. & Zhang, Y. Mucin glycans: a target for cancer therapy. Molecules 28, 7033 (2023).
Arike, L. & Hansson, G. C. The densely O-glycosylated MUC2 mucin protects the intestine and provides food for the commensal bacteria. J. Mol. Biol. 428, 3221–3229 (2016).
Sanz-Martinez, I., Pereira, S., Merino, P., Corzana, F. & Hurtado-Guerrero, R. Molecular recognition of GalNAc in mucin-type O-glycosylation. Acc. Chem. Res. 56, 548–560 (2023).
Liu, J., Jin, C., Cherian, R. M., Karlsson, N. G. & Holgersson, J. O-glycan repertoires on a mucin-type reporter protein expressed in CHO cell pools transiently transfected with O-glycan core enzyme cDNAs. J. Biotechnol. 199, 77–89 (2015).
Ye, J. et al. Core 2 mucin-type O-glycan inhibits EPEC or EHEC O157:H7 invasion into HT-29 epithelial cells. Gut Pathog. 7, 31 (2015).
Holmén Larsson, J. M., Thomsson, K. A., Rodríguez-Piñeiro, A. M., Karlsson, H. & Hansson, G. C. Studies of mucus in mouse stomach, small intestine, and colon. III. Gastrointestinal Muc5ac and Muc2 mucin O-glycan patterns reveal a regiospecific distribution. Am. J. Physiol. Gastrointest. Liver Physiol. 305, G357–G363 (2013).
Zhang, Y. et al. Mucin-type O-glycans: barrier, microbiota, and immune anchors in inflammatory bowel disease. J. Inflamm. Res. 14, 5939–5953 (2021).
Dupont, A., Lena, H., Klaus, B. & Hornef, M. W. Antimicrobial peptides and the enteric mucus layer act in concert to protect the intestinal mucosa. Gut Microbes 5, 761–765 (2014).
Cardoso, M. H., Meneguetti, B. T., Oliveira-Júnior, N. G., Macedo, M. L. R. & Franco, O. L. Antimicrobial peptide production in response to gut microbiota imbalance. Peptides 157, 170865 (2022).
Bamias, G., Kitsou, K. & Rivera-Nieves, J. The underappreciated role of secretory IgA in IBD. Inflamm. Bowel Dis. 29, 1327–1341 (2023).
Rondelli, V. et al. Mucin thin layers: a model for mucus-covered tissues. Int. J. Mol. Sci. 20, 3712 (2019).
Leon-Coria, A., Manish, K. & Chadee, K. The delicate balance between Entamoeba histolytica, mucus and microbiota. Gut Microbes 11, 118–125 (2020).
Einhorn, V., Haase, H. & Maares, M. Interaction and competition for intestinal absorption by zinc, iron, copper, and manganese at the intestinal mucus layer. J. Trace Elem. Med. Biol. 84, 127459 (2024).
Bayer, I. S. Recent advances in mucoadhesive interface materials, mucoadhesion characterization, and technologies. Adv. Mater. Interfaces 9, 2200211 (2022).
Kim, J. et al. N-glycan modifications with negative charge in a natural polymer mucin from bovine submaxillary glands, and their structural role. Polymers 13, 103 (2021).
Ding, L., Huang, Y., Cai, X. & Wang, S. Impact of pH, ionic strength and chitosan charge density on chitosan/casein complexation and phase behavior. Carbohydr. Polym. 208, 133–141 (2019).
Wang, J. & Zhuang, S. Chitosan-based materials: preparation, modification and application. J. Clean. Prod. 355, 131825 (2022).
Liu, X., Dong, Y., Wang, C. & Guo, Z. Application of chitosan as nano carrier in the treatment of inflammatory bowel disease. Int. J. Biol. Macromol. 278, 134899 (2024).
Ahmad, M., Ritzoulis, C. & Chen, J. Shear and extensional rheological characterisation of mucin solutions. Colloids Surf. B. Biointerfaces 171, 614–621 (2018).
Wang, L., Wu, Y., Li, J., Qiao, H. & Di, L. Rheological and mucoadhesive properties of polysaccharide from Bletilla striata with potential use in pharmaceutics as bio-adhesive excipient. Int. J. Biol. Macromol. 120, 529–536 (2018).
Menchicchi, B. et al. Structure of chitosan determines its interactions with mucin. Biomacromolecules 15, 3550–3558 (2014).
Yermak, I. M., Davydova, V. N. & Volod’ko, A. V. Mucoadhesive marine polysaccharides. Mar. Drugs 20, 522 (2022).
Wu, X. et al. Na+/K+ enhanced the stability of the air/water interface of soy hull polysaccharide and intestinal mucus. Int. J. Biol. Macromol. 245, 125206 (2023).
M. Ways, T. M., Lau, W. M. & Khutoryanskiy, V. V. Chitosan and its derivatives for application in mucoadhesive drug delivery systems. Polymers 10, 267 (2018).
Boedtkjer, E. & Pedersen, S. F. The acidic tumor microenvironment as a driver of cancer. Annu. Rev. Physiol. 82, 103–126 (2020).
Hajjar, S. & Zhou, X. pH sensing at the intersection of tissue homeostasis and inflammation. Trends Immunol. 44, 807–825 (2023).
Abu Elella, M. H. & Kolawole, O. M. Recent advances in modified chitosan-based drug delivery systems for transmucosal applications: a comprehensive review. Int. J. Biol. Macromol. 277, 134531 (2024).
Bal, K. et al. Recent progress in chitosan-based nanoparticles for drug delivery: a review on modifications and therapeutic potential. J. Drug Target. 33, 1–28 (2025).
Yang, F. et al. Oral administration of nanoemulsion of supercritical fluid extract of Angelica with quaternized chitosan alleviates ulcerative colitis in mice. Int. J. Biol. Macromol. 314, 144339 (2025).
de Oliveira Cardoso, V. M., Gremião, M. P. D. & Cury, B. S. F. Mucin-polysaccharide interactions: a rheological approach to evaluate the effect of pH on the mucoadhesive properties. Int. J. Biol. Macromol. 149, 234–245 (2020).
Chen, A. et al. Chain conformation, mucoadhesive properties of fucoidan in the gastrointestinal tract and its effects on the gut microbiota. Carbohydr. Polym. 304, 120460 (2023).
An, S. et al. Construction of Hohenbuehelia serotina polysaccharides-mucin nanoparticles and their sustain-release characteristics under simulated gastrointestinal digestion in vitro. Int. J. Biol. Macromol. 191, 1–8 (2021).
Yermak, I. M. et al. Mucoadhesive properties of sulphated polysaccharides carrageenans from red seaweed families Gigartinaceae and Tichocarpaceae. Int. J. Biol. Macromol. 142, 634–642 (2020).
Schattling, P., Taipaleenmäki, E., Zhang, Y. & Städler, B. A polymer chemistry point of view on mucoadhesion and mucopenetration. Macromol. Biosci. 17, 1700060 (2017).
Wang, X. et al. Acetylated polysaccharides: synthesis, physicochemical properties, bioactivities, and food applications. Crit. Rev. Food Sci. Nutr. 64, 4849–4864 (2024).
Kashapov, R. et al. Self-assembly of amphiphilic compounds as a versatile tool for construction of nanoscale drug carriers. Int. J. Mol. Sci. 21, 6961 (2020).
Käsdorf, B. T. et al. Mucin-inspired lubrication on hydrophobic surfaces. Biomacromolecules 18, 2454–2462 (2017).
Prasher, P. et al. Current-status and applications of polysaccharides in drug delivery systems. Colloid Interface Sci. Commun. 42, 100418 (2021).
Witten, J., Samad, T. & Ribbeck, K. Selective permeability of mucus barriers. Curr. Opin. Biotechnol. 52, 124–133 (2018).
Ho, D.-K. et al. Farnesylated glycol chitosan as a platform for drug delivery: synthesis, characterization, and investigation of mucus–particle interactions. Biomacromolecules 19, 3489–3501 (2018).
Ruiz-Pulido, G., Quintanar-Guerrero, D., Serrano-Mora, L. E. & Medina, D. I. Triborheological analysis of reconstituted gastrointestinal mucus/chitosan:TPP nanoparticles system to study mucoadhesion phenomenon under different pH conditions. Polymers 14, 4978 (2022).
McClements, D. J. Encapsulation, protection, and release of hydrophilic active components: Potential and limitations of colloidal delivery systems. Adv. Colloid Interface Sci. 219, 27–53 (2015).
Wijekoon, M. M. J. O., Mahmood, K., Ariffin, F., Mohammadi Nafchi, A. & Zulkurnain, M. Recent advances in encapsulation of fat-soluble vitamins using polysaccharides, proteins, and lipids: a review on delivery systems, formulation, and industrial applications. Int. J. Biol. Macromol. 241, 124539 (2023).
Vaziri, A. S., Alemzadeh, I., Vossoughi, M. & Khorasani, A. C. Co-microencapsulation of Lactobacillus plantarum and DHA fatty acid in alginate-pectin-gelatin biocomposites. Carbohydr. Polym. 199, 266–275 (2018).
Geng, W.-C., Jiang, Z.-T., Chen, S.-L. & Guo, D.-S. Supramolecular interaction in the action of drug delivery systems. Chem. Sci. 15, 7811–7823 (2024).
Watchorn, J., Stuart, S., Burns, D. C. & Gu, F. X. Mechanistic influence of polymer species, molecular weight, and functionalization on mucin–polymer binding interactions. ACS Appl. Polym. Mater. 4, 7537–7546 (2022).
Pecora, T. M. G. et al. Rheological behavior of a new mucoadhesive oral formulation based on sodium chondroitin sulfate, xyloglucan and glycerol. J. Funct. Biomater. 12, 28 (2021).
Periasamy, S. et al. Mucoadhesive role of tamarind xyloglucan on inflammation attenuates ulcerative colitis. J. Funct. Food 47, 1–10 (2018).
Joshi, A., Soni, A. & Acharya, S. In vitro models and ex vivo systems used in inflammatory bowel disease. Vitr. Models 1, 213–227 (2022).
Jung, T.-H., Han, K.-S., Park, J.-H. & Hwang, H.-J. Butyrate modulates mucin secretion and bacterial adherence in LoVo cells via MAPK signaling. PLoS One 17, e0269872 (2022).
Cai, Y. et al. SEDDS facilitate cinnamaldehyde crossing the mucus barrier: The perspective of mucus and Caco-2/HT29 co-culture models. Int. J. Pharm. 614, 121461 (2022).
Bu, X.-D., Li, N., Tian, X.-Q. & Huang, P.-L. Caco-2 and LS174T cell lines provide different models for studying mucin expression in colon cancer. Tissue Cell 43, 201–206 (2011).
Wu, C. et al. Modulation of intestinal epithelial permeability and mucin mRNA (MUC2, MUC5AC, and MUC5B) expression and protein secretion in Caco-2/HT29-MTX co-cultures exposed to aflatoxin M1, ochratoxin A, and zearalenone individually or collectively. Toxicol. Lett. 309, 1–9 (2019).
McCoy, R. et al. In vitro models for investigating intestinal host–pathogen interactions. Adv. Sci. 11, 2306727 (2024).
Cheong, K.-L. et al. Structure–function relationship of the brown seaweed Undaria pinnatifida laminaran: protein kinase C-mediated mucus secretion and gut barrier restoration. Carbohydr. Polym. 358, 123525 (2025).
Kootala, S. et al. Reinforcing mucus barrier properties with low molar mass chitosans. Biomacromolecules 19, 872–882 (2018).
Floor, E. et al. Development of a Caco-2-based intestinal mucosal model to study intestinal barrier properties and bacteria–mucus interactions. Gut Microbes 17, 2434685 (2025).
Zhao, T. et al. Influence of in vitro pectin fermentation on the human fecal microbiome and O-glycosylation of HT29-MTX cells. Int. J. Biol. Macromol. 284, 137710 (2025).
Giromini, C. et al. Role of short chain fatty acids to counteract inflammatory stress and mucus production in human intestinal HT29-MTX-E12 cells. Foods 11, 2022 (1983).
Kajiwara-Kubota, M. et al. Partially hydrolyzed guar gum increased colonic mucus layer in mice via succinate-mediated MUC2 production. npj Sci. Food 7, 10 (2023).
Hu, W. et al. Endotoxins induced ECM-receptor interaction pathway signal effect on the function of MUC2 in Caco2/HT29 co-culture cells. Front. Immunol. 13, 916933 (2022).
Barnett, A. M., Roy, N. C., Cookson, A. L. & McNabb, W. C. Metabolism of caprine milk carbohydrates by probiotic bacteria and Caco-2:HT29–MTX epithelial co-cultures and their impact on intestinal barrier integrity. Nutrients 10, 949 (2018).
Meldrum, O. W. & Yakubov, G. E. Journey of dietary fiber along the gastrointestinal tract: role of physical interactions, mucus, and biochemical transformations. Crit. Rev. Food Sci. Nutr. 65, 4264–4292 (2025).
Sharma, T. T., Rabizadeh, R. R., Prabhakar, V. S., Bury, M. I. & Sharma, A. K. Evolving experimental platforms to evaluate ulcerative colitis. Adv. Biol. 6, 2200018 (2022).
Ruff, W. E., Greiling, T. M. & Kriegel, M. A. Host–microbiota interactions in immune-mediated diseases. Nat. Rev. Microbiol. 18, 521–538 (2020).
Johansson, M. E. V. & Hansson, G. C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 16, 639–649 (2016).
Xia, B. et al. Mucin O-glycan-microbiota axis orchestrates gut homeostasis in a diarrheal pig model. Microbiome 10, 139 (2022).
Zang, P. et al. Alteration of gastrointestinal function and the ameliorative effects of Hericium erinaceus polysaccharides in tail suspension rats. Nutrients 17, 724 (2025).
Wang, Y. et al. Exploring effects of chitosan oligosaccharides on the DSS-induced intestinal barrier impairment in vitro and in vivo. Molecules 26, 2199 (2021).
Jiang, S., Xu, H., Zhao, C., Zhong, F. & Li, D. Oyster polysaccharides relieve DSS-induced colitis via anti-inflammatory and maintaining the physiological hypoxia. Int. J. Biol. Macromol. 238, 124150 (2023).
Li, M. et al. Combined Ganoderma lucidum polysaccharide and ciprofloxacin therapy alleviates Salmonella enterica infection, protects the intestinal barrier, and regulates gut microbiota. Food Funct. 14, 6896–6913 (2023).
Fan, X. et al. Intravital imaging of translocated bacteria via fluorogenic labeling of gut microbiota in situ. Proc. Natl. Acad. Sci. USA 122, e2415845122 (2025).
Xiao, H. et al. Tremella fuciformis polysaccharides ameliorated ulcerative colitis via inhibiting inflammation and enhancing intestinal epithelial barrier function. Int. J. Biol. Macromol. 180, 633–642 (2021).
Wang, Y. et al. Abelmoschus manihot polysaccharide fortifies intestinal mucus barrier to alleviate intestinal inflammation by modulating Akkermansia muciniphila abundance. Acta Pharm. Sin. B 14, 3901–3915 (2024).
Manning, J. C. et al. Lectins: a primer for histochemists and cell biologists. Histochem. Cell Biol. 147, 199–222 (2017).
Wu, A. M., Dudek, A. & Chen, Y. L. Recognition factors of Dolichos biflorus agglutinin (DBA) and their accommodation sites. Glycoconj. J. 40, 383–399 (2023).
Lastovickova, M., Strouhalova, D. & Bobalova, J. Use of lectin-based affinity techniques in breast cancer glycoproteomics: a review. J. Proteome Res. 19, 1885–1899 (2020).
Engevik, M. A. et al. Human Clostridium difficile infection: altered mucus production and composition. Am. J. Physiol. Gastrointest. Liver Physiol. 308, G510–G524 (2014).
Wen, X. et al. Pectin supplementation ameliorates intestinal epithelial barrier function damage by modulating intestinal microbiota in lipopolysaccharide-challenged piglets. J. Nutr. Biochem. 109, 109107 (2022).
Bergstrom, K. et al. Core 1- and 3-derived O-glycans collectively maintain the colonic mucus barrier and protect against spontaneous colitis in mice. Mucosal Immunol. 10, 91–103 (2017).
Nyström, E. E. L. et al. An intercrypt subpopulation of goblet cells is essential for colonic mucus barrier function. Science 372, eabb1590 (2021).
Singh, V. et al. Chronic inflammation in ulcerative colitis causes long-term changes in goblet cell function. Cell. Mol. Gastroenterol. Hepatol. 13, 219–232 (2022).
Ma, J. et al. Modified Gegen Qinlian decoction ameliorates DSS-induced chronic colitis in mice by restoring the intestinal mucus barrier and inhibiting the activation of γδT17 cells. Phytomedicine 111, 154660 (2023).
Nomura, E. et al. Characteristics of the mucosal surface on scanning electron microscopy in patients with remitting ulcerative colitis. Dig. Dis. Sci. 66, 3141–3148 (2021).
Taniguchi, M. et al. Sialylation shapes mucus architecture inhibiting bacterial invasion in the colon. Mucosal Immunol. 16, 624–641 (2023).
Meziu, E. et al. Visualization of the structure of native human pulmonary mucus. Int. J. Pharm. 597, 120238 (2021).
Almeqdadi, M., Mana, M. D., Roper, J. & Yilmaz, ÖH. Gut organoids: mini-tissues in culture to study intestinal physiology and disease. Am. J. Physiol. Cell Physiol. 317, C405–C419 (2019).
Mithal, A. et al. Generation of mesenchyme free intestinal organoids from human induced pluripotent stem cells. Nat. Commun. 11, 215 (2020).
Villegas-Novoa, C., Wang, Y., Sims, C. E. & Allbritton, N. L. Creation of a spatially complex mucus bilayer on an in vitro colon model. Sci. Rep. 14, 16849 (2024).
Zhang, D., Zhou, X., Zhou, W., Cui, S. W. & Nie, S. Intestinal organoids: a thriving and powerful tool for investigating dietary nutrients-intestinal homeostasis axis. Food Res. Int. 172, 113109 (2023).
Bozzetti, V. & Senger, S. Organoid technologies for the study of intestinal microbiota–host interactions. Trends Mol. Med. 28, 290–303 (2022).
Liu, Y. et al. Ginseng-derived nanocellulose alleviates murine colitis through modulation of inflammation, epithelial barrier and gut microbiome. Chem. Eng. J. 506, 160254 (2025).
Poletti, M., Arnauts, K., Ferrante, M. & Korcsmaros, T. Organoid-based models to study the role of host-microbiota interactions in IBD. J. Crohn’s. Colitis 15, 1222–1235 (2021).
Son, Y. S. et al. Maturation of human intestinal organoids in vitro facilitates colonization by commensal lactobacilli by reinforcing the mucus layer. FASEB J. 34, 9899–9910 (2020).
Yang, N. et al. A mucus layer derived from porcine intestinal organoid air–liquid interface monolayer attenuates swine enteric coronavirus infection by antiviral activity of Muc2. BMC Biol. 22, 297 (2024).
Farhang Doost, N. & Srivastava, S. K. A comprehensive review of organ-on-a-chip technology and its applications. Biosensors 14, 225 (2024).
Ashammakhi, N. et al. Gut-on-a-chip: current progress and future opportunities. Biomaterials 255, 120196 (2020).
Nelson, M. T. et al. Evaluation of human performance aiding live synthetically engineered bacteria in a gut-on-a-chip. ACS Biomater. Sci. Eng. 9, 5136–5150 (2023).
Thomas, D. P., Zhang, J., Nguyen, N.-T. & Ta, H. T. Microfluidic gut-on-a-chip: fundamentals and challenges. Biosensors 13, 136 (2023).
Wang, H., Cong, X., Mingqian, T. & Su, W. Advanced gut-on-chips for assessing carotenoid absorption, metabolism, and transport. Crit. Rev. Food Sci. Nutr. 65, 1344–1362 (2025).
Garcia-Gutierrez, E. & Cotter, P. D. Relevance of organ(s)-on-a-chip systems to the investigation of food-gut microbiota-host interactions. Crit. Rev. Microbiol. 48, 463–488 (2022).
Shin, Y. C., Than, N., Min, S., Shin, W. & Kim, H. J. Modelling host–microbiome interactions in organ-on-a-chip platforms. Nat. Rev. Bioeng. 2, 175–191 (2024).
Jing, B. et al. Chitosan oligosaccharides regulate the occurrence and development of enteritis in a human gut-on-a-chip. Front. Cell Dev. Biol. 10, 877892 (2022).
McCoy, R. et al. Mucus-on-a-chip: investigating the barrier properties of mucus with organic bioelectronics. J. Mater. Chem. B 13, 577–587 (2025).
Sharpen, J. D. A. et al. Transglutaminase 3 crosslinks the secreted gel-forming mucus component Mucin-2 and stabilizes the colonic mucus layer. Nat. Commun. 13, 45 (2022).
Sotres, J., Jankovskaja, S., Wannerberger, K. & Arnebrant, T. Ex-vivo force spectroscopy of intestinal mucosa reveals the mechanical properties of mucus blankets. Sci. Rep. 7, 7270 (2017).
Abrami, M. et al. Mucus structure, viscoelastic properties, and composition in chronic respiratory diseases. Int. J. Mol. Sci. 25, 1933 (2024).
Liu, C., Jiang, X., Gan, Y. & Yu, M. Engineering nanoparticles to overcome the mucus barrier for drug delivery: design, evaluation and state-of-the-art. Med. Drug Discov. 12, 100110 (2021).
Mura, P., Maestrelli, F., Cirri, M. & Mennini, N. Multiple roles of chitosan in mucosal drug delivery: an updated review. Mar. Drugs 20, 335 (2022).
Collado-González, M., González Espinosa, Y. & Goycoolea, F. M. Interaction between chitosan and mucin: fundamentals and applications. Biomimetics 4, 32 (2019).
Qi, Y. et al. Chitooligosaccharide reconstitutes intestinal mucus layer to improve oral absorption of water-soluble drugs. J. Control. Release 360, 831–841 (2023).
Sun, L. et al. Mucus-penetrating alginate-chitosan nanoparticles loaded with berberine hydrochloride for oral delivery to the inflammation site of ulcerative colitis. AAPS PharmSciTech 23, 179 (2022).
Menchicchi, B. et al. Biophysical analysis of the molecular interactions between polysaccharides and mucin. Biomacromolecules 16, 924–935 (2015).
Fernández-Lainez, C., aan de Stegge, M., Silva-Lagos, L. A., López-Velázquez, G. & de Vos, P. β(2 → 1)-β(2 → 6) branched graminan-type fructans and β(2 → 1) linear fructans impact mucus-related and endoplasmic reticulum stress-related genes in goblet cells and attenuate inflammatory responses in a fructan dependent fashion. Food Funct. 14, 1338–1348 (2023).
Song, W. et al. Modulating the gut microbiota is involved in the effect of low-molecular-weight Glycyrrhiza polysaccharide on immune function. Gut Microbes 15, 2276814 (2023).
Zeng, X. et al. Analysis of metagenome and metabolome disclosed the mechanisms of Dendrobium officinale polysaccharide on DSS-induced ulcerative colitis-affected mice. Int. J. Biol. Macromol. 277, 134229 (2024).
Wang, X. et al. Bacteroides methylmalonyl-CoA mutase produces propionate that promotes intestinal goblet cell differentiation and homeostasis. Cell Host Microbe 32, 63–78.e7 (2024).
Yang, J., Qin, K., Sun, Y. & Yang, X. Microbiota-accessible fiber activates short-chain fatty acid and bile acid metabolism to improve intestinal mucus barrier in broiler chickens. Microbiol. Spectr. 12, e02065-23 (2023).
Zhao, T. et al. Impact of structurally diverse polysaccharides on colonic mucin O-glycosylation and gut microbiota. npj Biofilms Microbiomes 9, 97 (2023).
Shuoker, B. et al. Sialidases and fucosidases of Akkermansia muciniphila are crucial for growth on mucin and nutrient sharing with mucus-associated gut bacteria. Nat. Commun. 14, 1833 (2023).
Hyun, G. H. et al. Mechanisms of interactions in pattern-recognition of common glycostructures across pectin-derived heteropolysaccharides by Toll-like receptor 4. Carbohydr. Polym. 314, 120921 (2023).
Zhang, X., Qi, C., Guo, Y., Zhou, W. & Zhang, Y. Toll-like receptor 4-related immunostimulatory polysaccharides: primary structure, activity relationships, and possible interaction models. Carbohydr. Polym. 149, 186–206 (2016).
Li, G. et al. Dietary polysaccharides from dragon fruit pomace, a co-product of the fruit processing industry, exhibit therapeutic potential in high-fat diet-induced metabolic disorders. Food Res. Int. 203, 115818 (2025).
Zhao, Z.-L. et al. Protective effect of mussel polysaccharide on cyclophosphamide-induced intestinal oxidative stress injury via Nrf2-Keap1 signaling pathway. Food Sci. Nutr. 11, 4233–4245 (2023).
Kurl, S. et al. Challenges, opportunities, and future prospects of polysaccharide-based nanoparticles for colon targeting: a comprehensive review. Carbohydr. Polym. Technolo. Appl. 6, 100361 (2023).
dos Santos, A. M. et al. Oral delivery of micro/nanoparticulate systems based on natural polysaccharides for intestinal diseases therapy: challenges, advances and future perspectives. J. Control. Release 334, 353–366 (2021).
Cui, M., Zhang, M. & Liu, K. Colon-targeted drug delivery of polysaccharide-based nanocarriers for synergistic treatment of inflammatory bowel disease: a review. Carbohydr. Polym. 272, 118530 (2021).
Acknowledgements
This work was supported by the Natural Science Foundation of Guangdong Province (2025A1515010078) and Key Research Project of Higher Education of Guangdong Province (2024ZDZX2084).
Author information
Authors and Affiliations
Contributions
Kit-Leong Cheong: writing—original draft, investigation, and visualization. Eric Biney and Malairaj Sathuvan: investigation, writing—review and editing, and validation. Min Wang, Hai-Jing Zhong, and Saiyi Zhong: supervision, funding acquisition, and writing—review and editing.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Cheong, KL., Biney, E., Wang, M. et al. Polysaccharides and the colon mucus barrier: a review of biophysical interactions and functional impacts. npj Sci Food 10, 98 (2026). https://doi.org/10.1038/s41538-026-00750-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41538-026-00750-6
