
Abstract
As immune checkpoint blockade induces durable responses in only a subset of patients, more effective immunotherapies are needed. Here we present bispecific antibody engagers, fusion proteins composed of a nanobody that recognizes immunoglobulin kappa light chains (VHHkappa) and a nanobody that recognizes either CTLA-4 or PD-L1. These fusions show strong antitumour activity in mice through recruitment of polyclonal immunoglobulins independently of specificity or isotype. The anti-CTLA-4 VHH-VHHkappa conjugate demonstrates superior antitumour activity compared with the conventional monoclonal anti-CTLA-4 antibody and reduces the number of intratumoural regulatory T cells in a mouse model of colorectal carcinoma. The anti-PD-L1 VHH-VHHkappa conjugate is less effective in the colorectal carcinoma model while still outperforming a conventional antibody of similar specificity. The potency of the anti-PD-L1 VHH-VHHkappa conjugate was enhanced by installation of the cytotoxic drug maytansine or a STING agonist. The ability of such fusions to engage the Fc-mediated functions of all immunoglobulin isotypes is an appealing strategy to further improve on the efficacy of immune checkpoint blockade, commonly delivered as a monoclonal immunoglobulin of a single defined isotype.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others
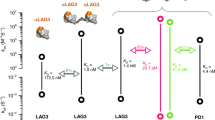

Data availability
All data required to evaluate the conclusions in this paper are included in the main text or Supplementary Information. All unique materials used, including nanobodies and nanobody drug adducts, are available from the authors upon reasonable request. The crystal structures of mouse and human PD-L1 were obtained from the Protein Data Bank (PDB IDs 6SRU and 5XXY, respectively). Source data are provided with this paper.
References
Jiang, X.-R. et al. Advances in the assessment and control of the effector functions of therapeutic antibodies. Nat. Rev. Drug Discov. 10, 101–111 (2011).
Park, H. I., Yoon, H. W. & Jung, S. T. The highly evolvable antibody Fc domain. Trends Biotechnol. 34, 895–908 (2016).
Liu, R., Oldham, R. J., Teal, E., Beers, S. A. & Cragg, M. S. Fc-engineering for modulated effector functions—improving antibodies for cancer treatment. Antibodies 9, 64 (2020).
Kretschmer, A., Schwanbeck, R., Valerius, T. & Rösner, T. Antibody isotypes for tumor immunotherapy. Transfus. Med. Hemotherapy 44, 320–326 (2017).
Mukai, K., Tsai, M., Starkl, P., Marichal, T. & Galli, S. J. IgE and mast cells in host defense against parasites and venoms. Semin. Immunopathol. 38, 581–603 (2016).
Ehrenstein, M. R. & Notley, C. A. The importance of natural IgM: scavenger, protector and regulator. Nat. Rev. Immunol. 10, 778–786 (2010).
DiLillo, D. J. & Ravetch, J. V. Fc-receptor interactions regulate both cytotoxic and immunomodulatory therapeutic antibody effector functions. Cancer Immunol. Res. 3, 704–713 (2015).
Dahan, R. & Ravetch, J. V. Co-targeting of adenosine signaling pathways for immunotherapy: potentiation by Fc receptor engagement. Cancer Cell 30, 369–371 (2016).
Fuentes-Antrás, J., Genta, S., Vijenthira, A. & Siu, L. L. Antibody–drug conjugates: in search of partners of choice. Trends Cancer 9, 339–354 (2023).
Sharma, P. & Allison, J. P. The future of immune checkpoint therapy. Science 348, 56–61 (2015).
Zou, W., Wolchok, J. D. & Chen, L. PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: mechanisms, response biomarkers, and combinations. Sci. Transl. Med. 8, 328rv324–328rv324 (2016).
Simpson, T. R. et al. Fc-dependent depletion of tumor-infiltrating regulatory T cells co-defines the efficacy of anti–CTLA-4 therapy against melanoma. J. Exp. Med. 210, 1695–1710 (2013).
Bulliard, Y. et al. Activating Fc γ receptors contribute to the antitumor activities of immunoregulatory receptor-targeting antibodies. J. Exp. Med. 210, 1685–1693 (2013).
Selby, M. J. et al. Anti-CTLA-4 antibodies of IgG2a isotype enhance antitumor activity through reduction of intratumoral regulatory T cells. Cancer Immunol. Res. 1, 32–42 (2013).
Vargas, F. A. et al. Fc effector function contributes to the activity of human anti-CTLA-4 antibodies. Cancer Cell 33, 649–663.e644 (2018).
Ingram, J. R. et al. Anti–CTLA-4 therapy requires an Fc domain for efficacy. Proc. Natl Acad. Sci. USA 115, 3912–3917 (2018).
Lax, B. M. et al. Both intratumoral regulatory T cell depletion and CTLA-4 antagonism are required for maximum efficacy of anti-CTLA-4 antibodies. Proc. Natl Acad. Sci. USA 120, e2300895120 (2023).
Dahan, R. et al. FcγRs modulate the anti-tumor activity of antibodies targeting the PD-1/PD-L1 axis. Cancer Cell 28, 285–295 (2015).
Hamers-Casterman, C. et al. Naturally occurring antibodies devoid of light chains. Nature 363, 446–448 (1993).
Ingram, J. R., Schmidt, F. I. & Ploegh, H. L. Exploiting nanobodies’ singular traits. Annu. Rev. Immunol. 36, 695–715 (2018).
Dougan, M. et al. Targeting cytokine therapy to the pancreatic tumor microenvironment using PD-L1–specific VHHs. Cancer Immunol. Res. 6, 389–401 (2018).
Liu, X., Balligand, T., Carpenet, C. & Ploegh, H. L. An armed anti-immunoglobulin light chain nanobody protects mice against influenza A and B infections. Sci. Immunol. 8, eadg9459 (2023).
Arvedson, T. et al. Targeting solid tumors with bispecific T cell engager immune therapy. Annu. Rev. Cancer Biol. 6, 17–34 (2022).
Goebeler, M.-E. & Bargou, R. C. T cell-engaging therapies—BiTEs and beyond. Nat. Rev. Clin. Oncol. 17, 418–434 (2020).
Guimaraes, C. P. et al. Site-specific C-terminal and internal loop labeling of proteins using sortase-mediated reactions. Nat. Protoc. 8, 1787–1799 (2013).
Ingram, J. R. et al. PD-L1 is an activation-independent marker of brown adipocytes. Nat. Commun. 8, 647 (2017).
Liu, R. et al. Structure of human phagocyte NADPH oxidase in the resting state. eLife 11, e83743 (2022).
Dranoff, G. et al. Vaccination with irradiated tumor cells engineered to secrete murine granulocyte-macrophage colony-stimulating factor stimulates potent, specific, and long-lasting anti-tumor immunity. Proc. Natl Acad. Sci. USA 90, 3539–3543 (1993).
Van Elsas, A., Hurwitz, A. A. & Allison, J. P. Combination immunotherapy of B16 melanoma using anti–cytotoxic T lymphocyte–associated antigen 4 (CTLA-4) and granulocyte/macrophage colony-stimulating factor (GM-CSF)-producing vaccines induces rejection of subcutaneous and metastatic tumors accompanied by autoimmune depigmentation. J. Exp. Med. 190, 355–366 (1999).
Hodi, F. S. et al. Immunologic and clinical effects of antibody blockade of cytotoxic T lymphocyte-associated antigen 4 in previously vaccinated cancer patients. Proc. Natl Acad. Sci. USA 105, 3005–3010 (2008).
Dougan, M. et al. IAP inhibitors enhance co-stimulation to promote tumor immunity. J. Exp. Med. 207, 2195–2206 (2010).
Baumeister, S. H., Freeman, G. J., Dranoff, G. & Sharpe, A. H. Coinhibitory pathways in immunotherapy for cancer. Annu. Rev. Immunol. 34, 539–573 (2016).
Behrens, L. M., van Egmond, M. & van den Berg, T. K. Neutrophils as immune effector cells in antibody therapy in cancer. Immunol. Rev. 314, 280–301 (2023).
Krummel, M. F. & Allison, J. P. CD28 and CTLA-4 have opposing effects on the response of T cells to stimulation. J. Exp. Med. 182, 459–465 (1995).
Sato, Y. et al. Fc-independent functions of anti-CTLA-4 antibodies contribute to anti-tumor efficacy. Cancer Immunol. Immunother. 71, 2421–2431 (2022).
Cha, J.-H., Chan, L.-C., Li, C.-W., Hsu, J. L. & Hung, M.-C. Mechanisms controlling PD-L1 expression in cancer. Mol. Cell 76, 359–370 (2019).
Kurino, T. et al. Poor outcome with anti-programmed death-ligand 1 (PD-L1) antibody due to poor pharmacokinetic properties in PD-1/PD-L1 blockade-sensitive mouse models. J. Immunother. Cancer 8, e000400 (2020).
Lopus, M. et al. Maytansine and cellular metabolites of antibody-maytansinoid conjugates strongly suppress microtubule dynamics by binding to microtubules. Mol. Cancer Ther. 9, 2689–2699 (2010).
Verhaar, E. R. et al. MICA-specific nanobodies for diagnosis and immunotherapy of MICA+ tumors. Front. Immunol. 15, 1368586 (2024).
Zhang, H., You, Q.-D. & Xu, X.-L. Targeting stimulator of interferon genes (STING): a medicinal chemistry perspective. J. Med. Chem. 63, 3785–3816 (2019).
Lioux, T. et al. Design, synthesis, and biological evaluation of novel cyclic adenosine–inosine monophosphate (cAIMP) analogs that activate stimulator of interferon genes (STING). J. Med. Chem. 59, 10253–10267 (2016).
Wu, Y.-t et al. Tumor-targeted delivery of a STING agonist improves cancer immunotherapy. Proc. Natl Acad. Sci. USA 119, e2214278119 (2022).
Nicolai, C. J. et al. NK cells mediate clearance of CD8+ T cell–resistant tumors in response to STING agonists. Sci. Immunol. 5, eaaz2738 (2020).
Brosius, I. I. I. et al. Mouse models of diabetic nephropathy. J. Am. Soc. Nephrol. 20, 2503–2512 (2009).
Wei, Q. & Dong, Z. Mouse model of ischemic acute kidney injury: technical notes and tricks. Am. J. Physiol. Ren. Physiol. 303, F1487–F1494 (2012).
Cheloha, R. W., Harmand, T. J., Wijne, C., Schwartz, T. U. & Ploegh, H. L. Exploring cellular biochemistry with nanobodies. J. Biol. Chem. 295, 15307–15327 (2020).
Vincke, C. et al. General strategy to humanize a camelid single-domain antibody and identification of a universal humanized nanobody scaffold. J. Biol. Chem. 284, 3273–3284 (2009).
Rashidian, M. et al. Predicting the response to CTLA-4 blockade by longitudinal noninvasive monitoring of CD8 T cells. J. Exp. Med. 214, 2243–2255 (2017).
Bannas, P., Hambach, J. & Koch-Nolte, F. Nanobodies and nanobody-based human heavy chain antibodies as antitumor therapeutics. Front. Immunol. 8, 1603 (2017).
Hoefman, S., Ottevaere, I., Baumeister, J. & Sargentini-Maier, M. L. Pre-clinical intravenous serum pharmacokinetics of albumin binding and non-half-life extended Nanobodies®. Antibodies 4, 141–156 (2015).
Hanke, L. et al. A bispecific monomeric nanobody induces spike trimer dimers and neutralizes SARS-CoV-2 in vivo. Nat. Commun. 13, 155 (2022).
Steeland, S., Vandenbroucke, R. E. & Libert, C. Nanobodies as therapeutics: big opportunities for small antibodies. Drug Discov. Today 21, 1076–1113 (2016).
Bothwell, A. L. et al. Somatic variants of murine immunoglobulin λ light chains. Nature 298, 380–382 (1982).
Popov, A. V., Zou, X., Xian, J., Nicholson, I. C. & Brüggemann, M. A human immunoglobulin λ locus is similarly well expressed in mice and humans. J. Exp. Med. 189, 1611–1620 (1999).
Ereño-Orbea, J. et al. Structural basis of enhanced crystallizability induced by a molecular chaperone for antibody antigen-binding fragments. J. Mol. Biol. 430, 322–336 (2018).
Grosso, J. F. & Jure-Kunkel, M. N. CTLA-4 blockade in tumor models: an overview of preclinical and translational research. Cancer Immun. 13, 5 (2013).
Kocak, E. et al. Combination therapy with anti–CTL antigen-4 and anti-4-1BB antibodies enhances cancer immunity and reduces autoimmunity. Cancer Res. 66, 7276–7284 (2006).
Wei, S. C., Duffy, C. R. & Allison, J. P. Fundamental mechanisms of immune checkpoint blockade therapy. Cancer Discov. 8, 1069–1086 (2018).
Dane, E. L. et al. STING agonist delivery by tumour-penetrating PEG-lipid nanodiscs primes robust anticancer immunity. Nat. Mater. 21, 710–720 (2022).
Osorio, J. C., Smith, P., Knorr, D. A. & Ravetch, J. V. The antitumor activities of anti-CD47 antibodies require Fc-FcγR interactions. Cancer Cell 41, 2051–2065. e2056 (2023).
Juneja, V. R. et al. PD-L1 on tumor cells is sufficient for immune evasion in immunogenic tumors and inhibits CD8 T cell cytotoxicity. J. Exp. Med. 214, 895–904 (2017).
Xie, Y. J. et al. Nanobody-based CAR T cells that target the tumor microenvironment inhibit the growth of solid tumors in immunocompetent mice. Proc. Natl Acad. Sci. 116, 7624–7631 (2019).
Acknowledgements
We thank the Vascular Biology Program (VBP) Bioimaging Core at Boston Children’s Hospital for training and support in the use of a PerkinElmer IVIS in vivo imaging system. We also thank the Center for Macromolecular Interactions at Harvard Medical School for their support and assistance with the SPR experiments. This research was supported by NIH grant R01AI182177 to X.L. and H.L.P., and by the NIH Director’s Pioneer Award DP1AI150593 to H.L.P. T.B. was supported by a fellowship from the Belgian American Educational Foundation and by a WBI World fellowship from Wallonie-Bruxelles International. R.K.A. was supported by the Cancer Research Institute Irvington Postdoctoral Fellowship to Promote Racial Diversity (reference number CRI3968).
Author information
Authors and Affiliations
Contributions
X.L., R.K.A. and H.L.P. conceived the idea. X.L., C.L., R.K.A. and H.L.P. designed the studies. X.L. prepared the nanobody conjugates and their drug adducts. X.L., C.L. and E.B. characterized the nanobody conjugates in vitro. X.L., C.L., R.K.A., E.B. and T.B. tested the nanobody conjugations and their drug adducts in vivo. X.L., C.L., R.K.A., E.B., T.B. and H.L.P. interpreted the data. X.L., C.L., R.K.A. and H.L.P. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
X.L. and H.L.P. have filed a patent covering the technologies described in this publication (international publication number WO2023141500A2). The other authors declare no competing interests.
Peer review
Peer review information
Nature Biomedical Engineering thanks Christian Klein, Michael Mitchell, John Wilson and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Individual tumor growth curves for the experiments presented in main Fig. 3.
a, Individual tumor growth curves for MC38 tumor-bearing mice (n = 6) treated intraperitoneally with the indicated VHHs at 5 mg/kg on day 1 post-tumor injection, followed by three times a week for 3 weeks as indicated by the arrows. b, Individual tumor growth curves for B16-F10 tumor-bearing mice (n = 6) treated intraperitoneally with the indicated VHHs at 5 mg/kg on day 1 post-tumor injection, followed by three times a week for 3 weeks as indicated by the arrows. The mice were vaccinated with GM-CSF–secreting B16 cells (GVAX) on day 0. c, Individual tumor growth curves for MC38 tumor-bearing mice (n = 6) treated intravenously with the indicated VHHs at 5 mg/kg or antibody at 12.5 mg/kg on day 7, 10, and 13 post-tumor injection as indicated by the arrows. d, Tumor growth (left) and survival curves (middle) for MC38 tumor-bearing mice (Data represent mean ± SD, n = 6) treated intravenously with the anti-CTLA-4 VHH (H11)-VHHkappa conjugate at 5 mg/kg and 2.5 mg/kg on day 7, 10, and 13 post-tumor injection as indicated by the arrows. † Mouse was sacrificed due to severe ulceration of its tumor. This is part of the study presented in Fig. 3f. Right: Individual tumor growth curves for mice treated with 2.5 mg/kg of the H11-VHHkappa conjugate. (Two-way ANOVA with Bonferroni’s correction for multiple comparisons was used to analyze the tumor growth curves. The two-sided Log-rank (Mantel-Cox) test was applied to analyze the survival curves.).
Extended Data Fig. 2 The anti-CTLA-4 VHH (H11)-VHHkappa conjugate and anti-CTLA-4 monoclonal 9H10 re-polarize the tumor microenvironment.
The H11-VHHkappa conjugate and 9H10 substantially alter the TME by depleting regulatory T cells. In addition, H11-VHHkappa induces an increase of tumor-infiltrating neutrophils, and an (non-significant, P = 0.06) reduction in tumor cell proliferation. Tumor cells were defined as live CD45− while tumor-infiltrating leukocytes were gated on live CD45+, myeloid cells were defined as live CD45+CD11b+. The myeloid compartment was further analyzed for expression of CD11c, MHCII, CD86 and Ly6G. Neutrophils were gated on live CD45+CD11b+Ly6Ghi. NK cells were gated on live CD45+NK1.1+. T cells were gated on live CD45+CD3+ and further analyzed for expression of CD8, CD4 CD25 and CD69. Activated CD4+ and CD8+ T cells were selected based on CD69 expression. Regulatory T cells were gated on live CD45+CD4+CD25+FoxP3+. Proliferation was measured by staining for Ki67. Results were analyzed by one way ANOVA with Bonferoni’s correction for multiple testing (Data represent mean ± SD, n = 3). Statistical significance was plotted as follows: * P < 0.05, ** P < 0.01 *** P < 0.001 and **** P < 0.0001.
Extended Data Fig. 3 Individual tumor growth curves for the experiment presented in main Fig. 5.
a, Individual tumor growth curves for MC38 tumor-bearing mice (n = 6) treated intraperitoneally with the indicated VHHs at 5 mg/kg on day 1 post-tumor injection, followed by three times a week for 3 weeks as indicated by the arrows. b, Individual tumor growth curves for B16-F10 tumor-bearing mice (n = 6) treated intraperitoneally with the indicated VHHs at 5 mg/kg on day 1 post-tumor injection, followed by three times a week for 3 weeks as indicated by the arrows. The mice were vaccinated with GM-CSF–secreting B16 cells (GVAX) on day 0. c, Individual tumor growth curves for wild-type and PD-L1 knockout MC38 tumor-bearing mice (n = 5) treated intraperitoneally with the indicated VHHs at 5 mg/kg on day 1 post-tumor injection, followed by three times a week for 3 weeks as indicated by the arrows. d, Individual tumor growth curves for MC38 tumor-bearing wild-type and LysMcrePD-L1fl/fl (with PD-L1 deficient myeloid cells) mice (n = 5) treated intraperitoneally with the indicated VHHs at 5 mg/kg on day 1 post-tumor injection, followed by three times a week for 3 weeks as indicated by the arrows.
Extended Data Fig. 4 Individual tumor growth curves and body weight curves for the experiments presented in main Fig. 6.
a, Individual tumor growth curves for MC38 tumor-bearing mice (n = 6) treated intravenously with A12-VHHkappa-DM1 or A12-VHHkappa-DM4 at 5 mg/kg on day 7, 10,13, and 17 post-tumor injection as indicated by the arrows. b, Individual tumor growth curves for MC38 tumor-bearing mice (n = 6) treated intravenously with indicated VHH-DM4 conjugates at 5 mg/kg on day 7, 10,13, and 17 post-tumor injection as indicated by the arrows. c, Individual tumor growth curves (upper panels) and body weight curves (lower panel, data represent mean ± SD) for B16-F10 tumor-bearing mice (n = 5) treated intravenously with indicated VHH-DM4 conjugates at 5 mg/kg on day 3, 6, 9, and 13 post-tumor injection as indicated by the arrows.
Extended Data Fig. 5 Conjugation of DM4 or a STING agonist to the anti-PD-L1 VHH (A12)-VHHkappa conjugate re-polarizes the tumor microenvironment.
A12-VHHkappa-DM4 treatment led to pronounced cell death in the tumor, and halted tumor cell proliferation. Inclusion of DM4 reduced the fraction of tumor-infiltrating cells (mostly of myeloid origin), particularly CD11c+MHCII+ cells. A12-VHHkappa-DM4 also reduced CD86 expression on CD11c+ cells, while an increase in neutrophil infiltration was detected. In the T cell compartment, CD4+ and CD8+ T cell activation was increased by A12-VHHkappa-DM4 treatment. A12-VHHkappa-STING agonist surprisingly increased the proliferation of tumor cells, and, as expected, increased the activation of CD4+ and CD8+ T cells in the tumor. Tumor cells were defined as live CD45− while tumor-infiltrating leukocytes were gated on live CD45+, myeloid cells were defined as live CD45+CD11b+. The myeloid compartment was further analyzed for expression of CD11c, MHCII, CD86 and Ly6G. Neutrophils were gated on live CD45+CD11b+Ly6Ghi. NK cells were gated on live CD45+NK1.1+. T cells were gated on live CD45+CD3+ and further analyzed for expression of CD8, CD4 CD25 and CD69. Activated CD4+ and CD8+ T cells were selected based on CD69 expression. Regulatory T cells were gated on live CD45+CD4+CD25+FoxP3+. Proliferation was measured by staining for Ki67. Results were analyzed by one way ANOVA with Bonferoni’s correction for multiple testing (Data represent mean ± SD, n = 3 for PBS-, A12-VHHkappa-DM4-, and A12-VHHkappa-STING agonist–treated groups, and n = 4 for the A12-VHHkappa conjugate–treated group). Statistical significance was plotted as follows: * P < 0.05, ** P < 0.01 *** P < 0.001 and **** P < 0.0001.
Extended Data Fig. 6 Individual tumor growth curves and body weight curves for the experiments presented in main Fig. 7.
a, Individual tumor growth curves for MC38 tumor-bearing mice (n = 6) treated intravenously with A12-VHHkappa-STING agonist or A12-VHHkappa-conjugate plus free STING agonist (equimolar amount) at 5 mg/kg on day 7, 10, and 14 post-tumor injection as indicated by the arrows. b, Individual tumor growth curves (upper panels) and body weight curves (lower panel, data represent mean ± SD) for B16-F10 tumor-bearing mice (n = 5) treated intravenously with A12-VHHkappa-STING agonist or A12-VHHkappa-conjugate plus free STING agonist (equimolar amount) on day 3, 7,11, and 15 post-tumor injection as indicated by the arrows. c, Individual tumor growth curves (upper panels) and body weight curves (lower panel, data represent mean ± SD) for MC38 tumor-bearing mice (n = 6) treated intravenously with A12-VHHkappa-STING agonist or A12-VHHkappa-conjugate plus free STING agonist (equimolar amount) at 10 mg/kg on day 7, 10, and 14 post-tumor injection as indicated by the arrows.
Extended Data Fig. 7 Evaluation of toxicity of VHHkappa conjugates and their drug adducts in mice.
a, Assessment of liver and renal toxicity of VHHkappa conjugates and their drug adducts by measuring alanine aminotransferase (ALT), aspartate aminotransferase (AST), creatinine, urea, and blood urea nitrogen (BUN) levels. BUN concentrations were calculated from urea concentrations using the formula: BUN (mg/dL) = [Urea] (mg/dL) / 2.14. Mice were intravenously treated with 5 mg/kg of VHHkappa conjugates or their drug adducts following the same dosing schedule as their therapeutic treatments: every three days for three doses (H11-VHHkappa conjugate and A12-VHHkappa-STING agonist) or four doses (A12-VHHkappa conjugate and A12-VHHkappa-DM4). Blood samples were collected three days after the final treatment (Data represent mean ± SD, n = 4, two-sided unpaired t test). b, Measurement of mouse interferon beta (IFN-β) levels in tumor homogenates and blood following treatment with A12-VHHkappa-STING agonist. Tumor-bearing mice were intravenously treated with A12-VHHkappa-STING agonist or A12-VHHkappa conjugate plus an equimolar amount of free STING agonist at 5 mg/kg on days 11 and 13. Tumors and blood samples were collected on day 14 for IFN-β measurement. IFN-β levels below the assay’s detection limit were recorded as 0 (Data represent mean ± SD, n = 3 for untreated and A12-VHHkappa-STING agonist–treated groups, and n = 4 for the A12-VHHkappa conjugate + STING agonist–treated group, two-sided unpaired t test).
Supplementary information
Supplementary Information
Supplementary Figs. 1–23 and Chemical synthesis.
Supplementary Data 1
Supplementary data for Figs. 3, 5–8, 10, 15 and 20.
Source data
Source Data Figs. 1–7 Source Data Extended Data Figs. 1–7
A single file containing all source data, with clearly named tabs for each Figure/Extended Data Figure item.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Liu, X., Le Gall, C., Alexander, R.K. et al. Nanobody-based bispecific antibody engagers targeting CTLA-4 or PD-L1 for cancer immunotherapy. Nat. Biomed. Eng 10, 39–55 (2026). https://doi.org/10.1038/s41551-025-01447-z
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41551-025-01447-z
This article is cited by
-
Benchmarking matters
Nature Biomedical Engineering (2025)
