
Abstract
Narratives about the motivations and conditions for mass violence as a persistent feature of conflict throughout human history have evolved in complexity and materiality. Victims of these events are key for understanding the evolution and transformative power of violent behaviour as it developed from simple intergroup conflict to more strategic mass violence. Here we present the results of a bioarchaeological study of 77 and biomolecular analysis of 25 individuals from a ninth-century BCE mass grave from Gomolava in the Carpathian Basin, Southeast Europe. The site is located at the interface of complex sociospatial relations, divergent cultural traditions and values, and competing ideologies of landscape use. We show that excessive lethal violence enacted mostly on women and children suggests a selective demographic bias. The people buried together shared few, even distant, genetic relationships, and so their killing presents striking evidence for an episode of cross-regional conflict and an underlying aggressive shift in power, violence and gender relations in the region. Gomolava provides evidence consistent with deliberate annihilation of select sections of a regional population as a motivation for mass violence behaviour in later prehistoric Europe. It also shines new light on the socioeconomic agency and importance of women and young individuals in later European prehistory.
Similar content being viewed by others
Main
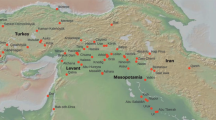
Violence is an integral facet of human behaviour that impacts on how societies function from individual rights to group values and ideologies. This has been shaping inquiry in the behavioural and social sciences for some time: why we fight, how we fight, who is targeted and how we are affected—whether killed, maimed, deprived of freedom, psychologically traumatized or otherwise impacted. Identifying reasons for violent interactions is complex and relies on examining cultural and material conditions specific to societies1. These range across environmental and economic constraints, resolving perceived offence or power aggrandisement as key motivations and consequences for inter- and intragroup violence2. The character and organization of mass graves has been recognized only more recently as a crucial source for understanding the core purposes of collective violence by humans3. Gomolava, located in Northern Serbia in the south Carpathian Basin, is one of the largest prehistoric single-event mass graves excavated so far in Europe (Fig. 1), and it is key to better understanding large-scale killing in later prehistory. Detailed study of this site provides new insights into how instrumental violence and social order intersect. It also extends our knowledge of the geographic and sociopolitical scope of conflicts in prehistory and how they were embedded within cultural and social processes4.

a, Map showing the location of Gomolava and the early first-millennium BC archaeological ceramic typo-chronology groups in the Southwest Carpathian Basin. b, Reconstruction of the burial event at Gomolava by S.N.
Our data reveal that the killing and burial of 77 individuals extended beyond a family or even a coresidential group of individuals, and the victims of violence were predominantly biologically unrelated women and children from a larger, regional population. Without having earlier precedents in known mass graves, this demographic bias reveals growing diversification of human violence and the roles and values of women and young individuals in communities within the region. Using an interdisciplinary biocultural approach, we explore the character and context of this mass killing event and the creation of an emotive place to bury the victims together 2,800 years ago.
Early Iron Age Gomolava was a place of memory. A tell site founded in the sixth millennium BCE5, it was repeatedly used across hundreds of generations as a marker in the social and physical landscape (Supplementary Note 1). It lies in the south Pannonian Plain, at the core of the Carpathian Basin. In the early first millennium BCE, it stood at the interface between regional cultural traditions (Fig. 1). To the north and west, the late Urnfield material culture, mortuary and settlement traditions remained dominant and included long-established, large-scale economic–political networks. Kalakača style pottery was used throughout the south Pannonian Plain, and the Gomolava settlement lay at the westernmost limit of its common use. They also used local styles of metalwork together with a new Thraco-Cimmerian style that originated in the western Eurasian Steppe. The South Pannonian Plain had been dominated by a network of massive enclosed sites in the later Bronze Age whose abandonment in the thirteenth to twelfth centuries BCE suggests sociopolitical collapse followed by increasingly mobile lifeways within, and limited migration outwards from, the plain6. Located beside the River Sava, Gomolava was one of a few tells that served as possible ancient place markers on river routes demarcating boundaries or places of ancestral importance in the social landscape7. A horizon of reoccupation of such older central sites identifiable through material culture and in 14C data and the establishment of new, smaller, enclosed sites in the ninth century BCE was closely contemporary to the burial at Gomolava8 (Fig. 1 and Supplementary Note 4). Material culture traditions also reveal increased mobility within trans-Carpathian networks to the east, including the arrival of mobile-pastoralist Mezőcsat groups to central parts of the plain, potentially from the Eurasian Steppe9,10. The killing event took place at an unsettled time when semi-sedentary communities were reconsolidating around tells and abandoned mega-forts as ancestral spaces and creating new enclosures10. Tensions between established and new groups that were following different lifeways may have given rise to conflict over land use and ownership; Gomolava was situated at a physical, political and conceptual flashpoint (Fig. 1).
The evolution of violence in prehistoric Europe
Large-scale violence can leave clear traces in the bioarchaeological record, including mass graves—a phenomenon that may date back approximately 10,000 years to hunter-gatherer societies11. Early collective violence involved relatively indiscriminate killing of small numbers of men, women and children and was probably the result of sporadic or localized conflicts such as raids. Farming and sedentism were introduced to Europe in the seventh millennium BCE, after which evidence for mass violence increases in scale and frequency12. During the Neolithic, conventions for who was actively involved in and most frequently affected by conflict narrowed and became more selective. Males, most likely as combatants, engaged in regional as well as local scales of war-like conflict. This was a by-product of increasing territoriality, population density and division of labour. It enabled the consolidation of larger-scale, increasingly centralized and institutionalized systems of violence and warfare13. From the mid-fourth millennium BCE, management of legitimate violence became permanently embedded in societies through institutionalization including more specialized warrior roles14,15. In the second millennium BCE, the advent of swords, shields, spears and battle axes made from bronze and the introduction of the horse to battlefields reshaped combat practices, although organic weapons, including clubs, remained in use. With this, the number of individuals potentially involved, the complexity of martial practices, potential for strategic instrumental violence and the geographic scope of violent conflict all expanded, resulting in battles that included several thousand combatants14,16. Armies may have coalesced for specific purposes in late second/early first millennium BCE in Europe, and it is likely that the killing at Gomolava occurred in such a framework before the subsequent emergence of standing armies. The nature of violence attested at Gomolava brings a new dimension to our understanding of violence in later prehistory by elucidating the complexity of gender and age factors underlying the purposes and impacts of violence17.
Wealth and conspicuous consumption
As preserved, the Gomolava grave measured a mere 2.9 m in diameter and 0.5 m in depth. It was a deliberately created, curated and protected space in a location that had held importance in the region for thousands of years7. Postholes around the perimeter of the burial pit suggest the reuse of an earlier feature or a modest memorialization of the burial, either of which emphasized the creation of a meaningful space that remained identifiable thereafter (Supplementary Note 1). Together with the lack of animal scavenging and the organized placement of articulated remains (Fig. 2 and Supplementary Note 2), this suggests considered intent. Small personal ornaments made of bronze were interred together with ceramic vessels suited to storing and consuming food and drink. Bones from between 50 and 100 animals were also deposited, including meat-bearing parts of cattle of various ages, adult pigs and a smaller number of sheep or goats. The articulated remains of most of a young cow were placed at the bottom of the grave pit below the articulated remains of the 77 humans. Symbolism surrounding food and structured deposition is further attested by broken quern stones placed on top of the grave and concentrations of burnt seeds, particularly in its centre7. Apart from the high number of individuals, this investment of resources and time at Gomolava contrasts with other known Early Iron Age mass burials, marking it as an exceptional and rich burial site, emphasizing the performative aspect of the burial event and commemoration18 (Figs. 1 and 2).

a, Burial plan of the human remains and finds from Gomolava mass burial 2 (by S.N. after Tasic 1972). b, Photograph of the Gomolava mass burial (reproduced courtesy of the Museum of Vojvodina). Note that no scale was included in the original photograph.
Gendered lifeways and deathways
Turning to the victims, the remains of all 77 individuals, curated at the Museum of Vojvodina, Novi Sad, were analysed bioarchaeologically (Supplementary Note 5 and Supplementary Table 5). Of these, 40 (51.9%) were juveniles (1–12 years), 11 (15.6%) were adolescents (13–17 years) and 24 (31.2%) were adults (18+ years), with only one (1.3%) articulated infant (<1 year) present (Table 1). Of the 72 individuals that could be biologically sexed either morphologically or through DNA or enamel peptide analysis (Methods; Supplementary Note 3 and Supplementary Tables 5 and 12), 51 (70.8%) were female, with males underrepresented in all age groups. Of the 24 sexed adults, 21 (87%) were female, as were 7 (70%) of the 10 sexed adolescents and 23 (62.2%) of the 37 sexed juveniles. The only articulated infant was a male. The absence or rarity of younger women at other mass burial sites has been noted with reference to their reproductive value, making their presence at Gomolava noteworthy19.
The predominance of women and younger individuals in the mass grave at Gomolava is exceptional in European prehistory18,20. The closest parallel is the fifth- to fourth-century BCE massacre at Fin Cop hillfort in England where victims recovered from a ditch were exclusively women and children21. This pattern at Gomolava is further supported by the presence of a second mass grave (mass grave 1) (Supplementary Notes 1 and 4). In this, 36 individuals were identified in 1954 in a pit truncated by river erosion, but there may originally have been up to 54 individuals, the majority of which were sexed female at the time7. Following on-site analysis, the bones were not retained. The orientation and placement of individuals within was less ordered, with the uppermost possibly simply thrown in. The date of the ceramics and shared features suggest the two graves were contemporary, potentially from the same event, but this cannot be tested. The burial pit for mass grave 1, demographics, inclusion of pottery, metalwork, grain and meat (including articulated animals) were strikingly similar to mass grave 2, the focus of this study.
A previous bioarchaeological analysis of the remains from mass grave 2 reiterated initial conclusions that had attributed the deaths to a pandemic22. However, our screening for pathogen DNA revealed no evidence of infectious agents22,23 (Supplementary Note 13). The analysis of single-nucleotide polymorphisms (SNPs) associated with health-related phenotypic traits identified alleles linked to lactase non-persistence, celiac disease susceptibility and malaria resistance in many individuals, reflecting patterns characteristic of early first-millennium BC populations of Central and Eastern Europe (Supplementary Note 12). The macroscopic analysis of the remains revealed the presence of skeletal changes indicative of non-specific systemic physiological stress (Supplementary Note 5 and Supplementary Table 6), but these did not result in increased frailty and reflected adaptation to stress. The most striking finding from our restudy substantially changes the interpretation of this site; there was extensive evidence for intentional, violent and frequently lethal trauma, mostly to the head. Peri-mortem (unhealed) injuries were found in 18.2% of individuals from mass grave 2 (Fig. 3, Supplementary Note 5 and Supplementary Table 7). This affected 22.5% of juveniles (aged between 5 and 12 years) and 13.8% of adolescents and adults, with all adult age groups and both sexes represented (Supplementary Note 5). In three individuals (3.9%), ante-mortem (healed) cranial trauma consistent with a potentially violence-related origin was noted, affecting 5% of juveniles and 2.8% of adolescents and adults. For comparison, in a recent population study of nearly 5,000 individuals from various periods from across Europe, only 5.5% showed evidence for cranial trauma as an indicator of violence24. Cranial injuries at Gomolava, both peri- and ante-mortem, involved mostly close contact and especially blunt force, which could have resulted from a number of implements or weapons with a rounded or flat profile as well as blunt force projectiles, such as sling-shot25. We also recorded a peri-mortem sharp force defence injury in one of the individuals (Sk42), suggesting a degree of resistance. Also present were a number of conventional peri-mortem projectile injuries (arrows and/or spears) (Supplementary Table 7), which suggest scenarios of victims trying to escape their killers. At least 20% of individuals showed signs of violence-related skeletal injuries.

a–c, Distribution and examples of cranial injuries recorded at Gomolava (a), including Sk26 in posterior-lateral view (b) and Sk33 in right-lateral view (c); both individuals had sustained extensive peri-mortem injuries. In both cases, the primary impact site is clearly recognizable as a void with missing bone, with secondary fractures radiating from the impact site (drawings by S.N.).
The true prevalence of trauma was probably much higher when we consider differential preservation due to a combination of previous glueing of cranial remains and the fact that not all attacks would leave marks on the skeleton26 especially those using sharp force that primarily targets soft tissue. Peri-mortem injuries dominate and are primarily located on the head but also affect the shoulder, arm and pelvis. The back, top and right side of the head are preferred targets, a pattern providing important insights into the nature of the confrontation resulting in the observed injuries. In most cases, the attacker and victim were not engaged in a face-to-face confrontation (as left lateral or anterior injuries are rare), nor were they necessarily evenly matched in strength or height when the blows were delivered27. Attackers were potentially taller than their victims or on horseback, targeting the superior aspect of the head (particularly notable in younger individuals; Supplementary Table 7), or the victims were in a compromised position, such as on the ground or fleeing, resulting in injuries to the superior and posterior aspects. Overall, the patterning reveals severe violence that was brutal, deliberate and efficient. Histotaphonomic analyses of computed tomography (CT) scans of eight bones from all levels of mass grave 2 demonstrated high levels of bioerosion (Supplementary Note 2). This is consistent with burial soon after death, suggesting that the killings occurred near Gomolava and that conditions allowed bodies to be collected immediately afterwards.
Relationships and social networks
To shed new light on the events that unfolded at Gomolava 2,800 years ago and on the victims, we generated low-coverage genomes (0.1–3.8× Depth of Coverage) for 25 of the 77 individuals in the grave. Damage patterns were consistent with ancient DNA and contamination levels were below 2% (Supplementary Tables 12–14). Genetic relatedness analysis revealed an almost complete lack of close (that is, first, second or third degree) relationships among the Gomolava individuals, with the exception of three more closely related individuals (Fig. 4a), with an adult female (Sk14) and her two daughters (Sk8 aged 7–8 years and Sk33 aged 7–9 years), who were confirmed as full siblings (Supplementary Note 8). The lack of close-kin relationships is further supported by a pairwise identity-by-descent (IBD) analysis, which revealed that merely 4% of individual pairs shared more than two IBD segments longer than 12 cM (Supplementary Table 20a and Supplementary Fig. 16). This indicates that most individuals in the assemblage were unrelated and did not even qualify as sixth-degree relatives28.

a, IBSrelate results. The lines show theoretically derived ranges of the joint expectation for each relationship. The circles show values for each pair of the analysed individuals. Inset: a single pedigree of a mother and her two daughters identified among the assemblage. b, Distribution of the sum of pairwise IBD sharing >12 cM for Gomolava and three previously published prehistoric mass graves (Koszyce, Potočani and Asparn-Schletz)29,30,31. Box plots show the median (centre line), interquartile range (box) and values within 1.5× the interquartile range (whiskers); points indicate individual data values, with outliers plotted beyond the whiskers. c, IBD sharing profile for non-closely related individuals in Gomolava. The plot shows a very close fit between the observed IBD sharing distribution (red bars) and the expected IBD sharing profile under a constant effective population size (dashed black line).
Given the general lack of biological relationships in the assemblage, it seems likely that we are dealing with a more or less random sample of a larger population, rather than an extended family or residents of a single, small settlement typical of the region at the time (Supplementary Note 8). To explore this further, we conducted a comparative analysis of IBD sharing patterns with the three other previously published mass graves from the wider region: the 7,000-year old mass burial at Asparn-Schletz29 in Austria, the approximately 6,200-year-old case of Potočani, Croatia30, and the approximately 5,000-year-old mass grave at Koszyce in southern Poland31. Similar to Gomolava, Asparn-Schletz and Potočani showed an absence of close genetic relatedness among those interred, with even distant relationships being rare, as reflected in reduced sharing of IBD segments >12 cM after excluding first-degree relatives. This pattern contrasts with the extended family structure observed at Koszyce, which consisted of mostly close and some more distant relatives (Fig. 4b, Supplementary Note 8 and Supplementary Table 20b). The abundance of adult females and juveniles at Gomolava, combined with the fact the individuals interred were largely unrelated, distinguishes Gomolava as a possibly select group based on age and gender.
Notions of kinship are diverse globally and culturally constituted, making it impossible to evaluate if people buried at Gomolava felt related or shared close bonds, but we can infer certain connections based on the manner of the burial31. In contrast to Koszyce, where related individuals were buried in close proximity, the three first-degree relatives at Gomolava were not interred together. We cannot tell if the people who buried them knew the deceased and buried them with others according to social or emotional, rather than biological, ties or whether they were unknown to them and therefore separated incidentally.
With limited information on Early Iron Age settlement numbers and size of sociopolitical units, demographic estimates based on archaeology are problematic. However, the size and distribution of settlements suggest that each was home—at least seasonally—to a population of tens to low hundreds, with numerous such units dispersed across the landscape forming translocal communities32. To gain insights into the size of the population from which the victims originated, we estimated the effective population size (Ne) based on patterns of IBD (Supplementary Note 11). The close alignment between inferred and expected number of shared IBD segments suggests that the effective size of the population remained stable over the timescale represented by IBD segment lengths, with an estimated range of 10,000–14,000 individuals. The smooth decay in the number of IBD segments with increasing segment length (Fig. 4c) is what we would expect under a constant-size model, without any recent population size changes. Our estimates suggest a sizeable meta-population in the low tens of thousands occupying and interacting within the South Pannonian Plain during this period (Supplementary Note 11). This is also consistent with the scarcity of shorter runs of homozygosity, which suggests a relatively large mating pool with a substantial effective population size (Supplementary Table 21).
Origins, connections and migration
Mass grave 2 contained Kalakača style ceramics as well as metalwork common to Southeast Europe (spectacle-brooches) and others of Thraco-Cimmerian style (hair locks and phalera) from the western Eurasian Steppe (Supplementary Note 7), indicating possible eastward connections33. Typologically, the objects date to the tenth to early eighth centuries BCE, which fits well with the mid-to-late ninth-century BCE radiocarbon dates we obtained for some of the human remains in the grave (Supplementary Note 4). Lead isotope and elemental data from 11 objects (Supplementary Note 7) indicated a relatively homogeneous composition, not biased by stylistic tradition, with metal sourced predominantly from the southern Alps but mixed with other sources—through recycling, for example—to create a local metal ‘pool’34. These results indicate that the ornaments were probably manufactured locally in the South Pannonian Plain, albeit with cultural/stylistic influences from the east.
To explore the origins and connections of those buried at Gomolava further, we carried out strontium isotope analyses, which, together with ancient DNA data, help shed light on their backgrounds. The strontium isotope ratios fall within a relatively wide range between 0.70814 and 0.71214, suggesting disparate origins. Eight individuals fall outside the local range for the immediate environs of Gomolava, as defined by strontium isotope measurements of plants from around the site (Supplementary Note 6). Seven of these fall within the baseline range for bioavailable strontium encompassing the surrounding region of the Pannonian Plain and hinterlands to the south, which also includes the Gomolava range (Fig. 5a and Supplementary Note 6). One individual, Sk53, stands out with an enamel strontium isotope ratio entirely outside the known range for the Pannonian Plain, indicating more distant natal origins35,36. These data suggest that, although many of the interred females grew up in the broad region around Gomolava, 35% of sampled individuals originated from tens of kilometres—or more—away from their place of burial.
The carbon and nitrogen stable isotope results show a similarly wide spread in values (Fig. 5b). This points to diverse foodways and subsistence strategies and supports the idea that we are dealing with a relatively heterogenous group that exploited isotopically diverse facets of their environment. This is also supported by the wide range of δ13C values that were observed in cattle and caprines, indicating that livestock grazed on diverse woodland and open pastures. This implies a wide range of pasturing practices and widespread distribution of these animals across the landscape, with different management regimes. Such patterns may indicate that, like humans, animals came from different settlement locations, whether by trade, rustling or the same social processes that concentrated the humans from different settlements together. Overall, the results indicate that we are dealing with a relatively heterogeneous group drawn from multiple settlements or from a network of one or more translocal communities.

a, Enamel strontium and carbon isotope results from Gomolava. Individual sex is represented by shape, with circles indicating females and triangles representing males. The ‘local’ strontium isotope baseline is indicated by the dotted lines and is based on strontium isotope measurements of the local plants, depicted on the right-hand side of the plots, b, Carbon and nitrogen isotope results from Gomolava. Different animal taxa are represented by colour: blue for sus, green for bos, magenta for ovis/capra and orange for cervus. Data can be found in Supplementary Tables 8 and 9.
To better understand the spatial and temporal relationships in the region of the people from mass grave 2, we explored their genetic ancestry using multidimensional scaling (MDS) and ancestry modelling37. As shown in Fig. 6a, all individuals form a relatively homogeneous cluster within the general genomic diversity of Bronze Age and Early Iron Age (approximately 2,800 to 800 BCE) individuals from Serbia, Hungary, Albania and Croatia, indicating that they derived from a similar ancestral pool to other Early Iron Age groups across the Balkans38. To formally test these relationships, we performed a qpWave37 analysis, which demonstrated that all individuals can be modelled as deriving from a single ancestral source, consistent with the visual clustering observed in the MDS37 (Supplementary Note 10 and Supplementary Table 22). Additional ancestry modelling using qpAdm37 indicates that their genetic makeup is best described as a mixture of three basal ancestries: ‘Serbia Iron Gates Mesolithic’, ‘Anatolia Neolithic’ and ‘Samara Yamnaya Early Bronze Age’ (Supplementary Note 10 and Supplementary Table 23). When compared with populations from relevant historical and geographic contexts, the Gomolava cluster most closely aligns with populations from Hungary during the Bronze Age (Fig. 6b).

a, MDS analysis based on genomic data showing the Gomolava individuals against a backdrop of other ancient individuals from West Eurasia. b, Ancestry modelling results using qpADM. The individually modelled results for the Gomolava individuals are shown in Supplementary Fig. 20.
Given the dearth of male individuals in the grave, it is difficult to draw any definite conclusions about the form of social organization at Gomolava. Other studies suggest that patrilocal practices were a common social norm in other parts of the Carpathian Basin during the Neolithic and Bronze Age39. The fact that many of the females fall outside the local strontium isotope range is consistent with exogamic practices (Fig. 5 and Supplementary Note 9). However, many of those individuals are juveniles or young adolescents, and without knowledge of marital age norms, it is difficult to establish whether this pattern reflects exogamic practices or simply a diverse group drawn from multiple residential communities or a combination of both. Bearing in mind that we sampled only about a third of the individuals in the grave, the general lack of shared IBD segments perhaps lends support to the latter scenario.
Discussion
Fragmented identities, collective memories
Fragmented identities, collective memories. Its mid-ninth century BCE date places the Gomolava mass graves in a period of heightened archaeologically visible activity and rejuvenation in the Pannonian Plain, marked by population influx and movement across an evolving and contested landscape. Amid tensions between practices stemming from sedentary and mobile lifeways, different communities established a potentially explosive set of conflicting ideologies of land use and ownership. Directly equating material culture with sociopolitical units is a well-recognized problem in archaeology, but sufficiently different combinations of objects, domestic and mortuary conventions exist to testify to the presence of meaningfully different groups interacting within the wider region. Competing attempts to introduce or enforce distinct lifeways and claim territory at a regional level (Fig. 1) may have triggered displacement of people.
Kinship and community may both be relational, not defined by biology40. For the people buried in mass grave 2, the evident diversity both in their places of childhood origin and in their diets suggests that relationships between the deceased were not based on long-term coresidency. Consequently, how and why the deceased were initially displaced and then gathered at this site has social implications. Some may have joined the translocal community or communities around Gomolava before the conflict through marriage (exogamy) or fostering. Others, whether pushed by the threat of attack, drawn to a place of perceived greater safety or even brought involuntarily as captives, may have experienced forced migration or displacement before the killing event near Gomolava41. We know it took place nearby because the histotaphonomy data show they were buried soon after death. The killing itself may have been coercive or punitive to impose hegemony or subdue resistance, making it an intentional, forward-looking act. Such systematic, instrumental violence has some precedent in Europe30, but the gendered bias and upscaling of violence recorded at Gomolava are remarkable for the period and region.
Lacking data for degrees of relatedness of typical coresidential communities in the Early Iron Age, the rarity of relatedness evident in mass grave 2 must be considered against the population estimate of 100–200 individuals for large-sized settlements of this period (Supplementary Note 1). The burial of so many unrelated children and adults indicates the improbability that those buried came from a single settlement. The strontium isotope data show that most of them probably came from local and more distant parts of a translocal network that encompassed hundreds of square kilometres in the south Pannonian Plain, but at least 2 of the 25 individuals (8%) came from farther afield. In light of this, we argue that Gomolava mass grave 2 was neither aimed simply at the destruction of a single local co-residential group, as at Koszyce around 2,800 BCE31, nor intended to reduce fighting capacity to subjugate such a group12. Instead, it was an expression of a conflict involving displacement of people and targeted killing in the southwest Pannonian Plain. The deliberate selection of such a large number of women and young individuals can probably not be reduced to an incidental selection of ‘soft’ or emotive targets. Contemporaneous as well as anthropological comparisons demonstrate the social, economic and martial influence of women42 and the position of women and younger individuals as primary rather than secondary social and economic actors43,44,45. The selection, gathering, killing and burial of this demographic—valuable as the future of communities to friends or as enslaved labour to foes—represents structured deposition. It is plausible that the underlying intention or symbolism referenced the act of hoarding valuable objects and depositing them together, often close to rivers, a common practice across prehistoric Europe46. Whether echoing or intentionally referencing this practice, the Gomolava collective burials went beyond utility and present multireferential, culturally situated, symbolic purposes46.
Precedents for a major killing event extending beyond nuclear families comes from the Neolithic sites of Asparn-Schletz and Potočani29,47. Like at Gomolava, the individuals buried in those mass graves were largely unrelated and interpreted as being drawn from one or more translocal communities. However, given the roughly equal numbers of males and females in the graves, Asparn-Schletz and Potočani lack evidence for selectivity in the acts of killing and are more akin to an indiscriminate massacre47. By contrast, at Gomolava, the victims were also preferentially selected for gender and age. The level of killing was probably also upscaled, as indicated by a larger number of dead and the presence of another closely—or precisely—contemporary mass grave (potentially bringing the total number of contemporary victims to over 130). A mixture of close contact and distance weaponry injuries, as well as the presence of trauma sustained in defence may indicate multiple violent stages or closely related violent events. Those coordinating the episode of violence behind mass grave 2 did not simply indiscriminately decimate numbers; their targeted acts entailed widespread genealogical disruption through demographic selection. The inclusion of a very small number of adult and adolescent males appears almost incidental, and their age distribution is similar to the much larger female cohort. The story of any other males from this group is different and unknown, and their absence from both mass graves leaves room only for speculation.
Taphonomic and dating evidence suggest a coherent and short-duration killing and depositional horizon (Supplementary Notes 2 and 4). Cross-culturally, women and children are often seen as valuable (for example, enslavement, procreation, labour and loot) and may be preferentially taken as captives, and targeting them at Gomolava was evidently a meaningful choice48,49. While it is not unusual for women and children to be affected by violence during times of conflict, a mass grave almost exclusively containing women and children is a rarer, often much more recent and geoculturally more exclusive phenomenon. Even then, the context of killing is more typically associated with sacrificial practices or regional-scale war and conquest-related massacres50. The demographics of those killed at Gomolava accentuate the sense that this was a purposeful, calculated act to fragment a network of regional relationships and lineages impacting a wide physical and social landscape. At the same time, acknowledging the value of the victims to their societies transfers value to the place they were assembled together in death. Whether the males absent from mass graves 1 and 2 conducted the killing event(s), were spared for reasons unknown or were themselves killed and interred elsewhere, it was those buried in the monuments that established their identity as places of commemoration and community value. Constructing this sense of place suggests that the mass graves would have shaped and focused the collective memory of the victims’ communities, as well as of people from across and passing through the region, potentially over several generations. Collective loss may fuel resentment, yet the violent psychogeographical connotations of Gomolava thereafter may have amplified a message of dominance and forced cohesion51.
The killing of women and children also signals their wider social and economic importance and societal contribution43. Cognisant of the potential value of unfree individuals for labour, their killing demonstrates that the perpetrators were willing to dispose of a perceived form of wealth for purposes beyond simple extermination. The isotopic evidence seems to support the idea that the women may have been more mobile, which is in line with exogamic practices, bearing in mind the absence of males. Whether such movement was coerced, arranged or negotiated by kin groups to enhance their status, it is clear that they and their nuclear families served as fundamental links in a wider social network. This network perspective emphasizes the regional-scale impact of preferentially targeting women and children for death, severing these links, in the Gomolava massacre. Furthermore, the removal of those buried at Gomolava from their respective communities through brutal killing, and the commemoration of this event, can both be read as powerful yet pragmatic attempts to rebalance power relations and assert dominance over land and resources, whether or not these acts were performed by the same people. Taken together, the killing event, the mortuary event and the resulting monument signal a chain of actions intended to forcibly resolve or eradicate conflict and rebalance power within or between communities. The disruption or subdual of multiple identities, networks and traditions through this act of large-scale violence distilled into a single grave thereby reflects a phenomenon engulfing a larger region. The choices manifested through Gomolava mass grave 2 shed new light on targeted gender- and age-selective killings as a way of enacting mass violence and assertion of power in prehistoric Europe.
Methods
Ethics and permission
All human and artefact remains sampled are in the keeping of the Museum of Vojvodina, Serbia. Samples for this study were obtained following all criteria set out in official written permits from the museum and in collaboration with co-authors working there. All osteoarchaeological work for this publication was carried out following the British Association for Biological Anthropology and Osteoarchaeology Code of Ethics and Code of Practice. Material not consumed during analysis will be returned to the Museum of Vojvodina.
Excavation
Mass grave 2 was unexpectedly identified while excavating the upper layer of the tell. Excavation followed standard procedures of the time, which included digging in non-stratigraphically defined spits or passes of regular depth. This resulted in the remains being removed in three layers. Some individuals had elements recovered from more than one layer, and some individuals, as recovered, included bones from other individuals. Specialized bioarchaeological recovery approaches had not been developed and so were not implemented, resulting in this partial mixing during excavation. The upper layer of the burial was partly disturbed, but apparently not truncated substantially. Some human bone—particularly from infants—was recovered alongside and stored with animal bone, indicating the possibility that fragile remains, particularly those of infants, were missed during excavation7.
Histotaphonomy
Eight human bone samples were scanned using X-ray CT at the University College Dublin X-ray CT Facility to investigate patterns of diagenesis, particularly bacterial bioerosion, which could provide insight into the taphonomic histories of bones and bodies52. The scans were undertaken using a Phoenix Nanotom M scanner with the following settings: 120 kV, 140 μA, a resolution of 30.83 μm, a scan time of 9 min, and 1,079 projection images. The fast scan option was used, and a 0.5-mm Cu filter applied. All samples came from long bone shafts in an attempt to control for intraskeletal variation in bioerosion52. Levels of bioerosion were assessed by using the Oxford Histological Index53, which scores levels of bioerosion on a scale from 0 (no internal microstructure remaining) to 5 (internal microstructure is mostly intact).
Osteoarchaeological analyses
The analysis followed standard bioarchaeological methods for age and sex assessment as well as palaeopathological diagnosis54. Age categories were defined as neonate (around time of birth), infant (<1 year), juvenile (1–12 years), adolescent (13–17 years), young adult (18–25 years), young middle adult (26–35 years), old middle adult (36–45 years), mature adult (45+ years) and adult (>17 years)55. Pathological changes recorded included degenerative joint changes in adult individuals, as broadly related to levels of joint wear and physical activity56; stress markers, including cribra orbitalia26 and dental enamel hypoplasia57 in all individuals as related to episodes of systemic stress; reactive new bone formation in all individuals as related to localized and systemic infection or inflammation, which may at times also result from systemic stress26; and skeletal trauma in all individuals, as related to social complexity, activity, environment and violent conflict58.
Sampling strategy for biomolecular analyses
We sampled 27 individuals (35% of the assemblage) from the mass grave, including 16 petrous bones and 11 teeth, selecting a broadly representative subset of the original burial population sufficient to maximize multiproxy potential of the study. Individuals were selected without demographic criteria from all three of the as-excavated burial strata to ensure broad representation across the burial context. The decision not to sample the entire assemblage was intentional, with conservation and future analyses in mind, and in line with current best-practice guidelines (following BABAO Code of Practice, British Association for Biological Anthropology and Osteoarchaeology). Where possible, we prioritized bones that were not firmly fixed naturally or had not been glued during previous conservation work, as sampling such remains would potentially be damaging and could render samples unusable (for example, breaking tooth roots). All 27 individuals were analysed for ancient DNA. Of these, 24 were analysed for strontium (⁸⁷Sr/⁸⁶Sr), oxygen (δ18O), nitrogen (δ15N) and carbon (δ¹³C) isotopes, and 5 were used for radiocarbon (¹⁴C) dating. Sex determination was performed on an additional 26 individuals using a minimally destructive mass spectrometry protocol that did not require additional destructive sampling.
Radiocarbon dating
Radiocarbon dating was conducted at the Oxford Radiocarbon Accelerator Unit. Collagen was extracted from the bone samples and purified using ultrafiltration59. The main steps in this process were: sequential reaction with HCl (0.5 M, overnight), NaOH (0.1 M, 30 min) and HCl (0.5 M, 15 min) at room temperature to demineralize and remove humic acids, before gelatinization (0.001 M HCl, 75 °C, 20 h), filtration to remove insoluble contaminants (EzeeTM filters) and ultrafiltration to remove small soluble contaminants (precleaned Sartious VivaspinTM Turbo 15, 30 kDa molecular weight cut-off). Extracted collagen was combusted using an elemental analyser, and a portion of gas sent to an isotope ratio mass spectrometer (for example, Sercon 20-22). The remaining CO2 was cryogenically collected and converted to graphite by reaction with hydrogen over an iron catalyst60. Dates of five humans and one faunal bone (cow) were measured on a Mini Carbon Dating System (MICADAS) and calculated following61 using a background correction specific to bone62 and an accelerator mass spectrometry-derived δ13C (Supplementary Note 4).
Mass spectrometry-based sex determination
Sex determination of skeletal remains was performed by mass spectrometry detection of sex-specific amelogenin peptides as described by Shaw et al.63, a modification of Stewart et al.64 using teeth from 26 non-adult individuals that could not be sexed osteologically. In brief, the detection of sex-chromosome-encoded endogenous AMELX and AMELY peptides derived from the dental enamel protein amelogenin allows the assignment of XX or XY karyotypes. Peptides were etched from enamel samples in 5% (v/v) HCl and purified using Pierce C18 resin-loaded spin columns, with the final elution in 60% acetonitrile/0.1% formic acid being lyophilized. Purified samples were dissolved in 0.1% (v/v) aqueous trifluoroacetic acid and analysed with a Dionex U3000 nanoLC coupled to a ThermoFisher LTQ FT Ultra Mass Spectrometer containing a nano-electrospray ionization source63. The presence (or absence) of AMELX and AMELY peptides in enamel extracts was determined by plotting chromatograms for the transition m/z 540.4 to 714.4 (characteristic for AMELX), and m/z 440.4 to 645.4 (characteristic for AMELY). Reconstructed ion chromatograms were generated, allowing confident sex assignment for all 26 samples.
Isotope analyses
Strontium (87Sr/86Sr), oxygen (δ18Oen) and carbon (δ13Cen) isotopes were measured in dental enamel (n = 24 individuals). Strontium isotope measurements were also conducted on baseline plant samples (n = 8) from locations within and around the site to assess the local range of variation. Sample processing was conducted at the Faculty of Archaeology, Leiden University, using standard protocols for multi-isotope analyses of archaeological enamel and modern plants (for details, see Laffoon et al.65). Strontium was extracted using ion exchange chemistry, and 87Sr/86Sr was measured on a TritonPlus thermal ionization mass spectrometer at the Vrije Universiteit Amsterdam using standard protocols66. Strontium isotope ratios were corrected for mass fractionation to an 86Sr/88Sr of 0.1194. 87Sr/86Sr are reported as mean ± 2 standard error of the mean, based on the analytical uncertainty calculated from 240 cycles of 8.1 s integration time (12 blocks of 20 cycles). Oxygen and carbon isotope values were measured on a Finnigan DeltaPlus isotope ratio mass spectrometer, interfaced with a GasBench II autosampler via reaction with orthophosphoric acid (H3PO4) (100%) at 45 °C for 24 h. Enamel stable isotope data were normalized to the Vienna Peedee Belemnite scale using an in-house carbonate reference material (VICS) calibrated against NBS19 and LSVEC-certified reference materials. For δ13C and δ15N analysis, bone chunks of mostly petrous bone were placed in 0.5 M EDTA (pH 7.5), with the solution regularly refreshed until the bone was fully demineralized67. Collagen isomorphs were then rinsed 15 times in distilled water with an overnight soak on the tenth rinse. Human collagens were analysed for δ13C, δ15N, %C and %N at Centre National de la Recherche Scientifique (CNRS) on a Thermo Flash 2000 EA interfaced with a Thermo DeltaV Advantage at Service de Spectrométrie de Masse Isotopique du Muséum (SSMIM, Paris). Faunal collagens were analysed for δ13C, δ15N, %C and %N on a vario PYRO cube Elemental Analyzer coupled to an Isoprime visION mass spectrometer in continuous flow mode at the Archaeology Stable Isotope Laboratory, University of Kiel. Stable isotope values were calibrated relative to V-PDB (Vienna PeeDee Belemnite, δ13C) and AIR (Ambient Inhalable Reservoir, δ15N) using international glutamic acid standards USGS40 (δ13C − 26.39 ± 0.04‰; δ15N − 4.52 ± 0.06‰) and USGS41a (δ13C − 37.63 ± 0.05‰; δ15N + 47.55 ± 0.15‰)68. Measurement uncertainty was surveyed via two internal standards, casein and cattle bone collagen (long-term averages calibrated to V-PDB and AIR with USGS40 and USGS41a).
Ancient DNA
Laboratory work
All samples were processed at the dedicated clean-lab facilities at the Globe Institute in Copenhagen, Denmark. Negative controls were included along the entire workflow to account for potential background contamination of the lab. Inside the clean lab, the samples were cleaned using a tissue dipped in a 5% bleach solution. The outermost layer was removed using a sterile disc drill to avoid surface contamination. The teeth were subsampled by cutting the tooth roots off and targeting the cementum69, while the petrous bones were sampled by isolating the otic capsule70. Subsequently, the samples were crushed into small chunks using a pestle and a mortar. DNA was extracted following standard protocols71, including a 10-min ‘predigestion’ step72. The DNA extracts were built into libraries using single-stranded library protocols both without USER treatment and following Uracil-Specific Excision Reagent (USER) treatment to reduce the effect of post-mortem DNA damage73. The libraries were sequenced first on two lanes of an Illumina HiSeq 2500 sequencer 75-bp single-end mode and then in three lanes of an Illumina NovaSeq SP flowcell run in 50-bp paired-end mode.
Data processing and authentication
Base calling was done with bcl2fastq2 v2.20.0, and libraries were demultiplexed by requiring an exact match to the eight-nucleotide indices assigned during preparation. Processing of the raw sequence data was conducted with the nf-core/eager pipeline v2.4.474. Adapter sequences were trimmed with AdapterRemoval v2.075, keeping only reads longer than 30 bp and with a minimum base quality score of 20. The resulting reads were aligned to the human reference genome hg19 using the bwa v0.7.576 with seeding disabled (-l 1024, -n 0.04), and a mapping quality filter of 30 was applied using SAMtools v1.3. To evaluate DNA damage patterns, we used DamageProfiler77, and contamination levels were estimated directly from the bam files using hapCon78 and contamMix v1.0.1179. Sequencing statistics and contamination estimates are presented in Supplementary Tables 13 and 14.
Imputation
We performed imputation of the genome using GLIMPSE v1.1.180, following the standard procedures outlined at https://odelaneau.github.io/GLIMPSE/docs/tutorials and using the 1000 Genomes phase 3 panel as a ref. 81. To prevent batch effects, we imputed each genome individually. We then filtered the imputed genomes following the guidelines outlined by Sousa da Mota et al.82, excluding all locations situated in regions identified to harbour repeats, and removed sites with QUAL <30 (Phred-scaled probability that the site has no variant).
Genetic sex determination and uniparental markers
Genetic sex was determined with Karyo_RxRy, which distinguishes between all sex chromosome karyotypes, as well as detecting autosomal aneuploidies such as trisomy 21 (ref. 83). Mitochondrial haplogroups were determined with Haplogrep284 as implemented in Haplocheck85. The Y-chromosome haplogroups were determined using Yleaf86, a workflow designed for Y-chromosome phylogeny placements using low-coverage ancient DNA data. Results are presented in Supplementary Tables 16 and 17.
Genetic relatedness and runs of homozygosity
We inferred the first-, second- and third-degree genetic relatedness between all pairs of individuals in our dataset using IBSrelate87 as implemented in NgsRelate v/288 and BREADR89. To evaluate the level of consanguinity at Gomolava, we estimated runs of homozygosity using Plink90, using the parameters outlined by Ceballos et al.91 (--homozyg-snp 50 --homozyg-kb 300 --homozyg-density 50 --homozyg-gap 100 --homozyg-window-snp 50--homozyg-window-het 1 --homozyg-window-threshold 0.05).
AncIBD and HapNe-LD
We used ancIBD92 on the imputed data to detect genomic segments that are IBD. In this analysis, we focused on the 1,240,000 SNPs and set a coverage cut-off, ensuring that individuals with at least 70% of those SNPs had an imputed genotype probability greater than 0.99. The identified IBD segments were refined by including a minimum segment length of 8 cM, and summary statistics were recorded for both the count and total length of segments exceeding 8, 12, 16 and 20 cM, along with the longest contiguous IBD block (Supplementary Table 20). Expected IBD sharing under various constant population sizes was calculated as described by Fernandes et al.93. Finally, we calculated the effective population size using HapNe-LD94 on the imputed data, which was translated into the census size following the methodology described by Pankratov et al.95.
MDS
To assess the genetic ancestry and affinities, we called pseudohaploid genotypes on the samples using samtools mpileup, filtering reads with a minimum base quality of 20 (-Q 20) at all 1,240,000 positions and randomly drawing one of them. We then merged our dataset with relevant published ancient individuals from the Allen Ancient DNA Resource v5496, to which we added the recently published ancient DNA data from Gerber et al.97. To avoid bias when co-analysing the captured and shotgun data98, we restricted the SNPs in the dataset to those recommended by Rohland et al.99. We used plink v1.9 to filter for missingness and removed all SNPs with missing call rates >90% (--geno 0.90) and individuals with missing genotyping rate >90% (--mind 0.9). The final ancient reference panel includes 869 ancient individuals covering 597,573 SNPs (Supplementary Table 27). On this dataset, we performed MDS by computing pairwise distances between all individuals using plink v1.9 with the parameters --distance square gz ‘flat-missing’. We then normalized these distances to the total number of SNPs and used the R function cmdscale to compute the ten first dimensions of the MDS.
Ancestry modelling
We used qpWave and the pAdm100 on the pseudohaploid genotypes as implemented in ADMIXTOOLS2 to estimate the minimum number of independent gene pools that explain our set of individuals and to model them as a mixture of contributions from n source populations.
Pathogen screening
To characterize the composition of the microbial communities, we aligned the host-removed sequences of each sample to a database comprising GTDB v202101, all mitochondrial and chloroplast genomes from NCBI, and CheckV v0.6102 using bowtie2 v2.5.2103 in very sensitive mode and allowing a maximum of 16 alignments per read. We used filter-bam 1.2.1 to exclude false-positive matches by filtering out alignments with an average nucleotide identity below 95%, an expected/observed breadth of coverage ratio below 70% and fewer than 500 aligned reads. The deamination rate at the 5′ end was computed for the retained alignments using metaDMG104 in local mode. Mapping statistics for alignments with at least 3% damage at the 5′ end and a MAP significance (how many sigmas the damage is away from 0 in metaDMG) of more than 2 are reported in Supplementary Table 28. To detect viral pathogens and parasites such as Plasmodium falciparum/vivax, we employed the metagenomicscreening workflow Pathopipe105 using a viral database of 10,179 taxa with 267,205 genomes, and a parasite database with 306 complete endoparasitic genomes from GenBank/RefSeq, respectively. After this initial screening, we filtered the resulting hits using the following criteria: (1) number of reads ≥20 for viruses and ≥200 for parasites; (2) ratio of observed/expected breadth of coverage ≥0.9; (3) average number of soft-clipped bases in alignment <5; (4) average edit distance ≤1; and (5) Unique Kmer rank equal to 1.
Metallurgical analysis
Samples of metal objects were taken at the National Museum of Vojvodina in Novi Sad, and all processing and analysis were completed at the National Centre for Isotope Geochemistry in the School of Earth Sciences at University College Dublin. Samples were taken either by drilling or cutting with a jeweller’s saw and subsequently manually cleaned to remove corrosion in drillings or on surfaces of cut fragments. Samples of 5–10 mg were weighed on an analytical scale. The wet chemistry protocol was developed following Balliana et al. Between 5 mg and 10 mg of sample were digested in 2 ml 8 M HNO3 and 20 μl HF on a hotplate overnight at ~100 °C. The solution was then diluted and aliquoted for chemical analysis and lead isotope analysis, respectively, after subsequent treatment. The first aliquot was diluted with 0.5 M HNO3 in multiple stages for the analysis of trace and major elements. The second aliquot was subjected to column chromatography with Sr-spec resin (SR-B25-A, Triskem) to purify lead. Interferences and contaminants were eluted before the collection of Pb in 6 M HCl. Samples were dried then treated with ~14 M HNO3, then dried again to finally be collected with 0.5 M HNO3.
Chemical composition analysis was performed on a Thermo iCAP Q Quadrupole ICP-MS in high matrix and collision cell mode, with He as collision cell gas (4.85 ml min−1), 20 runs with 10 sweeps each, 1 peak per mass and a dwell time of 20 ms (200 ms for As). For calibration, Sigma-Aldrich TraceCert multi-element solutions mix 1 and mix 2 were used in dilution between 1 ppb and 200 ppb. For Cu and Sn analysis, single-element standards (Alfa Aesar, Specpure) were mixed and diluted. Data evaluation was performed using the Qtegra software package. Average uncertainties are 1.5–2.5% (1 s.d.). The raw concentrations were normalized to 100% to avoid artificial differences in concentrations (totals >88% except G1741, G1740 and G1783 totals between 70% and 76%). Low totals may be caused by loss of material during weighing due to static, or by the presence of small amounts of light elements not analysed (for example, O and S); these do not affect the relative proportions of elements, assuming the metal is homogeneous. Lead isotope analysis was performed with a ThermoScientific Neptune multicollector inductively coupled plasma mass spectrometer. For mass bias correction, solutions were doped with Tl and corrected for 203Tl/205Tl of 0.418922. Data quality was monitored using the NIST SRM981 standard, which yielded values of 206Pb/204Pb of 16.9355 ± 0.0002, 207Pb/204Pb of 15.4905 ± 0.0002, 208Pb/204Pb of 36.6942 ± 0.0005, 208Pb/206Pb of 2.16671 ± 0.00001 and 207Pb/206Pb of 0.914681 ± 0.000007 (n = 6) on the day of analysis.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The DNA sequences reported in this Article have been deposited in the European Nucleotide Archive under the accession number PRJEB84956. All other data used in this study are available in the Supplementary Information.
References
Haas, J. The Anthropology of War (Cambridge Univ. Press, 1990).
Molloy, B., Bruyère, C. & Jovanović, D. in Rethinking Migrations in Late Prehistoric Eurasia (eds Fernández-Götz, M. et al.) 142–169 (British Academy, 2022); https://doi.org/10.5871/bacad/9780197267356.003.0007
Milner, G. R. Trouble in paradise. Legacy review of: War before civilization. By Lawrence H.Keeley, New York, NY: Oxford University Press. 1996. xiv + 245 pp. ISBN: 0-19-509112-4 (hardback). Am. J. Biol. Anthropol. https://doi.org/10.1002/ajpa.24924 (2024).
Koziol, K. M. Shattered mirrors: gender, age, and Westernized interpretations of war (and violence) in the past. In Bioarchaeology of Women and Children in Times of War (eds Martin, D.L. & Tegtmeyer C) 15–26 (Springer, 2017).
Brukner, B. Naselje Vinčanske grupe na Gomolavi (neolitski i ranoeneolitski sloj). Izveštaj sa iskopavanja 1967–1976, Rad Vojvođanskih Muzeja 26, 5–55 (1980).
Molloy, B. et al. Resilience, innovation and collapse of settlement networks in later Bronze Age Europe: new survey data from the southern Carpathian Basin. PLoS ONE 18, e0288750 (2023).
Tasić, N. An Early Iron Age collective tomb at Gomolava. Archaeologia Iugoslavica XIII 27–37 (1972).
Medović, P. & Medović, I. Gradina na Bosutu: naselje starijeg gvozdenog doba (Ildiko Medovic, 2012).
Gamba, C. et al. Genome flux and stasis in a five millennium transect of European prehistory. Nat. Commun. 5, 5257 (2014).
Metzner-Nebelsick, C. The first evidence for mounted nomads between the Caucasus and central Europe in the early 1st millennium BC. In Crossing Boundaries - Mounted Nomads in Central Europe, Their Eastern Roots and Connections; Tagungen des Landesmuseums für Vorgeschichte Halle (eds Meller, H. & Daim, F.) 125–149 (2022).
Mirazón Lahr, M. et al. Inter-group violence among early Holocene hunter-gatherers of West Turkana, Kenya. Nature 529, 394–398 (2016).
Meyer, C. Early massacres: mass violence in Neolithic Europe. In The Cambridge World History of Violence (eds Fagan, G. G. et al.) 299–319 (Cambridge Univ. Press, 2020).
Fibiger, L., Ahlström, T., Meyer, C. & Smith, M. Conflict, violence, and warfare among early farmers in Northwestern Europe. Proc. Natl Acad. Sci. USA 120, e2209481119 (2023).
Molloy, B. & Horn, C. Weapons, warriors and warfare in Bronze Age Europe. In The Cambridge World History of Violence (eds Fagan, G. G. et al.) 117–141 (Cambridge Univ. Press, 2020).
Horn, C. Trouble in paradise?—Violent conflict in funnel-beaker societies. Oxf. J. Archaeol. 40, 43–64 (2021).
Brinker, U. et al. Die menschlichen Skelettreste aus dem Tollensetal: Ein Vorbericht. Tod im Tollensetal-Forschungen zu den Hinterlassenschaften eines bronzezeitlichen Gewaltkonfliktes in Mecklenburg-Vorpommern. Teil 1, 191–208 (2014).
Redfern, R. Gendered violence in Iron Age and Roman Britain. In The Cambridge World History of Violence (eds Fagan, G. G. et al.) 320–341 (Cambridge Univ. Press, 2020).
Király, A., Sebők, K., Zoffmann, Z. K., Kovács, G. & Müller-Scheeßel, N. in Irreguläre Bestattungen in der Urgeschichte: Norm, Ritual, Strafe (ed. Müller-Scheeßel, N.) 307–326 (Dr. Rudolf Habelt GmbH, 2013).
Keeley, L. H. War Before Civilization (OUP USA, 1996).
Martin, D. L. & Tegtmeyer, C. Bioarchaeology of Women and Children in Times of War: Case Studies from the Americas (Springer, 2017).
Waddington, C. et al. Excavations at Fin Cop, Derbyshire: an Iron Age Hillfort in conflict?. Archaeol. J. 169, 159–236 (2012).
Farkas, G. & Marcsik, A. Das Sammelgrab von Gomolava (Jugoslavien) aus der Urzeit. Anthropologie 14, 93–96 (1976).
Zoffmann, Z. K. Anthropologische Charakteristik der Bevölkerung der Frûheisen zeitlichen Bosut-Kultur Aufgrund des Kollektivgrabes von Gomolava. Balcanica XXVIII 249–258 (1997).
Baten J, S. R. H. in The Backbone of Europe: Health, Diet, Work and Violence over Two Millennia. Cambridge Studies in Biological and Evolutionary Anthropology (eds Steckel R. H. et al.) 300–324 (Cambridge Univ. Press, 2018).
Fibiger, L. Misplaced childhood? In The Routledge Handbook of the Bioarchaeology of Human Conflict (eds Knüsel, C. & Smith, M.) 127–145 (Routledge, 2014).
Ortner, D. J. Identification of Pathological Conditions in Human Skeletal Remains (Academic Press, 2003).
Fibiger, L., Ahlström, T., Bennike, P. & Schulting, R. J. Patterns of violence-related skull trauma in Neolithic Southern Scandinavia. Am. J. Phys. Anthropol. 150, 190–202 (2013).
Caballero, M. et al. Crossover interference and sex-specific genetic maps shape identical by descent sharing in close relatives. PLoS Genet. 15, e1007979 (2019).
Gelabert, P. et al. Social and genetic diversity in first farmers of central Europe. Nat. Hum. Behav. https://doi.org/10.1038/s41562-024-02034-z (2024).
Novak, M. et al. Genome-wide analysis of nearly all the victims of a 6200 year old massacre. PLoS ONE 16, e0247332 (2021).
Schroeder, H. et al. Unraveling ancestry, kinship, and violence in a Late Neolithic mass grave. Proc. Natl Acad. Sci. USA 116, 10705–10710 (2019).
Furholt, M. Translocal communities—exploring mobility and migration in sedentary societies of the European Neolithic and Early Bronze Age. Praehist. Z. 92, 304–321 (2018).
Metzner-Nebelsick, C. in Rethinking Migrations in Late Prehistoric Eurasia (eds Fernández-Götz, M. et al.) 209–233 (Oxford Univ. Press, 2023).
Bruyère, C. et al. Trade, recycling and mixing in local metal management strategies of the later Bronze Age south Carpathian Basin: lead isotope and chemical analyses of hoarded metalwork. J. Archaeol. Sci. 164, 105957 (2024).
Giblin, J. Isotope Analysis on the Great Hungarian Plain: An Exploration of Mobility and Subsistence Strategies from the Neolithic to the Copper Age (Archaeolingua, 2020); https://www.archaeolingua.hu/book/isotope-analysis-great-hungarian-plain-exploration-mobility-and-subsistence-strategies
Cavazzuti, C. et al. Human mobility in a Bronze Age Vatya ‘urnfield’ and the life history of a high-status woman. PLoS ONE 16, e0254360 (2021).
Maier, R. et al. On the limits of fitting complex models of population history to F-statistics. eLife 12, e85492 (2023).
Allentoft, M. E. et al. Population genomics of Bronze Age Eurasia. Nature 522, 167–172 (2015).
Freilich, S. et al. Reconstructing genetic histories and social organisation in Neolithic and Bronze Age Croatia. Sci. Rep. 11, 16729 (2021).
Brück, J. Ancient DNA, kinship and relational identities in Bronze Age Britain. Antiquity 95, 228–237 (2021).
Driessen, J. in An Archaeology of Forced Migration: Crisis-Induced mobility and the Collapse of the 13th c. BCE Eastern Mediterranean 19–24 (Presses Universitaires de Louvain, 2018).
Linduff, K. M. & Rubinson, K. S. Are All Warriors Male?: Gender Roles on the Ancient Eurasian Steppe (AltaMira Press, 2008).
Pope, R. E. Gender and society. In The Oxford Handbook of the European Iron Age (eds Haselgrove, C. et al.) 1049–1078 (Oxford University Press, 2017).
Karmebäck, V. N., Wairore, J. N., Jirström, M. & Nyberg, G. Assessing gender roles in a changing landscape: diversified agro-pastoralism in drylands of West Pokot, Kenya. Pastoralism 5, 1–8 (2015).
Diversity of thought and data enrich archaeology. Nat. Hum. Behav. 8, 183–184 (2024).
Fontijn, D. Economies of Destruction: How the Systematic Destruction of Valuables Created Value in Bronze Age Europe, c. 2300–500 BC (Routledge, 2019).
Janković, I., Balen, J., Potrebica, H., Ahern, J. C. M. & Novak, M. Mass violence in Copper Age Europe: the massacre burial site from Potočani, Croatia. Am. J. Phys. Anthropol. 176, 474–485 (2021).
Cameron, C. M. Captives and culture change. Curr. Anthropol. 52, 169–209 (2011).
Martin, D. L. Hard times in dry lands: making meaning of violence in the ancient southwest. J. Anthropol. Res. 72, 1–23 (2016).
Greene, J. A. & Scott, D. D. Finding Sand Creek: History, Archeology, and the 1864 Massacre Site (Univ. Oklahoma Press, 2004).
Redfern, R. Iron Age ‘predatory landscapes’: a bioarchaeological and funerary exploration of captivity and enslavement in Britain. Cam. Archaeol. J. 30, 531–554 (2020).
Jans, M. M. E., Nielsen-Marsh, C. M., Smith, C. I., Collins, M. J. & Kars, H. Characterisation of microbial attack on archaeological bone. J. Archaeol. Sci. 31, 87–95 (2004).
Millard, A. R. in Handbook of Archaeological Sciences (eds Brothwell, D. R. & Pollard, A. M.) 637–647 (Wiley, 2001).
Buikstra, J. & Ubelaker, D. Standards for data collection from human skeletal remains. Arkansas archaeological survey research series (1994).
Mitchell, P. D. & Brickley, M. Updated Guidelines to the Standards for Recording Human Remains (Chartered Institute for Archaeologists, 2017).
Rogers, J. & Waldron, T. A Field Guide to Joint Disease in Archaeology (John Wiley & Sons, 1995).
King, T., Humphrey, L. T. & Hillson, S. Linear enamel hypoplasias as indicators of systemic physiological stress: evidence from two known age-at-death and sex populations from postmedieval London. Am. J. Phys. Anthropol. 128, 547–559 (2005).
Lewis, J. E. Identifying sword marks on bone: criteria for distinguishing between cut marks made by different classes of bladed weapons. J. Archaeol. Sci. 35, 2001–2008 (2008).
Brock, F., Higham, T., Ditchfield, P. & Ramsey, C. B. Current pretreatment methods for AMS radiocarbon dating at the Oxford Radiocarbon Accelerator Unit (ORAU). Radiocarbon 52, 103–112 (2010).
Dee, M. & Bronk Ramsey, C. Refinement of graphite target production at ORAU. Nucl. Instrum. Methods Phys. Res. B 172, 449–453 (2000).
Stuiver, M. & Polach, H. A. Discussion reporting of 14C data. Radiocarbon 19, 355–363 (1977).
Wood, R. E., Bronk Ramsey, C. & Higham, T. F. G. Refining background corrections for radiocarbon dating of bone collagen at ORAU. Radiocarbon 52, 600–611 (2010).
Shaw, B. et al. Antibody-based sex determination of human skeletal remains. iScience 26, 108191 (2023).
Stewart, N. A., Gerlach, R. F., Gowland, R. L., Gron, K. J. & Montgomery, J. Sex determination of human remains from peptides in tooth enamel. Proc. Natl Acad. Sci. USA 114, 13649–13654 (2017).
Laffoon, J. E., Davies, G. R., Hoogland, M. L. P. & Hofman, C. L. Spatial variation of biologically available strontium isotopes (87Sr/86Sr) in an archipelagic setting: a case study from the Caribbean. J. Archaeol. Sci. 39, 2371–2384 (2012).
Kootker, L. M., van Lanen, R. J., Kars, H. & Davies, G. R. Strontium isoscapes in The Netherlands. Spatial variations in 87Sr/86Sr as a proxy for palaeomobility. J. Archaeol. Sci. Rep. 6, 1–13 (2016).
Tuross, N., Fogel, M. L. & Hare, P. E. Variability in the preservation of the isotopic composition of collagen from fossil bone. Geochim. Cosmochim. Acta 52, 929–935 (1988).
Qi, H., Coplen, T. B., Geilmann, H., Brand, W. A. & Böhlke, J. K. Two new organic reference materials for delta13C and delta15N measurements and a new value for the delta13C of NBS 22 oil. Rapid Commun. Mass Spectrom. 17, 2483–2487 (2003).
Pinhasi, R. et al. Optimal ancient DNA yields from the inner ear part of the human petrous bone. PLoS ONE 10, e0129102 (2015).
Rohland, N. & Hofreiter, M. Ancient DNA extraction from bones and teeth. Nat. Protoc. 2, 1756–1762 (2007).
Damgaard, P. B. et al. Improving access to endogenous DNA in ancient bones and teeth. Sci. Rep. 5, 11184 (2015).
Rohland, N., Harney, E., Mallick, S., Nordenfelt, S. & Reich, D. Partial uracil–DNA–glycosylase treatment for screening of ancient DNA. Philos. Trans. R. Soc. Lond. B 370, 20130624 (2015).
Fellows Yates, J. A. et al. Reproducible, portable, and efficient ancient genome reconstruction with nf-core/eager. PeerJ 9, e10947 (2021).
Schubert, M., Lindgreen, S. & Orlando, L. AdapterRemoval v2: rapid adapter trimming, identification, and read merging. BMC Res. Notes 9, 88 (2016).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Neukamm, J., Peltzer, A. & Nieselt, K. DamageProfiler: fast damage pattern calculation for ancient DNA. Bioinformatics https://doi.org/10.1093/bioinformatics/btab190 (2021).
Huang, Y. & Ringbauer, H. hapCon: estimating contamination of ancient genomes by copying from reference haplotypes. Bioinformatics https://doi.org/10.1093/bioinformatics/btac390 (2022).
Fu, Q. et al. A revised timescale for human evolution based on ancient mitochondrial genomes. Curr. Biol. 23, 553–559 (2013).
Rubinacci, S., Ribeiro, D. M., Hofmeister, R. J. & Delaneau, O. Efficient phasing and imputation of low-coverage sequencing data using large reference panels. Nat. Genet. 53, 120–126 (2021).
Auton, A. & Salcedo, T. in Assessing Rare Variation in Complex Traits: Design and Analysis of Genetic Studies (eds Zeggini, E. & Morris, A.) 71–85 (Springer, 2015).
Sousa da Mota, B. et al. Imputation of ancient human genomes. Nat. Commun. 14, 3660 (2023).
Anastasiadou, K. et al. Detection of chromosomal aneuploidy in ancient genomes. Commun. Biol. 7, 14 (2024).
Weissensteiner, H. et al. HaploGrep 2: mitochondrial haplogroup classification in the era of high-throughput sequencing. Nucleic Acids Res. 44, W58–W63 (2016).
Weissensteiner, H. et al. Contamination detection in sequencing studies using the mitochondrial phylogeny. Genome Res. https://doi.org/10.1101/gr.256545.119 (2021).
Ralf, A., Montiel González, D., Zhong, K. & Kayser, M. Yleaf: software for human Y-chromosomal haplogroup inference from next-generation sequencing data. Mol. Biol. Evol. 35, 1291–1294 (2018).
Waples, R. K., Albrechtsen, A. & Moltke, I. Allele frequency-free inference of close familial relationships from genotypes or low-depth sequencing data. Mol. Ecol. 28, 35–48 (2019).
Korneliussen, T. S. & Moltke, I. NgsRelate: a software tool for estimating pairwise relatedness from next-generation sequencing data. Bioinformatics 31, 4009–4011 (2015).
Rohrlach, A. B. et al. BREADR: an R Package for the Bayesian estimation of genetic relatedness from low-coverage genotype data. J. Open Source Softw. 10, 7916 (2025).
Purcell, S. et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575 (2007).
Ceballos, F. C., Hazelhurst, S. & Ramsay, M. Assessing runs of Homozygosity: a comparison of SNP Array and whole genome sequence low coverage data. BMC Genomics 19, 106 (2018).
Ringbauer, H. et al. Accurate detection of identity-by-descent segments in human ancient DNA. Nat. Genet. 56, 143–151 (2024).
Fernandes, D. M. et al. A genetic history of the pre-contact Caribbean. Nature 590, 103–110 (2021).
Fournier, R., Tsangalidou, Z., Reich, D. & Palamara, P. F. Haplotype-based inference of recent effective population size in modern and ancient DNA samples. Nat. Commun. 14, 7945 (2023).
Pankratov, V. et al. Differences in local population history at the finest level: the case of the Estonian population. Eur. J. Hum. Genet. 28, 1580–1591 (2020).
Mallick, S. et al. The Allen Ancient DNA Resource (AADR): a curated compendium of ancient human genomes. Sci. Data 11, 182 (2024).
Gerber, D. et al. Interdisciplinary analyses of Bronze Age communities from Western Hungary Reveal complex population histories. Mol. Biol. Evol. 40, (2023).
Davidson, R. et al. Allelic bias when performing in-solution enrichment of ancient human DNA. Mol. Ecol. Resour. 23, 1823–1840 (2023).
Rohland, N. et al. Three assays for in-solution enrichment of ancient human DNA at more than a million SNPs. Genome Res. 32, 2068–2078 (2022).
Patterson, N. et al. Ancient admixture in human history. Genetics 192, 1065–1093 (2012).
Parks, D. H. et al. GTDB: an ongoing census of bacterial and archaeal diversity through a phylogenetically consistent, rank normalized and complete genome-based taxonomy. Nucleic Acids Res. 50, D785–D794 (2022).
Nayfach, S. et al. CheckV assesses the quality and completeness of metagenome-assembled viral genomes. Nat. Biotechnol. 39, 578–585 (2021).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Michelsen, C. et al. metaDMG—a fast and accurate ancient DNA damage toolkit for metagenomic data. Preprint at bioRxiv https://doi.org/10.1101/2022.12.06.519264 (2022).
Sikora, M. et al. The spatiotemporal distribution of human pathogens in ancient Eurasia. Nature 643, 1011–1019 (2025).
Balliana, E., Aramendía, M., Resano, M., Barbante, C. & Vanhaecke, F. Copper and tin isotopic analysis of ancient bronzes for archaeological investigation: development and validation of a suitable analytical methodology. Anal. Bioanal. Chem. 405, 2973–2986 (2013).
Acknowledgements
This research was funded through the ERC Consolidator Grant THE FALL OF 1200 BC project (GA #772753) awarded to B.M. We thank colleagues at the Museum of Vojvodina at Novi Sad for their unstinting support, advice and provision of research resources. We thank G. Kavanagh and S. Kehoe for their support with micro-CT scanning, F. Demeter, M. Amzirkov, J. Pendić and F. Molloy for additional sampling assistance, and A. Robinson for bioarchaeology and D. Jovanvić for archaeology advice. We also thank L. Cassidy and N. Carlin for their helpful comments on an earlier version of the manuscript.
Author information
Authors and Affiliations
Contributions
B.M., L.F. and H.S. conceptualized this work. L.F., M.I.O., J.K., J.L., C.A.M., D.M., A.K.K., C.B., T.B., C.B.R., R.L., L.A., D.R., N.J.O., B.S., S.T., C.W.-S., D.V.A, H.S. and B.M. generated and curated the data. L.F., M.I.O., J.K., J.E.L., C.A.M., C.B., T.B., C.B.R., R.L., Y.H., J.N., J.S.D., D.V.A, A.M., J.R.-M., H.R., H.S. and B.M. analysed and interpreted the data. L.F., M.I.O., J.R.-M., H.S. and B.M. consolidated the data analysis. L.F., M.I.O., J.E.L., C.A.M., C.B., B.M., J.K., C.B.R., R.L., S.T., S.N. and H.S. prepared the images. B.M. acquired funding. L.F. and M.I.O. wrote the original draft with input from B.M. and H.S. and written contributions or editorial input by all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Human Behaviour thanks the anonymous reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information (download PDF )
Supplementary Note 1. Cultural and archaeological context of the site of Gomolava. Supplementary Note 2. Histotaphonomic analysis of human bone samples. Supplementary Note 3. Mass-spectrometry-based sex determination. Supplementary Note 4. Site chronology. Supplementary Note 5. Skeletal trauma and stress indicators. Supplementary Note 6. Geospatial origins and diet patterns. Supplementary Note 7. Metalwork analysis. Supplementary Note 8. Biological relatedness. Supplementary Note 9. Uniparental markers. Supplementary Note 10. Ancestry estimation and modelling. Supplementary Note 11. Demographic inferences (hapNe-LD). Supplementary Note 12. SNPs associated with phenotypic traits. Supplementary Note 13. Pathogen screening (Includes Supplementary Tables 1–28 and Figs. 1–22).
Supplementary Tables 1–28 (download XLSX )
Supplementary Table 1. Results of the histotaphonomic analysis of human bone samples. Supplementary Table 2. Sex determinations of a subsample of 26 teeth determined using liquid chromatography–tandem mass spectrometry analysis of AMELX and AMELY peptides from tooth enamel. Supplementary Table 3. The chemical pretreatment and stable isotope data for the six radiocarbon-dated samples. Supplementary Table 4. The radiocarbon dating, calibration and modelling results for the samples dated from Gomolava. Supplementary Table 5. Age and sex distribution of individuals across various methods. Supplementary Table 6. Cribra orbitalia and linear enamel hypoplasia at Gomolava. Supplementary Table 7. Violence-related trauma at Gomolava. Supplementary Table 8. Enamel and plant isotope results. Supplementary Table 9. Carbon and nitrogen isotope results. Supplementary Table 10. Lead isotope analysis results. Supplementary Table 11. Chemical analysis results in weight %. Supplementary Table 12. Summary of individuals with genomic data. Supplementary Table 13. Sequencing statistics. Supplementary Table 14. Contamination estimates. Supplementary Table 15. Genetic sexing results and chr 21 trisomy analysis. Supplementary Table 16. mtDNA haplogroups of the newly reported individuals estimated using Haplogrep2. Supplementary Table 17. Y chromosomal haplogroups of the newly reported individuals using Yleaf. Supplementary Table 18. Genetic relatedness estimation using BREADR. Supplementary Table 19. Genetic relatedness using IBSrelate as implemented in ngsRelate. Supplementary Table 20. Pairwise IBD sharing estimated using ancIBD. Supplementary Table 21. Runs of homozygosity detected by Plink. Supplementary Table 22. qpWave for newly reported individuals using RIGHT1 and RIGHT2 (in parentheses) as reference. Supplementary Table 22b. P values for the qpWave models. qpWaves between Sk41 and Sk53 that show some evidence of ancestry heterogeneity, against all other individuals from Gomolava. Supplementary Table 23. qpAdm model for individuals and clustered group using distal sources. Supplementary Table 24. Effective population size through generations estimated using HapNe-LD. Supplementary Table 25: Results from HIrisPlex per individual. Supplementary Table 26a. Functional SNPs. SNP analysis for selected phenotypic traits. Supplementary Table 26b. Malaria-resistance-related alleles. Supplementary Table 27. Ancient comparative datasets analysed in this study. Supplementary Table 28. Metagenomic classification at the species level.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Fibiger, L., Iraeta-Orbegozo, M., Koledin, J. et al. A large mass grave from the Early Iron Age indicates selective violence towards women and children in the Carpathian Basin. Nat Hum Behav 10, 656–668 (2026). https://doi.org/10.1038/s41562-025-02399-9
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41562-025-02399-9
