
Abstract
Human activities drive a wide range of environmental pressures, including habitat change, pollution and climate change, resulting in unprecedented effects on biodiversity1,2. However, despite decades of research, generalizations on the dimensions and extent of human impacts on biodiversity remain ambiguous. Mixed views persist on the trajectory of biodiversity at the local scale3 and even more so on the biotic homogenization of biodiversity across space4,5. We compiled 2,133 publications covering 97,783 impacted and reference sites, creating an unparallelled dataset of 3,667 independent comparisons of biodiversity impacts across all main organismal groups, habitats and the five most predominant human pressures1,6. For all comparisons, we quantified three key measures of biodiversity to assess how these human pressures drive homogenization and shifts in composition of biological communities across space and changes in local diversity, respectively. We show that human pressures distinctly shift community composition and decrease local diversity across terrestrial, freshwater and marine ecosystems. Yet, contrary to long-standing expectations, there is no clear general homogenization of communities. Critically, the direction and magnitude of biodiversity changes vary across pressures, organisms and scales at which they are studied. Our exhaustive global analysis reveals the general impact and key mediating factors of human pressures on biodiversity and can benchmark conservation strategies.
Similar content being viewed by others

Main
Biodiversity change poses a critical threat to human societies from local to global scales, highlighting the urgent need for understanding the complex relationship between human pressures and their effects on ecosystems1. Human pressures, broadly classified in five main types—land-use change, resource exploitation, pollution, climate change and invasive species6—can enhance or reduce species diversity locally. Crucially, by impacting biodiversity at local scales, effects of human pressures can similarly impact biodiversity patterns among communities at broader spatial scales. This includes shifts in species composition among biological communities across a region, as well as increases and decreases in similarity between communities (homogenization and differentiation, respectively).
Despite decades of accumulating evidence of human impacts on biological communities, the trajectory of biodiversity in the Anthropocene remains unclear and attempts for syntheses have yielded mixed and debated results for both local diversity7,8,9,10,11,12 and composition of communities across space4,5,13,14. Understanding and generalizing the impacts of human pressures on biodiversity and how they are mediated by key factors, such as the spatial scale or type of pressure, remains a major challenge. Such information is critical if we are to understand whether the actions taken to prevent further loss and change of contemporary biodiversity are successful and to give insights into appropriate strategies to monitor the success of mitigation actions.
Crucially, previous research attempting to generalize biodiversity change has hitherto neglected two key elements. First, most past studies looked at biodiversity change across time using individual time series and did not contrast findings to reference controls9. Second, previous studies have rarely differentiated between changes in local diversity versus changes in variation in diversity across space. Unfortunately, the studies that have integrated these elements are generally restricted to a certain type of pressure or to a particular biome15,16,17,18,19. Consequently, we lack generalizations on the effects of human pressures on ecosystems and our understanding of biodiversity change with regard to its different dimensions remains incomplete. Thus, we are limited in our capacity to disentangle its underlying drivers. Given the multifaceted aspect of biodiversity and the plurality of drivers, organisms and spatial scales, the present lack of synthetic understanding and attribution of general impacts of human pressures on biological communities is hindering adequate actions and mitigation strategies20,21.
Here we compiled and analysed a large dataset to assess the impacts of human pressures on biodiversity, systematically contrasting impacted versus reference communities. We first identified the global trends of community homogenization and then the associated shifts in community composition and changes in local diversity. We studied these changes through a meta-analysis of distance-based unconstrained ordination plots broadly used to assess individual and case-specific effects of human pressures on community composition. We manually and systematically extracted datapoints from these ordination plots, each of these points representing the composition of an individual biological community (Supplementary Information section 1). This meta-analytical framework, first introduced by ref. 22, allowed us to discriminate between changes of homogeneity and shifts in composition of biological communities in relation to human pressures. By contrast to previous studies mostly restricted to biomonitoring time series, we focused on direct impact studies, considering any of the five most predominant anthropogenic pressures: land-use change, resource exploitation, pollution, climate change and invasive species1,6. For each study included, we compared impacted communities to the reference (control) scenario. Contrary to individual biodiversity time series, this allows direct quantification and comparison of the effects of human pressures23. Of the included studies, 32% are experimental, directly manipulating the human pressure to a reference control, whereas the remaining 68% of the studies do this impact-comparison in pairwise observational approaches.
To study the human impacts on community diversity across space, we collected 3,667 individual comparisons involving 49,401 reference communities and 48,382 impacted communities from 2,133 published studies. This global dataset includes all main groups of organisms (including plants, tetrapods, fish, insects, microbes and fungi) and is representative of the main biomes of the Earth (marine, freshwater and terrestrial). We focused and quantified changes associated with the five dominant human pressures, across several spatial scales from local to global (Fig. 1 and Extended Data Fig. 1). For each comparison, we calculated the log-response ratio—that is, the logarithm-transformed ratio of impacted to reference values—for different components of biodiversity change. First, we evaluated if the different impacted sites are more similar or dissimilar to each other than the reference sites (homogeneity: LRR homogeneity). Then, we looked at the change in species composition between impacted and reference sites (compositional shift: LRR shift). Doing so, we quantified the relative changes of the different dimensions of biodiversity across space in a standardized way. Further, we computed the change in local diversity as the log-response ratio of local diversity (LRR local diversity). We used mixed linear models to estimate the magnitude and significance of these changes and tested the effect of four groups of factors on their variation: biome, human pressure, group of organisms and spatial scale.

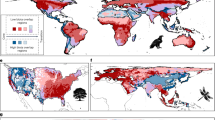
a, Global map of the 3,667 comparisons of diversity included in this study. b–e, Distribution of comparisons of diversity by type of biome (b), human pressures (c), groups of organisms (d) and spatial scale (e). These variables correspond to the four main factors tested in this study.
Contrary to general expectations, we find no evidence of systematic biotic homogenization in response to human pressures (Fig. 2a). The overall log-response ratio for homogeneity is close to zero, yet negative, which suggests biotic differentiation (LRR homogeneity = −0.062, 95% confidence interval (CI) = −0.012 to −0.113). Although the theory of biotic homogenization of communities under human pressure prevailed for a long time24, recent case studies show that biotic differentiation can be regularly observed14. Our exhaustive meta-analysis generalizes this observation, showing that the average impact of human pressures, across all published studies, is biotic differentiation. Critically, however, we find that spatial scale significantly mediates the effects of human pressures on community homogeneity (χ2 = 10.8, P = 0.029), showing from a wide range of contexts that both phenomena (biotic homogenization and differentiation) are widespread. Specifically, human pressures tend to homogenize communities at larger scales (positive LRR homogeneity; Fig. 2a) and differentiate them at smaller scales (negative LRR homogeneity). Large-scale biotic homogenization, when reported, is often linked to the redistribution of species and the facilitation of their dispersal over long distances by humans13,25,26,27,28,29. The fact that local studies show biotic differentiation can be explained by a finer sampling grain and better characterization of communities at small spatial scales, which can make biotic differentiation more apparent14,30. Furthermore, the well-known distance–dissimilarity relationship suggests that communities that are spatially closer are on average more similar31, and thus more prone to differentiation than at larger scales. Finally, stochastic effects and ecological drift can promote biotic differentiation32, and are likely to have more important roles in local impact studies in which strong pressures can completely destabilize communities by drastically reducing the number of individuals. In our systematic analysis, we indeed find a significant biotic differentiation in response to resource exploitation (LRR homogeneity = −0.117, 95% CI = −0.197 to −0.036) and pollution (LRR homogeneity = −0.071, 95% CI = −0.129 to −0.012), two types of human pressure capable of modifying ecosystems in a pronounced way over a short period of time, and thus increasing the importance of ecological drift in community assembly.

a, log-response ratio of community homogeneity (logarithm-transformed ratio of impacted to reference values, LRR homogeneity). b, log-response ratio of community composition shift (LRR shift). The global response (all data) is shown on the first row of each panel and is separated by factors in the following rows. The numbers in parentheses indicate the number of comparisons. For each category the dot represents the marginal mean computed from the model; dot size is proportional to number of studies included. The larger bar shows the 95% confidence interval and the thinner bar represents the 99% confidence interval.
By contrast, and in line with general expectations, we find a clear shift in community composition in response to human pressures (LRR shift = 0.564, 95% CI = 0.467 to 0.661) whose magnitude varies according to the type of biome (χ2 = 12.3, P = 0.002), pressure (χ2 = 42.5, P < 0.001), group of organisms (χ2 = 26.1, P = 0.001) and spatial scale (χ2 = 39.2, P < 0.001) considered (Fig. 2b). Our analysis shows unequivocally that community composition is impacted by human pressures. Such a strong shift can be directly and consistently linked to habitat changes benefiting certain species at the cost of others through environmental filtering and niche processes. Notably, we find that all five types of human pressures (land-use change, resource exploitation, pollution, climate change and invasive species) included in our analysis significantly shift the composition of biological communities (Fig. 2b). These five ubiquitous human pressures are clearly identified in the millennium ecosystem assessment and many studies have shown how they can impact the composition of biological communities since its publication28,33,34,35,36. Our results show that these human pressures systematically change the composition of communities and provide critical insights on the magnitude of effects across human pressures, supporting the notion that all human pressures need to be considered when attempting to bend the curve on biodiversity loss37. We find that habitat change and, above all, pollution have a particularly strong effect on community composition shifts. Yet, we acknowledge that experimental studies of these two human pressures may have compared reference controls to generally relatively high treatment levels (see Extended Data Figs. 2–7 for stratified analyses separating experimental from observational data) and that ranking human pressures can be strongly context- and metric-dependent37. We also show significant differences in composition shifts between groups of organisms. Microbes and fungi, which contain predominantly smaller species, have the highest shifts in the composition of their communities, whereas the effect is less pronounced for mammals, fish, amphibians and reptiles. It has been shown that smaller species, which generally exhibit higher diversity, shorter life cycles and higher dispersal rates relative to larger species, have higher rates of community composition change38,39. Here we provide evidence that this difference among groups is directly reflected in the magnitude of their response to human pressure. As for homogenization, spatial scale has an important role, with shifts in composition becoming increasingly marked as the spatial scale considered is reduced. Again, this result can be explained by better detection of rare species at finer spatial grain. However, directed shifts are also driven by the capacity of a new pool of species to colonize several impacted sites to establish similar communities, which is expected to be more likely at smaller spatial scales.
As changes in community diversity across space can be tightly coupled to changes in local diversity40, we further examined changes in local diversity in relation to human pressures. We extracted 1,139 comparisons of local diversity (taxonomic richness) between reference and impacted communities (Fig. 1a) for a subset of 727 publications and computed for each comparison the log-response ratio for this local diversity (LRR local diversity). Overall, we find clear evidence that sites impacted by human pressures have lower local diversity (LRR local diversity = −0.181, 95% CI = −0.291 to −0.071; Fig. 3). We find that the type of pressures (χ2 = 11.3, P = 0.023) and group of organisms (χ2 = 41.7, P < 0.001) significantly affected local diversity change. Similarly to the results for compositional shifts, pollution and habitat change are the strongest drivers of local diversity loss. Previous syntheses are in line with our finding on the impact of land-use change on local biodiversity16,41,42. However, contrary to community composition shifts, it is the largest organisms that are experiencing the strongest negative effects of human pressures for local diversity. We speculate that contemporary declines reported in vertebrate populations1,43 may be a manifestation of these pressures, given the intrinsic link between population size and risk of local extinction. The trajectory of local diversity in the Anthropocene is the subject of intense and long-standing debate7,8,9,10,12,44. These studies are built on time-series analysis, which generally lack an impact–reference comparison, may be limited by the number of sites and the accuracy of the measurements45, and must be based on adequate null model expectations23. We circumvent these challenges by systematically comparing impacted sites with reference sites (that is, control–impact design). In such designs, the control and impacted sets of sites are assumed to be comparable and any differences between the two treatments is attributed to a change in the impacted group relative to the reference group, which is considered as a stable baseline. Although this approach may be in some cases less sensitive than the gold-standard ‘before–after control–impact’ (BACI) design, which explicitly accounts for pre-existing differences between the impact and reference groups46, it remains by far the most widely used method to measure the real and direct effect of human pressures on biological communities (more than 95% of the studies considered had a control–impact design and less than 5% had a BACI design). With this impact-focused perspective, we quantify and recall the direct and adverse effects of human pressures on local biodiversity.

log-response ratio of local diversity (logarithm-transformed ratio of impacted to reference values, LRR local diversity). The global response (all data) shown on the first row is separated by factors in the following rows. The numbers in parentheses indicate the number of comparisons. For each category the dot represents the marginal mean computed from the model; dot size is proportional to the number of studies included. The larger bar shows the 95% confidence interval and the thinner bar represents the 99% confidence interval.
Finally, our results show a link between changes in local diversity and shifts in composition and homogenization of biological communities across space. Although an interdependency of these different aspects of biodiversity is theoretically predicted40, large-scale integrations are rare as many studies focused on one component only. We report that LRR homogeneity increases (Fig. 4a,b) and LRR shift decreases (Fig. 4c,d) with an increase of LRR local diversity (χ2 = 11.0, P < 0.001 and χ2 = 42.0, P < 0.001, respectively). In other words, a greater loss of species is associated with a stronger shift in composition and more differentiated communities. With a few exceptions, this pattern is highly consistent across biomes, types of pressure, groups of organisms and spatial scales (Fig. 4b,d). Inherent to any comparative study, the observed relationship does not allow us to deduce causality, yet a direct dependency between changes in local diversity, compositional shift and homogenization is not only in line with theoretical predictions, but corroborates the adverse effects of human pressures and their tangible repercussions on the various dimensions of biodiversity.

a, Scatterplot showing the relationship between the log-response ratio (logarithm-transformed ratio of impacted to reference values, LRR) of community local diversity and homogeneity (n = 1,139). The black line shows the relation estimated from the linear mixed model and the grey area its 95% confidence interval. The green lines represent the relation estimated for each biome separately (continuous, marine; dashed, terrestrial; dotted, freshwater). b, Model slopes estimated from subsets of each category. Error bar represents standard error. c, Scatterplot showing the relationship between the log-response ratio of community local diversity and composition shift (n = 1,139). The black line shows the relation estimated from the linear mixed model and the grey area its 95% confidence interval. d, Model slopes estimated from subsets of each category. Error bar represents standard error.
Bending the curve of contemporary biodiversity loss and change is one of the greatest challenges facing our society47,48. Ambitious targets have been proposed to reverse biodiversity change, yet the direction and magnitude of interdependent effects on different levels of biodiversity are still broadly debated. In particular, attributing the change in biodiversity to fundamental drivers has lagged behind21. By systematically assessing how the five main global human pressures impact biodiversity, we quantitatively attribute biodiversity change in impacted versus reference communities, integrating effects on both local diversity change and changes in community composition. Our comprehensive analysis provides a new and highly detailed picture of the state of knowledge available on the signal of human impacts on biodiversity and is thus an important benchmark for the development and assessment of future conservation strategies.
Methods
Search strategy
To detect studies reporting results of homogenization and compositional shift in response to human pressures (systematically comparing impact versus reference scenarios) we conducted an initial bibliographic search on 17 January 2022 on the Science Citation Index Expanded (SCI-EXPANDED) through the Web of Science platform using a combination of 231 relevant keywords (Supplementary Information section 2). The SCI-EXPANDED database was chosen for its functionalities, its generalist aspect and its good coverage of targeted disciplines49,50. This initial search generated 73,632 publications and was refined using a two-step procedure (Supplementary Information section 3). First, we searched the full text of all of these publications to detect all mentions of the principal coordinates analysis (PCoA) and non-metric multidimensional scaling (NMDS) methods, which are the main approaches to study and quantify among-community differentiation and composition shift. NMDS and PCoA explicitly aim to represent and preserve the pairwise dissimilarities between samples in the new reduced-dimensional space. Therefore, they were preferred to other ordination methods such as principal component analysis or (detrended) correspondence analysis whose focus is first on summarizing and maximizing the variance or correspondence structure of the data51. This step reduced the number of possibly relevant publications to 11,968. Second, we manually and individually evaluated the remaining publications to keep only those that met the minimum inclusion criteria for the meta-analysis. To be included, a publication had to feature at least one plot reporting the projection of biological communities in the first two dimensions of a PCoA or NMDS analysis computed from a community dissimilarity matrix, because both these methods strictly require a distance matrix as input, and are therefore preferred for the graphical representation of beta-diversity. The plot had to report at least two groups, the reference (least impacted) and the impacted groups, which could be visually separated by distinct colours or plotting symbols, respectively. Extracting these two groups from the same plot (coordinate system) allowed us to get quantifiable and comparable data on the magnitude and direction of impact effects directly from the original community level data. Moreover, as this type of plot is much more frequently reported than beta-diversity indices (for example, Sørensen index13), this approach makes it possible to include a much larger number and variety of studies. As PCoA and NMDS are directly derived from pairwise beta-diversity matrices, our approach reflects directly the standard beta-diversity indices on which they are built52, but the extracted information is limited to the first two dimensions of the representation22. We excluded restored sites (that is, degraded sites which have been subject to ecological restoration), studies on seed banks and studies with communities reconstructed from stomach content analyses, as they would either not be congruent with the impact framework nor reflect contemporary community change.
The final set of studies included 2,133 publications. A single publication may report one to several (1.72, on average) comparisons between different sets of impacted and reference communities, including comparative and experimental approaches. We extracted several records per publication when results were available for different types of pressure or different taxonomic groups, to systematically include and differentiate several comparisons (for example, effects of different pressures, such as warming and nutrient addition, respectively). We did not create multiple records for cases that were not unambiguously comparable. These cases include: (1) different levels of the same pressure across a gradient (for example, three categories of ‘low’, ‘medium’ and ‘high’ impact for the same pressure). In this case, we extracted the comparison between the two ends of the gradient (for example, low and high), assuming that, in case of experiment/manipulation, authors chose realistic scenarios. (2) Different taxonomic levels (for example, family, genus and species) of the same community. In this case, we took the finest available level (species if possible). (3) Different definitions of the same community (for example, including or excluding non-native species). In this case, we systematically and consistently prioritized the most ‘complete’ communities (that is, the definition including the largest diversity). When a study reported a comparison only for a subset of a community (for example, without non-native species) we included it only if this subset was consistent between the reference and the impacted sites. (4) Different seasons. In this case, we took only one season (with priority order: spring, summer, autumn, winter). (5) Different time points of a time series. In this case, we systematically took the latest point of the time series.
Each extracted comparison was assigned to one of the three main biomes: terrestrial, marine and freshwater. We grouped human pressures into five main categories (habitat change, pollution, resource exploitation, invasive species and climate change). Organisms were grouped into invertebrates, microbes (including bacteria, archaea and microeukaryotes), plants, fungi, fish, birds, reptiles and amphibians, mammals and others (including complex assemblages of several groups and composite organisms such as lichens). Finally, we classified spatial scales into five categories reflecting the order of magnitude of the geographical extent of the reference sites (that is, the distance between the two furthest reference sites, as the best proxy of how spatially aggregated/dispersed these communities are). These non-overlapping categories have following breaking points: point/microhabitat (sites less than 10 m apart), plot (less than 1 km apart), local (less than 100 km apart), regional (less than 1,000 km apart) and continental/global (more than 1,000 km apart).
Data extraction from the plots was achieved using Webplotdigitizer v.4.6 (ref. 53) in manual mode. If plots had no graduated axes for calibration, we assumed the data to be in an orthonormal space, converting the same number of pixels to the same value for both axes. The number of points extracted per graph ranged from 4 to 664 (mean = 26.7, s.d. = 36.5). In addition to point coordinates extracted from projection plots, we extracted mean local diversity (diversity index) in reference and impacted sites and more information about the study and its design. Further technical information on data collection and extraction is provided in Supplementary Information section 4.
Effect sizes
We measured the magnitude of homogenization and compositional shift under human pressure, across all the studies. To do so, we used the community coordinates extracted from the projected plots (PCoA and NMDS) to reconstruct a pairwise Euclidean distance matrix between communities from which we computed two effect sizes, following the approach by ref. 22. First, a measurement of the change in community similarity estimated as the log-transformed ratio of the mean distance in impacted communities \(\bar{{D}_{{\rm{I}}}}\) and the mean distance in reference communities \(\bar{{D}_{{\rm{R}}}}\). For clarity, this value is multiplied by −1 to express a change in homogeneity (LRR homogeneity) instead of heterogeneity, equation (1). Second, a measurement of the shift in community composition (LRR shift) estimated as the log-transformed ratio of the mean distance between impacted and reference communities \(\bar{{D}_{{\rm{B}}}}\) and the mean distance in impacted and reference communities \(\bar{{D}_{{\rm{W}}}}\), equation (2).
Given that these two effect sizes depend on the distances between communities in the analyses and that these distances can be altered by the inclusion of more groups or treatments in the plots, we conducted extra tests based on simulations to ensure the robustness (Supplementary Information section 5). Our results show that LRR homogeneity and LRR shift are generally robust to the inclusion of more groups in the PCoA and NMDS graphs from which they are derived.
For each record, the mean local diversity for the impacted and reference communities was also recorded (if reported as taxonomic richness). We computed a standardized effect size of the change in local diversity (LRR local diversity) by using the log-transformed ratio of the mean local diversity measured in the impacted communities \(\bar{{\alpha }_{{\rm{I}}}}\) and the mean local diversity in the reference communities \(\bar{{\alpha }_{{\rm{R}}}}\), equation (3).
The three effect sizes described here were not statistically tested individually (that is, within-study statistical testing). Statistical modelling and testing were instead conducted across studies, in which variability was accounted for in the interstudy synthesis, as described in the next section.
Statistical analyses
We used mixed models to test the effect of biome, pressure, group of organisms and spatial scale on the variation of LRR homogeneity, LRR shift and LRR local diversity. For each effect size, models included all the moderators together as fixed effects and two random effects (intercepts): publications and type of study (field observation, field manipulation or experimental). In all models, publication was used as a random effect to account for between-study heterogeneity. Within-study heterogeneity was not explicitly included because the number of extracted records per study was only 1.72, on average. The models were not weighted to limit the predominance of a few studies on the average effect sizes. As the sample size of LRR homogeneity and LRR shift is the number of pairwise distances between points in each original study and their relationship is exponential, weighting could strongly distort model results. Given the heterogeneity of our dataset, there is a risk that uncontrolled variables, irrespective of the number of datapoints, could bias results if the weighting scheme were to exacerbate their weight. Therefore, we adopted an unweighted approach which in this case is a more robust and more conservative strategy. However, we show that weighting the models using the square root of the sample size yields qualitatively similar results (Extended Data Figs. 8 and 9). Fixed effects were tested using Type II Wald χ2 tests. All statistical tests reported in this section are two-sided.
Additionally, for each response variable (LRR homogeneity, LRR shift and LRR local diversity), we fitted four independent mixed models including only one moderator at once and the same random effects as described above. From each model we computed estimated marginal means and their 95% and 99% CIs, respectively, for each level of the included factor. Estimated marginal means are an efficient approach to decompose the effect of a specific factor and they were used to produce Figs. 2 and 3. However, comparisons and interpretations of the relative effects of different human pressures must be made with care, as these pressures, which are very different in nature, are exerted on different scales (for example, local chemical pollution versus global warming) and to different degrees that were not measured here.
We used mixed models to test the relationship between LRR local diversity and LRR homogeneity and between LRR local diversity and LRR shift. These two regression models included a random intercept for the original publication. The effects of LRR local diversity on LRR homogeneity and LRR shift were tested using Type II Wald χ2 tests.
We assessed publication bias by inspecting funnel plots for potential asymmetry driven by small studies54. Additionally, we performed a file drawer analysis55,56 and a P-curve analysis57 to further confirm the robustness of our findings against selective publication. The results of these analyses (Supplementary Information section 6) indicate that our findings are robust and not unduly influenced by publication bias.
Additionally, we analysed the spatial layout of reference and impacted sites across a subset of studies (n = 200), specifically investigating differences between within-treatment distances (spatial homogeneity) and between within and between treatment distances (spatial shift). We found limited but significant evidence of spatial patterns in site distribution. However, these patterns did not correlate with the changes observed in community composition between reference and impacted sites (Supplementary Information section 7).
Statistical analyses were conducted with R v.4.0.3 (ref. 58). Mixed models were fitted using the glmmTMB package59 and marginal means estimated using the emmeans package60.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Articles were searched using the SCI-EXPANDED database accessed through the Web of Science platform (https://www.webofscience.com). Extracted data can be obtained from the GitHub project repository (https://github.com/fkeck/metabeta) and available at Zenodo (https://doi.org/10.5281/zenodo.14608770)61.
Code availability
Code to reproduce the analyses and graphical outputs are available on the GitHub project repository (https://github.com/fkeck/metabeta) and available at Zenodo (https://doi.org/10.5281/zenodo.14608770)61.
References
Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES, 2019).
Díaz, S. et al. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 366, eaax3100 (2019).
Primack, R. B. et al. Biodiversity gains? The debate on changes in local- vs global-scale species richness. Biol. Conserv. 219, A1–A3 (2018).
Dornelas, M. et al. Looking back on biodiversity change: lessons for the road ahead. Philos. Trans. R. Soc. B 378, 20220199 (2023).
Blowes, S. A. et al. Synthesis reveals approximately balanced biotic differentiation and homogenization. Sci. Adv. 10, eadj9395 (2024).
Mazor, T. et al. Global mismatch of policy and research on drivers of biodiversity loss. Nat. Ecol. Evol. 2, 1071–1074 (2018).
Vellend, M. et al. Global meta-analysis reveals no net change in local-scale plant biodiversity over time. Proc. Natl Acad. Sci. USA 110, 19456–19459 (2013).
Dornelas, M. et al. Assemblage time series reveal biodiversity change but not systematic loss. Science 344, 296–299 (2014).
Gonzalez, A. et al. Estimating local biodiversity change: a critique of papers claiming no net loss of local diversity. Ecology 97, 1949–1960 (2016).
Cardinale, B. J., Gonzalez, A., Allington, G. R. H. & Loreau, M. Is local biodiversity declining or not? A summary of the debate over analysis of species richness time trends. Biol. Conserv. 219, 175–183 (2018).
Leung, B. et al. Clustered versus catastrophic global vertebrate declines. Nature 588, 267–271 (2020).
Loreau, M. et al. Do not downplay biodiversity loss. Nature 601, E27–E28 (2022).
Petsch, D. K., dos Santos Bertoncin, A. P., Ortega, J. C. G. & Thomaz, S. M. Non-native species drive biotic homogenization, but it depends on the realm, beta diversity facet and study design: a meta-analytic systematic review. Oikos https://doi.org/10.1111/oik.08768 (2022).
Rolls, R. J. et al. Biotic homogenisation and differentiation as directional change in beta diversity: synthesising driver–response relationships to develop conceptual models across ecosystems. Biol. Rev. 98, 1388–1423 (2023).
Alkemade, R. et al. GLOBIO3: a framework to investigate options for reducing global terrestrial biodiversity loss. Ecosystems 12, 374–390 (2009).
Newbold, T. et al. Global effects of land use on local terrestrial biodiversity. Nature 520, 45–50 (2015).
Chaudhary, A., Verones, F., de Baan, L. & Hellweg, S. Quantifying land use impacts on biodiversity: combining species–area models and vulnerability indicators. Environ. Sci. Technol. 49, 9987–9995 (2015).
Schipper, A. M. et al. Projecting terrestrial biodiversity intactness with GLOBIO 4. Glob. Change Biol. 26, 760–771 (2020).
Staude, I. R. et al. Directional turnover towards larger-ranged plants over time and across habitats. Ecol. Lett. 25, 466–482 (2022).
Santangeli, A. et al. Mixed effects of a national protected area network on terrestrial and freshwater biodiversity. Nat. Commun. 14, 5426 (2023).
Gonzalez, A., Chase, J. M. & O’Connor, M. I. A framework for the detection and attribution of biodiversity change. Philos. Trans. R. Soc. B 378, 20220182 (2023).
Zhou, Z., Wang, C. & Luo, Y. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 11, 3072 (2020).
Kuczynski, L., Ontiveros, V. J. & Hillebrand, H. Biodiversity time series are biased towards increasing species richness in changing environments. Nat. Ecol. Evol. 7, 994–1001 (2023).
McKinney, M. L. & Lockwood, J. L. Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 14, 450–453 (1999).
Carvallo, G. O. & Castro, S. A. Invasions but not extinctions change phylogenetic diversity of angiosperm assemblage on southeastern Pacific Oceanic islands. PLoS ONE 12, e0182105 (2017).
Longman, E. K., Rosenblad, K. & Sax, D. F. Extreme homogenization: the past, present and future of mammal assemblages on islands. Glob. Ecol. Biogeogr. 27, 77–95 (2018).
Su, G. et al. Human impacts on global freshwater fish biodiversity. Science 371, 835–838 (2021).
Daru, B. H. et al. Widespread homogenization of plant communities in the Anthropocene. Nat. Commun. 12, 6983 (2021).
Capinha, C., Essl, F., Seebens, H., Moser, D. & Pereira, H. M. The dispersal of alien species redefines biogeography in the Anthropocene. Science 348, 1248–1251 (2015).
Chase, J. M. & Knight, T. M. Scale-dependent effect sizes of ecological drivers on biodiversity: why standardised sampling is not enough. Ecol. Lett. 16, 17–26 (2013).
Soininen, J., McDonald, R. & Hillebrand, H. The distance decay of similarity in ecological communities. Ecography 30, 3–12 (2007).
Zhou, Z., Zheng, M., Xia, J. & Wang, C. Nitrogen addition promotes soil microbial beta diversity and the stochastic assembly. Sci. Total Environ. 806, 150569 (2022).
Antão, L. H. et al. Climate change reshuffles northern species within their niches. Nat. Clim. Change 12, 587–592 (2022).
Marta, S., Brunetti, M., Manenti, R., Provenzale, A. & Ficetola, G. F. Climate and land-use changes drive biodiversity turnover in arthropod assemblages over 150 years. Nat. Ecol. Evol. 5, 1291–1300 (2021).
Rumschlag, S. L. et al. Consistent effects of pesticides on community structure and ecosystem function in freshwater systems. Nat. Commun. 11, 6333 (2020).
Magurran, A. E. et al. Divergent biodiversity change within ecosystems. Proc. Natl Acad. Sci. USA 115, 1843–1847 (2018).
Bellard, C., Marino, C. & Courchamp, F. Ranking threats to biodiversity and why it doesn’t matter. Nat. Commun. 13, 2616 (2022).
Schoener, T. W. Rate of species turnover decreases from lower to higher organisms: a review of the data. Oikos 41, 372–377 (1983).
Korhonen, J. J., Soininen, J. & Hillebrand, H. A quantitative analysis of temporal turnover in aquatic species assemblages across ecosystems. Ecology 91, 508–517 (2010).
Mouquet, N. & Loreau, M. Community patterns in source–sink metacommunities. Am. Nat. 162, 544–557 (2003).
Millard, J. et al. Global effects of land-use intensity on local pollinator biodiversity. Nat. Commun. 12, 2902 (2021).
Semenchuk, P. et al. Relative effects of land conversion and land-use intensity on terrestrial vertebrate diversity. Nat. Commun. 13, 615 (2022).
Living Planet Report 2022—Building a Nature-Positive Society (WWF, 2022).
Blowes, S. A. et al. The geography of biodiversity change in marine and terrestrial assemblages. Science 366, 339–345 (2019).
Valdez, J. W. et al. The undetectability of global biodiversity trends using local species richness. Ecography 2023, e06604 (2023).
Christie, A. P. et al. Quantifying and addressing the prevalence and bias of study designs in the environmental and social sciences. Nat. Commun. 11, 6377 (2020).
Mace, G. M. et al. Aiming higher to bend the curve of biodiversity loss. Nat. Sustain. 1, 448–451 (2018).
Leclère, D. et al. Bending the curve of terrestrial biodiversity needs an integrated strategy. Nature 585, 551–556 (2020).
Gusenbauer, M. & Haddaway, N. R. Which academic search systems are suitable for systematic reviews or meta-analyses? Evaluating retrieval qualities of Google Scholar, PubMed, and 26 other resources. Res. Synth. Methods 11, 181–217 (2020).
Foo, Y. Z., O’Dea, R. E., Koricheva, J., Nakagawa, S. & Lagisz, M. A practical guide to question formation, systematic searching and study screening for literature reviews in ecology and evolution. Methods Ecol. Evol. 12, 1705–1720 (2021).
Legendre, P. & Legendre, L. F. J. Numerical Ecology (Elsevier, 2012).
Anderson, M. J. et al. Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist. Ecol. Lett. 14, 19–28 (2011).
Rohatgi, A. Webplotdigitizer v.4.6 (2022).
Møller, A. P. & Jennions, M. D. Testing and adjusting for publication bias. Trends Ecol. Evol. 16, 580–586 (2001).
Orwin, R. G. A Fail-SafeN for effect size in meta-analysis. J. Educ. Stat. 8, 157–159 (1983).
Rosenberg, M. S. The file‐drawer problem revisited: a general weighted method for calculating fail‐safe numbers in meta‐analysis. Evolution 59, 464–468 (2005).
Simonsohn, U., Simmons, J. P. & Nelson, L. D. Better P-curves: making P-curve analysis more robust to errors, fraud, and ambitious P-hacking, a reply to Ulrich and Miller. J. Exp. Psychol. Gen. 144, 1146–1152 (2015).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2022).
Brooks, M. E. et al. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400 (2017).
Lenth, R. V. Emmeans: estimated marginal means, aka least-squares means. R package version 1.10.0 (2021).
Keck F. et al. fkeck/metabeta: v1.0 (v1.0). Zenodo https://doi.org/10.5281/zenodo.14608770 (2025).
Hedges, L. V., Gurevitch, J. & Curtis, P. S. The meta-analysis of response ratios in experimental ecology. Ecology 80, 1150–1156 (1999).
Acknowledgements
We thank H. Broadbent for his assistance in digitizing sampling maps. This study was supported by the FAN (Research Talent Development Fund of the UZH Alumni), the Ernst Göhner Foundation, the Gebauer Foundation and the Swiss National Science Foundation (grant no. 310030_197410). L.F. was supported by a stipend (Excellence Stipend for Doctoral Studies) from the Gesellschaft für Forschungsförderung Niederösterreich.
Funding
Open access funding provided by University of Zurich.
Author information
Authors and Affiliations
Contributions
F.K. designed the study, ran the initial search, performed the initial screening, conducted the analyses and led the writing of the paper. T.P. and F.K. performed the pilot test for minimum inclusion criteria. T.P. and F.A. had a central role in the writing of the paper. F.K., T.P., R.A., C.B., R.B., E.C., T.C., M.C., L.F., D.K., M.K., L.M., R.O., K.T., H.Z. and F.A. participated equally in data extraction and discussed and commented on the manuscript. Apart from F.K., T.P. and F.A., authors are listed in alphabetical order.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Henrique Pereira, Andy Purvis, William Kunin and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Distribution of comparisons of diversity across the four moderators.
a–c, Biomes. d–f, Groups of organisms. g–i, Human pressures. j–l, Spatial scale. For each moderator, the distribution is represented conditionally to the other moderators.
Extended Data Fig. 2 Location of diversity comparisons from observational studies and their distribution across biomes, pressures, organisms and scale.
a, Global map of the 2,501 comparisons of diversity included. b–e, Distribution of comparisons of diversity by type of biome, human pressures, groups of organisms and spatial scale. These variables correspond to the four main factors tested in this study.
Extended Data Fig. 3 Impacts of human pressures on homogeneity and shifts in composition of biological communities estimated from observational studies.
a, ratio of community homogeneity (logarithm-transformed ratio of impacted to reference values, LRR homogeneity). b, Log-response ratio of community composition shift (LRR shift). The global response (all data) is shown on the first row of each panel and is separated by factors in the following rows. The numbers between parentheses indicate the number of comparisons. For each category the dot represents the marginal mean computed from the model; dot size is proportional to number of studies included. The larger bar shows the 95% confidence interval and the thinner bar represents the 99% confidence interval.
Extended Data Fig. 4 Impacts of human pressures on local diversity estimated from observational studies.
Log-response ratio of local diversity (logarithm-transformed ratio of impacted to reference values, LRR local diversity). The global response (all data) shown on the first row is separated by factors in the following rows. The numbers between parentheses indicate the number of comparisons. For each category the dot represents the marginal mean computed from the model; dot size is proportional to number of studies included. The larger bar shows the 95% confidence interval and the thinner bar represents the 99% confidence interval.
Extended Data Fig. 5 Location of diversity comparisons from experimental studies and their distribution across biomes, pressures, organisms and scale.
a, Global map of the 1,166 comparisons of diversity included. b–e, Distribution of comparisons of diversity by type of biome, human pressures, groups of organisms and spatial scale. These variables correspond to the four main factors tested in this study.
Extended Data Fig. 6 Impacts of human pressures on homogeneity and shifts in composition of biological communities estimated from experimental studies.
a, Log-response ratio of community homogeneity (logarithm-transformed ratio of impacted to reference values, LRR homogeneity). b, Log-response ratio of community composition shift (LRR shift). The global response (all data) is shown on the first row of each panel and is separated by factors in the following rows. The numbers between parentheses indicate the number of comparisons. For each category the dot represents the marginal mean computed from the model; dot size is proportional to number of studies included. The larger bar shows the 95% confidence interval and the thinner bar represents the 99% confidence interval.
Extended Data Fig. 7 Impacts of human pressures on local diversity estimated from experimental studies.
Log-response ratio of local diversity (logarithm-transformed ratio of impacted to reference values, LRR local diversity). The global response (all data) shown on the first row is separated by factors in the following rows. The numbers between parentheses indicate the number of comparisons. For each category the dot represents the marginal mean computed from the model; dot size is proportional to number of studies included. The larger bar shows the 95% confidence interval and the thinner bar represents the 99% confidence interval.
Extended Data Fig. 8 Impacts of human pressures on homogeneity and shifts in composition of biological communities estimated using weighted regressions.
a, Log-response ratio of community homogeneity (logarithm-transformed ratio of impacted to reference values, LRR homogeneity). b, Log-response ratio of community composition shift (LRR shift). The global response (all data) is shown on the first row of each panel and is separated by factors in the following rows. The numbers between parentheses indicate the number of comparisons. For each category the dot represents the marginal mean computed from the model; dot size is proportional to number of studies included. The larger bar shows the 95% confidence interval and the thinner bar represents the 99% confidence interval. The weighting scheme applied is the inverse of the square root of the variance of the effect sizes, computed following Zhou et al.22.
Extended Data Fig. 9 Impacts of human pressures on local diversity estimated from experimental studies using weighted regression.
Log-response ratio of local diversity (logarithm-transformed ratio of impacted to reference values, LRR local diversity). The global response (all data) shown on the first row is separated by factors in the following rows. The numbers between parentheses indicate the number of comparisons. For each category the dot represents the marginal mean computed from the model; dot size is proportional to number of studies included. The larger bar shows the 95% confidence interval and the thinner bar represents the 99% confidence interval. The weighting scheme applied is the inverse of the variance of the effect size, computed following Hedges et al.62.
Supplementary information
Supplementary Information
Supplementary Figs. 1–7, Table 1 and references.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Keck, F., Peller, T., Alther, R. et al. The global human impact on biodiversity. Nature 641, 395–400 (2025). https://doi.org/10.1038/s41586-025-08752-2
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41586-025-08752-2
This article is cited by
-
Global habitat hotspots and extinction vulnerability of terrestrial vertebrates
Nature Sustainability (2026)
-
A review of urban energy resilience assessment under extreme climate conditions
International Journal of Environmental Science and Technology (2026)
-
Natural capital accounting for sustainable cities
Sustainability Science (2026)
-
What evidence exists on the impacts of human activities on biodiversity and carbon capacity in North-East Atlantic blue carbon ecosystems: a systematic map protocol
Environmental Evidence (2025)
-
Grassland degradation alters plant and soil biodiversity–multifunctionality relationships
Nature Plants (2025)
