
Abstract
Glioblastoma is the most common and aggressive primary brain cancer and shows minimal response to therapies. The immunosuppressive tumour microenvironment in glioblastoma contributes to the limited therapeutic response. Astrocytes are abundant in the central nervous system and have important immunoregulatory roles. However, little is known about their role in the immune response to glioblastoma1. Here we used single-cell and bulk RNA sequencing of clinical glioblastoma samples and samples from preclinical models, multiplexed immunofluorescence, in vivo CRISPR-based cell-specific genetic perturbations and in vitro mouse and human experimental systems to address this gap in knowledge. We identified an astrocyte subset that limits tumour immunity by inducing T cell apoptosis through the death receptor ligand TRAIL. Moreover, we identified that IL-11 produced by tumour cells is a driver of STAT3-dependent TRAIL expression in astrocytes. Astrocyte signalling through STAT3 and TRAIL expression were associated with a shorter time to recurrence and overall decreased survival in patients with glioblastoma. Genetic inactivation of the IL-11 receptor or TRAIL in astrocytes extended survival in mouse models of glioblastoma and enhanced T cell and macrophage responses. Finally, treatment with an oncolytic HSV-1 virus engineered to express a TRAIL-blocking single-chain antibody in the tumour microenvironment extended survival and enhanced tumour-specific immunity in preclinical models of glioblastoma. In summary, we establish that IL-11–STAT3-driven astrocytes suppress glioblastoma-specific protective immunity by inducing TRAIL-dependent T cell apoptosis, and engineered therapeutic viruses can be used to target this mechanism of astrocyte-driven tumour immunoevasion.
Similar content being viewed by others


Main
Glioblastoma is the most common and aggressive of brain tumours2. Little progress has been made over decades in the development of efficacious glioblastoma therapies. Consequently, the 5-year survival rate remains below 10% after surgical resection, radiotherapeutic and chemotherapeutic regimens2. Moreover, immune checkpoint blockade, which has seen success in the treatment of other malignancies, has not shown clinical benefit in glioblastoma3. This limited progress in the development of glioblastoma therapies reflects multiple challenges, including tumour cell and patient heterogeneity and the strong immunosuppressive nature of the tumour microenvironment (TME)1,4,5.
Multiple cell types participate in the regulation of the immune response in the central nervous system (CNS). For example, tumour-associated macrophage (TAM)6,7,8,9 and neutrophil10 subsets have been linked to the control of the immune response to glioblastoma. Astrocytes are the most abundant glial cells in the CNS and have established roles in development and homeostasis11. In the context of brain tumours, astrocyte-mediated responses in brain tumour growth have also been reported12,13,14,15,16,17,18,19. However, despite their importance in immune regulation in the CNS20, little is known about the role of astrocytes in the control of the immune response to glioblastoma21.
Here we describe an astrocyte subset in the glioblastoma TME that induces T cell apoptosis through tumour necrosis factor-related apoptosis-inducing ligand (TRAIL; encoded by TNFSF10) and impairs antitumour immunity. We also demonstrate that TRAIL+ astrocytes are induced in a STAT3-dependent manner by glioblastoma-secreted IL-11. We develop an oncolytic virus engineered to block TRAIL in the glioblastoma TME, which improved survival and enhanced tumour-specific T cell immunity. In summary, we identify a previously unknown role of astrocytes in the suppression of tumour-specific immunity and provide tools for their therapeutic modulation in glioblastoma.
TRAIL+ astrocytes are expanded in glioblastoma
To study glioblastoma-associated astrocytes, we first analysed a single-cell RNA-seq (scRNA-seq) dataset of glioblastoma samples and healthy tissue (control) samples22. In the glioblastoma samples, we detected a reduction in astrocyte clusters involved in homeostasis and a concomitant expansion of a subset of astrocytes (cluster 2) enriched for cell–cell interaction mechanisms with potential immunoregulatory roles (for example, through C3 (ref. 23), TGFβ12 or MIF18) (Fig. 1a–c and Extended Data Fig. 1a–f). Astrocytes from patients with accelerated glioblastoma recurrence (within 9 months after surgery) were enriched for pro-apoptotic signalling through TRAIL and displayed increased TRAIL expression levels (Fig. 1d,e, Extended Data Fig. 1g and Supplementary Data 1). Notably, glioblastoma-associated astrocytes in cluster 2 displayed the highest TRAIL expression levels (Fig. 1f, Extended Data Fig. 1h and Supplementary Data 2). Moreover, high TRAIL expression and extrinsic apoptosis signalling through death receptors were associated with shorter survival in The Cancer Genome Atlas (TCGA) dataset (Fig. 1g,h).

a–c, Uniform manifold approximation and projection (UMAP) plots of human astrocytes analysed by scRNA-seq in control and glioblastoma (GB) samples (n = 267 cells)22 (a), and depicting sample condition (b) and cluster proportions per condition, with the fold change in glioblastoma indicated in parentheses (c). d, Gene set enrichment analysis (GSEA) comparing astrocytes in early recurrence (≤9 months) and late recurrence (>9 months). FDR, false discovery rate; NES, normalized enrichment score. e,f, TNFSF10 expression in glioblastoma astrocytes by time to recurrence (e) and by cluster (f). g,h, Kaplan–Meier survival curve of glioblastoma in the TCGA dataset stratified by high or low TNFSF10 expression (top versus bottom quintile) (g) or by the expression of pathway components of the gene ontology term ‘extrinsic apoptotic signalling pathway via death domain receptors’ (GO:0008625) (top versus bottom quintile) (h). i, Schematic of snRNA-seq validation studies from frozen glioblastoma or control surgical specimens. j,k, UMAP of snRNA-seq of subclustered astrocytes in glioblastoma (n = 3,477 cells) (j) and cluster proportions of astrocytes per condition, with the fold change in glioblastoma indicated in parentheses (k). l, Transcriptional signature of cluster 2 (C2) glioblastoma-associated astrocytes shown in a in astrocyte clusters identified in snRNA-seq validation studies. m, Left, schematic of multiplexed iterative immunofluorescence (IF) microscopy protocol and automated analysis of normal brain (n = 5) and glioblastoma (n = 3) specimens. Middle, immunohistochemistry (IHC) image of EGFRvIII and haematoxylin and eosin counterstaining (top; scale bar, 100 µm) and close-up fluorescence images (bottom) with 4′,6-diamidin-2-phenylindol (DAPI), GFAP and TRAIL channels in representative control tissue (left; scale bar, 50 µm), glioblastoma peritumoural tissue (centre; scale bar, 50 µm) and tumoural tissue (right; 25 µm). Right, quantification of TRAIL+ astrocytes in peritumoural tissue and in malignant cells in tumour tissue. Data are the mean ± s.e.m. Unpaired two-tailed t-test used for statistical analysis. Paired two-tailed t-test in m. n indicates biologically independent samples. Schematics in d and i were created in BioRender. Lee, J. (2025) (https://BioRender.com/pn72nh0).
To validate these findings, we performed single-nucleus RNA-seq (snRNA-seq) on an independent collection of glioblastoma samples and control samples (Supplementary Table 1). Using copy-number analysis to distinguish non-malignant astrocytes from tumour cells, we identified four astrocyte populations (Fig. 1i,j and Extended Data Fig. 2a–i). Cluster 1 astrocytes were expanded in glioblastoma samples, and their transcriptional signature was the most similar to that of cluster 2 astrocytes identified by scRNA-seq in Fig. 1a–c (Fig. 1k,l). We also analysed human control and glioblastoma tissue by multiplexed iterative immunofluorescence microscopy, using EGFRvIII as a tumour cell marker (Fig. 1m). We detected TRAIL+ astrocytes in peritumoural areas of glioblastoma samples, whereas control brain tissue showed little TRAIL expression (Fig. 1m). A low proportion of glioblastoma cells also expressed TRAIL (Extended Data Fig. 2j).
We next investigated TRAIL-expressing astrocytes in the orthotopic mouse glioma GL261 model9. To overcome the technical limitations in transcriptionally distinguishing non-malignant astrocytes from glioma cells, we used Aldh1l1cre-ERT2/TdTomato immunocompetent mice as hosts to label putative astrocytes with TdTomato (Fig. 2a and Extended Data Fig. 3a). We identified five TdTomato+ reporter cell clusters by scRNA-seq (Fig. 2b,c and Extended Data Fig. 3b–h), and cluster 1 reporter cells were enriched in the GL261-implanted hemisphere (Fig. 2d and Extended Data Fig. 3i,j). Notably, cluster 1 cells expressed Tnfsf10 (Fig. 2e). Cluster 1 cells also displayed increased expression of cytokine, complement and hypoxia signalling pathways, but decreased calcium signalling and synaptic support functions (Extended Data Fig. 3k). As the capture of Tnfsf10 transcripts in scRNA-seq was sparse, we calculated a bioinformatic imputation score based on the transcriptional regulation of TRAIL and found that cluster 1 cells were most poised for TRAIL expression (Fig. 2f). Indeed, in flow cytometry validation studies, we detected increased TRAIL surface expression in TdTomato+ cells from GL261-bearing mice compared with sham-injected controls (Fig. 2g,h and Extended Data Fig. 4a–c). Note that this increase was independent of the response induced by CNS injury during tumour implantation (Extended Data Fig. 4d). We also detected an increased frequency in TRAIL+ reporter cells in a genetically defined model of glioma induced through the postnatal electroporation of CRISPR guide RNAs targeting Trp53, Pten and Nf1 (termed MK007) (Fig. 2h). In summary, TRAIL+ astrocytes are increased in the TME of human glioblastoma and in experimental mouse models.

a, Schematic of the GL261 glioma model in Aldh1l1cre-ERT2/TdTomato reporter mice used for scRNA-seq studies. b, UMAP of mouse TdTomato+ reporter cells analysed by scRNA-seq (n = 11,701 cells). c, Percentage of each hemisphere sample occupied by cluster 1 cells grouped by mouse. d, Differentially abundant cell neighbourhoods by condition calculated using miloR. Positive log2[fold change] shows enrichment in GL261-bearing hemispheres compared with controls. e, Feature plot of Tnfsf10 expression in reporter cells from GL261 tumours by scRNA-seq. f, TRAIL imputation score calculated using the CollecTri database of transcriptional regulation. Two-sided Mann–Whitney U-test with Bonferroni correction used for statistical analysis. g,h, Flow cytometry analyses of TdTomato+ reporter cells 15 days after implantation of GL261 tumours (g; n = 8 for sham, n = 9 for GL261) or 19 days after implantation of MK007 tumours (h; n = 6 each for both sham and MK007) in Aldh1l1cre-ERT2/TdTomato reporter mice. Sham surgery was performed as a control. The percentage of TRAIL+ reporter cells is shown. Data shown as the mean ± s.e.m. Unpaired two-tailed t-test used for statistical analysis. NS, not significant. n indicates biologically independent samples. Schematic in a was created in BioRender. Lee, J. (2025) (https://BioRender.com/pn72nh0).
TRAIL+ astrocytes limit T cell responses
TRAIL interacts with death receptor 4 (DR4) and DR5 to induce T cell apoptosis24. Previously, we reported that TRAIL+ astrocytes induce T cell apoptosis through DR5 in experimental autoimmune encephalomyelitis and multiple sclerosis25. Thus, we studied the role of TRAIL+ astrocytes in the control of T cell responses to glioblastoma. In multiplex immunostaining studies, we detected T cells positive for the apoptosis marker cleaved caspase-3 in the TME of human glioblastoma and mouse GL261 glioma samples (Fig. 3a,b). Notably, cleaved caspase-3+CD3+ T cells were localized closer to TRAIL+ astrocytes than cleaved caspase-3− T cells (Fig. 3c,d). In situ transcriptomic analyses highlighted the spatial correlation of TRAIL expression with reactive astrocyte programs (Extended Data Fig. 5a). Death receptor signalling programs were predominantly expressed at the glioblastoma leading edge, where astrocytes are located and where the correlation of DR5 and TRAIL expression was highest (Extended Data Fig. 5b). Collectively, these findings indicate that TRAIL+ astrocytes interact with DR5+ glioblastoma-infiltrating T cells.

a, Immunofluorescence images of apoptotic T cells in glioblastoma (n = 3). Dotted circles indicate CD3+ T cells. b, Immunofluorescence detection of TRAIL+ astrocytes and apoptotic T cells in GL261 tumours at day 15 (n = 3 mice). Arrow points to cleaved caspase-3+ apoptotic T cell. c,d, Distribution (c) and boxplots with median, interquartile range (IQR) ± 1.5 IQR, and minimal and maximal values (d) of distances between cleaved caspase-3+ and cleaved caspase-3− T cells to the nearest TRAIL+GFAP+ peritumoural astrocyte within 500 µm in glioblastoma (n = 3 samples). Two-sided, Bonferroni-corrected Mann–Whitney U-test. e, Schematic of lentiviral (LV) Gfap-driven Cas9 transduction for in vivo astrocyte-specific perturbation. f, Mouse survival in GL261 model after TRAIL inactivation in astrocytes. PBS injection and non-targeting (NT) sgRNA were used as controls (n = 20 sham, n = 29 NT sgRNA, n = 28 sgTnfsf10). Weight fold change from study initiation in surviving mice: sgTnfsf10 1.06 ± 0.034 g, sham 1.112 ± 0.026 g, NT sgRNA 1.064 ± 0.072 g (sgTnfsf10 versus sham, P = 0.9275; sgTnfsf10 versus NT sgRNA, P = 0.6543 by two-sided analysis of variance (ANOVA)). Data from three independent experiments. log-rank (Mantel–Cox) test. g, Cleaved caspase-3/7+Sytox− CD4+ and CD8+ T cells by flow cytometry in GL261 tumours at day 15 after TRAIL inactivation in astrocytes (n = 7 NT sgRNA, n = 9 sgTnfsf10). h, Quantification of GARC-1(77–85)-specific CD8+ T cells by flow cytometry at 15 days. i, Activation and exhaustion-stage analysis by TIM3 and SLAMF6 analysis in PD1+CD8+ T cells. j–m, GSEA analysis (P < 0.05) in RNA-seq of CD8+ (j) and CD4+ (k) T cells and of monocyte-derived (l) and microglia-derived (m) TAMs. Positive NES values indicate pathway enrichment in sgTnfsf10 versus NT sgRNA. n, Cleaved caspase-3/7 and caspase-8 levels by flow cytometry in CD4+ and CD8+ T cells co-cultured with astrocytes pretreated with GL261 TCM. Data from two independent experiments. o, CD4+ and CD8+ T cell apoptosis in T cells cultured with astrocytes pretreated with TCM and subjected to TRAIL blockade. p, Proximity ligation assay (PLA) for TRAIL and DR5 in co-cultures of T cells with astrocytes pretreated with TCM. Data shown as the mean ± s.e.m. Comparison by unpaired two-tailed t-test. n indicates biologically independent samples. Scale bars, 15 µm (a, middle and right), 25 µm (p), 30 µm (a, left), 50 µm (b). Schematic in n created in BioRender. Lee, J. (2025) (https://BioRender.com/pn72nh0).
To further investigate pro-apoptotic interactions between astrocytes and tumour-infiltrating T cells, we specifically inactivated Tnfsf10 in astrocytes in vivo using lentivirus-driven Cas9 and a Tnfsf10-targeting single guide (sgRNA) expressed under the control of the ABC1D gfa2 Gfap promoter26 (Fig. 3e and Extended Data Fig. 5c–e); a lentivirus encoding Cas9 and non-targeting sgRNA was used to control for nonspecific inflammation caused by the lentiviral vector. Astrocyte-specific TRAIL inactivation increased survival after orthotopic GL261 implantation (Fig. 3f), whereas deletion of TRAIL in GL261 cells had no effect on survival (Extended Data Fig. 5f). We validated these findings in the CRISPR-induced glioma model MK007, in which astrocyte-specific TRAIL inactivation led to increased survival (Extended Data Fig. 5g). Furthermore, analyses of T cell-deficient Rag2−/− mice established that the survival benefits resulting from abrogated TRAIL expression in astrocytes depended on T cells (Extended Data Fig. 5h). Astrocyte-specific TRAIL inactivation decreased CD4+ and CD8+ T cell apoptosis in the TME (Fig. 3g, and Extended Data Fig. 5i,j), and led to increased numbers of CD8+ T cells reactive to the tumour-associated antigen GARC-1(77–85), as determined using peptide-loaded major histocompatibility complex class I (MHCI) tetramers (Fig. 3h). Moreover, TRAIL inactivation increased the abundance of activated PD-1+SLAMF6−TIM3− CD8+ T cells (Fig. 3i and Extended Data Fig. 6a,b) and reduced the number of CD8+ T cells expressing TIGIT and CTLA4, which have been linked to T cell dysfunction27 (Extended Data Fig. 6c). Similarly, the expression of the regulator of terminal T cell exhaustion Tox28 was decreased after TRAIL inactivation (Extended Data Fig. 6d). Indeed, TRAIL inactivation increased IFNγ+ and TNF+CD8+ T cells, whereas CD4+ T cells showed increased IFNγ and reduced IL-10 production (Extended Data Fig. 6e,f). In support of a role for TRAIL in limiting tumour-specific T cell immunity, TRAIL inactivation in astrocytes induced transcriptional responses linked to glycolysis in CD8+ T cells and calcium signalling in CD4+ T cells, as detected by RNA-seq (Fig. 3j,k, and Supplementary Data 3 and 4), results indicative of increased T cell activation29. In in vitro studies, TRAIL also suppressed T cell receptor signalling, as indicated by the reduced nuclear translocation of NFAT (Extended Data Fig. 6g), a result in agreement with previously described non-apoptotic functions of TRAIL in early T cell activation30.
TRAIL inactivation in astrocytes also modulated microglia-derived and monocyte-derived TAM responses, which led to the upregulation of antigen presentation and co-stimulatory pathways, IFNγ and IL-12 signalling. Together, these results suggest that TRAIL inactivation improves the ability to promote T cell immunity (Fig. 3l,m and Supplementary Data 5 and 6). TAMs derived from microglia and CNS-recruited peripheral cells also expressed DR5, but at lower levels than T cells, and TRAIL inactivation in astrocytes did not affect TAM apoptosis (Extended Data Fig. 6h,i). However, TAM responses were modulated by TRAIL inactivation in astrocytes in a T cell-dependent manner (Extended Data Fig. 6j,k).
Finally, in the dendritic cell (DC) compartment, cDC2 and monocyte-derived DC subsets expressed DR5, whereas cDC1 subsets that are crucial for mounting CD8 responses expressed little DR5 (Extended Data Fig. 6l).
To investigate the mechanisms involved in the regulation of tumour-specific immunity by TRAIL+ astrocytes, particularly their ability to induce T cell apoptosis, we co-cultured in vitro-activated mouse CD4+ or CD8+ T cells with astrocytes pretreated with GL261 tumour-conditioned medium (TCM) or control medium (Extended Data Fig. 6m). TCM-pretreated astrocytes increased the activation of caspase-3 and caspase-7 (caspase-3/7) and caspase-8 in CD4+ and CD8+ T cells in a TRAIL-dependent manner, a result indicative of death-receptor-driven apoptosis (Fig. 3n,o). Analyses of astrocyte–T cell co-cultures using proximity ligation assays confirmed the presence of interactions between TRAIL expressed on TCM-pretreated astrocytes and DR5 expressed on T cells (Fig. 3p). These findings indicate that glioblastoma-secreted factors drive TRAIL+ astrocytes, which limit tumour-specific immunity by inducing extrinsic apoptosis in T cells.
STAT3 induces TRAIL expression in astrocytes
To study the regulation of TRAIL expression in astrocytes by glioblastoma-secreted factors, we first used an in vitro system. Treatment of primary mouse astrocytes with GL261 TCM induced Tnfsf10 transcription and increased TRAIL surface expression (Fig. 4a,b). Similarly, TCM from human glioblastoma U87 cells or primary glioblastoma cultures increased TNFSF10 expression in human primary astrocytes and transactivated the TNFSF10 promoter in luciferase reporter assays (Fig. 4c,d and Extended Data Fig. 7a).

a, Tnfsf10 expression in GL261 TCM-treated primary mouse astrocyte cultures (n = 12 control, n = 8 TCM). b, TRAIL expression in GL261 TCM-treated primary mouse astrocytes by flow cytometry (n = 3 control, n = 5 TCM). c, TNFSF10 expression in U87 TCM-treated primary human fetal astrocytes (n = 3 control, n = 4 TCM). d, Luciferase activity in U87 TCM-treated HepG2 cells expressing a TNFSF10 promoter reporter (n = 3 each). a.u., arbitrary units. e–g, Heatmap (e) and ingenuity pathway analysis (IPA) (f,g) from RNA-seq analysis of TRAIL+ versus TRAIL–TdTomato+ reporter cells sorted from GL261 tumours (n = 4 each). P < 0.05 for all pathways. h,i, STAT3 signalling score (M5897) in glioblastoma scRNA-seq (Fig. 1b) by time to recurrence (h) or by cluster (i). j, STAT3 signature (M9174) in astrocytes from validation glioblastoma snRNA-seq studies (Fig. 1j). k,l, STAT3 signalling score (M5897) in GL261 Aldh1l1cre-ERT2/TdTomato reporter cell scRNA-seq (Fig. 2b). m, Immunostaining of pSTAT3+TRAIL+ in peritumoural astrocytes and GL261 tumours at 15 days (n = 3 each). n, Representative immunofluorescence image of TRAIL+pSTAT3+ peritumoural astrocytes in glioblastoma (n = 3 glioblastoma; n = 5 control). Arrow highlights colocalization of nuclear marker DAPI and pSTAT3. o, STAT3 phosphorylation (Y705) by flow cytometry in primary mouse GL261 TCM-treated astrocytes (n = 3 control, n = 5 per TCM group). p, Effect of the STAT3 inhibitor (inhib.) BAY2353 on GL261 TCM-induced Tnfsf10 expression in primary mouse astrocytes (n = 6 each). q, Effect of STAT3 inhibition on TNFSF10 expression in U87 TCM-stimulated human astrocytes (n = 5 each). r, Effect of STAT3 inhibition on TRAIL surface expression in primary mouse astrocytes treated with GL261 TCM (n = 5 each). Data are the mean ± s.e.m. Unpaired two-tailed t-test used for statistical analyses. n indicates biologically independent samples. Scale bars, 50 µm (m, top), 25 µm (n), 10 µm (m, bottom).
To investigate the regulation of TRAIL expression in astrocytes, we performed RNA-seq on TRAIL+ and TRAIL− reporter cells isolated by flow cytometry from GL261-implanted Aldh1l1cre-ERT2/TdTomato reporter mice (Fig. 4e and Supplementary Data 7). Pathway and upstream regulator analyses showed that STAT3 signalling was increased in TRAIL+ astrocytes isolated from GL261-tumour-bearing mice (Fig. 4f,g). Of note, although calcium signalling-related genes were enriched in this astrocyte subset, stimulation of primary astrocytes with GL261 TCM did not induce significant Ca2+ influx (Extended Data Fig. 7b). A STAT3-driven transcriptional signature detected by scRNA-seq was also enriched in cluster 2 astrocytes, which were associated with a shorter time to recurrence (Figs. 1a and 4h,i), and in cluster 1 astrocytes expanded in patients with glioblastoma in our validation snRNA-seq studies (Figs. 1l and 4j). Moreover, TRAIL-expressing cluster 1 astrocytes in the GL261 glioma model also displayed an increased STAT3 transcriptional signature, as detected by scRNA-seq (Figs. 2d and, 4k,l). Indeed, in immunofluorescence studies, STAT3 phosphorylation and TRAIL expression colocalized in astrocytes in human glioblastoma and GL261 glioma samples (Fig. 4m,n). Spatial transcriptomic analyses showed a significant correlation between STAT3 signalling and TNFSF10 expression at the leading edge (Extended Data Fig. 7c). Finally, STAT3 phosphorylation was higher in TRAIL+ astrocytes than in TRAIL– astrocytes in the genetically engineered MK007 glioma model (Extended Data Fig. 7d). Together, these results suggest that STAT3 has a role in the regulation of TRAIL expression in glioma-associated astrocytes.
In agreement with the increase in STAT3 signalling in TRAIL+ astrocytes, we identified a correlation between STAT3 phosphorylation and TNFSF10 expression in public databases (Extended Data Fig. 7e). Indeed, we found multiple STAT3-binding sites in the TNFSF10 promoter (Extended Data Fig. 7f). Moreover, in functional validation studies, GL261 TCM induced STAT3 phosphorylation in primary mouse astrocytes in vitro (Fig. 4o and Extended Data Fig. 7g). Conversely, STAT3 inhibition abrogated the upregulation of TRAIL expression induced in mouse and human astrocytes by TCM (Fig. 4p–r). Thus, these findings indicate that glioblastoma-secreted factors induce TRAIL expression in astrocytes in a STAT3-dependent manner.
Glioblastoma-derived IL-11 drives TRAIL+ astrocytes
To identify glioblastoma-secreted molecules that induce STAT3-dependent TRAIL expression in astrocytes, we screened the TCGA database for STAT3-activating glioblastoma-secreted factors for which expression levels correlate with TRAIL expression. This analysis identified the cytokine IL-11 as a candidate inducer of TRAIL expression in astrocytes (Fig. 5a and Supplementary Data 8).

a, Pearson correlation of TNFSF10 in the TCGA glioblastoma dataset. b, IL11 expression in a glioblastoma scRNA-seq dataset4. OPC, oligodendrocyte progenitor cell; NPC, neural progenitor cell; AC, astrocyte; MES, mesenchymal. c, EGFRvIII immunohistochemistry and IL-11+ glioblastoma cell immunofluorescence. d, Peritumoural IL11RA+TRAIL+ astrocyte immunofluorescence in glioblastoma. e, Glioblastoma survival stratified by the first and fourth quartile IL11 expression (data from TCGA). f, IL-11 signalling score (WP2332) in GL261 Aldh1l1cre-ERT2/TdTomato reporter cell scRNA-seq (Fig. 2b). g, IL-11 immunofluorescence in GL261 at 15 days. h, IL-11 concentration in GL261-implanted brain lysates at 15 days (n = 3 each). i, Astrocyte STAT3 phosphorylation after treatment with recombinant mouse IL-11 (rmIL-11) (n = 3 control, n = 5 TCM). j–l, TNFSF10 expression by quantitative PCR (qPCR) (j,k) and flow cytometry (l) of primary mouse astrocytes treated with rmIL-11 (j: n = 23 control, n = 14 rmIL-11, l: n = 3 control, n = 5 rmIL-11) and human astrocytes treated with human IL-11 (rhIL-11) (k; n = 5 control, n = 11 rhIL-11). m–o, TNFSF10 expression by qPCR (m,n) and flow cytometry (o) in IL-11-treated mouse (m, n = 4 each) and human (n, n = 5 each) astrocytes with STAT3 inhibition. p,q, TNFSF10 expression in GL261 TCM-treated mouse (p) and U87 TCM-treated human (q) astrocytes and IL-11 neutralization (n = 5 each). r, GL261 survival after astrocyte Il11ra1 inactivation (n = 18 NT sgRNA, n = 28 sgIl11ra1). Weight fold change in survivors, sgIl11ra1 1.064 ± 0.028 g, NT sgRNA 1.035 ± 0 g (sgIl11ra1 versus NT sgRNA, P = 0.7015 two-tailed t-test). s,t, TRAIL+ (s) and pSTAT3+TRAIL+ (t) TdTomato+ reporter cells in GL261 at 15 days (n = 3 NT sgRNA, n = 5 sgIl11ra1). u, Apoptotic CD4+ and CD8+ tumour-infiltrating T cells (n = 5 each). v, Survival in IL11-overexpressing GL261 mice (n = 10 each). w–y, TRAIL+ (w) and pSTAT3+TRAIL+TdTomato+ reporter cells (x), and apoptotic T cells (y) in IL11-overexpressing GL261 mice at 12 days (n = 5 each). Data are the mean ± s.e.m. Unpaired two-tailed t-test. log-rank (Mantel–Cox) test (e,r,v). n indicates biologically independent samples. Scale bars, 25 µm (d, right) 50 µm (c, right; g, bottom; d, left bottom), 100 µm (c, left; d, left top), 500 µm (g, top).
IL-11 is a member of the IL-6 family and signals through its specific receptor chain IL11RA and gp130 (ref. 31). An analysis of human glioblastoma scRNA-seq and cell-type deconvoluted TCGA datasets showed that IL-11 expression was highest in malignant cells across all glioblastoma subtypes, whereas IL11RA was more broadly expressed in malignant and non-malignant compartments (Fig. 5b and Extended Data Fig. 8a–c). In spatially resolved transcriptomic analyses, we found associations between STAT3 signalling and TRAIL-induced apoptosis programs at the leading edge (Extended Data Fig. 8d), whereas IL-11 expression was most correlated with mesenchymal glioblastoma metaprograms32,33 (Extended Data Fig. 8e). In validation multiplex immunofluorescence studies, we detected IL11 expression in human glioblastoma and IL11RA expression in TRAIL+ astrocytes in the glioblastoma TME (Fig. 5c,d). Furthermore, high IL11 expression was associated with shorter patient survival in the TCGA dataset (Fig. 5e), and increased astrocyte IL11RA expression was associated with earlier glioblastoma recurrence (Extended Data Fig. 8f,g). Finally, in a scRNA-seq analysis, cluster 1 astrocytes in GL261-glioma-bearing mice displayed increased IL-11 signalling (Figs. 2d and 5f and Extended Data Fig. 8h). Moreover, we detected IL-11 expression in vivo in GL261 tumours by immunofluorescence (Fig. 5g). Secreted IL-11 was detectable in GL261 tumour lysates and in TCM from primary glioblastoma lines, GL261 or genetically engineered glioma lines (Fig. 5h and Extended Data Fig. 8i–k).
To further explore the regulation of TRAIL expression in astrocytes by glioblastoma-secreted IL-11, we stimulated in vitro mouse and human astrocytes with species-matched recombinant IL-11 (rIL-11). rIL-11, at levels comparable to those detected in GL261 TCM, activated STAT3 in astrocytes, transactivated the TNFSF10 promoter and induced TRAIL expression (Fig. 5j–l and Extended Data Fig. 8l,m). Notably, the induction of TRAIL expression by IL-11 was suppressed in mouse and human astrocytes through pharmacological inhibition of STAT3 (Fig. 5m–o). Similarly, IL-11 neutralizing antibodies abrogated the upregulation of Tnfsf10 expression induced by GL261 or U87 TCM in mouse and human astrocytes, respectively (Fig. 5p,q). Of note, rIL-11 stimulation did not increase Tnfsf10 expression in microglia, oligodendrocytes or neurons in culture (Extended Data Fig. 8n). Last, other STAT3-activating glioblastoma-secreted factors did not induce significant TRAIL expression in astrocytes (Extended Data Fig. 8o,p). These findings indicate that glioblastoma-produced IL-11 induces TRAIL+ astrocytes in a STAT3-dependent manner.
To evaluate the role of IL-11 on the suppressive activity of TRAIL+ astrocytes on T cells, we performed in vitro co-culture experiments. IL-11 blockade with specific antibodies suppressed CD4+ and CD8+ T cell apoptosis induced by TCM-treated astrocytes (Extended Data Fig. 9a). Conversely, IL-11-treated astrocytes induced TRAIL-dependent T cell apoptosis, which was abrogated by pretreatment with a STAT3 inhibitor (Extended Data Fig. 9b–d).
To study the relevance of glioma-produced IL-11 for the control of TRAIL+ astrocytes and T cells in vivo, we specifically inactivated Il11ra1 in astrocytes using lentivirus-delivered Cas9 and sgRNA as previously described26 (non-targeting sgRNA was used as a control). Il11ra1 inactivation in astrocytes increased survival after orthotopic GL261 implantation, and TRAIL+ and phosphorylated STAT3+ (pSTAT3+) Aldh1l1cre-ERT2/TdTomato reporter cells and apoptotic CD4+ and CD8+ T cells were reduced (Fig. 5r–u and Extended Data Fig. 9e,f). Conversely, IL-11 overexpression in GL261 cells decreased survival, and it augmented TRAIL expression and STAT3 activation in Aldh1l1cre-ERT2/TdTomato reporter cells and apoptosis in CD4+ and CD8+ T cells (Fig. 5v–y and Extended Data Fig. 9g). Of note, IL-11 overexpression did not alter GL261 cell growth in vitro, but IL-11-overexpressing cells showed a trend to grow faster in vivo (Extended Data Fig. 9g–i).
Finally, to investigate the regulation of IL-11 production in glioblastoma, we queried the IL11 promoter sequence, and detected multiple binding sites for the aryl hydrocarbon receptor (AHR), which has been shown to have important roles in glioblastoma growth, invasiveness and immunoevasion9,33,34 (Extended Data Fig. 9j). CRISPR–Cas9-driven Ahr inactivation in GL261 led to a significant reduction in IL-11 production (Extended Data Fig. 9k), whereas overexpression of a constitutively active AHR increased IL-11 expression (Extended Data Fig. 9l). The tryptophan metabolite kynurenine induces AHR signalling in multiple contexts, including glioblastoma9,34. The stimulation of GL261 cells with kynurenine induced Il11 expression in an AHR-dependent manner (Extended Data Fig. 9m). Similarly, kynurenine induced IL11 expression in human U87 glioblastoma cells (Extended Data Fig. 9n). Altogether, these findings indicate that IL-11 produced by glioma cells in an AHR-dependent manner induces TRAIL+ astrocytes, which suppress CD4+ and CD8+ T cell responses through the induction of apoptosis.
TRAIL blockade boosts tumour immunity
Oncolytic viruses are attractive tools for cancer immunotherapy because of their ability to selectively lyse tumour cells35. Moreover, oncolytic viruses can be engineered to express immunomodulatory molecules with the goal of reversing the immunosuppressive characteristics of the TME35. Thus, to disrupt the suppression of tumour-specific T cell immunity by TRAIL+ astrocytes, we used an oncolytic herpes simplex virus-1 (HSV-1) vector optimized for safety and potency36,37 (Supplementary Tables 2 and 3). We confirmed the ability of the HSV-1 vector to infect primary human glioblastoma cells and further engineered it to express a single-chain TRAIL-blocking antibody (scFv) in infected glioblastoma cells (HSV-1(anti-TRAIL)) (Fig. 6a and Extended Data Fig. 10a,b).

a, In vitro infection of primary human glioblastoma line BT145 with HSV-1(anti-TRAIL) at the indicated multiplicity of infection (m.o.i.). GFP fluorescence detection was indicative of virus-infected cells 72 h after infection. Interferon-defective Vero cells shown as positive control. Scale bar, 100 µm. b, Experimental design (top) of GL261–mNectin1-implanted mice randomized by tumour bioluminescence signal and treated with 3 × 106 plaque-forming units (p.f.u.) HSV-1 intratumourally 7 days after implantation. Tumour bioluminescence (bottom) 1 day before and 9 days after treatment with HSV-1 (n = 8 each). c, Survival of GL261–mNectin1-implanted mice treated with HSV-1 engineered to produce TRAIL-blocking single-chain antibody (scFv); mice treated with PBS, empty HSV-1 or HSV-1 expressing a non-targeting scFv were included as controls (n = 15 for PBS, n = 30 HSV-1(empty), n = 15 HSV-1(anti-TRAIL), n = 11 HSV-1(control scFv)). Body weight fold change from initiation of study in surviving mice, HSV-1(anti-TRAIL) 0.9079 ± 0.049 g, HSV-1(control scFv) 0.9344 ± 0 g, HSV-1(empty) 0.9887 ± 0.027 g (HSV-1(anti-TRAIL) versus HSV-1(empty), P = 0.2114; HSV-1(anti-TRAIL) versus HSV-1(control scFv), P = 0.9561 by two-sided ANOVA). d, Tumour-infiltrating cleaved caspase-3/7+CD4+ or CD8+ T cells in HSV-1-treated mice 17 days after GL261 implantation (n = 8 each). e–h, Frequency (e) and absolute count (f) of HSV-1-specific or of GARC-1(77–85)-specific (g,h) CD8+ T cells infiltrating the GL261 TME in HSV-1-treated mice 17 days after implantation (n = 8 each). i, Frequency of activated GARC-1(77–85)-specific PD-1+CD8+ T cells. j–m, Heatmap (left) and pathway analysis (right) of bulk RNA-seq data analysis from FACS-sorted tumour-infiltrating CD8+ T cells (j), CD4+ T cells (k), microglia-derived TAMs (l) and monocyte-derived TAMs (m) in HSV-1-treated mice 17 days after tumour implantation (n = 5 HSV-1(empty); n = 4 except monocytes, for which n = 3 for HSV-1(anti-TRAIL)); all pathways shown are P < 0.05. Data are the mean ± s.e.m. Paired two-tailed t-test (b). Survival analysis by log-rank (Mantel-Cox) test (c). Unpaired two-tailed t-test elsewhere. n indicates biologically independent samples.
We injected HSV-1(anti-TRAIL) intratumourally 7 days after the orthotopic implantation of GL261 cells expressing the HSV entry receptor mNectin-1 into immunocompetent B6 mice. HSV-1 not expressing any cargo (HSV-1(empty)) or expressing a nonspecific scFv (HSV-1(control scFv)) were used as controls. HSV-1(anti-TRAIL) treatment reduced tumour load (quantified on day 16 by in vivo bioluminescence imagining) and increased survival (Fig. 6b,c). HSV-1(anti-TRAIL) treatment also reduced CD4+ and CD8+ T cell apoptosis in the TME, as detected by flow cytometry, and increased the number of T cells in the TME, whereas the number of TAMs remained unchanged (Fig. 6d and Extended Data Fig. 10c,d). Moreover, HSV-1(anti-TRAIL) administration increased the proportion and number of tumour-specific and virus-specific CD8+ T cells in the TME detected using peptide-loaded MHCI tetramers (Fig. 6e–h). Notably, HSV-1(anti-TRAIL) treatment increased the abundance of tumour-reactive activated PD-1+SLAMF6−TIM3− CD8+ T cells in mice (Fig. 6i and Extended Data Fig. 10e–h), whereas expression of the exhaustion marker CTLA-4 was decreased (Extended Data Fig. 10i,j). TNF production was increased in TME-infiltrating CD8+ T cells from HSV-1(anti-TRAIL)-treated mice, whereas IL-10 production in CD4+ T cells was reduced (Extended Data Fig. 10k,l). These findings indicate that TRAIL blockade in the TME with HSV-1(anti-TRAIL) enhances tumour-specific T cell immunity by modulating both CD4+ and CD8+ T cell compartments.
Finally, we characterized by RNA-seq the effects of HSV-1(anti-TRAIL) treatment on T cells and on microglia-derived and monocyte-derived TAMs in the GL261 TME. We detected reduced apoptotic signalling and increased pro-inflammatory functions in CD8+ T cells of HSV-1(anti-TRAIL)-treated mice (Fig. 6j and Supplementary Data 9). Moreover, HSV-1(anti-TRAIL) administration upregulated the expression of transcriptional modules in CD4+ T cells associated with T helper 1-type responses and classical macrophage activation (Fig. 6k and Supplementary Data 10). In agreement with these findings, HSV-1(anti-TRAIL) treatment activated phagocytic, tumouricidal and immunogenic signalling pathways in microglia-derived and monocyte-derived TAMs, and IL-10 signalling was reduced (Fig. 6l,m and Supplementary Data 11 and 12). HSV-1(anti-TRAIL) treatment also enhanced bidirectional TAM–T cell crosstalk, as indicated by the increased expression of genes involved in T cell recruitment (Cxcl9, Cxcl10 and Cxcl11) and co-stimulation (Cd80 and Cd40) (Fig. 6m). In summary, TRAIL blockade in the TME using the engineered oncolytic virus HSV-1(anti-TRAIL) enhanced tumour-specific T cell and myeloid immune responses (Extended Data Fig. 10m).
Discussion
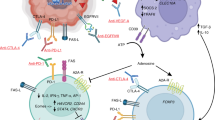
Astrocytes are the most abundant glial cells of the CNS and can regulate the immune response in the context of autoimmunity and other neurological conditions. However, despite their abundance and immunoregulatory functions25,26,38,39, little is known about the role of astrocytes in the control of glioblastoma-specific immunity. Here we described a subset of TRAIL+ astrocytes induced by tumour-secreted IL-11 in a STAT3-dependent manner, and suppressed glioblastoma-specific immunity by inducing apoptosis in CD4+ and CD8+ T cells.
TRAIL participates in the regulation of T cell responses through the induction of apoptosis through DR4 and DR5 signalling24 and through non-apoptotic mechanisms, such as the modulation of T cell receptor signalling and DC functions30,40,41,42. T cell apoptosis and dysfunction are detected in patients with glioblastoma7, even after immune checkpoint blockade3. We identified IL-11-induced TRAIL expression in astrocytes as a driver of T cell apoptosis in glioblastoma. In the context of CNS inflammation in experimental autoimmune encephalomyelitis and multiple sclerosis, TRAIL+ astrocytes can limit T cell-driven autoimmunity25. Thus, our studies suggest that these anti-inflammatory effects of TRAIL+ astrocytes are exploited by glioblastoma to evade tumour-specific immunity. TRAIL+ astrocytes probably synergize with other astrocyte subsets and cell types in the glioblastoma TME to suppress tumour-specific T cells through additional immunoregulatory mechanisms, such as the activation of immune checkpoint signalling, neuroimmune modulation and the production of immunosuppressive metabolites1,20. However, our functional studies support a substantial role for TRAIL+ astrocytes in the control of glioblastoma-specific immunity. Our findings also suggest that TRAIL+ astrocytes regulate microglia-derived and monocyte-derived TAMs, probably through indirect mechanisms such as cytokine-mediated T cell–TAM interactions, as previously described43. These additional effects of TRAIL+ astrocytes on TAMs and potentially other TME components are likely to further amplify T cell dysfunction.
The distinction of cancer cells and astrocytes in tissues through the use of individual protein markers is currently a technical limitation in the field. In this study, the subclonal neoantigen EGFRvIII was used as a tumour cell marker in combination with histological localization to identify glioblastoma-associated astrocytes. Indeed, TRAIL expression has also been reported in brain cancer cells44. Although we observed similar survival rates in TRAIL-deficient glioma models, TRAIL expression has been linked to cancer invasion and may play non-apoptotic roles45. Indeed, a positive correlation between DR5 expression in glioblastoma cells and survival outcomes46 suggests that TRAIL-triggered apoptosis in tumour cells may contribute to the antitumour response, which highlights challenges in the design of therapies based on targeting cancer cells with recombinant TRAIL while leaving T cells unharmed.
We showed that tumour-produced IL-11 drives TRAIL expression in glioblastoma-associated astrocytes in a STAT3-dependent manner. IL-11 can contribute to glioblastoma pathology through its cell autonomous effects on tumour cells, boosting tumour formation47 and metabolic fitness48. However, the role of IL-11 in the control of the immunosuppressive nature of the glioblastoma TME is unclear. Notably, IL-11 is reported to contribute to breast cancer pathology in a preclinical model in a non-tumour cell autonomous manner through its effects on the immune response in the TME49. Using in vivo cell-specific genetic perturbation approaches in combination with in vitro mouse and human experimental systems, we established that IL-11 produced by glioblastoma cells induces TRAIL+ astrocytes to suppress tumour-specific T cell immunity. Although our scRNA-seq and immunostaining analyses detected glioblastoma cells as the major source of IL-11, astrocytes50 and TAMs47 can also produce IL-11 in the CNS. Thus, additional sources of IL-11 in the glioblastoma TME besides tumour cells may also induce and stabilize TRAIL expression in astrocytes, thereby further promoting tumour immunoevasion. IL-11 may also participate in the regulation of STAT3+ astrocytes identified in the TME of brain metastases18. These effects of IL-11 on the regulation of TRAIL+ astrocytes may synergize with additional roles of IL-6 cytokine family members reported to operate in the glioblastoma TME to promote tumour pathology, such as IL-6, IL-27 and OSM51. Besides its effects on TRAIL expression, IL-11 may regulate additional STAT3-dependent mechanisms through which astrocytes limit the T cell response to tumours. Notably, IL-11-blocking antibodies are being developed for clinical use, with trials planned to treat pulmonary fibrosis and other fibrotic diseases (NCT05658107).
A challenging aspect for the design of efficacious therapies for glioblastoma is the strongly immunosuppressive nature of the TME1, which is a major barrier for the success of immunotherapeutic interventions. TME myeloid cells6,7,8,9, neurons52 and neutrophils10 can contribute to glioblastoma progression. STAT3 inhibitors have been tested as potential glioblastoma therapeutics, but their clinical benefits are not yet clear53. Attempts to therapeutically induce TRAIL-dependent apoptosis in glioblastoma cells have so far produced unsatisfactory clinical results because of tumour cell resistance to apoptosis and challenges associated with drug delivery across the blood–brain barrier54. Moreover, when compared with other forms of tumour cell death, apoptosis induces weaker tumour-specific immune responses40. In this context, oncolytic viruses offer opportunities for cancer immunotherapy, as they selectively lyse tumour cells while creating a pro-inflammatory microenvironment that induces protective T cell responses against viral and tumour antigens35. Oncolytic virotherapies have been evaluated for glioblastoma. For example, treatment with HSV-1 G207 or HSV-1 G47Δ showed evidence of positive clinical responses in paediatric recurrent/progressive high-grade glioma55 and adult glioblastoma56, respectively, linked to the lysis of tumour cells and increased immune infiltration of the TME. Moreover, survival of patients with glioblastoma after treatment with an optimized HSV-1 was recently associated with transcriptional signatures of immune activation and the expansion of specific T cell clones in the TME57. Here we engineered an oncolytic HSV-1, optimized for glioblastoma targeting and displaying enhanced safety mechanisms, to produce a TRAIL-blocking single-chain antibody in the TME. This engineered HSV-1 co-opted the T cell boosting abilities of HSV-1 while limiting TRAIL-driven CD4+ and CD8+ T cell apoptosis, which resulted in increased survival and tumour-specific T cell responses. In this respect, our findings resemble studies that used engineered bacteria for therapeutic lysis and TME remodelling of peripheral tumours, with the caveat that safety considerations prevent the use of engineered bacteria to modify the TME of brain tumours58.
In summary, we established that IL-11–STAT3-driven astrocytes suppress glioblastoma-specific protective immunity by inducing TRAIL-dependent T cell apoptosis, and we developed engineered therapeutic viruses to target this mechanism of astrocyte-driven tumour immunoevasion. Targeting TRAIL+ astrocyte-driven T cell apoptosis may provide an efficacious avenue to reinvigorate tumour-specific protective immunity and boost the therapeutic effects of oncolytic virotherapy57, vaccination59 or chimeric antigen receptor T cells60 in glioblastoma.
Methods
Mice
C57BL/6J mice (The Jackson Laboratory, 000664) were used as the genetic background for all in vivo experiments. Mice were housed in a conventional, pathogen-free facility at the Hale Building for Transformative Medicine under a standard 12-h light–dark cycle at 20–23 °C and 50% humidity with access to water and food ad libitum and in accordance with guidelines for care and experimentation defined by the Brigham and Women’s Hospital Institutional Animal Care and Use Committee (IACUC). Transgenic mouse genotypes were confirmed by PCR (Transnetyx). When Aldh1l1cre-ERT2/TdTomato reporter mice (The Jackson Laboratory, 031008 and 007909) were used, 200 mg kg–1 tamoxifen (Sigma-Aldrich, T5648) diluted in corn oil (Sigma-Aldrich, C8267) was intraperitoneally administered 2–4 weeks before starting the experiment, and TdTomato induction was confirmed by flow cytometry at the time of measurement. Where indicated, Aldh1l1cre-eGFP mice (The Jackson Laboratory, 026033) and Rag2–/– mice (The Jackson Laboratory, 008449) were used. Mice were used between the ages of 8 and 12 weeks. Mice were randomly assigned to experimental conditions. In oHSV treatment studies, treatment groups were balanced by randomizing mice prior to treatment based on tumour burden.
Human glioblastoma samples
Resected glioblastoma and healthy brain tissue samples were collected from patients with glioblastoma with full ethical approval and informed consent as approved by the Department of Oncologic Pathology at Dana Farber Cancer Institute and stored at −80 °C. De-identified clinical data, including patient age, sex, diagnosis, treatment history, isocitrate dehydrogenase (IDH) mutation status, O6-methylguanine-DNA methyltransferase (MGMT) promoter methylation status and cytogenetic report of copy number variations are provided in Supplementary Table 1. All patients were recruited through the Department of Oncologic Pathology at the Dana Farber Cancer Institute. Informed consent from both the patients and the family was obtained. Patients were chosen on the basis of a clinical diagnosis of glioblastoma, thus self-selection bias is not relevant.
snRNA-seq
To avoid transcriptional artefacts resulting from bias in the recovery of different cell types from frozen tissue, a previously described dissociation protocol for nucleus isolation was adapted61. In brief, frozen tissue was minced with a razor blade and mechanically homogenized in Nuclei EZ lysis buffer (Sigma-Aldrich, NUC101-1KT). After incubation for 5 min, the homogenate was filtered through a 70 µm strainer mesh. Two washing steps were performed with a 1% solution of BSA (Sigma-Aldrich, A3294-100G) in PBS (Thermo Fisher Scientific, 14190250), with the addition of 0.2U µl–1 RNase inhibitor (Lucigen, 30281-2). Centrifugation was performed at 500g for 5 min at 4 °C. To separate nuclei from cell debris, nucleus suspensions were incubated for 10 min with 10 µg ml–1 DAPI (Sigma, D9542) in wash buffer to stain DNA. Subsequently, DAPI+ nuclei were sorted by FACS and used for Drop-seq as described below.
Drop-seq
Drop-seq62 was used for scRNA-seq through the co-encapsulation of uniquely barcoded beads and single cells in nanolitre droplets before cell or nucleus lysis and transcriptome capture. Following a previously described protocol62, barcoded MACOSKO-2011-10 B beads (Chemgenes) linked to 108 uniquely barcoded oligonucleotides (5′-Bead–Linker-TTTTTTTAAGCAGTGGTATCAACGCAGAGTACJJJJJJJJJJJJNNNNNNNNTTTTTTTTTTTTTTTTTTTTTTTTTTTTTT-3′) were resuspended in a wash solution made of 10 mM Tris pH 8.0 (Sigma-Aldrich, T3038), 1 mM EDTA (Amresco, E177-100ML) and 0.01% Tween‐20 (Sigma-Aldrich, P9416) (TE-TW) at a density of 3 × 105 ml–1 and stored at 4 °C. Immediately before use, beads were centrifuged at 1,000g and resuspended in 1 ml lysis buffer composed of 570 µl OptiPrep (Sigma-Aldrich, D1556-250ML), 120 µl 20% Ficoll PM‐400 (Cytiva, 17-0300-10), 20 µl 10% Sarkosyl (Sigma-Aldrich, L7414-50ML), 40 µl of 0.5 M EDTA (Amresco, E177-100ML), 200 µl of 1 M Trizma hydrochloride solution pH 8.0 (Sigma-Aldrich, T2663-1L) and 50 µl of 1 M dithiothreitol (DTT; Thermo Fisher Scientific, P2325). Sorted cells or nuclei were taken up in 16% OptiPrep at a concentration of 2.5 × 105 ml–1 in 1 ml. If fewer cells or nuclei were available, Fluorinert FC-40 (Sigma Aldrich, F9755-250ML) was used as a backflow liquid. If fewer than 1 × 105 cells were available, bead concentrations were doubled to 6 × 105 ml–1 to increase the capture rate, which follows a Poisson distribution. Beads and cells were taken up in a 3 ml syringe (BD), adding a magnetic disk (VP Scientific, 772DP-N42-5-2) and using a magnetic stirrer (VP Scientific) to homogenize bead suspensions. QX200 Droplet Generation Oil for EvaGreen (Bio-Rad, 1864006) was taken up in a 10 ml syringe (BD). Three Legato 100 syringe pumps (KD Scientific) were loaded with oil, cell and bead syringes, and PE/2 polyethylene microtubing (Scientific Commodities, BB31695-PE/2) was used to connect them to the device input holes of polydimethylsiloxane (PDMS) devices placed on an inverted light microscope with an attached Fire-i 830b camera (Unibrain). Fire-i (Unibrain) was used to visualize rapidly flowing droplets with the lowest shutter speed and highest exposure and gain settings. The device was initially primed with oil before connecting cells and bead flows at flow rates of 4,500, 1,500 and 1,500 µl h–1, respectively. Output flow containing droplets was collected for 20–25 min or until the cell suspension was used up. For droplet breakage, approximately one-third of excess oil volume was added as 97% 1H,1H,2H,2H-Perfluoro-1-octanol (Sigma-Aldrich, 370533), along with 30 ml of 6× sodium chloride-sodium citrate (SSC) buffer (National Diagnostics, EC-873). Emulsions were shaken a few times and centrifuged at 1,000g for 1 min, resulting in a visible bead layer floating at the interface of the oil and SSC. In the subsequent steps, as much of the oil and SSC as possible was removed while topping up with fresh SSC and centrifuging to wash the beads before reverse transcription (RT) into cDNA. Beads were washed in 300 µl 1× RT buffer (Thermo Fisher Scientific, EP0742) before incubation in 200 µl RT mix composed of 75 µl H2O, 40 µl 5× RT buffer, 40 µl 20% Ficoll PM-400 (Cytiva, 17-0300-10), 20 µl 1 mM dNTPs (Thermo Fisher Scientific, R0194), 5 µl RNase inhibitor (Lucigen, 30281-2), 10 µl 50 µM template switch oligo (AAGCAGTGGTATCAACGCAGAGTGAATrGrGrG, IDT) and 10 µl (with 2,000 U) Maxima H Minus reverse transcriptase (Thermo Fisher Scientific, EP0742) for 30 min at room temperature and 90 min at 42 °C (with rotation in both cases). Reaction mixtures were then washed with 1 ml of a solution containing 10 mM Tris pH 8.0, 1 mM EDTA and 0.5% sodium dodecyl sulfate (Sigma-Aldrich, L3771-100G) (TE-SDS), twice with 1 ml of TE-TW and once with 1 ml 10 mM Tris pH 8.0. To remove bead-linked oligonucleotides that had not captured a mRNA molecule, beads were incubated in a mix of 20 µl 10× exonuclease I buffer (Thermo Fisher Scientific, EN0581), 170 µl H2O and 10 µl (with 200 U) exonuclease I (Thermo Fisher Scientific, EN0581) for 45 min at 37 °C with rotation. After washing once with TE-SDS, twice with TE-TW and once with 1 ml H2O, beads were counted using a haemocytometer chamber and divided into 4,000 beads per aliquot in PCR tubes. A PCR mix containing 24.6 µl H2O, 0.4 µl 100 µM SMART PCR primer (AAGCAGTGGTATCAACGCAGAGT) (IDT) and 25 µl 2× Kapa HiFi Hotstart Readymix (Kapa Biosystems, 7958927001) were added. PCR was performed in a thermocycler with the following program to amplify cDNA molecules: 95 °C (3 min); 4 cycles of 98 °C (20 s), 65 °C (45 s), 72 °C (3 min); 9 cycles of 98 °C (20 s), 67 °C (20 s), 72 °C (3 min); 72 °C (5 min); and 4 °C hold. PCR products were purified using AMPure XP beads (Beckman Coulter, A63881) at a 0.6× ratio according to manufacturer’s protocol, eluting all samples in 50 µl molecular-grade H2O (Thermo Fisher Scientific, AM9937). At this point, the distribution of fragment sizes in cDNA libraries was quantified using a 2100 Bioanalyzer (Agilent) instrument with a High Sensitivity DNA kit (Agilent, 5067-4626). Library concentration and quality were assessed using a rounded curve, with average sizes around 700 bp. In cases when the DNA concentration was not sufficient, additional PCR cycles with the following conditions were added as needed: 98 °C (20 s), 67 °C (20 s), 72 °C (3 min). Around 600 pg of cDNA library in 5 µl was used for fragmentation and addition of sample tags using a Nextera XT DNA Library Prep kit (Illumina, FC-131-1096). cDNA was added to 10 µl Nextera tagment DNA buffer and 5 µl Amplicon tagment mix. After incubation for 5 min at 55 °C, 5 µl Neutralize tagment buffer was added. After 5 min at room temperature, a mix containing 15 µl Nextera PCR mix, 8 µl H2O, 1 µl 10 µM New-P5-SMART PCR hybrid oligonucleotide (AATGATACGGCGACCACCGAGATCTACACGCCTGTCCGCGGAAGCAGTGGTATCAACGCAGAGT*A*C) (IDT) and 1 µl of a 10 µM sample-specific Nextera indexing oligonucleotide (Illumina, FC-121-1011) was obtained. Samples underwent PCR with the following program: 95 °C (30 s); 12 cycles of 95 °C (10 s), 55 °C (30 s), 72 °C (30 s); 72 °C (5 min); and 4 °C hold. cDNA was purified AMPure XP beads at a 0.6× ratio and eluted in 10 µl H2O. Library distribution was analysed using a 2100 Bioanalyzer and the molarity-weighted average fragment size was calculated. A Library Quantification kit (Kapa Biosystems, 07960140001) was used to determine library concentrations by qPCR according to standard protocols. Finally, 4 µl samples were pooled at an equimolar ratio of 5 nM. Finally, samples were sequenced using NovaSeq S2 with a Custom Read 1 primer (GCCTGTCCGCGGAAGCAGTGGTATCAACGCAGAGTAC), estimating around 25,000 reads per STAMP. STAMPs were estimated by multiplying the cell number used (determined by flow rate, run time and concentration) with the co-encapsulation probability (given by the product of the encapsulation probability of either cells or beads). The co-encapsulation probability of single units follows a Poisson distribution as a function of droplet volume and cell or bead concentration and was 2.5% for mouse samples and on average 1.4% for human nuclei.
PDMS device fabrication
Microfluidic devices were microfabricated by regular photolithography techniques at the Harvard Medical School Microfluidics/Microfabrication Core Facility. To generate a master mould with features of 125 µm height, a silicon wafer (University Wafer) was spin-coated with the epoxy-based negative photoresist SU-8 (Kayaku Advanced Materials) and using a Mask Aligner exposed to ultraviolet light through a photomask (CAD/Art) containing the Drop-seq pattern (https://mccarrolllab.org/dropseq). Spin coating and exposure conditions were determined following the SU-8 manufacturer’s protocol. The photoresist was then treated with SU8 developer, and isopropanol was used to rinse off undeveloped SU-8. Finally, the wafer was heated at 150 °C for 5 min. For replica moulding with PDMS, Sylgard 184 silicone elastomer and Dowsil 184 curing agent (Dow, DC4019862) were mixed at a 1:10 ratio. The mixture was poured into the master mould and air bubbles removed by placing in a vacuum degassing chamber for 2–3 h. Finally, the elastomer was cured at 65 °C for at least 4 h or preferably overnight. Next, 0.75-mm-wide holes were punched using a biopsy punch (Rapid Core). Devices were bonded to glass microslides (Fisher Scientific) by treatment with oxygen plasma (Technics Plasma Etcher 500-II) and baking at 150 °C for 10 min. Finally, channels were rendered hydrophobic by treatment with filter-sterilized Aquapel (Aquapel Glass Treatment) for 5 min. Aquapel was removed with pressurized oxygen and channels washed with Fluorinert FC-40 (Sigma-Aldrich, F9755-250ML) and again flushed with pressurized air. Finally, PDMS devices were baked at 65 °C overnight and sealed with Scotch tape until use.
Orthotopic GL261 model and delivery of lentivirus
A total of 1 × 105 GL261–Luc2 cells (American Type Culture Collection (ATCC)) were intracranially implanted. GL261 cells with passage number <10 were detached using Accutase (StemCell Technologies), and cell suspensions were prepared in serum-free DMEM. The surgical procedure was performed following IACUC guidelines for survival surgery. In brief, a small animal stereotaxic frame (David Kopf Instruments) was placed in a laminar flow cabinet, and a sterile field containing all surgical instruments and tools was established. An activated charcoal filter (VetEquip) was connected to the outflow to adsorb isoflurane. Mice were individually placed in an anaesthesia-induction chamber and a flow of isoflurane (Patterson Veterinary) at 2.5–3 l h–1 mixed with oxygen at 2.5 l h–1 was initiated. The top of the skull was shaved and disinfected using isopropyl swabs (Becton Dickinson) and application of the antiseptic Betadine (Purdue Products). For intracranial injection, the mouse skull was fixed on the stereotaxic frame using ear bars and a mouthpiece for incisor fixation and gas supplementation. Sufficient depth of anaesthesia was confirmed by a toe pinch, and mouse vital signs were monitored throughout the procedure. Following a 2 cm incision on the scalp, the injection coordinates were determined from bregma. A hole was drilled at the +1.8 mm (lateral) and +0.5 mm (anterior) position. A 25 µl glass syringe (Hamilton) was inserted until −3.5 mm depth and slowly retracted 0.5 mm to the final injection site at −3 mm depth. Next, 3 µl of GL261 cell suspension were gradually injected at a rate of 1 µl min–1. Control mice received a sham injection of 3 µl DMEM. For lentivirus administration, 10 µl containing 107 IU of lentivirus was injected at a rate of 2 µl min–1 at the same location of the tumour implant. After an additional minute, the needle was slowly retracted to avoid liquid backflow. The skin was sutured using Ethilon 4-0 non-absorbable filaments (Ethicon) in a simple interrupted pattern. Mice received one perioperative subcutaneous injection of the slow-release analgesic Buprenorphine-SR at a dose of 1 mg kg–1 body weight. Mouse weight was monitored daily, along with levels of activity and signs of wound inflammation or pain. For assessment of tumour burden, in vivo bioluminescence imaging was used. In brief, mice were anaesthetized in a gas chamber as described above, and 0.15 mg kg–1 d-luciferin (Cayman Chemical, 14681) in PBS was intraperitoneally injected. After 10 min, luminescence at 560 nm wavelength was captured multiple times at high sensitivity and 30 s exposure time inside an In-Vivo Xtreme Optical/X-ray imaging system (Bruker). Bruker MI SE v7 (Bruker) software was used for quantification of photons per second (p s–1) in the area of the mouse skull, using the highest intensity recording to capture the peak of bioluminescence. X-ray image overlays were used to limit ROI to mouse skull area. To account for background noise, the median value on the region’s perimeter was subtracted from total p s–1, resulting in net p s–1 as a final measure of tumour burden. In addition to standard humane end points based on clinical symptoms, the maximal tumour measurements permitted by IACUC protocols are a bioluminescence signal of >1 × 108 p s–1, which were not exceeded in any experiment.
In vivo generation of genetically engineered glioma model by electroporation
Electroporation was performed as previously described using a CRISPR–Cas9-based triple gene knockout approach63,64,65. In brief, postnatal day 0 (P0) pups were electroporated with a pX330 plasmid encoding Cas9 and guide sequences targeting Trp53, Pten and Nf1 injected at lambda −3.6 and dorsal–ventral −0.7 with 1 μg DNA in 1 μl. After injection, electric square pulses were laterally delivered using forceps-like electrodes (35 mV (VZ), 50 ms on, 950 ms off, 5 pulses). Mice were co-electroporated with pGlast-PBase and PBCAG-GFPt2aLuc vectors for visualization of tumours. Tumours were collected between days 80 and 100 and brains were digested and myelin removal was performed as described below. Cells were cultured in DMEM supplemented with 10% FBS, 100 U ml–1 penicillin and 100 μg ml–1 streptomycin for at least 3 passages to ensure removal of contaminating stromal cells. A total of 5 × 104 MK007 cells were orthotopically implanted by stereotaxic surgery as described above.
Cell culture
The GL261–Luc2 cell line (Caliper Life Sciences) was cultured in high-glucose DMEM (Gibco, 11965118) supplemented with 10% FBS (Gibco, 10-438-026), 1% penicillin–streptomycin (Thermo Fisher Scientific, 15140122) and 100 µg ml–1 G418 geneticin (Thermo Fisher Scientific, 10131027). U-87 MG cells (ATCC, HTB-14) were cultured in the same medium as GL261–Luc2 cells, except no selection antibiotic G418 was added. Patient-derived human glioblastoma lines BT145, BT482 and BT1018 were obtained through the Dana-Farber Cancer Institute Center for Patient Derived Models (available at models@dfci.harvard.edu) and were generated with prospective patient consent. Human glioblastoma lines were cultured using NeuroCult NS-A Proliferation Kit Human (StemCell Technologies, 05751) supplemented with 1% penicillin–streptomycin, 1:100 0.2% heparin (StemCell Technologies, 07980), 10 ng ml–1 recombinant FGF (Invitrogen, PHG0021) and 20 ng ml–1 EGF (PeproTech, AF-100-15). The HEK293FT cell line (Invitrogen, R70007) was chosen because of its high efficiency for lentivirus production and was cultured with DMEM/F12+GlutaMAX (Thermo Fisher Scientific, 10565-042) with 10% FBS, 1% penicillin–streptomycin and 100 µg ml–1 G418 geneticin (Thermo Fisher Scientific, 10131027). Hep G2 cells (ATCC, HB-8065) were chosen because it is reported to signal through IL-11, and cells were cultured in high-glucose DMEM supplemented with 10% FBS and 1% penicillin–streptomycin. Cell lines were authenticated and tested for mycoplasma by the suppliers. GL261 TCM was prepared as previously described9 by culturing 6 × 106 cells in 10 ml selection antibiotic-free culture medium for 24 h. Culture supernatants were then centrifuged and sterile filtered using a 22 µm filter to remove cellular debris, and aliquots stored at −80 °C until use. For AHR-activation experiments, GL261 cells were stimulated with 100 µM l-kynurenine (Sigma-Aldrich, K8625) or DMSO vehicle for 20 h, with or without pretreatment with the AHR inhibitor CH223191 (Tocris, 3858) for 90 min.
Generation of GL261 cell lines
In brief, 1.5 × 105 cells were seeded in 6-well plates and transfected by dropwise addition of DNA–lipid complexes made with 2.5 µg DNA at a DNA to Lipofectamine 3000 (Thermo Fisher Scientific, L3000008) ratio of 1:3 in Opti-MEM reduced serum medium+GlutaMAX (Thermo Fisher Scientific, 51985034). At 48 h after transfection, antibiotic selection was carried out with 800 µg ml–1 zeocin (Invivogen, ant-zn-1) for 2 weeks. Successfully transfected, antibiotic-resistant clones were further expanded and maintained at 300 µg ml–1 zeocin thereafter. For Il11 overexpression, parental GL261–Luc cells were transfected with a cDNA construct encoding mouse IL-11 and GFP (Sino Biological, MG50117-ACG) as described above and selected with hygromycin B (Cayman Chemical Company, 14291) at 500 µg ml–1. Next, polyclonal populations were FACS sorted to purify highly GFP-positive cells and maintained in 150 µg ml–1 hygromycin B. Finally, growth dynamics were calculated by seeding 20,000 cells and performing cell counting by flow cytometry after 2, 3, 4 or 7 days. Il11 expression was screened by qPCR and ELISA. Clone 2 was selected on the basis of it exhibiting unchanged growth and successfully overexpressing Il11. GL261 cells expressing mNectin-1 (GL261–mNectin1) were generated by lentiviral transduction of mNectin-1 cDNA followed by selection with 2 µg ml–1 puromycin (Gibco, A1113803). Single cells were obtained by limiting dilution and clones were screened for mNectin-1 expression by qPCR (Nectin1, Mm00445392_m1). A TRAIL-deficient GL261 line was generated by transfection of CRISPR–Cas9 lentivirus expression plasmid described below encoding the target sequence described below under the Gfap promoter and selected with zeocin. Ahr knockout GL261 cell line was generated by CRISPR–Cas9 gene editing, using two individual sgRNAs to target Ahr (sgRNA1, 5′-CGGCTTGCGCCGCTTGCGGC -3′; sgRNA2, 5′-AAACGTGAGTGACGGCGGGC-3′) designed using a web resource (https://www.synthego.com/products/bioinformatics/crispr-design-tool). These two sgRNAs were cloned into a sgOpti vector (Addgene, 85681). BsmBI sites and lentivirus particles of sgRNAs and lentiCas9-Blast (Feng Zhang, Addgene 52962) were generated in HEK293NT cells by co-transfecting with the packaging plasmids (pLenti-P2A and pLenti-P2B, LV003, Applied Biological Materials) using Lipofectamine 2000 (Thermo Fisher Scientific, 11668019) according to the manufacturer’s instructions. Lentiviral-particle-containing supernatants were collected 24 and 48 h after transfection. GL261 cells were transduced with the lentivirus in the presence of 5 µg ml–1 polybrene. At 48 h after transduction, cells were selected with blasticidin (5 µg ml–1) and puromycin (4 µg ml–1) for 2 weeks. The efficiency of Ahr knockout was validated by qPCR and immunoblot analyses. To generate GL261 cell lines with constitutively active AHR (AHR-CA) expression, the pBOS-Flag-AHR-CA plasmid (provided by M. Yamamoto66) was used to clone AHR-CA cDNA into the pLenti-CMV-GFP-Hygro (Addgene, 17446) XbaI-SalI site, replacing the GFP sequence. GL261 cells were infected with the lentivirus prepared as described above and were selected with hygromycin (200 μg ml–1) for 10 days. The expression of AHR-CA was verified by qPCR and immunoblot analyses.
Isolation of mouse CNS cells
For cell isolation from fresh mouse brain tissue, anaesthetized mice were perfused with 20 ml cold PBS. The brain was minced and enzymatically dissociated for 20 min, 37 °C and 100 r.p.m. shaking in a solution containing 0.66 mg ml–1 papain (Sigma-Aldrich, P4762) in HBSS without Mg or Ca (Thermo Fisher Scientific, 14175095), followed by a second incubation at the same conditions with an equal volume of DMEM supplemented with collagenase D (0.66 mg ml–1, Roche, 11088858001) and DNase I (8 U ml–1, Thermo Fisher Scientific, 90083). Using a 10 ml serological pipette, the suspension was dissociated and filtered through a 70 µm cell strainer (Fisher Scientific, 22363548) into a 50 ml tube. After centrifugation at 600g for 5 min, the pellet was resuspended in 10 ml of a solution containing 30% Percoll (GE Healthcare Biosciences) in PBS. Following gradient centrifugation for 25 min at 600g with the lowest acceleration and deceleration speeds, the layer of myelin accumulating at the top was aspirated. The cell pellet was washed with PBS and used for downstream applications.
Isolation of mouse splenocytes
Mouse spleens were dissected out of anaesthetized mice before cardiac perfusion, mechanically mashed through 70 µm nylon strainers (Thermo Fisher Scientific, 22-363-548) using a syringe plunger and washed with HBSS without Mg or Ca (Thermo Fisher Scientific, 14175095). Spleen tissue was incubated in 1 ml ACK lysis buffer (Gibco, A10492) for 5 min at 4 °C to lyse red blood cells, washed in HBSS, centrifuged at 500g and resuspended in PBS with 0.5% BSA (Sigma-Aldrich, A3294-100G) and 2 mM EDTA (VWR, E177-100ML) for downstream applications.
Lentivirus production for in vivo CRISPR–Cas9 deletion
We have previously described lentiviral backbones used perform Gfap promoter-driven genetic deletions in vivo26, which are derived from lentiCRISPRv2 (F. Zhang, Addgene plasmid 52961) and express Cas9 driven by the ABC1D gfa2 Gfap promoter67, whereas sgRNA is under U6 promoter control. To exchange 20 nucleotide-long sgRNA sequences to target different genes, we performed PCR-based restriction cloning using Phusion HF master mix (Thermo Fisher Scientific, F-548S) and the following primers: U6-PCR-F 5′-AAAGGCGCGCCGAGGGCCTATTT-3′, U6-PCR-R 5′-TTTTTTGGTCTCCCGGTGTTTCGTCCTTTCCAC-3′, crRNA-R 5′-GTTCCCTGCAGGAAAAAAGCACCGA-3′, crRNA-F 5′-AAAAAAGGTCTCTACCG(N)20GTTTTAGAGCTAGAAATAGCAAGTT-3′, where N20 marks the sgRNA substitution site. sgRNA design was performed using a combination of tools: Synthego (https://design.synthego.com//), CRISPick (SpyoCas9, Hsu (2013) tracrRNA, https://portals.broadinstitute.org/gppx/crispick/public) and the Sabatini/Lander list (Addgene 1000000096). The most upstream exon-targeting guide that was represented on the most sgRNA design platforms was picked. The sgRNA sequences used are as follows: NT sgRNA, 5′-GCACTACCAGAGCTAACTCA-3′; sgTnfsf10, 5′-ACTTCTCTCCACCCAGCTGC-3′; and sgIl11ra1, 5′-TTCATAGTCTACCGCTTGGC-3′. U6 and crRNA PCR products were purified using a QIAquick PCR Purification kit (Qiagen, 28104) and digested using the following restriction enzymes: DpnI (NEB, R0176S) and BsaI-HF (NEB, R3535/R3733). The U6 fragment and the pLenti backbone were also digested with AscI (NEB, R0558) and SbfI-HF (NEB, R3642). The crRNA fragment was additionally digested with SbfI-HF. Digested fragments were separated by gel electrophoresis on a 0.8% agarose gel (Life Technologies, 16500100) for 20 min at 135 V. DNA was extracted from gel fragments using a QIAquick Gel Extraction kit (Qiagen, 28704). Inserts were ligated into the lentivirus backbone overnight at 16 °C using a T4 DNA Ligase kit (NEB, M0202L). Finally, constructs were heat-shock-transformed into NEB Stable Esherichia coli (NEB, C3040) at 42 °C for 30 s and plated on ampicillin LB agar selection plates (MP Biomedicals, 113002221). After overnight incubation at 37 °C, single colonies were picked and plasmid DNA was isolated using a QIAprep Spin Miniprep kit (Qiagen, 27104). Successful sgRNA cloning was confirmed by Sanger sequencing (Azenta Life Science) of clones using the U6-PCR-F and crRNA-rev primers described above. DNA production was scaled up using a CompactPrep Maxi kit (Qiagen, 12863). Lentiviruses were prepared in HEK293FT cells following the instructions of ViraPower Lentiviral Packaging Mix (Thermo Fisher Scientific, K497500), with packaging and pseudotyping plasmids comprising pLP1 and pLP2, and pLP/VSVG, respectively. In brief, 5 × 106 HEK293FT cells cultured in antibiotic-free medium were transfected using Lipofectamine 2000 (Thermo Fisher Scientific, 11668019) complexes in Opti-MEM reduced serum medium+GlutaMAX (Thermo Fisher Scientific, 51985034). Overall, 3 µg of each helper plasmid and expression plasmid was used to form DNA–Lipofectamine complexes (12 µg DNA total, DNA to Lipofectamine ratio of 1:3), which were added dropwise to cell cultures. The following day, culture medium was replaced, and fresh medium was added. After 48 h, virus-containing supernatant was collected, filtered through 0.45 µm PVDF membranes and concentrated using Lenti-X Concentrator (Clontech, 631231) in accordance with the manufacturer’s protocol. Final pellets were resuspended in 1/500–1/1,000 of the original supernatant volume and aliquots stored at −80 °C. Lentiviral titration was performed using a qPCR Lentivirus Titration kit (ABM, LV900) with undiluted and 1:10–100 diluted samples.
Isolation, culture and stimulation of primary mouse astrocytes, microglia and oligodendrocytes
Primary mouse astrocytes were isolated from neonatal brains as previously described68. In brief, wild-type mouse pups aged 1–3 days were cryoanaesthetized and decapitated. Whole brains were extracted, and 2–3 brains pooled per culture. After centrifugation for 5 min at 500g, tissues were resuspended in 5 ml pre-warmed 0.25% Trypsin-EDTA (Thermo Fisher Scientific, 25200-072) and incubated at 37 °C for 10 min, followed by addition of culture medium to stop the dissociation reaction and then mechanical disaggregation by strong pipetting. The suspension was filtered through a 70 µm nylon strainer and centrifuged 5 min at 500g. Finally, the suspension was seeded onto T75 flasks (Falcon, 353136) that were coated at room temperature for 10 min with poly-l-lysine (Sigma-Aldrich, P4707) and washed with PBS. Mixed glial cultures were grown for at least 10 days before enrichment of astrocytes, microglia or oligodendrocytes per the following method. For isolation of microglia, mixed glial culture flasks were shaken at 150 r.p.m. and 37 °C for 30 min, and detached microglia were taken up from culture supernatants. New medium was added, and flasks were shaken at 180 r.p.m. overnight at 37 °C for isolation of oligodendrocyte progenitors (OPCs) as previously described69,70. The next day supernatants containing OPCs were incubated in Petri dishes for 20 min to remove remaining astrocytes or microglia and transferred to culture dishes coated with poly-d-lysine- (Thermo Fisher Scientific, A3890401) and ECM Gel (Sigma-Aldrich, E1270). OPCs were cultured in DMEM containing 1× B-27 supplement (Thermo Fisher Scientific, 17504044), 1× N2 supplement (Thermo Fisher Scientific, 17502-048), 1% penicillin–streptomycin (Thermo Fisher Scientific, 15140122), 1 mM sodium pyruvate (Thermo Fisher Scientific, 11360070), 50 ng ml–1 human NT3 (PeproTech, 450-03) and 10 ng ml–1 human PDGF-AA (PeproTech, 100-13A) for 1–4 days before differentiation to oligodendrocytes in DMEM containing 1% penicillin–streptomycin, 1 mM sodium pyruvate, 1× B-27 supplement, 1× N2 supplement, 50 ng ml–1 NT3, 20 ng ml–1 human CNTF (PeproTech, 450-13) and 40 ng ml–1 3,3′,5-triiodo-l-thyronine (Sigma-Aldrich, T5516) with no PDGF-AA for 4–7 days. Remaining mixed glia cultures were shaken at 240 r.p.m. for 2–4 h at 37 °C. Next, medium was washed out with PBS and 0.25% Trypsin-EDTA was used to detach astrocytes after incubation for around 5 min. Finally, astrocytes were taken up in culture medium and seeded in poly-l-lysine-coated experimental plates at 150 × 103 cells per well in 24-well plates. Culture medium used for astrocytes was made of DMEM/F12+GlutaMAX (Thermo Fisher Scientific, 10565-042) with 10% FBS (Gibco, 10-438-026), 1% penicillin–streptomycin (Thermo Fisher Scientific, 15140122), 100 mM HEPES (Gibco, 15630080) and 1× non-essential amino acids (Gibco, 11140050). Inhibition of STAT3 was performed using niclosamide (BAY2353) (Selleck Chemicals, S3030) at 2 µM in DMSO as vehicle (Sigma-Aldrich, 472301), as this concentration was reported to be effective but not cytotoxic71. Stimulation of astrocytes with tumour-conditioned medium or recombinant cytokines was performed overnight using 50 ng ml–1 mouse IL-11 (BioLegend, 756104), 100 ng ml–1 MIF (Novus Biologicals, NBP2-35190-10ug), 10 ng ml–1 EGF (PeproTech, 315-09), 100 ng ml–1 TGFα (PeproTech, 100-16A), 100 ng ml–1 TGFβ (R&D Systems, 7666-MB-005), 100 ng ml–1 IL-15 (PeproTech, 210-15) and 50 ng ml–1 IFN-γ (R&D Systems, 485-MI-100).
Isolation, culture and stimulation of primary mouse neurons
Primary neurons were isolated from mouse pups at 0–3 days of age based on previously published procedures72,73. In brief, brains were dissected into a Petri dish with HBSS (Thermo Fisher Scientific, 14175103) supplemented with 1 mM sodium pyruvate (Thermo Fisher Scientific, 11360070), 0.1% w/v glucose (Sigma-Aldrich, G-6152) in nuclease-free water (Thermo Fisher Scientific, AM9937) and 1% HEPES (Sigma-Aldrich, H-4034) followed by removal of meninges and the brainstem. Brains were then dissociated using a Worthington Papain Dissociation System (Worthington Biochemical, LK003150). In brief, albumin ovomucoid inhibitor was reconstituted in 32 ml EBSS, papain in 5 ml EBSS and DNase in 500 µl EBSS. Papain and DNase solutions were combined, and brain tissue was placed in 5.35 ml EBSS with papain and DNase, followed by 30 min of incubation at 37 °C. Tissue was mechanically dissociated by pipetting with a P1000, passed through a 70 µm strainer (Fisher Scientific, 22363548) and centrifuged at 200g for 5 min at room temperature. Cell pellets were resuspended in 2.7 ml EBSS with 0.3 ml reconstituted albumin ovomucoid inhibitor and 150 µl DNase in EBSS, then layered on top of 5 ml of ovomucoid inhibitor solution and centrifuged at 100g for 5 min at room temperature. Cells were taken up in MACS Neuro medium (Miltenyi Biotec, 130-093-570) supplemented with 2% MACS NeuroBrew-21 (Miltenyi Biotec, 130-093-566) and 1% penicillin–streptomycin (Thermo Fisher Scientific, 15140148) and plated at 1.5–2.5 × 105 cells per well in 24-well laminin-coated and poly-l-ornithine-coated culture plates (Westnet, 354659). Two days after plating, glia were eliminated by treating cultures for 48 h with 2 µM cytarabine (Millipore Sigma, PHR1787). Medium was then changed, and neuron cultures were used within 10–14 days after preparation.
In vitro co-culture of astrocytes and T cells
In vitro co-culture assays of astrocytes and T cells were performed in 24-well plates at a ratio of 1:2. In brief, 100 or 150 × 103 astrocytes per well were seeded 2 days before assay initiation and stimulated as detailed for around 16 h (overnight) before the addition of T cells. When blocking antibodies for TRAIL were used, they were added to astrocyte cultures for 1 h before T cell addition and in the co-culture medium. Blocking of IL-11 was performed during all of the overnight pre-stimulation step as well as during co-culture. Splenic T cells were isolated from female wild-type mice aged 8–10 weeks and purified by magnetic selection using Naive CD4 or CD8 T cell Isolation kits (Miltenyi Biotec, 130-104-453 or 130-096-543). Naive T cells were then activated by overnight stimulation with plate-bound anti-CD3 at 10 μg ml–1 (bioXcell, BE0001-1-A005mg) and 0.25 μg ml–1 anti-CD28 (bioXcell, BE0015-5-A005mg) at a density of 0.3 × 106 per well. The next day, activated T cells were collected, counted and resuspended in their respectively conditioned medium before being added onto astrocytes at a 1:2 ratio. T cell assays were performed in complete RPMI medium supplemented with 0.25 μg ml–1 anti-CD28 to maintain activation. After 3 h of co-cultivation, half of the culture medium was taken off for staining and measurement of caspase-8 cleavage using a Vybrant FAM Caspase-8 Assay kit (Thermo Fisher Scientific, V35119). After a total of 8 h, the rest of the culture supernatant containing T cells was taken off and stained to detect caspase-3/7 activation using a CellEvent Caspase-3/7 Green Flow Cytometry Assay kit (Thermo Fisher Scientific, C10740).
Functional blocking with monoclonal antibodies
We used the following monoclonal antibodies: TRAIL blocking antibody (Thermo Fisher Scientific, 16-5951-85) and its respective rat anti-IgG2aκ isotype control (Thermo Fisher Scientific, 16-4321-82) were used at 5 µg ml–1; human/mouse cross-reactive anti-IL11 blocking antibody (R&D Systems, MAB218) and its respective mouse anti-IgG2aκ Isotype control (Thermo Fisher Scientific, 14472485) were used at 10 µg ml–1.
Flow cytometry
For flow cytometry, single-cell suspensions were obtained as described above. A small aliquot (usually one-tenth of the sample) was taken after isolation from tissue and before staining and diluted 1:10–20 in PBS. This aliquot was run on the flow cytometer using the plate reader setting, which enables the analysis of defined sample volumes and therefore back calculation of absolute cell numbers per sample. The rest of the sample was first stained using a Live/Dead Fixable Aqua Dead Cell Stain kit for 405 nm excitation (Thermo Fisher Scientific, L34966) at 1:2,500 dilution and anti-CD16/CD32 Fc block (BD Biosciences, 553142), then washed in 0.5% BSA and 2 mM EDTA pH 8.0 in PBS (FACS buffer) and then stained extracellularly with a mix of fluorescently labelled antibodies in FACS buffer for 30 min in the dark at 4 °C, followed by another wash with FACS buffer. For intranuclear staining, cell pellets were fixed for 20 min at room temperature in fixation buffer with a FOXP3 transcription factor staining kit (eBioscience, 00-5523-00), followed by a washing step and staining for 30 min at room temperature in the provided wash buffer. Finally, samples were washed with FACS buffer and analysed on a BD FACSymphony Analyzer (BD Biosciences). Data were analysed using FlowJo (v10.9). The following antibodies were used for T cell analysis: BUV496 anti-SLAMF6 (BD Biosciences, 750046), BUV563 anti-CD11b (BD Biosciences, 741242), BUV661 anti-CD45 (BD Biosciences, 612975), BUV737 anti-PD-1 (BD Biosciences, 749306), BUV805 anti-LAG-3 (BD Biosciences, 748540), BUV650 anti-CD3 (BD Biosciences, 740530), BV711 anti-TIGIT (BD Biosciences, 744214), BV750 anti-TNF (BD Biosciences, 566365), BV786 anti-TIM3 (BD Biosciences, 747621), FITC anti-CD8a (BioLegend, 100804), PerCP-Vio 700 anti-DR5 (Miltenyi Biotec, 130-102-478), PE anti-CTLA4 (BioLegend, 106306) or BV421 anti-CTLA4 (BioLegend, 106312), PE/Dazzle 594 anti-IL-10 (BioLegend, 505034), PE/Cy5 anti-IL2 (BioLegend, 503824), PE/Cy7 anti-CD4 (BioLegend, 100422), AF700 anti-CD39 (Invitrogen, 56-0391-82) and APC/Cy7 anti-IFNγ (BD Biosciences, 561479). GL261-specific CD8+ T cells were stained with an APC-conjugated H-2D(b) tetramer loaded with GARC-1(77–85) peptide (AALLNKLYA) (NIH Tetramer Core). For detection of PE-conjugated HSV-1-specific CD8+ T cells, H-2Kb tetramer with HSV-1 glycoprotein B peptide SSIEFARL (Immudex, JD2670) was used. The following antibodies were used for the detection of apoptosis: BUV737 anti-CD11b (BD Biosciences, 612800), BV421 anti-CD11a (BD Biosciences, 740020), BV605 anti-CX3CR1 (BioLegend, 149027), BV650 anti-CD3 (BD Biosciences, 740530), BV711 anti-CD8a (BD Biosciences, 752634), BV785 anti-Ly6C (BioLegend, 128041), Cellevent Caspase-3/7 Flow Cytometry Assay kit (Thermo Fisher Scientific, C10740) as described below, PerCP-Vio 700 anti-DR5 (Miltenyi Biotec, 130-102-478), PE/Cy7 anti-CD4 (BioLegend, 100422), APC anti-CD45 (Thermo Fisher Scientific, 17-0451-83) and APC/Cy7 anti-CD49d (BioLegend, 103635, 1:20). The following antibodies were used for measuring TRAIL expression and pSTAT3 in TdTomato+ reporter cells: BUV661 anti-CD45 (BD Biosciences, 612975), BUV737 anti-CD11b (BD Biosciences, 612800), BV421 anti-CD11a (BD Biosciences, 740020), BV605 anti-CX3CR1 (BioLegend, 149027), BV785 anti-Ly6C (BioLegend, 128041), PE/Cy7 anti-TRAIL (BioLegend, 109311, 1:10), APC anti-pSTAT3 (Y705) (Thermo Fisher Scientific, MA5-36902, 1:25) and APC/Cy7 anti-CD49d (BioLegend, 103635, 1:20). The following antibodies were used for the detection of DCs: BUV661 anti-CD45 (BD Biosciences, 612975), BUV 563 anti-CD11c (BD Biosciences, 749040), BUV737 anti-CD11b (BD Biosciences, 612800), BV785 anti-I-A/I-E (BioLegend, 107645), BUV395 anti-F4/80 (BD Biosciences, 565614), BV570 anti-Ly6C (BioLegend, 128030), BV421 anti-CD103 (BD Biosciences, 562771), PE/Dazzle 594 anti-CD172a (BioLegend, 144015) and PerCP-Vio 700 anti-DR5 (Miltenyi Biotec, 130-102-478, 1:10) as described74. The following antibodies were used for the analysis of TAM phenotypes: BUV395 anti-I-A/I-E (BD Biosciences, 743876), BUV563 anti-CD86 (BD Biosciences, 741285), BUV661 anti-CD45 (BD Biosciences, 612975), BUV737 anti-CD11b (BD Biosciences, 612800), BV605 anti-CD80 (BD Biosciences, 563052), BV711 anti-CD163 (BioLegend, 155325), BV785 anti-Ly6C (BioLegend, 128041), PE/Cy7 anti-CD68 (BioLegend, 137015), PE anti-F4/80 (BioLegend, 123110), PE/Cy5 anti-CD11c (BioLegend, 117316), APC/Cy7 anti-CD49d (BioLegend, 103635, 1:20), BV421 anti-CD11a (BD Biosciences, 740020), AF700 anti-CD39 (Thermo Fisher Scientific, 56-0391-82) and PerCP-Vio 700 anti-DR5 (Miltenyi Biotec, 130-102-478). All antibodies were used at 1:100 dilution unless otherwise specified.
FACS
Single-cell suspensions were stained with a mix of antibodies in FACS buffer for 30 min at 4 °C as described above. All antibodies were diluted 1:100 in FACS buffer. Before sorting, cell suspensions were filtered through tubes with 35 µm cell strainer caps (Falcon, 352235). The following antibodies were used for sorting: BV421 anti-CD11a (BD Biosciences, 740020), BV605 anti-CX3CR1 (BioLegend, 149027), BV650 anti-CD11b (BD Biosciences, 563402), BV711 anti-CD8a (BD Biosciences, 752634), BV785 anti-Ly6C (BioLegend, 128041), FITC anti-CD3 (BioLegend, 100204), PE/Cy7 anti-CD4 (BioLegend, 100422), APC anti-CD45 (Thermo Fisher Scientific, 17-0451-83) and APC/Cy7 anti-CD49d (BioLegend, 103635, 1:20). CD4+ T cells were sorted as live+ CD45+CD11b–CD3+CD4+. CD8+ T cells were sorted as live+ CD45+CD11b−CD3+CD8+ cells. Microglia were sorted as live+ CD45+CD11b+Ly6Clow/–CX3CR1+CD49dlowCD11alow. Monocytes were sorted as CD45+CD11b+Ly6ChighCD49dhighCD11ahigh. Cells were sorted using a FACSAria IIU (BD Biosciences). Sorting from Aldh1l1cre-ERT2/TdTomato reporter mice was performed by gating TdTomato+ cells against wild-type animals.
Flow cytometry analysis of T cells
For flow cytometry of T cells, magnetic separation was used to negatively select CNS-infiltrating T cells with a Pan T-cell Isolation Kit II mouse (Miltenyi Biotec, 130-095-130). T cells were then reactivated in a complete RPMI containing 50 ng ml–1 phorbol 12-myristate 13-acetate (PMA) (Sigma Aldrich, P8139), 1 μM ionomycin (Sigma Aldrich, I3909-1ML), 1:1,500 each of GolgiStop and GolgiPlug (BD Biosciences, 554724) and 1:100 BUV395 anti- LAMP1 (BD Biosciences, 565533) for 4 h at 37 °C. T cell culture medium was composed of RPMI 1640 (Gibco, 11875119) supplemented 10% FBS, 1% penicillin–streptomycin, 50 µM 2-metcaptoethanol (Sigma-Aldrich, M6250), 1% non-essential amino acids (Thermo Fisher Scientific, 11140050), 10 mM HEPES (Thermo Fisher Scientific, 15630130), 1 mM sodium pyruvate (Thermo Fisher Scientific, 11360070) and 1× GlutaMAX (Thermo Fisher Scientific, 35050061). T cells were then incubated with 50 nM dasatinib (Selleck Chemicals, S1021) to improve tetramer staining and 1:200 FcBlock (BD Biosciences, 553142) for 20 min at 37 °C before the addition of fluorescently labelled MHC tetramers at a final dilution of 1:50 in 2% FBS for staining at room temperature for 1 h in the dark. Washing steps were performed with FACS buffer and centrifugation at 600g for 5 min at 4 °C. Surface markers were stained in FACS buffer for 30 min at 4 °C, followed by fixation and permeabilization with BD Cytofix/Cytoperm (BD Biosciences, 554715) for 20 min at 4 °C according to the supplied protocol. Intracellular markers were then stained with an antibody solution prepared in permeabilization buffer. Stained T cells were stored at 4 °C in FACS buffer until analysis.
Apoptosis analysis
Apoptosis was analysed by flow cytometry using a Cellevent Caspase-3/7 Flow Cytometry Assay kit (Thermo Fisher Scientific, C10740), staining samples with caspase-3/7 detection agent for 30 min at 37 °C. To ensure consistent staining, Sytox AADvanced Dead Cell Stain was added to each tube exactly 1 min before measurement by flow cytometry. To measure death-receptor-induced apoptosis, a Vybrant FAM Caspase-8 Assay kit (Thermo Fisher Scientific, V35119) was used per the manufacturer’s protocol.
RNA extraction and real-time qPCR
For quantitative analysis of gene transcription from cultured cells, cellular RNA was extracted using a RNeasy kit (Qiagen, 74004). In brief, cells were lysed in 350 µl RLT buffer and stored at −80 °C until extraction following the manufacturer’s instructions. RNA was transcribed to cDNA using a High-Capacity cDNA Reverse Transcription kit (Life Technologies, 4368813). Quantitative measurement of gene expression was performed by qPCR using Taqman Fast Universal PCR master mix (Life Technologies, 4367846), MicroAmp Optical 384-well plates (Thermo Fisher Scientific, 43-098-49) and a ViiA 7 Real-Time PCR system (Applied Biosciences). The following Taqman probes were used: ActB (Mm02619580_g1), Tnfsf10 (Mm01283606_m1), Tox (Mm00455231_m1), Il11 (Mm00434162_m1), Cyp1b1 (Mm00487229_m1), IL11 (Hs01055414_m1) and CYP1B1 (Hs00164383_m1). qPCR data were analysed using the ΔΔCt method by normalizing the expression of each gene for each replicate by the average of the housekeeping gene (ActB) and then normalizing the treatment group to the control group to determine the relative change. For in vivo quantification of transgene expression, minced brain tissue was homogenized using BeadBug homogenizer tubes filled with 2.8 mm steel balls (Sigma, Z763829) and using an AllPrep DNA/RNA Mini kit (Qiagen, 80204). In brief, 600 µl RLT Plus supplemented with 40 mM dithiothreitol (Sigma-Aldrich, 646563-10X.5ML) was used per 30 mg of tissue. Mechanical dissociation was performed on a PowerLyzer 24 with two 20-s cycles at 2,500 r.p.m. and 30 s of dwell time. Tubes were centrifuged on a countertop centrifuge at 13,000g for 5 min. An AllPrep DNA/RNA Mini kit was used to isolate RNA following the manufacturer’s instructions, with the addition of DNAse I treatment of RNA columns to eliminate genomic DNA (Qiagen, 79254). RNA was transcribed to cDNA as described above. Samples were normalized to 25 ng µl–1 cDNA and anti-TRAIL monoclonal antibody expression was quantified by qPCR as follows. For transgene amplification the following primer pairs and probe pairs were used: anti-TRAIL_fwd: TCTGTCTGCTTCTGTTGGCG, anti-TRAIL_rev: GGCTTCCCCCAATTTCTGTTG, anti-TRAIL_probe: AGCGAGTGACACTCTCCTGTCG with 5′ FAM as reporter and 3′ IowaBlack FQ as quencher (Integrated DNA Technologies). Expression levels were normalized by subtracting Ct values of housekeeping gene (ActB; Thermo Fisher Scientific, Mm02619580_g1) and expression calculated using the formula \({2}^{-{C}_{{\rm{t}}}{\rm{normalized}}}\).
Bioinformatic RNA-seq analysis
For bulk RNA-seq, the raw fastq files are processed using Kallisto75, which aligned the read pairs to the GRCm38 reference genome and quantified the gene expression of each gene based on Gencode (v.M37)76. The tximport77 package was used to organize the output from Kallisto, and differential expression analysis was performed using DESeq2 (v.4.3). For scRNA-seq analysis, sequenced reads from pooled samples were trimmed and demultiplexed using Trim Galore (v.0.6.5), assigned to their associated cell barcodes and unique molecular identifiers (UMIs) and aligned to the ENSEMBL reference genomes for mouse and human. Drop-seq software was used to correct bead synthesis errors and to quantify gene expression62. The Seurat package (v.4.3) was used to normalize count matrices, remove doublets, correct batch effects and integrate samples into one dataset. The SCTransform function was used to normalize and scale expression matrices, stabilize variance, regress out variance due to mitochondrial genes and identify the 2,000 most variable genes. Next, principal component analysis was performed to identify the parameters for dimensionality reduction. For clustering, we used the Louvain algorithm with multilevel refinement. We applied the MAST algorithm for differential expression analysis between clusters as part of the FindAllMarkers function of Seurat. Differential expression analyses between specific groups were performed with FindMarkers. For pathway analysis, GSEA (Broad Institute, v.4.3.2) was run using a pre-ranked list with the average expression log2[fold change] as the ranking metric. Curated gene signatures from the Molecular Signatures Database (MSigDB; v.2023.1) were used for the identification of relevant pathways. We show analysis of Hallmark gene sets (h.all) and a large collection of curated gene sets (c2.all), which can be found on the MSigDB website. IPA (Qiagen) was performed using gene identifiers and the average expression log2[fold change], filtering to include 2,000–3,000 genes at most. We performed core analysis and made use of pathway enrichment and upstream regulator tools. Any signature scores shown were calculated using the AddModuleScore function in Seurat. Mouse astrocytes were identified using a canonical marker expression score (Slc1a2, Slc1a3, Aldoc, Gja1, Aldh1l1, S100b and Vim) above 0. Differential abundance testing for mouse scRNA-seq data was conducted using miloR78 (v.1.8.1), constructing a k-nearest neighbours graph and assigning cells to a neighbourhood (k = 20, d = 9). Differential abundance testing was performed in a generalized linear model framework between tumour and control samples using testNhoods (robust = TRUE, fdr.weighting = “graph-overlap”). Grouping of neighbourhoods was performed using the groupNhoods function (overlap=1, max.lfc.delta=5). Groups were then assigned to scRNA-seq clusters and the log fold change were plotted for each neighbourhood and cluster. For TCGA bulk RNA-seq cell-type deconvolution, the gene expression data of TCGA-glioblastoma was downloaded from the GDC Data Portal (National Cancer Institute) and the de-convolution algorithm BayesPrism was used to extract cell-type-specific expression profiles from each sample79. For imputation of TRAIL expression, the CollecTri database of the transcriptional regulatory networks and the decoupleR package80 were used. For visualization, Seurat, dittoSeq (v.4.3) or ggplot2 (v.3.4.3) was used. Volcano plots were made using EnhancedVolcano (v.3.17). replotGSEA function was also used (https://github.com/PeeperLab). CellChat was used to predict cell–cell interactions81. All R-based analyses were performed using RStudio (v.4.3.0) with R (v.4.3.1), and final figures were edited using Adobe Illustrator (v.29.4).
ELISA
Protein-level detection of mouse or human IL-11 in TCM was performed using ELISA with a Mouse IL11 ELISA kit (Thermo Fisher Scientific, EMIL11) or a Human IL11 ELISA kit (Thermo Fisher Scientific, EHIL11), respectively, following the manufacturer’s protocol and read out using a GloMax Explorer Multimode microplate reader (Promega) at 450 nm. For detection of IL-11 in ex vivo tumour tissues, mice were intracardially perfused as described above and tumour tissue was dissected, weighed and minced, followed by enzymatic digestion with papain and collagenase D as described above, except for the addition of 1× Halt Protease and phosphatase inhibitor cocktail (Thermo Fisher Scientific, 78440). Following digestion, samples were centrifuged at 10,000g and supernatants separated for ELISA.
Glioblastoma survival analysis
Glioblastoma survival was analysed using the cBioPortal dataset glioblastoma multiforme (TCGA, Firehose Legacy), and patient survival data were downloaded for 166 samples with transcriptional data available (mRNA expression z scores relative to all samples (log RNA Seq V2 RSEM). Statistical analysis and survival curves were produced in R using the survminer package. In addition, the Gepia2 database was used for survival analysis based on gene or signature expression in the ‘glioblastoma’ dataset and to detect correlation with similarly expressed genes82, which were classified using PantherDB.
Human primary astrocyte culture
Primary human fetal astrocytes were isolated as previously described83. Second trimester fetal brain tissues were obtained from Centre Hospitalier Universitaire Sainte-Justine or from the Birth Defects Research Laboratory, University of Washington with maternal written consent and under local ethic boards’ approval. Isolation of glial cells was carried out through mechanical and chemical digestion. Primary fetal astrocytes were maintained in DMEM/F12 (Thermo Fisher Scientific, 11320033) with 10% FBS (Wisent Bio Products, 080-450), 1% penicillin–streptomycin (Thermo Fisher Scientific, 15140122) and 1% GlutaMAX (Thermo Fisher Scientific, 35050061) and cultured at 37 °C with 5% CO2. For in vitro stimulation conditions, human primary astrocytes were seeded at 80,000 cells per well. The cell medium was aspirated, cells were washed with 37 °C PBS and the fresh, appropriately conditioned medium was then added into wells. Human astrocytes were stimulated for 6 h with U87 TCM (see above) and 50 ng ml–1 recombinant human IL-11 (PeproTech, 200-11). At the end of the treatment, total RNA was isolated and transcribed and analysed by qPCR as described above. In experiments involving primary patient-derived glioblastoma cell lines, commercially acquired human astrocytes (ScienCell, 1800) cultured in DMEM and 10% FBS were used.
Immunofluorescence staining of tissue sections
Following perfusion with ice-cold PBS, mouse brains were dissected and fixed in 4% paraformaldehyde solution (Electron Microscopy Sciences, 15714S) at 4 °C overnight, then dehydrated in 30% sucrose (Sigma-Aldrich, S0389) for 1 week. Fixed brains were then grossly dissected to embed in Tissue-Tek OCT compound (Sakura Finetek USA, 4583) and stored at −80 °C. Tissue blocks were cut on a Leica cryostat CM1950 (Leica) at 8 µm and mounted on Tissue Path Superfrost Plus Gold slides (Thermo Fisher Scientific, 15-188-48) and frozen at −20 °C. Immunofluorescence staining was performed by permeabilization with 1× BD Perm/Wash buffer (BD Biosciences, 554723) for 10 min at room temperature, followed by blocking of nonspecific binding with serum-free protein block (Agilent, X090930-2) for 10 min at room temperature. Primary and secondary antibodies were diluted in BD Perm/Wash buffer and tissue slides incubated with primary staining solution at 4 °C overnight, washed with BD Perm/Wash buffer 3 times and then incubated with secondary staining solution for 1 h. After staining, ProLong Gold Antifade mountant (Thermo Fisher Scientific, P36930) was added onto the tissue, and the section was sealed with High Precision 1.5H cover slips (Azer Scientific, ES0107052) and nail polish (Electron Microscopy Sciences, 72180). For pSTAT3 staining, tissue permeabilization was performed with 0.5% Triton-X (Sigma-Aldrich, T8787-100ML) and 4% BSA in PBS for 1 h at room temperature. Primary antibody staining was performed in the same solution overnight at room temperature, followed by washing in 1× BD Perm/Wash buffer, secondary staining for 1 h at room temperature in 1× BD Perm/Wash buffer, followed by DAPI (Sigma-Aldrich, D9542-1MG) staining for 5 min. Mounting and slide sealing were performed similarly for all stainings. Confocal fluorescence microscopy was performed using a Zeiss LSM710 (Zeiss) at ×20 magnification. The following antibodies were used: mouse anti-GFAP-AF488 (eBioscience, 53-9892-82), chicken anti-GFAP (Abcam, ab4674), donkey anti-chicken-AF405 (Jackson ImmunoResearch, 703-475-155), rabbit anti-pSTAT3 (Cell Signaling Technology, 9145 T), donkey anti-rabbit-AF647 (Thermo Fisher Scientific, A31573), goat anti-TRAIL (Invitrogen, PA5-47073), donkey anti-goat-AF568 (Thermo Fisher Scientific, A11057), rat anti-CD3-FITC (eBioscience, 11-0032-80), rabbit anti-cleaved caspase-3 (Asp175) (Cell Signaling Technology, 9661T), rabbit anti-IL11RA1 (Abcam, ab125015) and rat anti-CD68 (Bio-Rad, MCA1957T). HSV-infected glioblastoma lines were imaged for GFP fluorescence using a Leica DMi8 Widefield microscope (Leica).
Multiplex immunofluorescence human glioblastoma
Multiplexed immunofluorescence and immunohistochemistry labelling were performed similarly to a previously described iterative indirect immunofluorescence imaging (4i) protocol84. In brief, formalin-fixed paraffin-embedded tissue sections of 2 µm thickness were mounted on adhesive glass slides in an adaptable chamber system (Ibidi sticky slides, Gräfelfing) before deparaffinization and heat-induced antigen retrieval at pH 9. As described previously84, each immunofluorescence imaging cycle consisted of blocking, indirect immunofluorescence staining and DAPI staining (Carl Roth), imaging on a Keyence BZ- X810 compact fluorescence microscope (Keyence) in imaging buffer and antibody elution. After five iterations, subsequent immunohistochemistry for EGFRvIII and GFAP and haematoxylin counterstaining were appended (adhering to standard operating procedures of the Institute of Neuropathology in Freiburg) on the same sections, and stepwise imaging was performed in brightfield modus at an identical location on the same microscope. Images of immunofluorescence labelling and immunohistochemistry labelling (with inverted greyscale intensity) were aligned and integrated on the basis of nuclear staining in DAPI and haematoxylin. This approach resulted in a 15-plexed source file for downstream analysis of each sample, created in Image J (v.1.53t with Java 1.8.0_345 (64-bit)). The following antibodies were used for the 6 cycles: cycle 1, GFAP (Dako, Z0334), CD3 (Bio-Rad, MCA1477T) and DR4 (R&D Systems, MAB347); cycle 2, IL-11 (LSBio, LS-C177617-50), OLIG2 (Biotechne, AF2418) and DR5 (R&D Systems, MAB6311); cycle 3, IL11RA (Abcam, ab125015), TRAIL (R&D Systems, AF375) and IBA1 (Synaptic Systems, 234 308); cycle 4, pSTAT3 (Cell Signaling Technology, 9145); cycle 5, cleaved caspase-3 (Cell Signaling Technology, 9664); and cycle 6, EGFRvIII (Biozol, GTX02628).
Immunofluorescence image analysis
Image analysis was conducted using the QuPath application (v.0.3.2 with Java 16.0.2). Analysis of multiplexed immunofluorescence images of human glioblastoma are described first. In brief, comprehensive training of automated analysis tools was carried out across samples. First, regions of interest were restricted to parenchymal or tumoural regions (including endothelial structures but excluding erythrocyte-filled luminal areas and other areas of clearly nonspecific antibody binding using a pixel classifier). Next, automated segmentation was performed using the ‘Watershed cell detection’ algorithm referring to the DAPI nucleus signal. Automated cell-type classification was performed using the ‘train object classifier’ algorithm, whereby positive or negative cells were manually selected until the accuracy of the classifier was sufficient. GFAP was used to identify tumour cells and astrocytes, IBA1 for myeloid cells and CD3 for T cells. To identify TRAIL+ astrocytes, we additionally trained an object classifier to detect cells positive for GFAP and TRAIL within non-malignant regions determined by histopathology. Apoptotic T cells were classified by training an object classifier for cells positive for CD3 and cleaved caspase-3. The distance to the nearest TRAIL+ astrocyte was calculated using the ‘detect centroid distances 2D’ function in QuPath. The number of specific detections was then divided by the surface area of the analysed image (after exclusion of nonspecific signals by pixel classification). Detection measurements and calculated distances were exported and analysed in R. For analysis of immunofluorescence images of mouse brain, we used the ‘cell detection’ and ‘train object classifier’ algorithms and quantified the number of detections over analysed area. Data points are biological replicates, except for distance measurements, for which each detected cell was plotted from n = 3 glioblastoma specimens from patients.
Proximity ligation assay
Proximity ligation assay (PLA) was performed using a Duolink In Situ Red Starter Kit Goat/Rabbit (Sigma-Aldrich, DUO92105-1KT) as previously described85. In brief, primary mouse astrocytes were isolated as described above. Splenic T cells were isolated by magnetic separation with a Pan T cell isolation kit II, mouse (Miltenyi Biotec, 130-095-130) and activated with anti-CD3 and anti-CD28 as described above. Around 5 × 104 astrocytes were plated in poly-l-lysine-coated chamber culture slides (Celltreat, 229168) 1 day before overnight stimulation with control or GL261 TCM. Culture medium was washed out and 2 × 105 activated T cells were added to the co-cultures for 4 h. Medium was carefully aspirated and cells were fixed in 4% paraformaldehyde for 20 min at room temperature, followed by two washes with PBS and permeabilized with 100% methanol (Sigma-Aldrich, 34860-4L-R) at −20 °C for 30 min. Samples were blocked with Duolink blocking solution for 1 h at room temperature, then stained with primary antibodies goat anti-TRAIL (Invitrogen, PA5-47073) and rabbit anti-DR5 (Abcam, ab8416) overnight at 4 °C; normal rabbit IgG was used as isotype control (Cell Signaling Technology, 2729S). Samples were washed with 5% BSA, then stained with PLA probe-conjugated secondary antibodies for 1 h at 37 °C, followed by washing twice with 1× Duolink Buffer A. Ligation was performed as instructed by the Duolink kit for 30 min at 37 °C, followed by two washes with buffer A, and ligation as instructed by the kit for 100 min at 37 °C. After final washes in 1× Duolink buffer B twice, and 0.01× Duolink buffer B, cells were stained with 1 µg ml–1 DAPI (Sigma-Aldrich, D9542) in 0.01× buffer B. Slides were mounted with ProLong Gold Antifade mounting medium (Fisher Scientific, P36930) and sealed using a coverglass (Fisher Scientific, 12-548-5M) and nail polish (Electron Microscopy Sciences, 72180) and imaged using Zeiss LSM880 confocal microscope (Zeiss, LSM880) at ×10 magnification.
Promoter activity assay
Human TRAIL promoter activity was assayed using a Secrete-Pair Gaussia Luciferase Assay (Genecopoeia, LF062). In brief, HepG2 cells were transiently transfected with a human TNFSF10 promoter reporter plasmid (Genecopoeia, TRAIL-HPRM37867-PG02) by use of Lipofectamine 3000 (Thermo Fisher Scientific, L3000008) at a DNA to Lipofectamine ratio of 1 µg: 1.5 µl. At 24 h after transfection, cells were stimulated with U87 TCM or recombinant human IL-11 (PeproTech, 200-11) for 24 h. Supernatants were collected, and any cell debris removed by centrifuging at 500g for 5 min. Luciferase activity assays were performed following the manufacturer’s protocol and analysed using a GloMax Explorer Multimode microplate reader (Promega). High-sensitivity buffer was used to detect the effect of IL-11 on TRAIL expression, whereas U87 TCM was measured using the stable detection buffer.
Analysis of calcium signalling
To analyse calcium signalling in astrocytes, the Ca2+ indicator Fluo-4 AM (Thermo Fisher Scientific, F14201) was used. In brief, primary mouse astrocytes were plated 2 days before measurement at a density of 105 per well as described above. On the day of measurement, Fluo-4-AM was dissolved in DMSO, dispersed with an equal volume of 20% w/v Pluronic F-127 (Sigma-Aldrich, P2443) and diluted to 4 µM in DMEM. Cells were washed with HBSS and incubated in Fluo-4-AM loading solution for 1 h at 37 °C, then washed again with HBSS and left to rest for 30 min at 37 °C to allow for de-esterification. First, a baseline signal pre-stimulation was recorded as described below. Next, astrocytes were stimulated with control growth medium, growth medium with 2 µM ionomycin as a positive control or with GL261 TCM. All media used during measurement were supplemented with 1.8 mM CaCl2 to provide extracellular calcium, and samples were measured immediately after stimulation. Signals were read out every 6 s for 240 s using a GloMax Explorer Multimode microplate reader (Promega) at 475 nm excitation and 495/505 detection and normalized to the baseline signal.
NFAT translocation analysis
To measure nuclear translocation of NFAT2, naive splenic CD4 T cells were isolated by magnetic selection as described above and activated with 10 µg ml–1 plate-bound anti-CD3 and 0.25 µg ml–1 soluble anti-CD28, with or without 10 µg ml–1 plate-bound recombinant TRAIL (R&D Systems, 1121-TL-010) in complete RPMI for 30 min. T cells were then stained with FITC anti-CD3 (1:100, Thermo Fisher Scientific, 11-0032-82), followed by fixation and permeabilization with eBioscience FOXP3/Transcription Factor Staining buffer set (Thermo Fisher Scientific, 00-5523-00) following the manufacturer’s instructions and intracellular staining with PE NFATc1 (1:50, BioLegend, 649606) and DAPI (Sigma-Aldrich, D9542) for 1 h. Around 5,000 cells of each condition were measured on an Imagestream X Mark II (Cytek Biosciences), and the Similarity function in IDEAS software (Amnis, v.6.2) was used to calculate the overlap between DAPI and NFAT2 signals.
Generation of HSV-1(anti-TRAIL)
Anti-TRAIL mouse monoclonal antibody (clone: 75411.11; VWR) was sequenced by liquid chromatography and tandem mass spectrometry at Rapid Novor (Kitchener). VH and VL regions were assembled in a scFv format fused to a mIgG1 Fc domain to generate the TRAIL-blocking single-chain antibody. Sequence of the VH region: QVTLKESGPGILQPSQTLSLTCTFSGFSLTTHGMGVGWIRQPAGKGLEWLANIWWDDDKFHSPSLKNRLTISKDTSNNQAFLRITNVDTADAAIYYCARISWERAYYFDYWGQGVMVTVSSAETTAPSVYPLAPGTALKSNSMVTLGCLVKGYFPEPVTVTWNSGALSSGVHTFPAVLQSGLYTLTSSVTVPSSTWSSQAVTCNVAHPASSTKVDKKIVPRECNPCGCTGSEVSSVFIFPPKTKDVLTITLTPKVTCVVVDISQNDPEVRFSWFIDDVEVHTAQTHAPEKQSNSTLRSVSELPIVHRDWLNGKTFKCKVNSGAFPAPIEKSISKPEGTPRGPQVYTMAPPKEEMTQSQVSITCMVKGFYPPDIYTEWKMNGQPQENYKNTPPTMDTDGSYFLYSKLNVKKETWQQGNTFTCSVLHEGLHNHHTEKSLSHSPG; sequence of the VL region: DVQMTQSPSFLSASVGERVTLSCRASQNVDTYLDWYQQKLGEAPKLLIYNTNNLHTGIPSRFSGSGSGTDYTLTISSLQPEDVATYFCLQRNRWLTFGSGTKLEIKRADAAPTVSIFPPSMEQLTSGGATVVCFVNNFYPRDISVKWKIDGSEQRDGVLDSVTDQDSKDSTYSMSSTLSLTKVEYERHNLYTCEVVHKTSSSPVVKSFNRNEC. The expression construct encoding the single-chain antibody was cloned between the CAG promoter and the rabbit β-globin pA and inserted using gateway recombination in the UL3/UL4 intergenic region of ONCR-142 DNA in a bacterial artificial chromosome (BAC). The excision of the BAC vector sequence was achieved by Cre–loxP recombination. BAC-removed viruses were plaque-purified in the Vero-SF cell line before expansion, purification by centrifugation, determination of titre and aliquoting into vials.
Oncolytic virus therapy
All virus stocks were thawed and prepared immediately before dosing and kept on ice until administration. For survival and efficacy studies, treatment and control groups were randomized on the basis of the tumour bioluminescence signal at day 6 after GL261–mNectin1 tumour implantation such that both groups displayed similar average signals. On day 7, mice were intratumourally injected with 3 × 106 p.f.u. HSV-1. Animals were monitored daily for clinical signs and body weight, and given dietary support as needed. Mice were euthanized in accordance with experimentation protocols once a body weight decrease of 20% occurred. Survival differences were calculated by log-rank (Mantel–Cox) test.
Spatial transcriptomic re-analysis
10x Visium spatial transcriptomic data from human glioblastoma were analysed using the SPATA2 R package86. Spatially weighted Pearson correlation analysis of our pathways or genes of interest were calculated for all tumour samples. For Ivy Glioblastoma Atlas Project data analysis, expression data for genes of interest were downloaded using the ivygapSE package (v.1.26) and replotted using R. Sample W20-2-1-L.03 was excluded because of its TNFRSF10B expression being identified as an extreme outlier using the identify_outliers function in the rstatix package (v.0.7.2).
Statistical analysis
GraphPad Prism 10 (GraphPad Software, v.10.0.2) was used for most bar graph visualizations and to calculate significant differences. Sample sizes (n) denote biological replicates and not technical repeated measurements of the same sample. For in vitro studies, replicates indicate independent biological samples randomly allocated to experimental groups. For in vivo studies, replicates indicate individual mice on a C57Bl/6J background allocated randomly to experimental groups. No statistical methods were used to predetermine sample sizes. Sample sizes were chosen on the basis of previous studies9,25. Difference testing between groups was done using a two-sided Student’s t-test or a one-way ANOVA followed by Tukey’s post hoc test as indicated in the figure legends for each experiment. For mouse survival studies, Kaplan–Meier curves are shown, and the log-rank (Mantel–Cox) test was performed to assess significance of survival differences between groups. A P value < 0.05 was considered significant. Statistical significance was indicated as follows: NS, P > 0.05; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Sequencing data have been deposited into the Gene Expression Omnibus under the accession number GSE246293. Gepia2 is available at http://gepia2.cancer-pku.cn/. CPTAC-3 data87 are accessible from https://www.cbioportal.org/. TCGA/GTEx survival data are available from http://gepia2.cancer-pku.cn/. Protein classification is available from https://pantherdb.org/. Gene ontology pathway analysis is available from https://amigo.geneontology.org/amigo/search/ontology. Source data are provided with this paper.
References
Andersen, B. M. et al. Glial and myeloid heterogeneity in the brain tumour microenvironment. Nat. Rev. Cancer 21, 786–802 (2021).
van den Bent, M. J. et al. Primary brain tumours in adults. Lancet 402, 1564–1579 (2023).
Zhao, J. et al. Immune and genomic correlates of response to anti-PD-1 immunotherapy in glioblastoma. Nat. Med. 25, 462–469 (2019).
Neftel, C. et al. An integrative model of cellular states, plasticity, and genetics for glioblastoma. Cell 178, 835–849 (2019).
Patel, A. P. et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 344, 1396–1401 (2014).
Chen, Z. et al. A paracrine circuit of IL-1β/IL-1R1 between myeloid and tumor cells drives genotype-dependent glioblastoma progression. J. Clin. Invest. 133, e163802 (2023).
Friebel, E. et al. Single-cell mapping of human brain cancer reveals tumor-specific instruction of tissue-invading leukocytes. Cell 181, 1626–1642 (2020).
Karimi, E. et al. Single-cell spatial immune landscapes of primary and metastatic brain tumours. Nature 614, 555–563 (2023).
Takenaka, M. C. et al. Control of tumor-associated macrophages and T cells in glioblastoma via AHR and CD39. Nat. Neurosci. 22, 729–740 (2019).
Maas, R. R. et al. The local microenvironment drives activation of neutrophils in human brain tumors. Cell 186, 4546–4566 (2023).
Khakh, B. S. & Deneen, B. The emerging nature of astrocyte diversity. Annu. Rev. Neurosci. 42, 187–207 (2019).
Heiland, D. H. et al. Tumor-associated reactive astrocytes aid the evolution of immunosuppressive environment in glioblastoma. Nat. Commun. 10, 2541 (2019).
Kim, J.-K. et al. Tumoral RANKL activates astrocytes that promote glioma cell invasion through cytokine signaling. Cancer Lett. 353, 194–200 (2014).
Loeffler, S., Fayard, B., Weis, J. & Weissenberger, J. Interleukin-6 induces transcriptional activation of vascular endothelial growth factor (VEGF) in astrocytesin vivo and regulates VEGF promoter activity in glioblastoma cells via direct interaction between STAT3 and Sp1. Int. J. Cancer 115, 202–213 (2005).
Mega, A. et al. Astrocytes enhance glioblastoma growth. Glia 68, 316–327 (2020).
Okolie, O. et al. Reactive astrocytes potentiate tumor aggressiveness in a murine glioma resection and recurrence model. Neuro-Oncology 18, 1622–1633 (2016).
Perelroizen, R. et al. Astrocyte immunometabolic regulation of the tumour microenvironment drives glioblastoma pathogenicity. Brain 145, 3288–3307 (2022).
Priego, N. et al. STAT3 labels a subpopulation of reactive astrocytes required for brain metastasis. Nat. Med. 24, 1024–1035 (2018).
Wurm, J. et al. Astrogliosis releases pro-oncogenic chitinase 3-like 1 causing MAPK signaling in glioblastoma. Cancers 11, 1437 (2019).
Lee, H. G., Wheeler, M. A. & Quintana, F. J. Function and therapeutic value of astrocytes in neurological diseases. Nat. Rev. Drug Discov. 21, 339–358 (2022).
Placone, A. L., Quinones-Hinojosa, A. & Searson, P. C. The role of astrocytes in the progression of brain cancer: complicating the picture of the tumor microenvironment. Tumour Biol. 37, 61–69 (2016).
Couturier, C. P. et al. Glioblastoma scRNA-seq shows treatment-induced, immune-dependent increase in mesenchymal cancer cells and structural variants in distal neural stem cells. Neuro-Oncology https://doi.org/10.1093/neuonc/noac085 (2022).
Rosberg, R. et al. Hypoxia-induced complement component 3 promotes aggressive tumor growth in the glioblastoma microenvironment. JCI Insight 9, e179854 (2024).
Janssen, E. M. et al. CD4+ T-cell help controls CD8+ T-cell memory via TRAIL-mediated activation-induced cell death. Nature 434, 88–93 (2005).
Sanmarco, L. M. et al. Gut-licensed IFNγ+ NK cells drive LAMP1+TRAIL+ anti-inflammatory astrocytes. Nature 590, 473–479 (2021).
Wheeler, M. A. et al. MAFG-driven astrocytes promote CNS inflammation. Nature 578, 593–599 (2020).
McLane, L. M., Abdel-Hakeem, M. S. & Wherry, E. J. CD8 T cell exhaustion during chronic viral infection and cancer. Annu. Rev. Immunol. 37, 457–495 (2019).
Khan, O. et al. TOX transcriptionally and epigenetically programs CD8+ T cell exhaustion. Nature 571, 211–218 (2019).
Chang, C. H. et al. Posttranscriptional control of T cell effector function by aerobic glycolysis. Cell 153, 1239–1251 (2013).
Lehnert, C. et al. TRAIL-receptor costimulation inhibits proximal TCR signaling and suppresses human T cell activation and proliferation. J. Immunol. 193, 4021–4031 (2014).
Cook, S. A. & Schafer, S. Hiding in plain sight: interleukin-11 emerges as a master regulator of fibrosis, tissue integrity, and stromal inflammation. Annu. Rev. Med. 71, 263–276 (2020).
Greenwald, A. C. et al. Integrative spatial analysis reveals a multi-layered organization of glioblastoma. Cell 187, 2485–2501 (2024).
Opitz, C. A. et al. An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature 478, 197–203 (2011).
Sadik, A. et al. IL4I1 is a metabolic immune checkpoint that activates the AHR and promotes tumor progression. Cell 182, 1252–1270 (2020).
Ma, R., Li, Z., Chiocca, E. A., Caligiuri, M. A. & Yu, J. The emerging field of oncolytic virus-based cancer immunotherapy. Trends Cancer 9, 122–139 (2023).
Kennedy, E. M. et al. Design of an interferon-resistant oncolytic HSV-1 incorporating redundant safety modalities for improved tolerability. Mol. Ther. Oncolytics 18, 476–490 (2020).
Haines, B. B. et al. ONCR-177, an oncolytic HSV-1 designed to potently activate systemic antitumor immunity. Cancer Immunol. Res. 9, 291–308 (2021).
Clark, I. C. et al. Barcoded viral tracing of single-cell interactions in central nervous system inflammation. Science 372, eabf1230 (2021).
Sanmarco, L. M., Polonio, C. M., Wheeler, M. A. & Quintana, F. J. Functional immune cell–astrocyte interactions. J. Exp. Med. 218, e20202715 (2021).
Kalkavan, H., Rühl, S., Shaw, J. J. P. & Green, D. R. Non-lethal outcomes of engaging regulated cell death pathways in cancer. Nat. Cancer 4, 795–806 (2023).
Suter, T. et al. The brain as an immune privileged site: dendritic cells of the central nervous system inhibit T cell activation. Eur. J. Immunol. 33, 2998–3006 (2003).
Chyuan, I. T., Tsai, H. F., Wu, C. S., Sung, C. C. & Hsu, P. N. TRAIL-mediated suppression of T cell receptor signaling inhibits T cell activation and inflammation in experimental autoimmune encephalomyelitis. Front. Immunol. 9, 15 (2018).
Chen, D. et al. CTLA-4 blockade induces a microglia–Th1 cell partnership that stimulates microglia phagocytosis and anti-tumor function in glioblastoma. Immunity 56, 2086–2104 (2023).
Nakamura, M. et al. APO2L/TRAIL expression in human brain tumors. Acta Neuropathol. 99, 1–6 (2000).
Ishimura, N., Isomoto, H., Bronk, S. F. & Gores, G. J. Trail induces cell migration and invasion in apoptosis-resistant cholangiocarcinoma cells. Am. J. Physiol. Gastrointest. Liver Physiol. 290, G129–G136 (2006).
Kuijlen, J. M. et al. TRAIL-receptor expression is an independent prognostic factor for survival in patients with a primary glioblastoma multiforme. J. Neurooncol. 78, 161–171 (2006).
Li, J. et al. PI3Kγ inhibition suppresses microglia/TAM accumulation in glioblastoma microenvironment to promote exceptional temozolomide response. Proc. Natl Acad. Sci. USA 118, e2009290118 (2021).
Stuart, S. F. et al. The interleukin-11/IL-11 receptor promotes glioblastoma survival and invasion under glucose-starved conditions through enhanced glutaminolysis. Int. J. Mol. Sci. 24, 3356 (2023).
Janiszewska, M. et al. Subclonal cooperation drives metastasis by modulating local and systemic immune microenvironments. Nat. Cell Biol. 21, 879–888 (2019).
Zhang, Y. et al. Interleukin-11 potentiates oligodendrocyte survival and maturation, and myelin formation. J. Neurosci. 26, 12174–12185 (2006).
Hara, T. et al. Interactions between cancer cells and immune cells drive transitions to mesenchymal-like states in glioblastoma. Cancer Cell 39, 779–792 (2021).
Winkler, F. et al. Cancer neuroscience: state of the field, emerging directions. Cell 186, 1689–1707 (2023).
Groot, J. et al. A first-in-human Phase I trial of the oral p-STAT3 inhibitor WP1066 in patients with recurrent malignant glioma. CNS Oncol. 11, CNS87 (2022).
Thang, M., Mellows, C., Mercer-Smith, A., Nguyen, P. & Hingtgen, S. Current approaches in enhancing TRAIL therapies in glioblastoma. Neurooncol. Adv. 5, vdad047 (2023).
Friedman, G. K. et al. Oncolytic HSV-1 G207 immunovirotherapy for pediatric high-grade gliomas. N. Engl. J. Med. 384, 1613–1622 (2021).
Todo, T. et al. Intratumoral oncolytic herpes virus G47 for residual or recurrent glioblastoma: a phase 2 trial. Nat. Med. 28, 1630–1639 (2022).
Ling, A. L. et al. Clinical trial links oncolytic immunoactivation to survival in glioblastoma. Nature 623, 157–166 (2023).
Wu, M. R., Jusiak, B. & Lu, T. K. Engineering advanced cancer therapies with synthetic biology. Nat. Rev. Cancer 19, 187–195 (2019).
Mitchell, D. A. et al. Tetanus toxoid and CCL3 improve dendritic cell vaccines in mice and glioblastoma patients. Nature 519, 366–369 (2015).
Majzner, R. G. et al. GD2-CAR T cell therapy for H3K27M-mutated diffuse midline gliomas. Nature 603, 934–941 (2022).
Habib, N. et al. Massively parallel single-nucleus RNA-seq with DroNc-seq. Nat. Methods 14, 955–958 (2017).
Macosko, E. Z. et al. Highly parallel genome-wide expression profiling of individual cells using nanoliter droplets. Cell 161, 1202–1214 (2015).
Kilian, M. et al. T-cell receptor therapy targeting mutant capicua transcriptional repressor in experimental gliomas. Clin. Cancer Res. 28, 378–389 (2022).
John Lin, C.-C. et al. Identification of diverse astrocyte populations and their malignant analogs. Nat. Neurosci. 20, 396–405 (2017).
Yamamoto, H. et al. Genome editing of Nf1, Pten, and Trp53 in neonatal mice induces glioblastomas positive for oligodendrocyte lineage transcription factor 2. J. Toxicol. Pathol. 34, 359–365 (2021).
Ito, T. et al. A constitutively active arylhydrocarbon receptor induces growth inhibition of jurkat T cells through changes in the expression of genes related to apoptosis and cell cycle arrest. J. Biol. Chem. 279, 25204–25210 (2004).
Lee, Y., Messing, A., Su, M. & Brenner, M. GFAP promoter elements required for region-specific and astrocyte-specific expression. Glia 56, 481–493 (2008).
Gutierrez-Vazquez, C. & Quintana, F. J. Protocol for in vitro analysis of pro-inflammatory and metabolic functions of cultured primary murine astrocytes. STAR Protoc. 3, 101033 (2022).
Petersen, M. A. et al. Fibrinogen activates BMP signaling in oligodendrocyte progenitor cells and inhibits remyelination after vascular damage. Neuron 96, 1003–1012 (2017).
Armstrong, R. C. Isolation and characterization of immature oligodendrocyte lineage cells. Methods 16, 282–292 (1998).
Natarajan, R. et al. STAT3 modulation to enhance motor neuron differentiation in human neural stem cells. PLoS ONE 9, e100405 (2014).
Beaudoin, G. M. 3rd et al. Culturing pyramidal neurons from the early postnatal mouse hippocampus and cortex. Nat. Protoc. 7, 1741–1754 (2012).
Goshi, N., Morgan, R. K., Lein, P. J. & Seker, E. Correction to: A primary neural cell culture model to study neuron, astrocyte, and microglia interactions in neuroinflammation. J. Neuroinflammation 19, 49 (2022).
Bowman-Kirigin, J. A. et al. The conventional dendritic cell 1 subset primes CD8+ T cells and traffics tumor antigen to drive antitumor immunity in the brain. Cancer Immunol. Res. 11, 20–37 (2023).
Bray, N. L., Pimentel, H., Melsted, P. & Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 34, 525–527 (2016).
Frankish, A. et al. Gencode 2021. Nucleic Acids Res. 49, D916–D923 (2021).
Soneson, C., Love, M. I. & Robinson, M. D. Differential analyses for RNA-seq: transcript-level estimates improve gene-level inferences. F1000Res. 4, 1521 (2015).
Dann, E., Henderson, N. C., Teichmann, S. A., Morgan, M. D. & Marioni, J. C. Differential abundance testing on single-cell data using k-nearest neighbor graphs. Nat. Biotechnol. 40, 245–253 (2022).
Chu, T., Wang, Z., Pe’er, D. & Danko, C. G. Cell type and gene expression deconvolution with BayesPrism enables Bayesian integrative analysis across bulk and single-cell RNA sequencing in oncology. Nat. Cancer 3, 505–517 (2022).
Badia, I. M. P. et al. decoupleR: ensemble of computational methods to infer biological activities from omics data. Bioinform. Adv. 2, vbac016 (2022).
Jin, S. et al. Inference and analysis of cell–cell communication using CellChat. Nat. Commun. 12, 1088 (2021).
Tang, Z., Kang, B., Li, C., Chen, T. & Zhang, Z. GEPIA2: an enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 47, W556–W560 (2019).
Durafourt, B. A., Moore, C. S., Blain, M. & Antel, J. P. Isolating, culturing, and polarizing primary human adult and fetal microglia. Methods Mol. Biol. 1041, 199–211 (2013).
Gut, G., Herrmann, M. D. & Pelkmans, L. Multiplexed protein maps link subcellular organization to cellular states. Science 361, eaar7042 (2018).
Alam, M. S. Proximity ligation assay (PLA). Curr. Protoc. Immunol. 123, e58 (2018).
Ravi, V. M. et al. Spatially resolved multi-omics deciphers bidirectional tumor-host interdependence in glioblastoma. Cancer Cell 40, 639–655 (2022).
Wang, L. B. et al. Proteogenomic and metabolomic characterization of human glioblastoma. Cancer Cell 39, 509–528 (2021).
Acknowledgements
This work was supported by grants NS102807, ES02530, ES029136 and AI126880 from the NIH; RG4111A1 and JF2161-A-5 from the NMSS; RSG-14-198-01-LIB from the American Cancer Society; and PA-1604-08459 from the International Progressive MS Alliance. C.F.A. was supported by a scholarship from the German Academic Exchange Service (DAAD). B.M.A. was supported by the Training Program in Nervous System Tumors (K12CA090354) from the NCI/NIH, the Career Enhancement Program (CEP) for the SPORE at Harvard Cancer Center (P50CA165962) from the NCI/NIH, the Post-Doctoral Fellowship in Translational Medicine from the PhRMA Foundation, a T32 Cancer Neuroscience Training Grant at Harvard Cancer Center (T32CA27386) and Career Development Award-2 from the Veterans Affairs Medical Center (1IK2BX006568-01). M.K. was supported by a Leopoldina Research Fellow Grant of the German Academy of Sciences. L.F. was funded by Gemeinnützige Hertie-Stiftung (medMS-Doktorandenprogramm). H.-G.L. was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2021R1A6A3A14039088), and The Gene Lay Institute of Immunology and Inflammation of Brigham and Women’s Hospital, Massachusetts General Hospital, and Harvard Medical School (FP-0034491). J.E.K. was supported by a T32 Cancer Neuroscience training grant (T32CA27386). J.-H.L. was supported by the Basic Science Research Program funded by National Research Foundation of Korea (NRF)/Ministry of Education (2022R1A6A3A03071157), and long-term postdoctoral fellowship funded by Human Frontier Science Program (LT0015/2023-L). T.I. was supported by the EMBO postdoctoral fellowship (ALTF: 1009-2021). C.M.P. was supported by the National Multiple Sclerosis Society (FG-2307-42209) and the Mayer Foundation. M.L. was supported by NIH (F31AI176618). M.A.W. was supported by the NINDS, the NIMH, the NIDA and the NCI (R01MH130458, R01MH132632, R01DA061199, R00NS114111 and T32CA207201). We thank all members of the Quintana Laboratory for their advice; R. Krishnan for technical assistance with flow cytometry and FACS; D. Ling for technical assistance with confocal microscopy; I. A. Glass for help with deriving human primary astrocytes; S. Feau, J. Jacques, P. Grezsik and M. Floyd for antibody sequencing, cloning and viral production; and V. Haridas for technical assistance with imaging flow cytometry.
Author information
Authors and Affiliations
Contributions
C.F.A. and F.J.Q. designed the study. C.F.A., B.M.A., F.G., M.K., L.M.S., F.P., J.M.R., H.-G.L., G.P., J.E.K., J.-H.L., T.I., C.M.P., L.S., J.M., A.S., M.L., E.N.C., A.P., L. Li and K.F. performed or assisted with in vitro and in vivo experiments. Z.L. and C.F.A. performed bioinformatic analyses. C.F.A., I.C.C. and M.A.W. performed and aided with microfluidics for scRNA-seq experiments. M.D. and L.F. performed multiplex immunofluorescence of glioblastoma samples. F.G., C.A.S., C.Q., L. Lerner and F.J.Q. designed and generated engineered oncolytic viruses. K.L.L., D.A.R., E.A.C. and M.P. provided glioblastoma samples. B.D., J.L., D.H.S. and J.P.A. provided unique reagents. C.F.A. and F.J.Q. wrote the manuscript with input from co-authors. F.J.Q. directed and supervised the study.
Corresponding author
Ethics declarations
Competing interests
C.A.S., L. Lerner and C.Q. were employees of Oncorus during the performance of some of these studies. The other authors declare no competing interests.
Peer review
Peer review information
Nature thanks Ido Amit, Michelle Monje, Michael Platten and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 scRNA-seq analysis of astrocytes in GBM.
(a) Cluster markers in GBM astrocyte scRNA-seq. (b) UMAP plot colored by patient of tissue origin. (c) Astrocyte cluster proportions by time to recurrence. (d) UMAP plot colored and split by astrocyte cluster. (e) Gene ontology analysis showing overrepresentation of biological pathways (PantherDB) for differentially expressed genes in each astrocyte cluster. Cluster 0 had insufficient DEGs (n = 2) for overrepresentation analysis. (f) CellChat interaction prediction analysis of cluster 2 astrocytes and different TME compartments analysed by scRNA-seq22. (g,h) Volcano plots of differentially expressed genes in all astrocytes (g) or cluster 2 astrocytes and cluster 4 astrocytes (h) comparing patients with early (≤ 9 months) vs late recurrence (> 9 months) as indicated. n indicates biologically independent samples.
Extended Data Fig. 2 Validation of GBM-associated astrocyte analyses in an independent patient cohort.
(a) Transcript capture in snRNA-seq from frozen GBM tissues. (b) Results of copy number variation (CNV) calling algorithm CONICSmat used for exclusion of malignant GBM cells by chromosome 7 polysomy and chromosome 10 monosomy in validation snRNA-seq studies. (c) Cell type calling by use of canonical markers in validation snRNA-seq studies. Clusters 0 and 2 were selected for analysis of astrocytes. (d) Proportion of each cell type captured in validation snRNA-seq studies. (e) Elbow plot showing Principal Component Analysis (PCA) in GBM snRNA-seq astrocyte data. (f) UMAP of validation snRNA-seq analysis of non-malignant cells (n = 6,502 nuclei; Control = 1,154 nuclei, GBM = 5,348 nuclei). (g) Astrocyte cluster proportions by sample. (h) Cluster distribution of subclustered astrocytes in snRNA-seq. (i) Heatmap of cluster markers from GBM astrocyte snRNA-seq validation studies. (j) TNFSF10 expression in GBM cells analysed by scRNA-seq4. OPC, oligodendrocyte progenitor cell; NPC, neural progenitor cell; AC, astrocyte; MES, mesenchymal. n indicates biologically independent samples.
Extended Data Fig. 3 scRNA-seq analysis of astrocytes in the GL261 model.
(a) In vivo bioluminescence signal used to quantify luciferase-expressing GL261 tumour size at 7 and 14 days after implantation. Net photons per second (P/s) after background correction are shown. Boxplot shows interquartile range with mean, minimal and maximal values. (b-d) Violin plots of quality control metrics for GL261 Aldh1l1cre-ERT2/TdTomato reporter cell scRNA-seq showing number of detected genes (b), number of counts (c), and percent of mitochondrial reads (d). (e) Elbow plot of principal component analysis. (f) Heatmap of cluster markers. (g) Feature plots showing expression of canonical astrocyte markers Gfap, Aldoc, Slc1a2, Slc1a3, Aldh1l1, S100b, Gja1 and Vim in mouse scRNA-seq. (h,i) UMAP plot split by condition (h) and cluster proportions by sample (i). (j) MiloR differential abundance results showing the fold change (FC) of neighborhood abundance in GL261 vs. Control samples. Neighborhoods are grouped by cluster. (k) GSEA pathway analysis of cluster 1 cells in GL261 scRNA-seq data. NES, normalized enrichment score. n indicates biologically independent samples.
Extended Data Fig. 4 In vivo detection of TRAIL+ astrocytes.
(a,b) Flow cytometry gating strategy for analysis of TdTomato+ TRAIL+ cells in Aldh1l1Cre-ERT2/TdTomato reporter mice (a) and analysis of TRAIL expression in TdTomato+/− fractions (b). Paired two-tailed t-test. (c) Immunofluorescence analysis of the tumour border in GL261 tumour-bearing Aldh1l1eGFP reporter mice at day 15 (left). The fluorescence signals of DAPI, TRAIL, GFP astrocyte reporter are shown along with a quantification of TRAIL+ cells within GFP+ or GFP- fractions (right). (d) Immunofluorescence analysis and quantification of TRAIL+ astrocytes in naïve or sham-injured mice after 15 days. Data shown as mean ± SEM. Unpaired two-tailed t-test used for statistical analysis. n indicates biologically independent samples.
Extended Data Fig. 5 Analysis of T-cell apoptosis in GBM and TME characterization after Tnfsf10 knockdown in astrocytes.
(a) Reanalysis of spatial transcriptomics data in GBM86. Spatially-weighted correlations of TRAIL imputation score from Fig. 2f and non-malignant reactive astrocyte signature32. (b) Spatially annotated bulk RNA-seq from Ivy Glioblastoma Atlas showing signature score of death receptor signalling (R-HSA-73887) split by anatomical niche (left). Pearson correlation analysis of TNFSF10 and TNFRSF10B expression split by anatomical niche. (c) Validation by flow cytometry of Tnfsf10-targeting lentivirus-driven genetic perturbation in TdTomato+ Aldh1l1Cre-ERT2/TdTomato reporter cells isolated from naïve mice (n = 8 NTsgRNA, n = 10 sgTnfsf10). (d) Control measurement of TRAIL in a GFP+ GL261 line by flow cytometry 15 days after implantation following the lentivirus-driven genetic perturbation in astrocytes (n = 3 NTsgRNA, n = 2 sgTnfsf10). (e) Analysis by qPCR of off-target Tnfsf10 deletion effects in sorted T cells and myeloid cells effects following lentivirus administration as shown in Fig. 3f. (f) Survival analysis following implantation of CRISPR control-edited or Tnfsf10-deleted GL261 lines (n = 4 GL261-NTsgRNA, n = 6 GL261-sgTnfsf10), (g) Survival analysis of mice implanted with genetically engineered glioma model MK007 following TRAIL inactivation in astrocytes as shown in Fig. 3f (n = 10 each). (h) Survival analysis of GL261-implanted Rag2−/− mice following TRAIL inactivation in astrocytes (n = 5 each). (i,j) Flow cytometry quantification related to Fig. 3h of cleaved caspase-3/7+ single-positive (i) and caspase-3/7+ SYTOX+ double-positive (j) CD4+ or CD8+ T cell populations. Data shown as mean ± SEM. Unpaired two-tailed t-test used for statistical analysis. Survival analysis by Log-rank (Mantel-Cox) test for (f-h). n indicates biologically independent samples.
Extended Data Fig. 6 Analysis of T-cell activation and myeloid cells in after Tnfsf10 knockdown in astrocytes.
(a,b) Activation/exhaustion profiling by flow cytometry of tumour-infiltrating PD-1+ CD8+ (a) or PD-1+ CD4+ (b) T cells following genetic perturbation of TRAIL in astrocytes (n = 7 NTsgRNA, n = 9 sgTnfsf10). (c) Flow cytometry analysis of T-cell exhaustion markers in tumour-infiltrating CD8+ (left) or CD4+ (right) T cells following genetic perturbation of TRAIL in astrocytes (n = 7 NTsgRNA, n = 9 sgTnfsf10). (d) Tox expression measured by qPCR in GL261-infiltrating CD4 or CD8 T cells following genetic inactivation of TRAIL in astrocytes. (e,f) Flow cytometry analysis and representative plots of cytokines after restimulation with PMA/Ionomycin in tumour-infiltrating CD8+ (e) or CD4+ (f) T cells following genetic perturbation of TRAIL in astrocytes (n = 7 NTsgRNA, n = 9 sgTnfsf10). (g) Nuclear translocation of NFAT2 analysed by imaging flow cytometry in splenic CD4+ T cells activated in vitro with anti-CD3/CD28 alone or in combination with recombinant TRAIL as indicated. Violin plot of similarity score between DAPI and NFAT2 analysed using the Mann-Whitney U-test with Sidak’s correction (left) and representative images (right) are shown. (h) DR5 expression median fluorescence intensity (MFI) and histogram distribution in T cell and myeloid compartments from GL261-bearing mice following genetic perturbation with control NTsgRNA (n = 7 mice). (i) Percentage of apoptotic microglia and monocyte-derived macrophages (MDM) in GL261-bearing mice 15 days following genetic inactivation of TRAIL in astrocytes. (j,k) Differential expression analysis by bulk RNA-seq in microglia (j) and flow cytometry analysis of microglia and MDM isolated from GL261-bearing Rag2-/- mice undergoing sgTnfsf10 or NTsgRNA genetic perturbation of astrocytes (k). (l) DR5 median fluorescence intensity and histogram in GL261-infiltrating dendritic cell subtypes. (m) Purity analysis of mixed glia culture, using GFAP, CD45, CD11b, O4, Ter119 as lineage markers. Data shown as mean ± SEM. Unpaired two-tailed t-test. n indicates biologically independent samples.
Extended Data Fig. 7 Analysis of pSTAT3+ TRAIL+ astrocytes.
(a) TNFSF10 expression in human astrocytes stimulated overnight with tumour-conditioned or control medium from three patient-derived GBM lines (BT145, BT482, BT1018). (b) Analysis of calcium signalling in astrocytes by use of Fluo-4-AM calcium indicator in primary mouse astrocytes stimulated in vitro with the indicated conditions. (c) Pearson correlation of STAT3 signalling programs (M5897) and TNFSF10 expression in Ivy Glioblastoma Atlas bulk RNA-seq data split by anatomical structure. (d) pSTAT3 measurement by flow cytometry in TRAIL+ or TRAIL- astrocytes from Aldh1l1Cre-ERT2/TdTomato reporter mice implanted with MK007. (e) TNFSF10 expression and STAT3 (Y705) phosphorylation in GBM samples from CPTAC-3 dataset. Only patients that ultimately succumbed to GBM and not any other morbidity were analysed. Boxplot with median, interquartile range (IQR) ± 1.5IQR, and minimal/maximal values is shown. (f) STAT3 binding sites within 2 kb upstream of the transcription start site (TSS) in the human TNFSF10 predicted by MULAN. (g) pSTAT3 (Y705) in GL261 TCM-treated murine astrocytes. Data shown as mean ± SEM. Unpaired two-tailed t-test used. n indicates biologically independent samples.
Extended Data Fig. 8 IL-11 signalling in GBM-associated astrocytes.
(a,b) IL11 expression per cell type (a) and GBM subtype (b) in TCGA after cell-type deconvolution. (c) IL11 (left) and IL11RA expression by cell type in scRNA-seq22. (d) Spatially annotated bulk RNA-seq from Ivy Glioblastoma Atlas showing expression correlation of STAT3 signalling program (M5897) and TRAIL-driven apoptosis signalling (M23448) split by anatomical structure. (e) Spatial transcriptomics analysis of human GBM performed with SPATA286 showing spatially-weighted correlation of IL11 expression with GBM subtype metaprograms32. (f) IL11RA expression by time to recurrence in GBM-associated astrocytes analysed by scRNA-seq in Fig. 1b. (g) IL11RA expression in GBM-associated astrocytes determined by scRNA-seq. (h) Il11ra1 expression in GL261-associated astrocytes determined by scRNA-seq. (i-k) IL-11 quantified by ELISA in conditioned media from primary human GBM lines (i, n = 4 each), primary mouse astrocytes or GL261 cultures (j, n = 3 astrocytes, n = 6 GL261), or genetically-engineered glioma models (k, n = 4 each). (l) pSTAT3 (Y705) in primary murine astrocytes stimulated with rmIL-11 for 15 mins. (m) TNFSF10 promoter luciferase reporter activity in HepG2 cells treated with rhIL-11 for 24 or 48 h. (n) Tnfsf10 expression measured by qPCR in purified cultures of either microglia, oligodendrocytes, or neurons overnight with recombinant IL-11. (o) IL15 and TGFB1 expression in GBM cells analysed by scRNA-seq4. (p) Tnfsf10 expression measured by qPCR in primary mouse astrocyte cultures stimulated with the indicated cytokines. Data shown as mean ± SEM. Unpaired two-tailed t-test and two-way ANOVA used. n indicates biologically independent samples.
Extended Data Fig. 9 Effect of IL-11 on astrocyte function and regulation of IL-11 expression in glioblastoma cells.
(a) Cleaved caspase-3/7+ and cleaved caspase-8+ CD4/8+ T cells in co-cultures with GL261 TCM pre-treated astrocytes with IL-11 neutralization (n = 6 CD4+ groups; n = 3 CD8+ anti-Isotype; n = 4 CD8+ anti-IL-11). (b) Cleaved caspase-3/7+ and cleaved caspase-8+ CD4/8+ T cells co-cultured with rmIL-11-treated astrocytes (n = 11 CD4+ control; n = 6 CD4+ rmIL-11; n = 7 CD8+ control; n = 4 CD8+ rmIL-11). (c) Percentage of cleaved caspase-3/7+ (left) and cleaved caspase-8+ (right) CD4+ or CD8+ T cells co-cultured with rmIL-11 pre-treated astrocytes and TRAIL-blocking antibodies (n = 6 CD4+; n = 3 CD8+ anti-Isotype; n = 4 CD8+ anti-TRAIL). (d) Percentage of cleaved caspase-3/7+ CD4+ T cells co-cultured with astrocytes pre-treated with rmIL-11 in combination with STAT3 inhibitor or vehicle. TRAIL-blocking or control antibodies, or recombinant TRAIL were added at start of co-culture. (n = 6 each). (e) Validation of astrocyte-specific Il11ra1 knockdown by immunofluorescence. (f) Absolute cell count of the indicated compartments in GL261-bearing mice following astrocyte-specific Il11ra1 or non-targeting control inactivation. (g,h) IL-11 concentration in TCM (g) (n = 4 empty vector; n = 2 IL-11 OE) and growth dynamics (h) (n = 6 empty vector; n = 2 IL-11 OE) of empty vector transfected or IL-11 overexpressing (OE) GL261 cells (n = 3 each). (i) Relative increase in bioluminescence tumour size between days 6 and 11 in control or IL-11-overexpressing tumours. (j) Analysis of the Il11 promoter showing multiple AHR consensus binding sites. (k,l) Expression of Il11 (left) and Cyp1b1 (right) in GL261 cells harboring control- or AHR-deletion (k) or a constitutively activated form of AHR (l). (m,n) Mouse GL261 (m) or human U87 (n) GBM cells stimulated with L-Kynurenine (Kyn) alone or in combination with AHR inhibitor CH223191 (AHRi). Data shown as mean ± SEM. Unpaired two-tailed t-test and two-way ANOVA used. n indicates biologically independent samples.
Extended Data Fig. 10 FACS analysis of T-cell function in HSV-1-treated mice.
(a) Apoptosis in T cells co-cultured with IFNγ-activated astrocytes and anti-TRAIL single-chain antibody, which was incorporated into HSV-1α-TRAIL (n = 5 anti-Isotype, n = 3 anti-TRAIL). (b) RT-qPCR detection of anti-TRAIL scFv sequence from RNA isolated from brains of GL261-bearing mice 24 h after treatment with HSV-1α-TRAIL. (c) Immunofluorescence analysis of CD68+ myeloid cell infiltration into GL261 tumours following treatment with HSV-1Empty or HSV-1α-TRAIL. (d) Absolute T cell count from GL261-bearing mouse brains following treatment with HSV-1Empty or HSV-1α-TRAIL. (e,f) Quantification of activated/exhausted tumour-infiltrating HSV-1 MHC-I tetramer+ PD-1+ CD8+ (e) or GARC-177-85 tetramer+ PD-1+ CD8+ (f) following HSV-1 treatment (n = 8 each). (g,h) Quantification of activated/exhausted tumour-infiltrating PD-1+ CD8+ (g) or PD-1+ CD4+ (h) T cells following HSV-1 treatment (n = 8 each). (i,j) FACS analysis of T-cell exhaustion markers in tumour-infiltrating CD8+ (i) or CD4+ (j) T cells with relevance for anti-tumour immunity in the context of TRAIL blockade (n = 8 each). (k,l) FACS analysis (left) and exemplary plots (right) of cytokine expression in tumour-infiltrating CD8+ (k) or CD4+ (l) T cells (n = 8 each). Data shown as mean ± SEM. Unpaired two-tailed t-test used. (m) Graphical summary. In this study, we describe the induction of TRAIL+ astrocytes in GBM. TRAIL+ astrocytes are regulated by GBM-derived IL-11 via STAT3 signalling and limit anti-tumour immunity by inducing death receptor-mediated apoptosis in T cells. Treatment with an oncolytic HSV engineered to express TRAIL blocking single-chain antibodies in the TME improves the anti-tumour response. n indicates biologically independent samples. Schematic in (m) created in BioRender. Lee, J. (2025) (https://BioRender.com/pn72nh0).
Supplementary information
Supplementary Information
Supplementary Tables 1–3 and supplementary references.
Supplementary Data
Supplementary Data 1–12.
Source data
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Faust Akl, C., Andersen, B.M., Li, Z. et al. Glioblastoma-instructed astrocytes suppress tumour-specific T cell immunity. Nature 643, 219–229 (2025). https://doi.org/10.1038/s41586-025-08997-x
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41586-025-08997-x
This article is cited by
-
Immunosuppressive mechanisms and therapeutic interventions shaping glioblastoma immunity
Nature Cancer (2026)
-
131I/MTX-loaded engineered outer membrane vesicles for sequentially coordinated radio-metabolic-immunotherapy of glioma
Journal of Nanobiotechnology (2025)
-
Neuroimmune interactions in health and disease
Nature Immunology (2025)
-
Advanced nanotheranostic approaches for targeted glioblastoma treatment: a synergistic fusion of CRISPR-Cas gene editing, AI-driven tumor profiling, and BBB-modulation
Medical Oncology (2025)
-
TRAILblazing Astrocytes: Glioblastoma’s Covert Immunosuppressive Agents
Neuroscience Bulletin (2025)
