
Abstract
Cancer-associated fibroblasts (CAFs) have a pivotal cancer-supportive role, yet CAF-targeted therapies are lacking1,2. Here, using spatial transcriptomics and single-cell RNA sequencing, we investigate the role of nicotinamide N-methyltransferase (NNMT) in high-grade serous ovarian cancer. Mechanistically, NNMT-induced H3K27me3 hypomethylation drives complement secretion from CAFs, attracting immunosuppressive myeloid-derived suppressor cells (MDSCs) to the tumour. Nnmt knockout in immunocompetent mice impairs tumour growth in syngeneic ovarian, breast and colon tumour models through enhanced CD8+ T cell activation. Using high-throughput screening, we develop a potent and specific NNMT inhibitor that reduces the tumour burden and metastasis in multiple mouse cancer models and restores immune checkpoint blockade efficacy by decreasing CAF-mediated recruitment of MDSCs and reinvigorating CD8+ T cell activation. Our findings establish NNMT as a central CAF regulator and a promising therapeutic target to mitigate immunosuppression in the tumour microenvironment.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


Data availability
All data generated or analysed during this study are included in the Article and its Supplementary Information. Raw data have been made publicly available at the Gene Expression Omnibus (GEO) under the following accessions: spatial transcriptomics (GSE295028), scRNA-seq (GSE292661), RNA-seq (GSE124014)6 and ChIP–seq (GSE124015)6. Processed scRNA-seq data are available at Zenodo76 (https://doi.org/10.5281/zenodo.15238384). Immune deconvolution was performed using the SpatialDecon (v.3.18) package using the SafeTME reference matrix19 available online (https://rdrr.io/bioc/SpatialDecon/man/safeTME.html). The MSigDB v2023 Molecular Signatures Database was used to remap gene symbols to human or mouse orthologues (https://www.gsea-msigdb.org/gsea/msigdb). The UniProt reference proteome used for global proteomics was: UP000000589_10090. Gene set databases included the GOBP (https://www.gsea-msigdb.org/gsea/msigdb/human/genesets.jsp?collection=GO:BP) collection and a custom gene set derived from ref. 3 as outlined in Supplementary Table 7. The pan-cancer scRNA-seq CAF data are available online (https://gist-fgl.github.io/sc-caf-atlas/, ref. 21; and https://chenxisd.shinyapps.io/pancaf/, ref. 22). The coordinates and structure factor files have been deposited in the Protein Data Bank (PDB) under accession code 9ATV. Source data are provided with this paper.
Code availability
The code used in this study has been made available at GitHub. The package code for scRNA-seq analysis is available at https://github.com/yan-cri/scRICA. The Code for fibroblast detection is available at https://github.com/jannameldorf/HE-fibroblast-detection, and the code for monocyte detection is available at https://github.com/apiffko/IF.mono.quantif. Both codes have been archived at Zenodo (https://doi.org/10.5281/zenodo.15257915, ref. 77; and https://doi.org/10.5281/zenodo.14982759, ref. 78, respectively).
References
Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 16, 582–598 (2016).
Sahai, E. et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 20, 174–186 (2020).
Chhabra, Y. & Weeraratna, A. T. Fibroblasts in cancer: unity in heterogeneity. Cell 186, 1580–1609 (2023).
Bagaev, A. et al. Conserved pan-cancer microenvironment subtypes predict response to immunotherapy. Cancer Cell 39, 845–865 (2021).
Kandalaft, L. E., Dangaj Laniti, D. & Coukos, G. Immunobiology of high-grade serous ovarian cancer: lessons for clinical translation. Nat. Rev. Cancer 22, 640–656 (2022).
Eckert, M. A. et al. Proteomics reveals NNMT as a master metabolic regulator of cancer-associated fibroblasts. Nature 569, 723–728 (2019).
Ulanovskaya, O. A., Zuhl, A. M. & Cravatt, B. F. NNMT promotes epigenetic remodeling in cancer by creating a metabolic methylation sink. Nat. Chem. Biol. 9, 300–306 (2013).
Zhang, L. et al. Accumulation of nicotinamide N-methyltransferase (NNMT) in cancer-associated fibroblasts: a potential prognostic and predictive biomarker for gastric carcinoma. J. Histochem. Cytochem. 69, 165–176 (2021).
Song, M. et al. High stromal nicotinamide N-methyltransferase (NNMT) indicates poor prognosis in colorectal cancer. Cancer Med. 9, 2030–2038 (2020).
Sartini, D. et al. Nicotinamide N-methyltransferase in non-small cell lung cancer: promising results for targeted anti-cancer therapy. Cell Biochem. Biophys. 67, 865–873 (2013).
Zhao, H., Li, R., Chen, Y., Yang, X. & Shang, Z. Stromal nicotinamide N-methyltransferase orchestrates the crosstalk between fibroblasts and tumour cells in oral squamous cell carcinoma: evidence from patient-derived assembled organoids. Oncogene 42, 1166–1180 (2023).
Ogawa, M. et al. Tumor stromal nicotinamide N-methyltransferase overexpression as a prognostic biomarker for poor clinical outcome in early-stage colorectal cancer. Sci. Rep. 12, 2767 (2022).
Yang, J. et al. Overexpression of nicotinamide n-methyltransferase mainly covers stroma of colorectal cancer and correlates with unfavorable survival by its product 1-MNA. J. Cancer 12, 6170–6181 (2021).
Russell, D. L., Brown, H. M. & Dunning, K. R. ADAMTS proteases in fertility. Matrix Biol. 44–46, 54–63 (2015).
Geisler, S. B. et al. Obscurin-like 1, OBSL1, is a novel cytoskeletal protein related to obscurin. Genomics 89, 521–531 (2007).
Diehl, V. et al. The role of decorin and biglycan signaling in tumorigenesis. Front. Oncol. 11, 801801 (2021).
De Martino, D. & Bravo-Cordero, J. J. Collagens in cancer: structural regulators and guardians of cancer progression. Cancer Res. 83, 1386–1392 (2023).
Reis, E. S., Mastellos, D. C., Ricklin, D., Mantovani, A. & Lambris, J. D. Complement in cancer: untangling an intricate relationship. Nat. Rev. Immunol. 18, 5–18 (2018).
Danaher, P. et al. Advances in mixed cell deconvolution enable quantification of cell types in spatial transcriptomic data. Nat. Commun. 13, 385 (2022).
Duraiswamy, J. et al. Myeloid antigen-presenting cell niches sustain antitumor T cells and license PD-1 blockade via CD28 costimulation. Cancer Cell 39, 1623–1642 (2021).
Luo, H. et al. Pan-cancer single-cell analysis reveals the heterogeneity and plasticity of cancer-associated fibroblasts in the tumor microenvironment. Nat. Commun. 13, 6619 (2022).
Ma, C. et al. Pan-cancer spatially resolved single-cell analysis reveals the crosstalk between cancer-associated fibroblasts and tumor microenvironment. Mol. Cancer 22, 170 (2023).
Maniati, E. et al. Mouse ovarian cancer models recapitulate the human tumor microenvironment and patient response to treatment. Cell Rep. 30, 525–540 (2020).
Roby, K. F. et al. Development of a syngeneic mouse model for events related to ovarian cancer. Carcinogenesis 21, 585–591 (2000).
Veglia, F., Sanseviero, E. & Gabrilovich, D. I. Myeloid-derived suppressor cells in the era of increasing myeloid cell diversity. Nat. Rev. Immunol. 21, 485–498 (2021).
Bronte, V. et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat. Commun. 7, 12150 (2016).
Guo, R.-F. & Ward, P. A. Role of C5a in inflammatory responses. Annu. Rev. Immunol. 23, 821–852 (2005).
Markiewski, M. M. et al. Modulation of the antitumor immune response by complement. Nat. Immunol. 9, 1225–1235 (2008).
Kumar, V., Lee, J. D., Clark, R. J. & Woodruff, T. M. Development and validation of a LC-MS/MS assay for pharmacokinetic studies of complement C5a receptor antagonists PMX53 and PMX205 in mice. Sci. Rep. 8, 8101 (2018).
Litvinchuk, A. et al. Complement C3aR inactivation attenuates tau pathology and reverses an immune network deregulated in tauopathy models and Alzheimer’s disease. Neuron 100, 1337–1353 (2018).
An, L.-L. et al. Complement C5a induces PD-L1 expression and acts in synergy with LPS through Erk1/2 and JNK signaling pathways. Sci. Rep. 6, 33346 (2016).
Ferrari, K. J. et al. Polycomb-dependent H3K27me1 and H3K27me2 regulate active transcription and enhancer fidelity. Mol. Cell 53, 49–62 (2014).
Hornburg, M. et al. Single-cell dissection of cellular components and interactions shaping the tumor immune phenotypes in ovarian cancer. Cancer Cell 39, 928–944 (2021).
Maeda, M. et al. Cancer cell niche factors secreted from cancer-associated fibroblast by loss of H3K27me3. Gut 69, 243–251 (2020).
Tyan, S.-W. et al. Breast cancer cells induce stromal fibroblasts to secrete ADAMTS1 for cancer invasion through an epigenetic change. PLoS ONE 7, e35128 (2012).
van Haren, M. J. et al. Esterase-sensitive prodrugs of a potent bisubstrate inhibitor of nicotinamide N-methyltransferase (NNMT) display cellular activity. Biomolecules 11, 1357 (2021).
Policarpo, R. L. et al. High-affinity alkynyl bisubstrate inhibitors of nicotinamide N-methyltransferase (NNMT). J. Med. Chem. 62, 9837–9873 (2019).
Chen, D. et al. Novel propargyl-linked bisubstrate analogues as tight-binding inhibitors for nicotinamide N-methyltransferase. J. Med. Chem. 62, 10783–10797 (2019).
Babault, N. et al. Discovery of bisubstrate inhibitors of nicotinamide N-methyltransferase (NNMT). J. Med. Chem. 61, 1541–1551 (2018).
Hanson, Q. M. et al. Target class profiling of small-molecule methyltransferases. ACS Chem. Biol. 18, 969–981 (2023).
Barrows, R. D. et al. Potent uncompetitive inhibitors of nicotinamide N-methyltransferase (NNMT) as in vivo chemical probes. J. Med. Chem. 65, 14642–14654 (2022).
Tseng, D. et al. Anti-CD47 antibody–mediated phagocytosis of cancer by macrophages primes an effective antitumor T-cell response. Proc. Natl Acad. Sci. USA 110, 11103–11108 (2013).
Lavie, D., Ben-Shmuel, A., Erez, N. & Scherz-Shouval, R. Cancer-associated fibroblasts in the single-cell era. Nat. Cancer 3, 793–807 (2022). 2022 37.
Kerdidani, D. et al. Lung tumor MHCII immunity depends on in situ antigen presentation by fibroblasts. J. Exp. Med. 219, e20210815 (2022).
Kanzaki, R. & Pietras, K. Heterogeneity of cancer-associated fibroblasts: opportunities for precision medicine. Cancer Sci. 111, 2708–2717 (2020).
Yang, D., Liu, J., Qian, H. & Zhuang, Q. Cancer-associated fibroblasts: from basic science to anticancer therapy. Exp. Mol. Med. 55, 1322–1332 (2023).
Kennel, K. B., Bozlar, M., De Valk, A. F. & Greten, F. R. Cancer-associated fibroblasts in inflammation and antitumor immunity. Clin. Cancer Res. 29, 1009–1016 (2023).
Chen, Y., McAndrews, K. M. & Kalluri, R. Clinical and therapeutic relevance of cancer-associated fibroblasts. Nat. Rev. Clin. Oncol. 18, 792–804 (2021).
Weigert, M. et al. A cell atlas of the human fallopian tube throughout the menstrual cycle and menopause. Nat. Commun. 16, 372 (2025).
Hao, Y. et al. Dictionary learning for integrative, multimodal and scalable single-cell analysis. Nat. Biotechnol. 42, 293–304 (2024).
Wu, T. et al. clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innovation 2, 100141 (2021).
Iyer, S. et al. Genetically defined syngeneic mouse models of ovarian cancer as tools for the discovery of combination immunotherapy. Cancer Discov. 11, 384–407 (2021).
Trimboli, A. J. et al. Pten in stromal fibroblasts suppresses mammary epithelial tumours. Nature 461, 1084–1091 (2009).
Schwörer, S. et al. Hypoxia potentiates the inflammatory fibroblast phenotype promoted by pancreatic cancer cell–derived cytokines. Cancer Res. 83, 1596–1610 (2023).
Shen, F. W. et al. Cloning of Ly-5 cDNA. Proc. Natl Acad. Sci. USA 82, 7360–7363 (1985).
Cordero, A. B., Kwon, Y., Hua, X. & Godwin, A. K. In vivo imaging and therapeutic treatments in an orthotopic mouse model of ovarian cancer. J. Vis. Exp. 42, e2125 (2010).
Zhang, G.-L., Zhang, Y., Cao, K.-X. & Wang, X.-M. Orthotopic injection of breast cancer cells into the mice mammary fat pad. J. Vis. Exp. 143, e58604 (2019).
Downs, E. C., Robertson, N. E., Riss, T. L. & Plunkett, M. L. Calcium alginate beads as a slow-release system for delivering angiogenic molecules in vivo and in vitro. J. Cell. Physiol. 152, 422–429 (1992).
Nielsen, T. B., Yan, J., Luna, B. & Spellberg, B. Murine oropharyngeal aspiration model of ventilator-associated and hospital-acquired bacterial pneumonia. J. Vis. Exp. 136, e57672 (2018).
Wang, L. et al. Mild hypothermia alleviates complement C5a-induced neuronal autophagy during brain ischemia–reperfusion injury after cardiac arrest. Cell. Mol. Neurobiol. 43, 1957–1974 (2023).
Francke, A., Herold, J., Weinert, S., Strasser, R. H. & Braun-Dullaeus, R. C. Generation of mature murine monocytes from heterogeneous bone marrow and description of their properties. J. Histochem. Cytochem. 59, 813 (2011).
Rappsilber, J., Mann, M. & Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2, 1896–1906 (2007).
Müller-Reif, J. B. et al. A new parallel high-pressure packing system enables rapid multiplexed production of capillary columns. Mol. Cell. Proteom. 20, 100082 (2021).
Demichev, V., Messner, C. B., Vernardis, S. I., Lilley, K. S. & Ralser, M. DIA-NN: neural networks and interference correction enable deep proteome coverage in high throughput. Nat. Methods 17, 41–44 (2020).
Ariotta, V. et al. H&E image analysis pipeline for quantifying morphological features. J. Pathol. Inform. 14, 100339 (2023).
Stringer, C., Wang, T., Michaelos, M. & Pachitariu, M. Cellpose: a generalist algorithm for cellular segmentation. Nat. Methods 18, 100–106 (2021).
Kowal, M., Żejmo, M., Skobel, M., Korbicz, J. & Monczak, R. Cell nuclei segmentation in cytological images using convolutional neural network and seeded watershed algorithm. J. Digit. Imaging 33, 231–242 (2020).
Hasan, M. R., Hassan, N., Khan, R., Kim, Y.-T. & Iqbal, S. M. Classification of cancer cells using computational analysis of dynamic morphology. Comput. Methods Programs Biomed. 156, 105–112 (2018).
Pachitariu, M. & Stringer, C. Cellpose 2.0: how to train your own model. Nat. Methods 19, 1634–1641 (2022).
Gao, P. et al. A graph convolutional network-based screening strategy for rapid identification of SARS-CoV-2 cell-entry inhibitors. J. Chem. Inf. Model. 62, 1988–1997 (2022).
Jafari, R. et al. The cellular thermal shift assay for evaluating drug target interactions in cells. Nat. Protoc. 9, 2100–2122 (2014).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D 60, 2126–2132 (2004).
Vagin, A. A. et al. REFMAC5 dictionary: organization of prior chemical knowledge and guidelines for its use. Acta Crystallogr. D 60, 2184–2195 (2004).
Pino, L. K. et al. The Skyline ecosystem: informatics for quantitative mass spectrometry proteomics. Mass Spectrom. Rev. 39, 229–244 (2020).
Metz, K. S. et al. Coral: clear and customizable visualization of human kinome data. Cell Syst. 7, 347–350 (2018).
Heide, J. et al. Data for ‘NNMT inhibition in cancer-associated fibroblasts restores antitumour immunity’. Zenodo https://doi.org/10.5281/zenodo.15238384 (2025).
Heide, J. et al. Code for ‘NNMT inhibition in cancer-associated fibroblasts restores antitumour immunity’. Zenodo https://doi.org/10.5281/zenodo.15257915 (2025).
Heide, J. et al. Code for ‘NNMT inhibition in cancer-associated fibroblasts restores antitumour immunity’. Zenodo https://doi.org/10.5281/zenodo.14982759 (2025).
Acknowledgements
J.H. and A.P. are recipients of the Walter Benjamin scholarship from the German Research Foundation (Deutsche Forschungsgemeinschaft (DFG)) (459327389 to J.H., 455353745 to A.P.). This work was supported by the National Cancer Institute/National Institutes of Health (NCI/NIH) RO1CA211916 and R35CA264619 to E.L.; R01CA262508 to R.R.W.; and R00CA259224 to S.S.; with additional funding from NCI/NIH under Chemical Biology Consortium contract no. HHSN261200800001E and intramural funds for the NCATS, as well as the NCI Experimental Therapeutics (NExT) program (603888 to E.L.). We thank the members of the ArsenalBio team for snRNA-seq libraries and sequencing data: L. Berthoin, R. Sit, S. Sabri, S. Panagiotopoulou, B. Gavin and G. Zheng; the staff at the University of Chicago Human Tissue Resource Center (RRID: SCR_019199), the Flow Cytometry Core (RRID: SCR_017760), the Integrated Small Animal Imaging Research Resource (RRID: SCR_017923), the Genomics Facility (RRID: SCR_019196), Pharmaron, the Microscopy Core (P30CA014599), the University of Chicago Metabolomics Platform (RRID: SCR_022932), and the Animal Research Center (RRID: SCR_021806) for technical support, particularly for the support from A. Solanki; all other contributing and supporting members of the Division of Cancer Treatment and Diagnosis (DCTD) at the NCI and the NCI Chemical Biology Consortium for their contributions to the project; G. Isenberg for editing the manuscript; and all of the patients who donated tissue for this study.
Author information
Authors and Affiliations
Contributions
E.L. conceived the project. J.H., A.P., A.J.B., K.Y., S.S., H.L., G.M.S., R.R.W. and E.L. did the experimental planning. J.H., M.F.A., E.T., M.T. and R.A. conducted the in vitro experiments. J.H., A.P., M.F.A., N.L. and K.Y. performed the in vivo studies. L.S. and M.M. performed the proteomic analysis and M.F.A., H. Shah and J.H. the metabolomic experiments. J.H., L.S., K.Z., Y.L. and Q.L. conducted the bioinformatic analysis. A.J.B., J.H. and E.L. selected patient samples and performed the spatial transcriptomics experiments. A.J.B. reviewed HGSOC cases and performed immunohistochemistry scoring. L.D. wrote the code for IHC/IF analysis. S.P., K.K., L.L., T.G.Y., K.C.-C.C., J.H.S., Q.M.H., M.S., H. Sun, G.M.S., M.D.H. and E.L. developed the NNMT inhibitor, and A.O., A.W. and R.K.S. performed X-ray crystallography. J.H., A.P. and E.L. wrote the manuscript with input from all of the authors.
Corresponding author
Ethics declarations
Competing interests
J.H., S.P., M.F.A., K.K., L.L., T.G.Y., K.C.-C.C., H. Sun, A.W., J.H.S., Q.M.H., M.S., G.M.S., M.D.H. and E.L. are listed as inventors on a patent application describing NNMT inhibitors filed by the NIH and The University of Chicago. The E.L. laboratory receives research grant funding from Abbvie through the University of Chicago. R.R.W. has stock and other ownership interests with Boost Therapeutics, Immvira, Reflexion Pharmaceuticals, Coordination Pharmaceuticals, Magi Therapeutics and Oncosenescence; he has served in a consulting or advisory role for Aettis, AstraZeneca, Coordination Pharmaceuticals, Genus, Merck Serono, Nano Proteagen, NKMax America and Shuttle Pharmaceuticals; he has received research grant funding from Varian and Regeneron through the University of Chicago; he has received compensation, including travel, accommodations or expense reimbursement from AstraZeneca, Boehringer Ingelheim and Merck Serono. The other authors declare no competing interests.
Peer review
Peer review information
Nature thanks Stefan Knapp and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Spatial transcriptomics of ovarian cancer stroma identifies NNMT upregulation during ovarian cancer progression.
a, Principal component analysis (PCA) of spatial transcriptomics data of cancer-adjacent ovarian stroma (n = 10), primary ovarian cancer stroma (n = 28), and omental metastases (n = 16). Among the 29 ovarian cancer samples on the tissue microarray (TMA), one stroma sample was not collected due to the low abundance of stroma, and one sample was excluded because the stroma primarily contained vessels (highlighted with an arrow). b, Analysis of NNMT RNA expression according to anatomic region (Kruskal-Wallis test with Dunn’s multiple comparisons test). Sample size as described in (a). c, Immunohistochemistry (IHC): TMA was stained for NNMT, and matched samples of cancer-adjacent ovarian stroma, primary ovarian tumour, and omental metastases of HGSOC are shown (n = 3 patients). d, The TMA was stained for NNMT and scored in NNMTlow (n = 16), NNMTint (n = 13) and NNMThigh (n = 24). The NNMT IHC score was compared with NNMT RNA expression levels from the respective sample (Kruskal-Wallis test with Dunn’s multiple comparisons test). e, CD45 deconvolution analysis of spatial transcriptomics of CD45 + CD163- immune cells in NNMTlow/int (n = 9) and NNMThigh stroma (n = 8) collected from ovarian tumour samples (n = 17). Immune cell area of interest (AOI) was only collected when more than 50 CD45+ CD163− immune cells were present. Treg = regulatory T cells, monocytes C = classical monocytes, monocytes NC I = nonclassical/intermediate monocyte, mDCs = myeloid dendritic cells, pDCs = plasmacytoid dendritic cells, NK = natural killer cells, B naïve = naïve B cells, mast = mast cells. f, CD45 deconvolution analysis of spatial transcriptomics: Proportions of different cell subsets were compared between NNMTlow/int (n = 9) and NNMThigh (n = 8) stroma (Unpaired t-test with Welch correction). g, Representative cores show the co-staining for CD8/Granzyme B and DAPI (aqua) from Fig. 1g. Arrows highlight CD8+ Granzyme B+ cells. Data are shown as mean ± SEM.
Extended Data Fig. 2 scRNA-seq analysis reveals NNMT expression across all CAF subtypes.
a, Dot plot showing canonical marker expression across different cell types in scRNA-seq data from omental metastases of chemotherapy-naïve HGSOC patients (n = 7). Genes used for CAF annotation are listed in Supplementary Table 5. SMC = smooth muscle cells, Adipo/Endo cells = adipocytes/endothelial cells. b, Distribution of patient samples across all major cell types. Total cell numbers per cell type are shown in brackets. CN = chemotherapy naïve. c, Gene ontology enrichment analysis based on the Biological Processes dataset of the identified CAF subsets: inflammatory CAFs (iCAFs), vascular CAFs (vCAFs), proliferative CAFs (prolCAFs), antigen-presenting CAFs (apCAFs), oxidative phosphorylation CAFs (OXPHOS CAFs), myofibroblastic CAFs (myCAFs), neural CAFs, and developmental CAFs (devCAF) (Hypergeometric test with Benjamini-Hochberg FDR correction). ag = antigen. d, Top 200 overexpressed genes of the respective CAF subtypes from (c) are highlighted (scaled mean expression) (two-tailed Wilcoxon Rank Sum test with Benjamini-Hochberg FDR correction) (Supplementary Table 6). e, Gene Set Enrichment Analysis of established CAF subtype signatures in the spatial transcriptomics data of NNMThigh (n = 24) versus NNMTlow (n = 16) ovarian cancer stroma. Gene sets are based on Chhabra and Weeraratna (2023)3 (Supplementary Table 7). FDR = false discovery rate, NES = normalized enrichment score. f, scRNA-seq dataset published by Luo et al.21 (https://gist-fgl.github.io/sc-caf-atlas/; https://creativecommons.org/licenses/by/4.0/). Uniform Manifold Approximation and Projection (UMAP) of cancer types (left panel), cell type annotation (middle panel), NNMT expression (right panel). OVCA = ovarian cancer, PDAC = pancreatic ductal adenocarcinoma, LUAD = lung adenocarcinoma, BLCA = urinary bladder cancer, GC = gastric cancer, BRCA = breast cancer, LC = liver cancer, PRAD = prostate cancer, CRC = colorectal cancer, ICC = intrahepatic cholangiocarcinoma, THCA = thyroid cancer. g, scRNA-seq dataset published by Ma et al.22 (https://chenxisd.shinyapps.io/pancaf/; https://github.com/SGDDNB/ShinyCell (GPL-3.0)). UMAP of cancer types (left panel), CAF subtype annotation (middle panel), NNMT expression (right panel). BRCA = breast cancer, LIHC = liver hepatocellular carcinoma, PRAD = prostate adenocarcinoma, OVCA = ovarian cancer, CRC = colorectal cancer, UCEC = uterine corpus endometrial carcinoma, SMC = smooth muscle cells, iCAF = inflammatory CAFs, pCAF = proliferative CAFs, mCAF = matrix CAFs, meCAF = metabolic CAFs.
Extended Data Fig. 3 Nnmt−/− mice display increased numbers of CD8+ T cells in the tumour.
a, Nnmt−/− mice: Exon 2 of Nnmt and flanking splicing regions were constitutively deleted using CRISPR-Cas9 gene editing technology in mouse zygotes. Knockout was verified by PCR, which showed a 475 base pair (bp) gene product due to exon 2 deletion. P = primer. b, Omental tumour weights 16 days after intraperitoneal (i.p.) injection of ID8 ovarian cancer cells into Nnmt+/+ and Nnmt−/− mice (n = 8 per group) (two-tailed unpaired Student’s t-test). c, ID8 omental tumours from (b) were formalin-fixed and H&E stained. The left panel shows a healthy omentum from a tumour-naïve female C57BL/6 mouse with a milky spot containing immune cells. d, Flow cytometry gating strategy to identify CD4+/CD8+ T cells, F4/80+ macrophages, Ly6G+ neutrophils, and Ly6Chigh monocytes and their PD-L1, C5aR and cytokine expression. e, Splenocytes from tumour-naïve Nnmt+/+ (n = 9) and Nnmt−/− (n = 7) mice were stimulated with PMA/Ionomycin and flow cytometry was performed (two-tailed unpaired Student’s t-test). f, Blood samples were collected from tumour-naïve Nnmt+/+ and Nnmt−/− mice and flow cytometry was performed (n = 6 per group) (two-tailed unpaired Student’s t-test). g, Flow cytometry of subcutaneous (s.c.) MC38 colon cancers from Nnmt+/+ and Nnmt−/− mice. CD8+ T cell cytokine production was quantified after PMA/ionomycin stimulation (n = 6 per group) (two-tailed Mann-Whitney test). h, E0771-LMB cells were administered via retro-orbital intravenous (i.v.) injection into Nnmt+/+ (n = 5) and Nnmt−/− (n = 6). After 12 days, lungs were formalin-fixed, and metastases size was assessed by H&E staining. Mean metastases size represents the average size of the five largest metastases per mouse (two-tailed unpaired Student’s t-test). i, E0771-LMB cells were administered via retro-orbital i.v. injection into Nnmt+/+ and Nnmt−/− mice (n = 9 per group). After 10 days, lungs were stimulated with PMA/Ionomycin and CD8+ T cell cytokine production was quantified using flow cytometry (two-tailed unpaired Student’s t-test). Data are shown as mean ± SEM. One experiment representative of at least two independent experiments is shown for panels f, g, and h.
Extended Data Fig. 4 NNMT is upregulated in the cancer stroma of different murine cancer models.
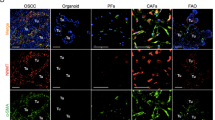
a-f, HGS-2 intrabursal and omental tumours, E0771-LMB mammary fat pad (m.f.p.) tumours or lung metastases following intravenous (i.v.) injection, MC38 subcutaneous (s.c.) tumours, and ID8 intraperitoneal (i.p.) omental tumours were formalin-fixed and immunofluorescently stained for DAPI (blue), fibroblast markers (podoplanin or α-SMA, green), and NNMT (red). The median fluorescent intensity (MFI) of NNMT was measured in tumour-free stroma from the corresponding anatomical site (e.g., ovary or omentum) and compared to the tumour stroma (n = 3 mice per group) (two-tailed unpaired Student’s t-test). g, Nnmt+/+ and Nnmt−/− mice underwent myeloablative irradiation, followed by bone marrow (BM) transplantation from either Nnmt+/+ or Nnmt−/− donor mice (see Fig. 2g, MC38 cancer model). Four weeks after bone marrow transfer, blood samples were collected to confirm successful engraftment by flow cytometry (n = 8 per group, ordinary one-way ANOVA with Tukey’s multiple comparisons test). h, Nnmt+/+ and Nnmt−/− mice underwent myeloablative irradiation, followed by BM transplantation from either Nnmt+/+ or Nnmt−/− donor mice (ID8 cancer model). Eight weeks after bone marrow transfer, blood samples were collected to confirm successful engraftment by flow cytometry (n = 12 mice per group) (ordinary one-way ANOVA with Tukey’s multiple comparisons test). i, Fibroblasts were isolated from the mammary fat pad of tumour-naïve Nnmt−/− mice, and NNMT expression was restored via lentiviral transduction. Western blot showing NNMT in NNMT-expressing mammary fibroblasts, in human CAFs isolated from ovarian cancer samples and Nnmt−/− control cells. For gel source data, see Supplementary Fig. 2. Data are shown as mean ± SEM.
Extended Data Fig. 5 NNMT upregulation in fibroblasts activates complement factor secretion.
a, Immunoblotting of NNMT in NNMT-expressing NIH-3T3 (3T3-NNMT) and control (3T3-CTRL) fibroblasts and immortalized and primary human CAFs isolated from ovarian cancer tissue. b, Gene Set Enrichment Analysis (GSEA) of 3T3-NNMT compared to 3T3-CTRL cells using the Gene Ontology Biological Processes (GOBP) gene set. NES = normalized enrichment scores. c, ID8 omental tumours from Nnmt+/+ and Nnmt−/− mice (n = 7 per group) were analysed by flow cytometry 16 days after intraperitoneal (i.p.) cancer cell injection (two-tailed unpaired Student’s t-test). d, Transwell migration of splenocytes isolated from Nnmt+/+ and Nnmt−/− mice toward conditioned media (CM) collected from 3T3-NNMT and 3T3-CTRL cells (n = 3 technical replicates per group) (ordinary one-way ANOVA with Dunnett’s multiple comparisons test). e, CM was collected after 24 h from 3T3-NNMT, 3T3-NNMT-C3−/− and 3T3-CTRL cells grown in low-methionine media, and the C3 protein levels were measured by ELISA and normalized to total cell protein (n = 3 technical replicates) (Kruskal-Wallis test with Dunn’s multiple comparisons test). f, Subcutaneous co-injection of E0771-LMB breast cancer cells with 3T3-NNMT or 3T3-NNMT-C3−/− fibroblasts into Nnmt+/+ C57BL/6 mice. After nine days, tumours were formalin-fixed and stained for DAPI (blue), Ly6C (green), and PD-L1 (red). Tumour-infiltrating monocytes were quantified using an AI-based algorithm (n = 8 per group) (two-tailed unpaired Student’s t-test). g-h, MC38 subcutaneous (s.c.) tumours of Nnmt+/+ and Nnmt−/− mice were analysed by flow cytometry 12 days after cancer cell injection (n = 8 per group) (two-tailed unpaired Student’s t-test). i, Pearson correlation analysis of C5aR and PD-L1 MFI in Ly6Chigh monocytes of Nnmt+/+ MC38 tumours (n = 8). j, Whole blood from Nnmt+/+ mice and Nnmt−/− mice was drawn ten days after intravenous (i.v.) injection of E0771-LMB cancer cells, and flow cytometry was performed (n = 8 per group) (two-tailed unpaired Student’s t-test). k, Whole blood was drawn from Nnmt+/+ mice and Nnmt−/− mice, and flow cytometry was performed (n = 12 per group) (two-tailed unpaired Student’s t-test). l, Lungs of Nnmt+/+ (n = 7) and Nnmt−/− (n = 8) mice bearing E0771-LMB metastases were analysed by flow cytometry (two-tailed unpaired Student’s t-test). m, Pearson correlation analysis of C5aR and PD-L1 MFI in Ly6Chigh monocytes of lungs of Nnmt+/+ mice bearing E0771-LMB metastases (n = 6). Data are shown as mean ± SEM. One experiment representative of at least two independent experiments is shown for panels a, d, e, and k.
Extended Data Fig. 6 NNMT upregulation in fibroblasts reduces H3K27me3 methylation.
a, scRNA-seq data analysis of CAFs from Luo et al.21, including CAFs from ovarian, breast, colon, and other cancers. Pearson correlation coefficients of log-normalized gene expression values are shown for genes and NNMT across approximately 800,000 single CAFs (visualized using RAWGraphs 2.0). NNMT expression positively correlates with 1,265 genes, with complement factors highlighted in red. b, Fibroblasts were isolated from the ovaries of tumour-naïve Nnmt−/− mice, and NNMT expression was restored via lentiviral transduction. Western blot analysis confirmed NNMT expression in NNMT-expressing ovarian fibroblasts and Nnmt−/− control cells. c, Expression of several CAF markers in NNMT-expressing 3T3 fibroblasts (3T3-NNMT) and control (CTRL) cells was analysed by flow cytometry (n = 3 technical replicates) (two-tailed unpaired Student’s t-test). MFI = mean fluorescence intensity, FMO = fluorescence minus one. d, Integrative Genomics Viewer visualization of H3K27me3 peaks for the CAF markers C3, Mylk, Fbln1, Saa3, and their adjacent genes (Genome Reference Consortium GRCm38, y-axis scale 0-36). e, Cell size (FSC-A) of 3T3-NNMT and 3T3-CTRL cells was measured by flow cytometry (n = 3 technical replicates) (two-tailed unpaired Student’s t-test). f, 3T3-NNMT and 3T3-CTRL cells were grown in either low- or high- L-methionine media for 24 h before RNA extraction, and C3 and Cfb expression levels were measured by qPCR (n = 3 technical replicates) (ordinary one-way ANOVA with Tukey’s multiple comparisons test). Data are shown as mean ± SEM. One experiment representative of at least two independent experiments is shown for panels b, c, e, and f.
Extended Data Fig. 7 High-throughput screening identifies a specific NNMT inhibitor.
a, Overview of the methyltransferase (MTase)-GLO assay used for high-throughput screening of compound libraries to identify an NNMT inhibitor (NNMTi). SAM = S-adenosylmethionine, NAM = nicotinamide, 1-MNA = 1-methylnicotinamide, SAH = S-adenosylhomocysteine. b, Cellular thermal shift assay (CETSA). CAOV3 cells were treated with different concentrations of the hit compound and then heated, lysed, and immunoblotted for NNMT. c, The MTase-GLO assay was used to test non-specific inhibition of related MTases (MT) by the hit compound. PNMT = phenylethanolamine N-methyltransferase, HNMT = histamine N-methyltransferase, GNMT = glycine N-methyltransferase, GAMT = guanidinoacetate N-methyltransferase, COMT = catechol O-methyltransferase. d, Blood samples were collected from Nnmt+/+ or Nnmt−/− mice, and relative 1-MNA levels were measured by mass spectrometry (n = 5 per group) (two-tailed Mann-Whitney test). e, CAOV3 cells were treated with different concentrations of the hit compound. After 24 h, 1-MNA concentrations were measured in the cell supernatant by mass spectrometry (n = 5 technical replicates). f, NNMT and substrates were mixed with different concentrations of NNMTi/NNMTi-D, and the abundance of S-adenosylhomocysteine (SAH) was measured by MTase-GLO (n = 3 technical replicates per group). g, HiBiT-NNMT-expressing K562 cells were treated with different concentrations of NNMTi or NNMTi-D and the HiBiT-NNMT bioluminescent signal was measured by cellular thermal shift assay (CETSA) (n = 2 technical replicates). TS = target stabilization. h, CAOV3 cells were treated with different concentrations of NNMTi (n = 2 technical replicates) or NNMTi-D (n = 1 technical replicate). After 24 h, 1-MNA concentration was measured in the cell supernatant by mass spectrometry. i, Inhibition of 659 protein kinases was measured in the presence of 10 μM NNMTi (Supplementary Table 10) (n = 2 technical replicates). The size of the red dots represents the degree of inhibition level of 329 kinases. TK = tyrosine kinase, TKL = tyrosine kinase-like, STE = serine/threonine kinase, CK1 = casein kinase 1, AGC = protein kinase A, G, and C family, CAMK = calcium/calmodulin-dependent protein kinase, CMGC contains CDKs = cyclin-dependent kinases, MAPKs = mitogen-activated protein kinases, GSKs = glycogen synthase kinases, CLKs = Cdc2-like kinases. The illustration was created using CORAL75. j, Indicated enzymes were treated with 10 μM NNMTi and unspecific enzyme inhibition was measured (n = 2 technical replicates). Left panel: Rxn Biology MTase Panel. Right panel: Eurofin Safety Screen. Data are shown as mean ± SEM. One experiment representative of at least three independent experiments is shown for panels f-h.
Extended Data Fig. 8 Effect of NNMT inhibitor treatment on H3K27me3 methylation, colony formation, proliferation, and migration of several cancer cell lines.
a, ID8-F3, E0771-LMB, OVCAR8, and OVCAR4 cancer cells were treated with DMSO or NNMTi at indicated concentrations in 10 μM L-methionine media for 48–96 h. After 48 h, the treatment media was renewed, and protein lysates were immunoblotted for H3K27me3. b, ID8-F3, MC38, E0771-LMB, and OVCAR8 cancer cells were treated with DMSO or NNMTi at indicated concentrations in 10 μM L-methionine media. The treatment media was renewed every 72 h, and colony formation was assessed after 8–12 days (n = 3 technical replicates). c, ID8-F3, MC38, E0771-LMB, and OVCAR8 cancer cell lines were treated at the indicated concentrations with DMSO, NNMT inhibitor (NNMTi) (left panels) or the less active distomer (NNMTi-D) (right panels) for 2–4 days in 10 μM L-methionine media, and proliferation was assessed (n = 5 technical replicates) (ordinary two-way ANOVA with Dunnett’s multiple comparisons test). d, ID8-F3, MC38, E0771-LMB, and OVCAR8 cancer cells were treated at the indicated concentrations with DMSO, NNMT inhibitor (NNMTi) (left panels) or the less active distomer (NNMTi-D) (right panels) in 10 μM L-methionine media for 24–72 h until the cells reached confluency. Cells were then scratched, and wound width was measured over time (n = 5 technical replicates) (ordinary two-way ANOVA with Dunnett’s multiple comparisons test). Data are shown as mean ± SEM. One experiment representative of at least two independent experiments is shown for panels a-d.
Extended Data Fig. 9 Development of a bioavailable NNMTi.
a, Human ovarian CAFs were treated with either NNMTi (5 µM) or DMSO in 5 µM L-methionine media, and 1-MNA levels were measured by mass spectrometry at the indicated time points (n = 3 technical replicates) (ordinary one-way ANOVA with Šídák’s multiple comparisons test). b, NNMT-expressing and Nnmt−/− control ovarian fibroblasts were treated for 72 h with NNMTi or DMSO. CAF marker expression (α-SMA, left) and cell size (FSC-A, right) were analysed by flow cytometry (n = 3 technical replicates) (ordinary one-way ANOVA with Šídák’s multiple comparisons test). MFI = mean fluorescence intensity. c-d, C57BL/6 mice (n = 3 per group) received a single treatment via oral gavage (p.o.) with varying concentrations of (c) NNMTi or (d) the less active distomer (NNMTi-D), and compound plasma levels were measured by mass spectrometry at the indicated time points. e-f, C57BL/6 mice (n = 3 per group) received a single p.o. treatment with varying concentrations of (e) NNMTi or (f) NNMTi-D, and 1-MNA plasma levels were measured by mass spectrometry. g, Human or murine NNMT, substrates, and assay reagents were mixed with different concentrations of NNMTi, and S-adenosylhomocysteine (SAH) levels were measured by MTase-GLO. Data are shown as mean ± SEM. One experiment representative of three independent experiments is shown for panel b.
Extended Data Fig. 10 NNMT inhibitor treatment reduces tumour burden and metastasis, and decreases the abundance of established CAF subtypes in vivo.
a, Growth curve of orthotopic mammary fat pad (m.f.p.) E0771-LMB breast tumours treated with NNMTi (n = 9) or vehicle (n = 10). b, Primary tumour weights after intrabursal injection of BPPNM ovarian cancer cells and intraperitoneal (i.p.) treatment with NNMTi or vehicle (n = 8 per group). c, MC38 subcutaneous (s.c.) tumours were treated once by intratumoral (i.t.) NNMTi, NNMTi-D or vehicle injection 13 days after cancer cell injection. Tumours were removed 8 h after treatment, and tumour 1-MNA levels measured (n = 4 mice per group) (ordinary one-way ANOVA with Šídák’s multiple comparisons test). d, E0771-LMB breast cancer cells were injected via retro-orbital intravenous (i.v.) route into C57BL/6 mice (day 0). Mice were treated by oropharyngeal inhalation (o.p.) with either NNMTi, NNMTi-D, or vehicle. 13 days post-injection, metastasis size was assessed by H&E staining. Mean metastasis size represents the average of the five largest lesions per mouse (n = 5 mice per group) (ordinary one-way ANOVA with Dunnett’s multiple comparisons test). e, Growth curve of s.c. MC38 tumours in Nnmt−/− mice, treated with NNMTi (n = 8) or vehicle (n = 7). f, Gating strategy for flow cytometry to identify CAFs. g, Flow cytometry of m.f.p. E0771-LMB tumours treated with NNMTi (n = 10) or vehicle (n = 8), as shown in (a). Result for H3K27me3low CAFs is shown. h, Flow cytometry of intrabursal HGS-2 tumours treated with NNMTi or vehicle, as shown in (a) (n = 8 per group). i, Flow cytometry was performed 18 days after injection of orthotopic m.f.p. E0771-LMB breast tumours treated with NNMTi or vehicle via i.p. injection (n = 9 per group). j, Omental tumours were H&E stained 19 days after intrabursal injection of HGS-2 ovarian cancer cells and treatment with NNMTi (n = 10) or vehicle (n = 7). An AI-based algorithm analysed average CAF density (left) and size (right). CAFs are outlined in red. Data are shown as mean ± SEM. One experiment representative of at least two independent experiments is shown for panels d and i. Panel a, e (two-way ANOVA with Šídák’s multiple comparisons test); b, g, h, i, j (two-tailed unpaired Student’s t-test).
Extended Data Fig. 11 NNMT inhibitor treatment increases CD8+ T cell activation in vivo and enhances immunotherapy efficacy.
a-b, MC38 s.c. tumours treated i.t. with NNMTi or NNMTi-D were analysed by flow cytometry 15 days after cancer cell injection. Uniform Manifold Approximation and Projection (UMAP) visualizes marker intensity (a) and a total of 240,000 CD45+ immune cells (b, top panel), consisting of 120,000 cells from NNMTi-treated tumours and 120,000 from NNMTi-D-treated tumours (b, bottom panels). c, CD8+ T cell (blue) and Ly6Chigh PD-L1high monocyte clusters (red) are highlighted in the UMAP from panel (b). The proportions of Ly6Chigh PD-L1high monocytes and CD8+ T cells (blue) were quantified for each tumour (n = 6 per group) (two-tailed unpaired Student’s t-test). d, MC38 tumours treated as described in (a-b) were stimulated with PMA/ionomycin, and cytokine production was quantified by flow cytometry (conventional gating) (n = 8 per group) (two-tailed unpaired Student’s t-test). e, Flow cytometry of orthotopic HGS-2 tumours treated with NNMTi or vehicle (n = 8 per group). CD8+ T cell cytokine production was quantified after PMA/ionomycin stimulation. f, Growth curve of MC38 s.c. tumours in Nnmt+/+ and Nnmt−/− mice treated with anti-PD-1 or IgG control (n = 6 mice per group) (ordinary two-way ANOVA with Tukey’s multiple comparisons test). g, Growth curve of orthotopic m.f.p. E0771-LMB breast cancers treated with NNMTi or NNMTi-D and anti-PD-1 or IgG control (n = 9 or 10 per group) (ordinary two-way ANOVA with Šídák’s multiple comparisons test). h, Growth curve of orthotopic m.f.p. E0771-LMB breast cancers treated with NNMTi or NNMTi-D and anti-CD47 or IgG control (n = 9 or 10 per group) (ordinary two-way ANOVA with Tukey’s multiple comparisons test). Data are shown as mean ± SEM.
Supplementary information
Supplementary Information (download PDF )
Supplementary Methods (synthesis, purity and chiral analyses of NNMTi (NCGC00685960) and NNMTi-D (NCGC00685961)), and Supplementary Figs. 1 and 2.
Supplementary Tables (download XLSX )
Supplementary Tables 1–12.
Source data
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Heide, J., Bilecz, A.J., Patnaik, S. et al. NNMT inhibition in cancer-associated fibroblasts restores antitumour immunity. Nature 645, 1051–1059 (2025). https://doi.org/10.1038/s41586-025-09303-5
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41586-025-09303-5
This article is cited by
-
Cross-talk between cancer-associated fibroblasts and dormant cancer cells: current status and promising therapeutic potential
Cancer Cell International (2025)
