
Abstract
Celiac disease (CeD) is an autoimmune disorder triggered by dietary gluten. While HLA-DQ2/8-mediated presentation of gliadin peptides is required for disease, the mechanisms that underlie the loss of oral tolerance to gluten remain incompletely understood. Long-noncoding RNAs (lncRNAs) have been increasingly recognized as regulators of immune function, yet their role in oral tolerance has not been previously explored. Here, using a screen designed to identify lncRNAs responsive to T cell activation and enriched for CeD-associated GWAS variants, we identified lnc13 as a top candidate. In HLA-DQ8 transgenic mice lacking lnc13, unmanipulated gluten ingestion led to molecular signatures resembling human CeD and hallmark features of loss of oral tolerance to gluten: increased IFN-γ+ lymphocytes, IL-12+ myeloid cells, cytotoxic intraepithelial immune cells and crypt hyperplasia in the small intestine. Mechanistically, lnc13 binds specific DNA regulatory regions and limits immune cell responsiveness to proinflammatory signals. In particular, lnc13 restrains IL-15-driven differentiation of CD8+ natural killer-like lymphokine-activated killer cells (an IL-15-dependent pathway strongly implicated in CeD). These findings establish lnc13 as a critical noncoding modulator of oral gluten tolerance.
Similar content being viewed by others

Main
Dietary antigens such as wheat gluten are typically well tolerated. In patients with celiac disease (CeD), however, peptides derived from gliadin provoke chronic immune activation that leads to small intestinal mucosal damage and systemic inflammation1,2,3,4,5. While the human leukocyte antigens (HLA) DQ2 and DQ8 are necessary for antigen presentation to T cells in CeD, these alleles alone are not sufficient for disease development4,6,7. The underlying mechanisms that govern the breakdown of oral gluten tolerance remain incompletely defined.
Genome-wide association studies (GWAS) have identified numerous non-HLA single-nucleotide polymorphisms (SNPs) linked to CeD risk, with ~94% of variants mapping to noncoding regions of the genome3,8,9. It is now recognized that much of the genome is pervasively transcribed into noncoding RNAs, including long-noncoding RNAs (lncRNAs), which share many features with mRNAs (for example, 5′ caps, splicing and polyadenylation) but do not encode proteins10,11. Approximately 100,000 lncRNA genes have been annotated in humans, vastly outnumbering protein-coding genes12.
LncRNAs have been implicated in diverse regulatory processes across development, immunity and disease13. SNPs located within lncRNA loci can disrupt their expression, structure or molecular interactions, leading to altered regulatory function13,14,15,16,17,18. Notably, GWAS SNPs are disproportionately enriched in lncRNAs compared to nontranscribed intergenic regions, suggesting that noncoding variants may exert phenotypic effects through RNA-based mechanisms10.
Previously, we identified lnc13, a CeD-associated lncRNA that regulates inflammatory gene expression in macrophages and harbors a CeD-associated SNP haplotype block16; however, lnc13’s physiological function in oral tolerance (particularly in vivo) has remained unknown due to the lack of a knockout model.
In this study, we screened lncRNAs upregulated in gliadin-specific T cells and prioritized those overlapping CeD GWAS SNPs. We identified lnc13 as a highly conserved, inducible regulator. To test its physiological function in the context of oral gluten tolerance, we generated an lnc13 knockout mouse and crossed it with HLA-DQ8 transgenic mice lacking endogenous major histocompatibility complex (MHC) class II. Upon gluten exposure, lnc13−/−-DQ8 mice exhibited hallmark immunological and histological features of CeD. Mechanistically, we show that lnc13 represses responsiveness of CD8+ intraepithelial lymphocytes (IELs) to IL-15, a cytokine implicated in CeD pathology. These findings highlight lnc13 as a key genetic regulator of gluten tolerance and immune restraint in the gut.
Results
Lnc13 is a candidate regulator of oral gluten tolerance
Gliadin peptides from ingested gluten are presented by antigen-presenting cells on HLA-DQ2 or HLA-DQ8, leading to activation of gliadin-specific T cells6. If left unchecked, these hyperactivated T cells initiate a cascade of inflammatory responses that culminate in tissue pathology4. We reasoned that regulation of T cells is a critical component of oral gluten tolerance and therefore we hypothesized that lncRNA candidates would (1) respond to mucosal T cell activation and (2) contain CeD-associated GWAS SNPs.
To identify such lncRNAs, we profiled RNA expression in gliadin-specific T cells from human CeD small intestines stimulated via the TCR. This analysis revealed 449 lncRNAs that were significantly upregulated and 168 that were downregulated. Notably, only three of these differentially expressed lncRNAs overlapped with CeD GWAS SNPs: IL21-AS1, LINC02929 and lnc13, all of which were upregulated following stimulation (Fig. 1a).

a, Volcano plot showing differentially expressed long-noncoding RNAs (lncRNAs) in gluten-specific CD4+ T cells from human small intestine activated by TCR for 3 h (data from GSE146441). n = 22 donors. lncRNAs upregulated (red) or downregulated (blue) are highlighted (abs|log2 fold change (FC)| > 2, adjusted P < 0.01). CeD-associated GWAS lncRNAs are shown in black. b, Cross-species sequence conservation (human-to-mouse alignment) of the three top candidate lncRNAs overlapping CeD-associated GWAS loci: IL21-AS1, LINC02929 and lnc13. c,d, Expression of lnc13 (C; P = 9.44537 × 10−7) (c) and its RNA-degrading enzyme DCP2 (D; P = 5.69524E-10) (d) in human gluten-specific CD4+ T cells at 0 and 3 h post-TCR stimulation (GSE146441). n = 22 donors. e, Time-course expression of human lnc13 in CD4+ T cells over 5 days of TCR activation (GSE96538), showing peak expression at days 3–4. n = 3 donors. P = 0.00078864. f, ChIP-seq analysis showing TF binding upstream of lnc13 in T cells. Middle: ReMap atlas of TF peaks; bottom: RELA ChIP-seq tracks in resting and 3-h-stimulated CD4+ T cells (GSE99319), indicating NF-κB recruitment. g, RNA expression of lnc13 in CD4+ T cells from WT and Rela-conditional knockout mice following 4 h of TCR stimulation (GSE239700), demonstrating NF-κB dependence. n = 4. P = 0.0017125. Differential expression (a,c–e,g) was computed using DESeq2 with adjusted P values that reflect Benjamini–Hochberg multiple comparisons correction. P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. In scatter-plots (c–e,g), lines represent mean ± s.e.m.
Among these, LINC02929 (an uncharacterized transcript) exhibited the smallest fold change. IL-21-AS1, a promoter antisense lncRNA, has been studied in other autoimmune diseases, where it regulates IL-21 expression in cis13,19; however, IL21-AS1 showed the least evolutionary conservation between humans and mice (Fig. 1b), diminishing its suitability for in vivo modeling.
In contrast, lnc13 fulfilled all key criteria: it harbored a CeD-associated haplotype block, was robustly induced upon gliadin-specific T cell activation and exhibited high sequence conservation across species. We therefore prioritized lnc13 as the leading candidate for further investigation.
Lnc13 levels increased markedly following 3 h of gliadin-specific T cell stimulation, while its degrading enzyme DCP2 (ref. 16) decreased in parallel (Fig. 1c,d and Extended Data Fig. 1a). Human T cells showed delayed upregulation of lnc13, with peak expression (21.5-fold increase) occurring 3–4 days after TCR activation (Fig. 1e). This pattern of expression was largely inversely correlated with DCP2 and resembled the late-phase kinetics of FOXP3, a key regulator of immune restraint (Extended Data Fig. 1b,c). By contrast, proinflammatory transcripts such as TBX21 and IFNG exhibited earlier or mid-phase induction (Extended Data Fig. 1d,e). Both gluten-specific T cell and general peripheral T cell TCR stimulation upregulated lnc13 to similar magnitudes and kinetics (Extended Data Fig. 1f).
To understand how lnc13 is transcriptionally regulated, we examined transcription factor (TF) binding at its promoter using ChIP-seq datasets. We identified multiple inducible TF peaks, notably for RELA, a member of the NF-κB family, which binds near a potential lnc13 upstream regulatory element following T cell activation (Fig. 1f). Given that NF-κB activity is elevated in CeD and broadly pleiotropic, its downstream effectors such as lnc13 may represent more selective therapeutic targets20,21. Indeed, conditional deletion of Rela in activated T cells abolished lnc13 expression (Fig. 1f), confirming that lnc13 is an NF-κB-dependent gene induced during T cell activation. Overall, we identified lnc13 as a top regulator of oral gluten tolerance that is dynamically induced by NF-κB upon T cell activation.
lnc13 loss breaks gluten tolerance in HLA-DQ8 mice
The lnc13 transcript harbors the CeD-associated haplotype block marked by lead SNP rs917997 (ref. 16). The risk allele of rs917997 alters lnc13’s secondary structure and impairs its regulatory function in macrophages16. While lnc13 is highly conserved between humans and mice, the immediate sequences surrounding the SNPs of this haplotype block do not align perfectly between species, precluding precise allele modeling in mice. To mimic the systemic loss-of-function effect of the human SNP, we generated a full lnc13 knockout (KO) mouse (Extended Data Fig. 2a and see Supplementary Table 2 for Cas9 gRNA sequences).
Lnc13 is located near Il18rap but exists as an independent transcript, and its genomic locus lacks enhancer-associated chromatin features (Extended Data Fig. 2b–d). In bone-marrow-derived macrophages (BMDMs), lnc13 is downregulated upon LPS stimulation, and its deletion does not alter Il18rap expression under either resting or activated conditions (Supplementary Fig. 1a,b). These findings confirm that the KO is specific and does not disrupt neighboring gene expression. Notably, lnc13−/− BMDMs showed heightened expression of inflammatory genes such as Il6, Il1b and Nos2 following LPS stimulation (Supplementary Fig. 1c), suggesting that lnc13 restrains inflammation.
To test whether lnc13 regulates oral tolerance to gluten, we crossed the lnc13 KO onto HLA-DQ8 transgenic mice, which express the human DQ8 allele under its native promoter and lack endogenous murine MHC class II; thus every CD4+ T cell TCR in the mouse is educated against HLA-DQ8 (refs. 22,23) (Supplementary Fig. 2a,b). These mice were weaned on a gluten-free diet and switched at 10 weeks old to standard chow containing gluten (Fig. 2a). To normalize exposure, all mice were additionally gavaged with 20 mg of unmanipulated gliadin every other day for 45 days, similar to previously published protocols22,24.

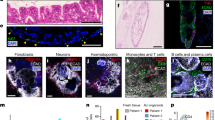
a, Schematic of the gluten challenge protocol in HLA-DQ8 transgenic mice. Mice were weaned onto a gluten-free diet, then switched to standard chow and gavaged with 20 mg gliadin every other day for 45 days. b,c, Relative RNA expression of type 1 cytokines Ifng (b; P = 0.00351387) and Il12b (c; P = 0.00207621) in terminal ileum tissue from gluten-free or gluten-challenged mice, measured by RT–qPCR. Relative RNA levels are calculated in reference to Rpl32 housekeeping gene. n = 3–6 mice, across four independent experiments. d, lnc13 expression in terminal ileum from gluten-free versus gluten-challenged DQ8 mice, showing decreased expression upon gluten exposure. Relative RNA levels are calculated in reference to Rpl32 housekeeping gene. n = 3–6, across four independent experiments. P = 0.00023608. e–g, Representative flow cytometry plots (left) and quantification (right) showing frequencies of IFN-γ+ cells from live CD45+TCRb+B220− cells, where quantifications show those further gated on CD8+CD4− (P = 0.00013518) or CD4+CD8− (P = 0.00047811) (e), frequencies of IL-12+CD11b+ cells from live CD45+TCRb−B220− cells (P = 0.00031403) (f) and frequencies of FOXP3+ cells (P = 0.03630467) from live CD45+TCRb+B220−CD4+CD8− cells (g) in the small intestinal lamina propria (Lp) of gluten-challenged lnc13−/−-DQ8 and WT-DQ8 mice. n = 6 mice, across four independent experiments. h, Pathway enrichment analysis of DEGs in whole ileum (‘tissue’) and Lp lymphocytes (LPLs) from lnc13−/−-DQ8 versus WT-DQ8 mice after gluten challenge, and duodenal biopsies from patients with CeD versus healthy controls (GSE134900). Shared pathways include immune response and cytokine-mediated signaling. i, Expression of selected proinflammatory and B cell-associated genes in the ileum of lnc13−/−-DQ8 versus WT-DQ8 mice by RT–qPCR. Relative RNA levels (ΔΔCt) are calculated in reference to Rpl32 and then normalized to the average of WT-DQ8 samples. n = 6 across three independent experiments. P values for Il1a, Il21, Csf2, Il2, Tnfa, Il27, Il13, Il1b, Il17a, Hprt, Il7, Cd138 and Blimp1 are 0.006489, 0.004429, 0.001036, 0.000873, 0.030904, 0.023022, 0.0517, 0.0078, 0.035474, 0.299992923, 0.61115255, 0.20111989 and 0.02767208, respectively. An unpaired, two-tailed, Student’s t-test was used for comparison (b–g,i). *P < 0.05,**P < 0.01, ***P < 0.001, NS, not significant. In scatter-plots (b,c,e–g,i), samples from lnc13−/−-DQ8 (KO-DQ8) mice are represented by red dots and those from lnc13+/+-DQ8 (WT-DQ8) mice are represented by blue dots. Lines represent mean ± s.e.m. Image of mouse in a generated by BioRender; Yang-Fischer, R. https://biorender.com/hz505m6 (2026).
The mucosa of patients with CeD is associated with a type 1 immune profile, characterized by IFN-γ and IL-12 (ref. 4). Remarkably, in the small intestines of lnc13−/−-DQ8 mice, gluten challenge led to increased expression of Ifng and Il12b (Fig. 2b,c). Notably, lnc13 levels were decreased in gluten-challenged wild-type (WT)-DQ8 mice compared to gluten-free controls (Fig. 2d), mirroring findings in biopsies from patients with CeD16. Because CeD arises from coordinated interactions among CD4+ helper T (TH) cells, cytotoxic CD8+ IELs, B cells and myeloid cells4, we next examined whether lnc13 deficiency affected each of these key immune populations involved in gluten-driven inflammation.
Flow cytometric analysis revealed that gluten-challenged lnc13−/−-DQ8 mice exhibited increased frequencies of IFN-γ+CD4+ and CD8+ T cells in the small intestinal lamina propria compared to WT-DQ8 controls (Fig. 2e and Supplementary Fig. 3a), consistent with a TH1-skewed inflammatory response. We also observed an increase in IL-12-producing CD11b+ myeloid cells (Fig. 2f and Supplementary Fig. 3b), in agreement with independent human studies that have reported enrichment of IL-12 signaling pathways in lamina propria dendritic cells and macrophages from patients with active celiac disease, as well as elevated circulating IL-12 levels25,26. Indeed, human patients with CeD express increased levels of IL12Aand IL12B transcripts (Supplementary Fig. 3c). Further profiling identified increased IL-12 production within dendritic cells, a population previously implicated in breaking gluten tolerance27 (Supplementary Fig. 3a,d). Together with established evidence that IL-12 is predominantly produced by myeloid cells, these findings support the biological relevance of increased IL-12 production observed in our model.
In contrast to observations in human CeD, where increased numbers of FOXP3+ regulatory T (Treg) cells have been reported in the lamina propria28, we observed a reduction in FOXP3+ Treg cells in the lamina propria of gluten-challenged lnc13−/−-DQ8 mice (Fig. 2g). This difference likely reflects distinctions between reductionist mouse models of oral tolerance and the complex, polygenic nature of human disease, where Treg cell expansion may represent a compensatory but functionally impaired response28,29. Notably, multiple mouse models of loss of oral tolerance have similarly reported diminished or failed Treg cell expansion as a hallmark of disease induction, underscoring the relevance of this phenotype in experimental settings27,30,31,32. Together, these findings indicate that lnc13 is essential for maintaining immune homeostasis in response to dietary gluten.
To assess global molecular changes, we performed RNA sequencing (RNA-seq) on whole ileal tissue and lamina propria lymphocytes (LPLs) from gluten-challenged WT and lnc13−/−-DQ8 mice. Pathway analysis revealed substantial overlap between differentially expressed genes (DEGs) in lnc13-deficient mice and those in human CeD biopsies (Fig. 2h). We also observed increased expression of proinflammatory cytokines implicated in CeD, such as Il1a, Il1b, Il21, Csf2, Il2, Il27, Il13 and Tnfa (Fig. 2i). Cross-species DEG correlation confirmed strong similarity between the lnc13−/−-DQ8 mouse and human CeD transcriptomes, especially for inflammatory genes like Cxcl5, Saa2 and Oas2 (Extended Data Fig. 3a).
In the LPL compartment, loss of lnc13 was associated with upregulation of B cell activation and antibody secretion pathways (Fig. 2h). Consistent with this, we observed an elevated expression of Blimp1 and Sdc1 (CD138), markers of plasma cells (Fig. 2i). Immunoglobulin (Ig) locus genes were similarly upregulated in both lnc13-deficient mice and patients with CeD (Extended Data Fig. 3b). Serologically, lnc13−/−-DQ8 mice produced higher levels of anti-gliadin, anti-deamidated gliadin and anti-tTG (Extended Data Fig. 3c), hallmarks of humoral autoimmunity in CeD.
Altogether, these data demonstrate that lnc13 is critical for maintaining gluten tolerance and preventing the inflammatory responses that characterize CeD.
Lnc13 restrains cytotoxic intraepithelial lymphocytes
In CeD, the breakdown of oral tolerance leads to epithelial inflammation, driven in part by cytotoxic IELs4. To evaluate whether loss of lnc13 promotes this pathology, we analyzed IELs isolated from the small intestines of gluten-challenged WT-DQ8 and lnc13−/−-DQ8 mice.
RNA-seq of bulk IELs revealed several immune-related pathways uniquely altered in this compartment, including those related to defense response to bacteria and interspecies interaction, none of which was prominent in lamina propria datasets (Figs. 2h and 3a). These changes suggested a distinct, lnc13-dependent inflammatory environment within the epithelium.

a, Pathway enrichment analysis of DEGs from whole small intestinal tissue (‘tissue’) and IEL fraction of lnc13−/−-DQ8 versus WT-DQ8 mice after gluten challenge and duodenal biopsies from patients with CeD versus healthy controls (GSE134900). Enriched pathways in IELs include antibacterial defense, interspecies interaction and immune response. Scale bar shows −log10(FDR) of enriched pathways in each dataset. b, Representative flow cytometry plots (left) and quantification (right) showing increased frequencies of CD8α+CD45+ cells of total live cells in the IEL compartment of lnc13−/− versus WT mice. n = 4, P = 0.00121286. c–i, Elevated cytotoxicity markers shown in various IEL populations: GZMB expression (c; P = 3.50839 × 10−6 for %, P = 0.00027095 for MFI) and CD107a (LAMP-1)+ CD107b (LAMP-2)+ double-positive (d, P = 0.00021012) cells of live CD45+B220− TCRgd−TCRβ+CD8α+CD8β+ IELs; GZMB expression of live CD45+B220−TCRgd−TCRβ−NKG2D+ intraepithelial (IE) NK cells (e, P = 0.00477969 for %, P = 0.00477969 for MFI); CD107a+CD107b+ double-positive cells of live CD45+B220−TCRgd−TCRβ+CD8α+CD8β− IELs (f, P = 0.00161301); T-bet expression of live CD45+B220−TCRgd−TCRβ−NK1.1+NKG2D+ intraepithelial NK cells (g, P = 0.00308031 for %, P = 0.00205277 for MFI); and NKG2D+ cells of live CD45+B220−TCRgd−TCRβ+CD8α+CD8β+ IELs (h, P = 0.00181557). i, % GZMB+NKG2D+ double-positive CD8ab+ T cells (P = 0.02508331). For c,e,g, representative histogram plots are shown on the left with dashed lines denote isotype controls, red histogram represent cells from lnc13−/−-DQ8 mice and blue histogram represent cells from WT-DQ8 mice. The middle section of these panels show quantification of cells expressing these markers, with gating based on isotype controls. On the right section of these panels, quantification of MFI is shown. For d,f,h, representative pseudocolor plots are shown on the left, while quantifications of marker-positive cells are shown on the right. Across scatter-plots in c–i, samples from lnc13−/−-DQ8 (KO-DQ8) mice are represented by red dots and those from lnc13+/+-DQ8 (WT-DQ8) mice are represented by blue dots. n = 6 biological replicates, across four experiments. j, Volcano plot of DEGs from sorted CD8αβ+TCRαβ+ IELs post-gluten challenge. Top upregulated genes include T cell activation markers (Phlda1 and Atf3), homing receptors (S1pr1, S1pr3 and Ccr7), co-activation receptors (Tnfrsf4/OX40, Tnfsf11/RANKL, Tnfrsf9/4-1BB and Il1r2) and NK-like genes (Klra7, Klrb1b and Klri1). Differential expression was computed using DESeq2 to derive P values. k, Pathway enrichment of CD8αβ+ T IEL DEGs from lnc13−/−-DQ8 versus WT-DQ8 mice after gluten challenge (n = 3 mice), alongside pseudobulk DEG profiles from human patient with CeD CD8+ T cells versus controls, and human ‘natural effector’ (NE) CD8+ T cells versus all other CD8+ subsets (GSE252762). Shared pathways include T cell activation, proliferation and lymphomagenesis. Unpaired, two-tailed, Student’s t-test was used for comparison (b–i). *P < 0.05,**P < 0.01, ***P < 0.001, ****P < 0.0001; lines represent mean ± s.e.m.
Consistent with a CeD-like phenotype, CD8+ T cells were markedly increased in the epithelium of lnc13-deficient mice (Fig. 3b), along with an expansion of CD8αβ+TCRαβ+ IELs expressing GZMB and CD107a/b (LAMP-1/2), markers of cytotoxic granule release (Fig. 3c,d and Supplementary Fig. 4). These CD8αβ+ cytotoxic IELs are strongly implicated in CeD tissue damage4. We also detected elevated inflammatory marker expression in other IEL subsets, including: CD107a/b in CD8αα+ T cells (Fig. 3f) and T-bet and GZMB in NK cells (Fig. 3e–g).
CD8+ IELs in CeD often acquire an NK-like lymphokine-activated killer (LAK) phenotype, characterized by expression of NKG2D33. In lnc13−/−-DQ8 mice, we observed increased frequencies of NKG2D+CD8αβ+ IELs and of dual NKG2D+GZMB+ cytotoxic IELs (Fig. 3h,i), mirroring pathogenic IEL profiles seen in patients with CeD.
To investigate transcriptional signatures underpinning these phenotypes, we performed RNA-seq on sorted CD8αβ+ IELs from gluten-challenged WT and lnc13-deficient DQ8 mice (Fig. 3j and Supplementary Table 3). Genes most differentially upregulated in lnc13−/− CD8+ IELs included activation markers (Phlda1 and Atf3), T cell trafficking receptors (S1pr1, S1pr3 and Ccr7), critical T cell co-activation receptors (Tnfsf11/RANKL, Tnfrsf4/OX40, Tnfrsf9/4-1BB and Il1r2), cell-cycle regulator (Cdkn3) and NK-associated genes (Klra7, Klri1 and Klrb1b). Pathway analysis indicated enhanced signatures of T cell proliferation, activation and lymphomagenesis (Fig. 3j), the latter of which aligns with the known risk of enteropathy-associated T cell lymphoma in CeD34.
To assess translational relevance, we compared our murine transcriptomes to published single-cell RNA-seq of human small intestinal CD8+ T cells28. Gene expression changes in lnc13−/− CD8αβ+ IELs closely mirrored those found in natural effector CD8+ T cells from patients with active CeD, which are absent in healthy controls or gluten-free treated patients (Fig. 3k). Together, these data demonstrate that lnc13 restrains the differentiation and activation of cytotoxic and inflammatory IELs, and that its loss drives the emergence of a CeD-like intraepithelial immune landscape28,33.
We next performed a systematic characterization of immune cell populations and cytokine profiles. In peripheral blood, cytokine analysis revealed elevated TNF and IFN-γ, whereas CRP and IL-6 remained unchanged, consistent with a type 1-skewed but nonsystemically inflamed state (Extended Data Fig. 4a–d). This distinction is expected, as lnc13 represents only one CeD-associated locus, whereas human CeD reflects the combined effects of many genetic and environmental risk factors3. Peripheral immune alterations were modest and largely not biologically meaningful, with minimal changes in circulating IFN-γ+ T cells and preserved T cell development (Extended Data Fig. 4e–u). Thus, lnc13 deficiency does not drive systemic autoimmunity but instead results in mucosa-restricted inflammation, mirroring human CeD.
Overall we identify that lnc13 is necessary to restrain expansion of cytotoxic and inflammatory intraepithelial lymphocytes.
Loss of lnc13 leads to CeD-like pathology in mice
Although HLA-DQ8 transgenic mice mount an immune response to gluten, they typically do not develop overt small intestinal pathology in response to dietary gluten alone23,27. Given the pronounced immune activation observed in lnc13−/−-DQ8 mice, we hypothesized that lnc13 deficiency might provide an additional inflammatory stimulus to trigger celiac-like histological changes.
This hypothesis is supported by a previous study showing that transgenic overexpression of IL-15 in DQ8 mice can sensitize the intestinal mucosa to gluten-induced injury, leading to recapitulation of CeD pathology22. Of note, while CeD in humans primarily affects the proximal small intestine, gluten is thought to accumulate in the distal small intestine of mice, such as the terminal ileum, due to differences in gastrointestinal physiology and gluten metabolism24,35.
We therefore first examined the terminal ileum. After 45 days of gluten exposure, lnc13−/−-DQ8 mice exhibited significantly lower villus-to-crypt ratios compared to WT-DQ8 controls (Fig. 4a–c). Of note, we observed crypt hyperplasia, a hallmark of early CeD pathology that reflects ongoing mucosal inflammation and epithelial remodeling (Fig. 4d). Consistent with CeD-like IEL infiltration, crypt hyperplasia was accompanied by a significant increase in IEL numbers, which inversely correlated with the villus-to-crypt ratio (Fig. 4e,f). The increased IEL numbers agrees with our earlier flow cytometry result showing increased frequency of CD8+ IELs in mice lacking lnc13 (Fig. 3b).

a, Representative H&E-stained histological sections of terminal ileum from gluten-challenged WT-DQ8 and lnc13−/−-DQ8 mice. Crypt thickness indicated by black brackets. Black scale bar, 200 μm. b,c, Quantification of: villus-to-crypt (V/C) ratios (n = 10–12 mice, across five independent experiments; P = 0.00073755) (b) and villous atrophy (VA) scores in terminal ileum gut bundles (n = 5–9 mice; P = 0.02808226) (c). VA scoring: 0, no atrophy; 1, mild; 2, clear atrophy. Blinded scoring performed by a pathologist. Only samples from (A) that had sufficient coverage for a confident call by the pathologist were scored. d, Measurement of crypt depth in terminal ileum, showing crypt hyperplasia in lnc13−/−-DQ8 mice (n = 10–12 mice, across five independent experiments; P = 0.0054398). e, EL counts per 100 intestinal epithelial cells (IECs) in terminal ileum, increased in lnc13−/−-DQ8 mice (n = 10–12 mice, across five independent experiments). f, Correlation between V/C ratio and IEL density across individual mice. Pearson two-tailed correlation test showing inverse correlation of IEL counts per 100 IECs and V/C ratio. n = 22 mice. P = 6.54842 × 10−5. g,h, Swiss roll histological quantification in distal small intestines (ileum) (g; P = 0.00030838 for V/C ratio; P = 0.04160064 for crypt length) and proximal small intestines (jejunum) (h; P = 0.00023621 for V/C ratio; P = 0.0441443 for crypt length): left, V/C ratio; middle, villus height; right, crypt depth. Analysis confirms crypt hyperplasia extends into proximal small intestine in gluten-challenged lnc13−/−-DQ8 mice (n = 3–10 mice). i, Schematic of gluten withdrawal protocol to assess pathology reversibility. j, Quantification of V/C ratio in gluten-challenged mice switched to a gluten-free diet. Gluten-induced pathology in lnc13−/−-DQ8 mice is reversible (n = 6–8). Statistical comparisons: unpaired, two-tailed Student’s t-test. In scatter-plot, lines represent mean ± s.e.m., except c (median). *P < 0.05,**P < 0.01, ***P < 0.001, ****P < 0.0001. Image of mouse in i generated by BioRender; Yang-Fischer, R. https://biorender.com/hz505m6 (2026).
We next used Swiss roll sections of the entire small intestine, enabling unbiased histological analysis across the proximal-to-distal axis. Using this approach, we found that crypt hyperplasia extended into the proximal small intestine of lnc13−/−-DQ8 mice (Fig. 4g,h and Supplementary Fig. 5). This pathology was observed only in mice expressing HLA-DQ8, challenged with gluten, and lacking lnc13, supporting a model in which these three factors together are required to induce histological features of CeD.
In humans, crypt hyperplasia typically precedes villous atrophy, which emerges after prolonged inflammation36. Thus, the changes we observed in lnc13−/−-DQ8 mice after 45 days of gluten exposure likely represent early-stage CeD-like pathology. Finally, we tested whether these changes were reversible. Upon switching mice to a gluten-free diet, we observed resolution of histological abnormalities (Fig. 4i,j), meeting a critical standard for CeD mouse modeling24.
To determine directly whether the immune phenotypes observed in gluten-challenged lnc13−/−–DQ8 mice require gluten exposure, we performed a full immune characterization of WT-DQ8 and lnc13−/−–DQ8 mice maintained on a gluten-free diet with sham gavage for 45 days (Extended Data Fig. 5a). Across all compartments examined, we did not observe any differences between genotypes. In the lamina propria, Treg cell frequencies were unchanged (Extended Data Fig. 5b), and the percentages of IFN-γ+CD8+ and CD4+ T cells (Extended Data Fig. 5c) were comparable between WT-DQ8 and KO-DQ8 mice. Similarly, among IELs, we detected no differences in GZMB+CD8αβ+ T cells (percent or mean fluorescence intensity (MFI); Extended Data Fig. 5d), GZMB+ NK cells (Extended Data Fig. 5e), CD107a/b+CD8αβ+ IELs (Extended Data Fig. 5f), CD107a/b+CD8αα+ IELs (Extended Data Fig. 5g) or NKG2D+CD8+ IELs (Extended Data Fig. 5h). Peripheral lymph nodes likewise showed no differences in IFN-γ+CD8+ (Extended Data Fig. 5i) or IFN-γ+CD4+ T cells (Extended Data Fig. 5j), nor in FoxP3+ Treg cell frequencies (Extended Data Fig. 5k).
We also performed a full immune characterization in WT-DQ8 and lnc13−/−-DQ8 mice that was first challenged with gluten, and then reverted to a gluten-free diet for 45 days with sham treatments every other day (Extended Data Fig. 6a). Similarly, mice challenged with gluten and subsequently reverted to a gluten-free diet exhibited normalization across all immune compartments examined (Extended Data Fig. 6), including IEL activation markers, lamina propria effector T cells and peripheral T cell subsets.
Together, these results demonstrate that the immune abnormalities observed during gluten challenge (including heightened IEL activation, increased effector function and reduced Treg cell frequencies) fully normalize when gluten is removed. Thus, the immunopathology in lnc13−/−-DQ8 mice is gluten-dependent rather than an intrinsic or persistent consequence of lnc13 deficiency. These findings demonstrate that loss of lnc13 in the context of HLA-DQ8 and dietary gluten induces key features of CeD, including crypt hyperplasia, IEL infiltration and gluten dependence with reversibility on a gluten-free diet.
Lnc13 restrains CD8αβ+ IEL responsiveness to IL-15 signaling
Given the robust inflammation observed in both the lamina propria and epithelial compartments of lnc13−/−-DQ8 mice after gluten challenge, we hypothesized that lnc13 normally dampens immune cell responsiveness to inflammatory cytokines. Notably, lnc13 expression is one of the highest in CD8+ T cells (Fig. 5a and Extended Data Fig. 7a,b), prompting us to focus our mechanistic studies on this population.

a, Relative expression of lnc13 in murine CD4+ T cells, CD8+ T cells and B cells, measured by RT–qPCR. lnc13 is most highly expressed in CD8+ T cells (n = 4; P = 0.00700709 comparing CD4+ versus CD8+ T cells, P = 0.00795608 comparing CD8+ T cells and B cells). b, RT–qPCR quantification of lnc13 and Dcp2 expression in CD8α+ IELs cultured for 48 h with medium alone, soluble IL-15 (sIL-15) or IL-15 superagonist (IL-15SA; IL-15 pre-complexed with IL-15Rα). IL-15 stimulation leads to downregulation of lnc13 and upregulation of its degrading enzyme Dcp2. lnc13 and Dcp2 expression, by RT–qPCR, was first normalized to Rpl32. ΔΔCt was calculated relative to average of unstimulated samples (for lnc13) or IL-15SA stimulated samples (for Dcp2). n = 5 mice per experiment. For comparisons of lnc13 expression medium versus IL-15 and medium versus IL15ra, as well as Dcp2 medium versus IL-15 and medium versus Il15rap, P = 4.70678 × 10−6, 2.74899 × 10−6, 2.47 × 10−5 and 4.7847 × 10−7, respectively. c, Schematic model proposing that IL-15 signaling activates STAT5 to promote Dcp2 expression, leading to degradation of lnc13. d, Time-course of NKG2D expression in WT CD8a+ IELs cultured with or without IL-15SA. NKG2D induction becomes robust by day 5, marking NK-like differentiation. e, Purified CD8a+ T IELs were stimulated with IL-15SA for 5 days. CD8+ IELs from lnc13−/− mice exhibit enhanced cytotoxicity and NK marker expression. Representative flow plots (left) and quantification (right) showing: NKG2D+GZMB+ double-positive live cells. n = 4 mice across three independent experiments. P = 0.00028843. Samples from lnc13−/− mice are represented by red dots and those from lnc13+/+ mice are represented by blue dots. f, After 5 days of IL-15SA stimulation, representative histogram of GZMB and NKG2D of WT-DQ8 (blue) and lnc13−/−-DQ8 (red) is shown. Dashed line represents isotype control. Error bars represent mean ± s.e.m. Unpaired, two-tailed Student’s t-test was used for comparisons. **P < 0.01, ***P < 0.001, ****P < 0.0001.
To identify relevant cytokine cues, we stimulated murine immune cells with key CeD-associated cytokines: IL-15, TNF, IFN-γ and IL-6 (Extended Data Fig. 7c). While TNF had no effect on lnc13 expression, IFN-γ and IL-6 upregulated lnc13, consistent with STAT1 and STAT3 binding at a potential lnc13 upstream regulatory element in available ChIP-seq data (Extended Data Fig. 7c,d). By contrast, IL-15 stimulation led to a downregulation of lnc13 (Extended Data Fig. 7c).
IL-15 plays a central role in CeD, particularly in driving IEL cytotoxicity and tissue destruction33; however, the molecular intermediates linking IL-15 stimulation to pathogenic T cell differentiation remain incompletely defined. We confirmed that lnc13 does not directly regulate IL-15 expression (Extended Data Fig. 8a–c), suggesting that its function lies downstream of IL-15 signaling.
To model IL-15 signaling more accurately in vitro, we isolated small intestine CD8+ IELs from WT mice and stimulated them with either soluble IL-15 (sIL-15) or an IL-15 superagonist (IL-15SA), which mimics physiological presentation via complexing with IL-15Rα37 (Fig. 5c). Both forms of IL-15 stimulation resulted in downregulation of lnc13 and upregulation of Dcp2, its RNA-degrading nuclease (Fig. 5b). These changes occurred within 4 h (Extended Data Fig. 8d). We next noticed that, at the Dcp2 promoter, there exist substantial binding of STAT5, the canonical activated TF of IL-15 signaling38,39,40,41 (Extended Data Fig. 8e,f) and Dcp2 expression is dependent on STAT5 activity (Extended Data Fig. 8g). These results provide an explanation for the upregulated Dcp2 expression by IL-15 (Fig. 5c) and a mechanism for the decreased lnc13 levels upon IL-15 stimulation in CD8+ T cells.
To further investigate the expression dynamics of lnc13, we next stimulated naive CD4+ and naive CD8+ T cells with IL-15, TCR, IL-15 + TCR or IL-12. In contrast to IL-15, which significantly reduced lnc13 levels in CD8+ T cells, IL-12 stimulation had no detectable effect on lnc13 expression in either naive CD8+ or CD4+ T cells (Extended Data Fig. 8h,i). Moreover, lnc13 expression in CD4+ T cells was not responsive to IL-15 stimulation (Extended Data Fig. 8h). In both CD4+ T cells and CD8+ T cells, while TCR signaling increased lnc13 expression, lnc13 levels were induced much higher in CD4+ T cells than CD8+ T cells (Extended Data Fig. 8h,i). In CD4+ T cells, TCR + IL-15 also increased lnc13 expression to a similar level as TCR signaling alone. In CD8+ T cells, however, IL-15 + TCR resulted no change in lnc13 levels, indicating the suppression of lnc13 by IL-15 signaling and induction of lnc13 by TCR signaling canceled each other (Extended Data Fig. 8h,i). To understand this divergence, we examined Dcp2, the lnc13-degrading enzyme and found that while IL-15 induces Dcp2 expression in peripheral CD8+ T cells, IL-15 does not induce Dcp2 in CD4+ T cells (Extended Data Fig. 8j). This cell-type-specific Dcp2 regulation may reflect differences in chromatin accessibility and enhancer usage between CD4+ and CD8+ lineages. As controls, we observed that for canonical T cell activation cytokines Ifng and Il2, TCR and TCR + IL-15 strongly induced their expression (Extended Data Fig. 8k–n).
We next confirmed that human lnc13 is similarly degraded by IL-15 in CD8+ T cells (Extended Data Fig. 9a). We purified human CD8+ T cell subsets and found that effector memory and central memory CD8+ T cells expressed much higher levels of lnc13 than naive CD8+ T cells (Extended Data Fig. 9b). We next examined the expression of lnc13 upon stimulation in human naive, effector memory and central memory CD8+ T cells (Extended Data Fig. 9c–e). lnc13 in naive human CD8+ T cells responded similar as in mice, whereas IL-15 reduced lnc13 levels, TCR stimulation increased lnc13 levels and TCR + IL-15 had no effect on lnc13 levels (Extended Data Fig. 9c). In effector memory CD8+ T cells, whereas TCR stimulation had no effect, IL-15 stimulation and IL-15 + TCR stimulation reduced lnc13 levels (Extended Data Fig. 9d). In central memory CD8+ T cells, lnc13 levels remained largely unchanged across all conditions, with only a minimal increase under combined IL-15 + TCR (Extended Data Fig. 9e).
To assess functional consequences, we cultured WT CD8+ IELs with IL-15SA and monitored NKG2D expression over time. NKG2D, a cytotoxic NK receptor, was strongly induced by day 5 (Fig. 5d). When we repeated the experiment using lnc13−/− IELs, we observed a significantly higher proportion of cells expressing IFN-γ, NKG2D and GZMB, hallmarks of NK-like, IL-15-driven LAK cells and resembling phenotypes seen in vivo post-gluten challenge (Fig. 5e,f and Extended Data Fig. 9f). Notably, knockout of lnc13 did not alter the expression of Il18rap in CD8+ T cell at resting or upon stimulation validating that these effects are not artifacts due to disruption of the adjacent Il18rap gene (Extended Data Fig. 9g).
These results establish that lnc13 constrains the differentiation of CD8+ IELs into cytotoxic LAK cells by limiting responsiveness to IL-15. Loss of lnc13 lowers the threshold for IL-15-induced activation, potentially contributing to the inflammatory and cytotoxic phenotype observed in vivo.
Lnc13 associates with DNA regulatory elements in CD8+ T cells
To understand how lnc13 regulates IL-15 responsiveness, we performed transcriptomic analysis of CD8α+ IELs stimulated in vitro with IL-15SA for 48 h (Fig. 6a and Supplementary Table 4). Top upregulated genes in KO (for example, Tiam1, Maf, Cxxc5, Brca2, Bcl9, Bcl2l2, Fcho1, Bmf and Junb) are consistent with enhanced T cell activation, proliferation and epigenetic remodeling, in line with the heightened responsiveness observed to in vivo gluten challenge CD8+ T IELs and in CD8+ T IELs from patients with CeD (Figs. 3k and 6a). Pathway enrichment analysis revealed that, with IL-15SA, KO vs WT DEGs are enriched for response to stimulus, signal transduction, leukocyte activation, IL-2/IL-15 signaling, Notch signaling, NK activation and lymphoma/CeD signatures (Fig. 6b). These are consistent with the broad transcriptomic signatures from post-gluten challenge in vivo CD8+ IEL KO versus WT DEGs and celiac disease CD8+ T IEL DEGs (Fig. 3k). In contrast, the no-stimulation KO–WT comparison shows these terms as largely attenuated in significance (TCR, IFN-γ, response to stimulus, signal transduction and cell cycle) or nonsignificant (such as leukocyte activation, IL-2–IL-15 signaling, inflammatory response, CeD and lymphoma), indicating that the in vivo CD8+ T IEL transcriptomic changes are partly recapitulated by IL-15 stimulation ex vivo (Fig. 6b–d).

a, Volcano plot of DEGs from CD8α+ IELs isolated from WT or lnc13−/− mice and cultured with IL-15SA for 48 h. n = 3 biological replicates. Key upregulated genes in lnc13−/− cells include T cell activation (Tiam1 and Junb), immune activation (Maf, Cxxc5 and Ifngr2) and proliferation (Brca2, Mapk7, Bcl9, Bcl2l2 and Bmf). Differential expression was computed using DESeq2 to derive P values. b, Pathway enrichment analysis of DEGs from IL-15SA-stimulated CD8α+ IELs (‘+IL-15’) and control unstimulated CD8a+ IELs (unstim). n = 3 biological replicates. Enriched terms in ‘+IL-15’ but not in ‘unstim’ include signal transduction, leukocyte activation, T cell response in CeD, NK cell activation and IL-2/IL-15 signaling pathway. c, Heatmap showing expression of selected IL-15-responsive genes (for example, Junb, Pak1, Bcl2l1 and Cbl) and IL-15 signaling pathway components (for example, Il2rb, Jack3, Gab2 and Rela), which are also marked with yellow circles in RNA-seq of WT and lnc13−/− activated with IL-15. d, Pathway enrichment analysis of lnc13 KO-DQ8 vs WT-DQ8 DEGs from CD8ab+ T IELs isolated after gluten challenge (‘in vivo gluten’) and lnc13 KO versusWT DEGs from CD8a+ T cell stimulated for 48 h with IL-15SA (‘ex vivo + IL-15’) and control unstimulated CD8a+ T cells (‘ex vivo unstim’). ‘In vivo gluten’ DEGs show significant enrichment of IL-15–IL-2 signaling pathway along with those associated with RANKL, IL-12, IL-6, TNF, IL-4/IL-5 and IFN-γ, whereas those from ‘ex vivo + IL-15’ show only significant enrichment for IL-15–IL-2, Notch and MAPK. DEGs from ‘ex vivo unstim’ do not show significant enrichment of any of these pathways. Scale bar shows −log10(FDR) of the mentioned pathways. e, Heatmap of selected shared DEGs between CD8α+ IELs stimulated with IL-15SA in vitro (left) and CD8αβ+ IELs isolated in vivo after gluten challenge (right). Many IL-15-responsive genes upregulated in vitro are also elevated in vivo upon lnc13 loss. On the far right, genes are functionally classified into proliferation (green), immune activation (purple), metabolism (yellow) or circadian rhythm (blue). For c and e, the numbers after ‘rep:’ indicate paired littermate mice from the same cage. f, Enrichment of TF motifs associated with DEGs in lnc13-deficient IELs (IL-15SA condition). Multiple enriched TFs (outer circle) are known to interact with the NuRD chromatin remodeling complex (highlighted by connecting lines). g, Schematic of RAP–DNA sequencing approach to map endogenous lnc13 chromatin association in primary CD8+ T cells. h, RAP-DNA-seq track (n = 2, overlaid), WT RAP versus KO RAP (as negative control), showing strong enrichment of lnc13 at its own locus, confirming pulldown specificity. i–k, RAP-DNA-seq coverage tracks showing association of lnc13 with Maf (i) Cxxc5 (j) and Gab2 (k). Each track is normalized by both bin size and total read counts, akin to the RPKM normalization analysis in RNA-seq (h–k). Tracks from both replicates were overlaid together. l, Bar graph showing the percent overlap of lnc13 KO versus WT ‘ex vivo + IL-15’ DEGs with RAP peaks separated based on how close these peaks are to the gene body of the DEGs: ±5 kb, ±50 kb or ±500 kb. m, Model. In resting CD8+ T cells, lnc13 associates with NuRD to repress IL-15 pathway genes. Upon IL-15 stimulation, lnc13 is degraded, lifting repression and permitting NK-like LAK differentiation.
We also observed upregulation of signaling components associated with IL-15 signaling, such as Il2rb, Jak3, Gab2, Rela, Stat5a, Mapk8 and Pik3cg, as well as IL-15 response genes, such as Junb, Pak1, Arhgdia, Bcl2l1 and Cbl (Fig. 6c). Moreover, we observed that 32.6% of the in vivo lnc13−/−-DQ8 versus WT-DQ8 CD8+ T IELs after gluten challenge DEGs (Fig. 3j) correspond to IL-15-responsive genes in WT cells, and this proportion increases to 42.0% when using the KO IL-15 response set (Extended Data Fig. 10a and Supplementary Tables 5 and 6). Several of the DEGs in both the in vivo and ex vivo +IL-15SA datasets overlapped and interestingly, many of the upregulated genes are associated with proliferation and immune activation while downregulated genes are associated with metabolism and circadian rhythm (Fig. 6e). These data suggest that a significant proportion of the in vivo transcriptomic changes (KO-DQ8 versus WT-DQ8, post-gluten) are IL-15-responsive genes.
Notably, the in vivo DEGs also show enrichment for signaling pathways driven by multiple cytokines beyond IL-2/IL-15, including TNF, IL-12, IFN-γ, RANKL, IL-6 and IL-4/IL-5, whereas the ex vivo IL-15-stimulated DEGs are enriched primarily for IL-15/IL-2-associated pathways (along with Notch and MAPK, which act downstream of IL-15 signaling) (Fig. 6d). None of these signatures was detected in the unstimulated condition, indicating that the enhanced cytokine responses in lnc13-deficient cells remain specific to the activating signal (Fig. 6d). Together, these results suggest that ex vivo IL-15 stimulation captures the core IL-15-responsive program regulated by lnc13, whereas the full in vivo transcriptional phenotype reflects the systems-level integration of additional cytokine inputs within the intestinal microenvironment. These results suggest that lnc13 deficiency lowers the activation threshold of CD8+ T IELs, enabling synergistic responses to IL-15 and other inflammatory cytokines, which together drive the amplified effector and inflammatory signatures observed in vivo. Indeed, many of these cytokine pathways (for example, TNF, IL-12 and IFN-γ) are known to amplify or modulate specific aspects of IL-15 signaling, further supporting this integrated model42,43,44.
We next independently validated a selection of key lnc13 DEGs presented from both in vivo gluten and ex vivo IL-15-stimulated RNA-seq datasets (Fig. 3j and Fig. 6a) by RT–qPCR in human CD8+ T cells with lnc13 knockdown versus control siRNA knockdown that were unstimulated or stimulated with human IL-15 (Extended Data Fig. 10b,c). These results show consistent upregulation of these genes upon lnc13 knockdown when stimulated with IL-15, but the differences were largely abolished without IL-15 stimulation.
To determine how lnc13 controls expression of these targets, we performed TF motif analysis of lnc13-regulated genes. This revealed significant enrichment for TFs associated with the NuRD (nucleosome remodeling and deacetylase) complex, consistent with our previous work showing that lnc13 recruits NuRD to repress gene expression16 (Fig. 6f).
We next used RNA antisense purification (RAP) followed by DNA sequencing (RAP–DNA-seq) to map lnc13 chromatin binding sites in primary CD8+ T cells, where RAP in lnc13 KO CD8+ T cells served as negative controls for background sequencing noise (Fig. 6g). Lnc13 enrichment at its own locus confirmed the specificity of our pulldown (Fig. 6h). We then performed differential enrichment analysis, and significant peaks are shown in Integrated Genome Viewer (IGV) (Fig. 6i–k) for representative genes (Maf, Cxxc5 and Gab2). These genes are upregulated in lnc13 KO CD8+ IELs after IL-15 stimulation, illustrating a direct correspondence between binding and expression effects. Notably, binding patterns differed among targets: Maf and Gab2 exhibited sharp enrichment at promoter regions, whereas Cxxc5 displayed more diffuse binding across the gene body. Such broad binding patterns are consistent with previous reports demonstrating that, in certain contexts, lncRNAs can exert regional regulatory influence rather than binding at discrete, sharply defined sites45,46.
We next compared the genomic coordinates of RAP peaks with DEGs from the CD8a+ IELs stimulated with IL-15SA RNA-seq dataset. Considering the possibility of lnc13 to act via both promoter-proximal and distal enhancer interactions, we evaluated overlap using three proximity windows around each gene body: ±5 kb, ±50 kb and ±500 kb. We found that 14.1% of DEGs overlapped nearby (±5 kb) RAP peaks, 32.1% overlapped within ±50 kb and 83.6% overlapped within ±500 kb (Fig. 6l), representing 33% of total RAP peaks (Extended Data Fig. 10d). Thus, a substantial proportion of lnc13-regulated genes are physically associated with lnc13-bound loci, consistent with cis-regulatory targeting.
These data demonstrate that lnc13 binds specific DNA regulatory elements in CD8+ T cells and represses expression of IL-15-responsive genes, a function likely mediated via NuRD recruitment. Together, this provides a molecular mechanism for how lnc13 restrains cytotoxic effector differentiation and preserves oral tolerance to dietary antigens (Fig. 6m and Supplementary Fig. 6).
Discussion
We demonstrate here that the long-noncoding RNA lnc13 is a key regulator of oral tolerance to gluten. In mice humanized with HLA-DQ8, genetic ablation of lnc13 resulted in enhanced proinflammatory and cytotoxic immune responses, mucosal pathology and molecular signatures reminiscent of human CeD. These findings provide causal evidence that a noncoding RNA, previously associated with CeD risk by GWAS, acts as a modulator on intestinal immune activation.
Our results help address a long-standing paradox in CeD: while a significant portion of the population carry HLA-DQ2 or HLA-DQ8, only a subset develops CeD7. Rather than acting as a deterministic trigger, our data suggest that lnc13 functions to raise the threshold for gluten-driven immune activation. Loss of lnc13 lowers this threshold, rendering the immune system more susceptible to inflammation in response to dietary gluten. Consistent with this model, the immune and histological alterations observed (manifested primarily as crypt hyperplasia, a well-recognized early stage of human CeD before villous atrophy) align with the polygenic nature of the disease, in which no single GWAS variant is expected to generate severe pathology in isolation. Indeed, it remains unclear whether any single genetic perturbation can fully recapitulate the full spectrum of human CeD pathology in mice.
Notably, lnc13−/−-DQ8 mice develop gluten-dependent and reversible intestinal pathology in the absence of adjuvants or cytokine overexpression, a level of physiological fidelity that is uncommon among existing CeD models. That deletion of a single noncoding RNA, representing one CeD-associated locus, elicits molecular, immunologic and histopathologic hallmarks of the human disease underscores both the specificity and biological significance of this regulatory pathway. The lnc13−/−-DQ8 mice constitute a genetic model in which loss of a noncoding, GWAS-linked regulator, together with HLA-DQ8 and dietary gluten, lowers the tolerance threshold and enable gluten-dependent CeD-like immune activation and early pathology.
A complex milieu of inflammatory signals, especially cytokines, drive CeD pathology4. In CD8+ T cells, which are critical in CeD pathogenesis, we show that lnc13 desensitizes these cells to inflammatory signals, in particular IL-15, a cytokine central to CeD pathogenesis. Mechanistically, lnc13 associates with DNA regulatory elements of effector genes, including those in the IL-15 pathway, and represses their expression, likely via NuRD complex recruitment. In the absence of lnc13, CD8αβ+ IELs become hyperresponsive to inflammatory signals, including IL-15, adopt an NK-like cytotoxic phenotype and recapitulate effector states observed in active CeD. Notably, IL-15 itself is not a GWAS hit and is not uniformly elevated in all patients24, suggesting that altered sensitivity to inflammatory signals, rather than cytokine abundance alone, may represent a critical determinant of pathology.
Beyond CD8+ T cells, lnc13 deficiency also affects broader immune homeostasis, including effects in Treg cells, B cells and myeloid cells. These alterations likely cooperate to shape intestinal pathology, particularly during early disease stages characterized by crypt hyperplasia and epithelial remodeling before overt villous atrophy. Because lnc13 is expressed across multiple immune compartments that collectively maintain mucosal tolerance, the use of a global knockout allowed us to capture the integrated consequences of its loss, analogous to the systemic loss-of-function associated with human risk alleles; however, future studies using cell type-specific deletions will be important to determine whether lnc13 functions independently or synergistically across lymphocytes, myeloid cells and epithelial compartments to maintain tolerance.
The reversibility of intestinal pathology in lnc13−/−-DQ8 mice upon gluten withdrawal further underscores the physiological relevance of this model. Although hundreds of non-HLA CeD GWAS loci have been identified, few have been functionally interrogated in vivo under disease-relevant conditions. In this context, lnc13−/−-DQ8 mice provide a tractable genetic platform to examine how individual noncoding risk loci modulate immune thresholds rather than dictate disease outcomes outright, and to explore therapeutic strategies aimed at restoring immune homeostasis.
Finally, lnc13 is also implicated in inflammatory bowel disease GWAS loci, suggesting that its function may extend beyond gluten-specific responses to broader regulation of mucosal immunity47. More generally, the experimental framework used here (integrating noncoding genetics, in vivo modeling and mechanistic dissection) offers a template for decoding how modest-effect noncoding variants contribute to complex inflammatory diseases.
Methods
Ethics statement
All animal experiments were approved by the Institutional Animal Care and Use Committee (protocol number AABH6558) of Columbia University and were performed in accordance with the Guide for the Care and Use of Laboratory Animals in an AAALC-accredited facility. All procedures were conducted under protocols approved by Columbia University.
De-identified human peripheral CD8+ T cells were obtained commercially (STEMCELL Technologies). These samples were provided de-identified by the vendor and therefore did not constitute human participant research requiring institutional review board approval. Donor consent was obtained by the vendor according to their institutional policies.
Mouse
All mice used in this study are on C57BL/6J background. Mouse colonies were maintained under specific pathogen-free conditions at Columbia University. Mice were housed at 68–79 °F and 30–70% humidity in a 12-h light–dark cycle with ad libitum access to food and water. Transgenic HLA-DQ8 mice lacking mouse MHC-II (DQ8) was a gift from B. Jabri, at the time in University of Chicago. Lnc13−/− mice were generated as follows. Three pairs of CRISPR sgRNAs were complexed with CRISPR Cas9 and injected into fertilized mouse eggs in the B6C BAF1 background (Supplementary Table 2 lists gRNA sequences and lnc13 genotyping primers). For each pair, one sgRNA targets near the beginning of the deletion and the other at the end. The entire length of lnc13, excluding regions of Il18rap, were deleted. Eggs were implanted into pseudopregnant female mice. Pups with the correct deletion were confirmed by PCR and Sanger sequencing. Mice were backcrossed onto C57BL/6 background for at least nine generations. The lnc13 KO mice were then crossed onto mice having the HLA-DQ8 transgene, while lacking endogenous MHC-II. These mice are maintained by breeders with genotype lnc13+/− HLA-DQ8tg/tg MHC-II−/− to allow littermate controls in our experiments to account for variations in microbiome and development. For most experiments, littermate (age-matched) and co-housed (sex-matched) mice are used to account for variations in microbiome.
Gluten challenge
Gluten challenge was followed essentially as previously published with a few exceptions24. Mice were bred on breeder chow and pups were weaned into gluten-free diet (AIN-76A, Envigo) until they were 10 weeks old. Subsequently, these mice are switched back onto normal chow containing gluten. Mice were gavaged every other day for 45 days with 20 mg of crude gliadin (Sigma G3375). For sham gluten challenge, mice were gavaged with PBS every other day. For gluten withdrawal, gluten-challenged mice were switched back to gluten-free diet and sham challenged every other day with PBS.
Histology analysis
Terminal ileum gut bundles were prepared as previously published24. Specifically, for whole intestine Swiss rolls, the entire small intestine (from just before sphincter to just within cecum) was extracted after killing the mice. Then, large chunks of feces were removed by gentle pushing with tweezers and finger. Then, the insides of small intestine were washed by injection of 1× PBS from the stomach end and again pushed all the way through with tweezers and finger. Then, small intestines were rolled with the proximal end in the middle. Swiss rolls were fixed in 10% neutral buffered formalin for 24–48 h before transferring to 70% ethanol. Processing, paraffin embedding, sectioning and hematoxylin and eosin (H&E) staining were carried out by Columbia University Molecular Pathology Shared Resource.
H&E slides were scanned using Leica Aperio AT2 slide scanner. Counting of IELs and measurements of villi and crypt lengths were carried out in a blinded fashion on Aperio ImageScope software. Pathological scoring of villous atrophy was carried out in a blinded fashion by a trained, experienced pathologist. A score of 2 denotes villous atrophy, a score of 1 means mild atrophy and a score of 0 means no noticeable signs of villous atrophy.
ELISA
Mice peripheral blood was obtained by tail bleeding under heat lamp. Then ~50–100 ml of blood was allowed to coagulate at 20 °C for 30 min and spun down for 5 min at 1,800g at 4 °C. Sera were collected and stored at −80 °C. ELISA for peripheral blood antibodies were carried out as previously described24. In brief, purified gliadin (Sigma G3375) was digested by pepsin (Sigma P7000) in 0.2 N HCl. Then, the pH was adjusted to 8.0 with NaOH to further digest by trypsin (Sigma T4799) and then by pancreatin (Sigma P1750). Undigested fragments were fractioned and the digested fragments were isolated to be loaded into ELISA plates. Deamidated gliadin (SurModics A19500) and tTG (Sigma T5398) were purchased and loaded into ELISA plates. Donkey anti-mouse IgG antibody with peroxidase (Jackson ImmunoResearch 715-035-150) were used for detection of anti-gliadin, anti-deamidated gliadin and anti-tTG ELISA. Development of signal was carried out using OptEIA Regent Set A (BD Biosciences 550536) and signal was read as optical density at 450 nm.
For cytokine levels, sera were examined by ELISA kits for TNF (BD Biosciences 560478), IFN-γ (BD Biosciences 555138), IL-6 (BD Biosciences 555240) and CRP (Proteintech KE10085) as directed by the manufacturer.
IEL and LPL preparation
IEL and LPLs were obtained as previously described48. In brief, small intestines were collected. Then, the feces were cleaned and epithelial cells were dissociated with several washes of PBS containing FBS, EDTA and dithiothreitol (DTT). These epithelial fractions were collected and immune cells were further enriched with 40% Percoll in 1× PBS and spun for 20 min at 2,000 rpm at 20 °C with acceleration at 2 and no brake. For isolation of small intestine epithelial cells, the upper fraction of Percoll separation was taken. The lower fraction/pellet contains the IEL fraction. Then, the rest of the small intestine was digested with a combination of Liberase (Sigma 5401119001) and DNase I (Sigma 11284932001) in HBSS (with magnesium and calcium). The digested intestines were then mashed through a cell filter and the final lamina propria cells were enriched for immune cells with Percoll to obtain the LPL fraction. IELs and LPLs were first stimulated with PMA/ionomycin/BFA (anti-CD107a and anti-CD107b flow cytometry antibodies are also added at this step) for 4 h at 37 °C in complete RPMI to stain for intracellular cytokines.
Peripheral immune cell isolation
Spleens were collected and mashed through a 70-μm cell strainer, and the splenocytes were incubated with RBC lysis buffer at 20 °C for 3 min. RBC lysed splenocytes were then processed for flow cytometry or first stimulated with PMA/Ionomycin/BFA for 4 h at 37 °C in complete RPMI. Inguinal lymph nodes were also mashed through a 70-μm cell strainer. Lymph node cells were then processed for flow cytometry or first stimulated with PMA/ionomycin/BFA for 4 h at 37 °C in complete RPMI. Peripheral blood was obtained by tail vein bleeding or cardiac puncture. Around 200 μl of blood was immediately transferred to a FACS tube with 5 ml 0.5 M EDTA on ice. Red blood cells were lysed with RBC lysis buffer. Blood cells were then stained for flow cytometry or first stimulated with PMA/ionomycin/BFA for 4 h at 37 °C in complete RPMI. Thymuses were directly smashed through a 70-μm cell strainer.
Cell culture
Primary murine BMDMs were cultured as previously described49. Specifically, BMDMs were cultured in DMEM containing 10% heat-inactivated FBS and 10% L929 conditioned medium. Cells were treated with or without 10 ng ml−1 LPS (Sigma L8274).
Peripheral CD8+ T cells were isolated by negative selection using the MojoSort Mouse CD8+ T cell Isolation kit (BioLegend 480007) from spleen and peripheral lymph nodes. Peripheral CD4+ T cells were isolated with MojoSort Mouse CD4+ T cell Isolation kit (BioLegend 480005). CD8+ T cells were cultured in RPMI-1640 containing 10% heat-inactivated FBS (Peak Serum PS-SB4), 1× non-essential amino acids (Thermo 11140050), 1× β-mercaptoethanol (Thermo 21985023), 1× HEPES (Thermo 15630080), 1× GlutaMAX (Thermo 35050061), 1× Pen–Strep (Thermo 15140122) and 1× sodium pyruvate (Thermo 11360070). CD8+ T cells were cultured with or without 100 ng ml−1 soluble IL-15 (PeproTech 210-15). CD8+ T cells were also activated by plate-bound aCD3 (Tonbo 40-0031) and soluble aCD28 (Tonbo 70-0281). De-identified human peripheral CD8+ T cells were obtained from STEMCELL Technologies and cultured as above, but stimulated with human IL-15 (Thermo 200-15). Sorted CD8a+ T IELs were cultured as with peripheral CD8+ T cells with or without 100 ng ml−1 soluble IL-15 or 100 ng ml−1 IL-15SA (Sino Biological CT113-M08H).
RAW 264.7 cells (ATCC) were cultured in DMEM containing 10% heat-inactivated FBS and treated with or without 100 ng ml−1 of TNF (PeproTech 315-01 A), IFN-γ (BioLegend 575302), IL-6 (PeproTech 216-16) or soluble IL-15.
Flow cytometry
The following conjugated antibodies were purchased: CD45.2 BV786 (BD 563686), CD45 redFluor710 (Tonbo Biosciences 80-0451), TCRβ BV711 (BD Biosciences 563135), CD8α vFluor450 (Tonbo 75-1886), CD8β PE-TexasRed (Biotium BNC942003), CD8α APC (Tonbo 20-0081), TCRγδ PE (eBiosciences 12-5711-81), NK1.1 FITC (eBioscience 11-5941-82), streptavidin BUV395 (BD 564176), NKG2D biotin (BioLegend 115703), Eomes Alexa647 (eBiosciences 51-4875-80), T-bet PE-Cy7 (eBioscience 25-5825-80), CD107a PE (BioLegend 121611), CD107b APC (Thermo A15449), IFN-γ PE-Cy7 (Tonbo 60-7311), GZMB FITC (BioLegend 515403), IL-12 p40/p70 APC/Fire 750 (BioLegend 505217), CD4 APC-eFluor780 (eBioscience 47-0042-82), FOXP3 eFluor450 (eBioscience 48-5773-82), B220 PE-Cy5 (eBiosciences 15-0452-81), CD11b violetFluor 500 (Tonbo 85-0112), CD103 PE-Cy7 (BioLegend 121425), FITC isotype control (BioLegend 400107), PE-Cy7 isotype control (Tonbo 60-4301), biotin isotype control (BioLegend 400903), human CD8+ BUV737 (BD OptiBuild 749367), human CD45RA APC-Cy7 (BioLegend 304127) and human CCR7 BV605 (BD Horizon 563711). Representative gating strategies for LPLs and IELs are shown in supplementary materials. Dead cells were stained with Live-or-Dye 350/488 fixable staining kit (Biotium). Flow cytometry staining and data collection were conducted as previously described50. For CD107a/b detection, the cells were stained as previously described24. In brief, CD107a and CD107b antibodies were added to the BMA/iono/BFA stimulation, as described below, before staining the rest of the markers. Intracellular cytokines were fixed with eBioscience IC Fixation Buffer (Thermo 00-8222-49) and washed with 1× eBioscience Permeabilization Buffer (Thermo 00-8333-56). TFs were stained with eBioscience Foxp3/Transcription Factor Staining Buffer set (Thermo 00-5523-00).
Fluorescent-activated cell sorting
FACS was used to sort various cell populations. For CD8a T IELs, we first utilized anti-CD8a-biotin (Tonbo 30-0081) and streptavidin nanobeads (BioLegend) to enrich for CD8a+ cells from the IEL fraction. Then, cells were sorted for CD8a violetFluor 450 (Tonbo 75-1186). For CD8αβ T cells, the whole IEL fraction was sorted for CD8α APCs and CD8β PE-TexasRed. For peripheral CD4+ and CD8+ T cells, spleen and peripheral lymph nodes were first collected, followed by RBC lysis buffer (Sigma R7757). Mouse naive CD4+ cells were sorted on CD4+CD8−CD44loCD62Lhi, naive CD8+ T cells were sorted on CD4−CD8+CD44loCD62Lhi. Mouse B cells were sorted from whole splenocytes on CD45+B220+ cells. For lamina propria myeloid cells, the LPL fraction was sorted for CD45+TCRb−B220−CD11b+ cells.
RNA antisense purification
RAP was performed as previously described16 using 25 million primary CD8+ T cells, followed by 2 mM DSG and 3% formaldehyde crosslinking. In brief, lysed whole cells were fractionated to obtain intact nuclei. Nuclei were lysed and incubated with antisense single-stranded 5’ biotinylated DNA probes tiling the entire lnc13 transcript but not binding to Il18rap (Supplementary Table 2 lists probe sequences). RAP probes are ordered as 90–120-nt oligonucleotides from IDT and designed as previously described16 such that sequences that significantly match to other regions of the genome or significantly cover known repetitive sequences are not used. Streptavidin-magnetic beads were used to enrich for lnc13 and bound chromatin was eluted by RNaseH digestion. Lnc13-bound chromatin were examined by sequencing. DNA-seq library was generated using NEBNext Ultra II DNA Library Prep kit from Illumina. Around 40 million 150-bp paired-end reads were generated by next-generation sequencing (Azenta). Raw FASTQ reads were initially removed of adaptor by CutAdapt and aligned to the mm39 genome by Burrows–Wheeler Aligner (BWA). PCR duplicates, low-quality mapped reads and reads mapping to the probe sequences were removed. Then, the relative signal was calculated, in 150-bp windows with 75-bp step size, using DeepTools bamCoverage and normalized to both bin sizes and number of reads, akin to RPKM method in RNA-seq51. These generated tracks are viewed in IGV to obtain track views shown in Fig. 6. To obtain significantly bound regions, we next calculated the average signal in 2.5-kb bin sizes across the entire genome using DeepTools multibigwigsummary to call significantly bound regions comparing the signal from WT lnc13 RAP to KO lnc13 RAP, which served as a negative control for nonspecific binding of probes or background sequencing noise (|log2FC| > 1, P < 0.005). The original RAP-seq paper utilized bin sizes of 100 kb for Xist as Xist is expected to exhibit long distance coverage across the entire chromosome45. Here, we sought to examine lnc13 binding at a higher resolution to examine binding to specific loci in the genome. Significant peaks were shown in a bed format in IGV, shown in Fig. 6.
To find peaks that overlapped with lnc13 DEGs, we first generated a .bed file of all ex vivo IL-15SA stimulated lnc13 KO versus WT IEL RNA-seq DEGs. We then generated three separate.bed files each expanding the original gene body coordinates by ±5 kb, ±50 kb or ±500 kb. Then, BEDtools was used to find significant RAP peaks that completely overlapped lnc13 DEGs depending on these genomic distances52. Finally, Venny v.2.1.0 was used to generate Venn diagrams.
Human lnc13 knockdown
To generate siRNAs against human lnc13, we first cloned the entire human lnc13 region (primers are shown in Supplementary Table 2). Each primer also contained T7 polymerase sites (NEB M0251S) to allow in vitro transcription to make double-stranded RNA of the entire human lnc13 region. Subsequently, purified double-stranded RNAs were digested into 17-21nt siRNAs by incubation with RNase III (NEB M0245). Around 1 μg of pooled siRNA were nucleofected, with a home-made nucleofection buffer, into de-identified human CD8+ T cells (STEMCELL Technologies) for lnc13 knockdown22. Control knockdown siRNA (siNeg) was ordered from IDT (51-01-14-04)
To prepare human CD8+ T cells for knockdown, they were first activated by plate-bound aCD3 (Tonbo 40-0037), aCD28 (Tonbo 40-0289) and human IL-15 (Peprotech) for 48 h to expand. Then, these cells were nucleofected with human siRNA. Then, cells are transferred to medium alone (unstimulated) or medium containing human IL-15 100 ng ml−1 for 48 h before collecting the cells for RNA.
RNA extraction and cDNA generation
For whole-tissue RNA, samples were extracted and flash-frozen in liquid nitrogen. Then, samples were homogenized with a mortar and pestle containing small amounts of liquid nitrogen to keep the samples frozen. Homogenized samples were then transferred to TRIzol. TRIzol RNA extraction was then performed as directed by the product protocol. For extraction of RNA from cells, we utilized Quick-RNA Miniprep kit (Zymo). In certain cases, cDNA was generated using oligonucleotide dT primers and a GoScript Reverse Transcriptase kit (Promega) as directed using equal amount of RNA between samples of comparison.
qPCR
qPCR was carried out with a NEB Luna qPCR master Mix (NEB M3003E). For analysis, Ct values from gene of interest were normalized to housekeeping gene Ct values in the same sample to obtain ΔCt values and account for minor differences in loading between samples. In most cases, to facilitate representation of fold-change differences between samples, samples were further normalized by the ΔΔCt method, whereby average of one sample was used to normalize the other samples. The exact method of normalization is detailed in the figure legends of every quantitative PCR with reverse transcription (RT–qPCR) experiment.
Oligonucleotide generation
All oligonucleotides (listed in Supplementary Table 2) were generated by oligonucleotide synthesis companies, such as Integrated DNA Technologies.
RNA-seq library preparation and bioinformatics
For library preparation of IELs, LPLs and whole tissue (small intestine ileum), a NEBNext UltraII RNA library Prep kit (Illumina) was used. For the library preparation of CD8a+ T IELs cultured in IL-15SA for 48 h, CD8a+ T IELs cultured in medium alone for 48 h or sorted CD8ab T IELs after gluten challenge, NEBNext Single Cell/Low Input RNA library Prep kit (Illumina) was used.
All samples were sequenced at around 20 million reads per sample by Azenta. Reads were aligned using RNA-STAR53. Gene counts were counted using FeatureCounts54 and differential expression was conducted using DESeq2 (ref. 55) accounting for variation between cages (analysis considered each WT–KO pair that were same sex, littermates and co-housed). G:profiler and Enrichr were used to find statistically significant pathways affected56,57. For Fig. 2h, whole tissue or LPL RNA-seq DEGs (P < 0.05, |log2FC| > 1) were used for pathway analysis. A list of human-mouse homologs were provided by Jax for comparison between mouse and human genes. To obtain the data presented in Extended Data Fig. 3a, the fold change (KO versus WT) of mouse lnc13 ileum DEGs (log2FC > 1 P < 0.05) was plotted against their human homolog’s fold change (celiac versus HC). Then, a Pearson’s correlation was applied. For Fig. 3a, IEL DEGs (P < 0.05, |log2FC| > 1) were used for pathway analysis. For Fig. 3j,k and Supplementary Table 3, small intestine CD8ab T IEL DEGs (P < 0.05 |log2FC| > 1) were used for volcano plot and pathway analysis. For Fig. 6a,b and Supplementary Table 4, DEGs for CD8a T IEL IL-15SA (P < 0.05, |log2FC| > 1) and a minimal read count >10 was used for volcano plot and pathway analysis. For Fig. 6b,d DEG for unstimulated CD8a+ T IEL (P < 0.05, |log2FC| > 1) was used for pathway analysis. For Extended Data Fig. 10a, IL-15 inducible DEGs for WT and KO (P < 0.05, |log2FC| > 1) was used with Venny v.2.1.0 to generate Venn diagram with DEGs of in vivo after gluten challenge CD8ab T cells. For Fig. 6c,e, subsets of statistically significant genes (P < 0.05) were shown. For Fig. 6f, predicted TF associations of transcriptomic analysis (P < 0.05) were determined using g:profiler. Association of these predicted TFs (top 21 most significant motifs) with NuRD complex was found by STRING analysis (physical association) of each TF with each component of NuRD complex58.
Publicly available RNA-seq analysis
Publicly available RNA-seq data were trimmed and aligned as above, except a custom GTF file was provided to make a custom RNA-STAR alignment16. In particular, the entirety of human (hg38 chr2:102452045-102454691) and mouse lnc13 (mm39 Chr1:40588014-40590867) region was provided as an addition to the GENCODE GTF file. DESeq2 was used to perform normalization and statistics for publicly available RNA-seq data. For Fig. 1a and Supplementary Table 1, differentially expressed lncRNAs were filtered based on P < 0.01 and |log2FC| > 2. For Fig. 2h, human RNA-seq (Padj < 0.05, |log2FC| > 1) were used for pathway analysis. For Fig. 3h, human CD8+ T cell and CD8+ NE pseudobulk RNA-seq DEGs (Padj < 0.05, |log2FC| > 1) were used for pathway analysis. For IL-12A and IL-12B expression in celiac samples, DESeq2 normalized IL-12A and IL-12B counts were further normalized against average of healthy control samples. For stranded RNA-seq analysis, RNA-seq was aligned by RNA-STAR and optioned to output strand-specific bam files, which were visualized in IGV. Gene Expression Omnibus accession numbers of data generated from publicly available sequencing data are noted in the figures or figure legends.
Biological replicates
All biological replicates were taken from distinct samples. Where appropriate, the number of independent experiments are noted.
Statistics and reproducibility
No statistical methods were used to predetermine sample size but sample sizes are similar to those reported in previous publications22,48. No data were excluded from the analyses. The experiments were not randomized. Data collection and analysis were not performed blind to the conditions of the experiments except where explicitly stated (histological scoring). Data distribution was assumed to be normal but this was not formally tested. Statistical analyses were performed using GraphPad Prism unless otherwise stated. Biological replicates represent independent samples derived from distinct animals or cell preparations.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All generated next-generation sequencing data are uploaded to the NCBI Gene Expression Omnibus under accession number GSE298129. Source data are provided wih this paper.
References
Green, P. H. R. & Cellier, C. Celiac disease. N. Engl. J. Med. 357, 1731–1743 (2007).
Tjon, J. M.-L., van Bergen, J. & Koning, F. Celiac disease: how complicated can it get?. Immunogenetics 62, 641–651 (2010).
Abadie, V., Sollid, L. M., Barreiro, L. B. & Jabri, B. Integration of genetic and immunological insights into a model of celiac disease pathogenesis. Annu. Rev. Immunol. 29, 493–525 (2011).
Iversen, R. & Sollid, L. M. The immunobiology and pathogenesis of celiac disease. Annu. Rev. Pathol. Mech. Dis. 18, 47–70 (2023).
Laurikka, P., Kivelä, L., Kurppa, K. & Kaukinen, K. Review article: systemic consequences of coeliac disease. Aliment Pharmacol. Ther. 56, S64–S72 (2022).
Kim, C.-Y., Quarsten, H., Bergseng, E., Khosla, C. & Sollid, L. M. Structural basis for HLA-DQ2-mediated presentation of gluten epitopes in celiac disease. Proc. Natl Acad. Sci. USA 101, 4175–4179 (2004).
Karell, K. et al. HLA types in celiac disease patients not carrying the DQA1*05-DQB1*02 (DQ2) heterodimer: results from the European Genetics Cluster on Celiac Disease. Hum. Immunol. 64, 469–477 (2003).
Buniello, A. et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 47, D1005–D1012 (2019).
Dubois, P. C. A. et al. Multiple common variants for celiac disease influencing immune gene expression. Nat. Genet. 42, 295–302 (2010).
Hangauer, M. J., Vaughn, I. W. & McManus, M. T. Pervasive transcription of the human genome produces thousands of previously unidentified long intergenic noncoding RNAs. PLoS Genet. 9, e1003569 (2013).
Mattick, J. S. et al. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 24, 430–447 (2023).
Zhao, L. et al. NONCODEV6: an updated database dedicated to long non-coding RNA annotation in both animals and plants. Nucleic Acids Res. 49, D165–D171 (2021).
Tian, R. & Ghosh, S. Mechanisms AND functions of lncRNAs linked to autoimmune disease risk alleles. Adv. Immunol. 161, 1–15 (2024).
Castellanos-Rubio, A. & Ghosh, S. Disease-associated SNPs in inflammation-related lncRNAs. Front. Immunol. 10, 420 (2019).
Castellanos-Rubio, A. & Ghosh, S. Functional implications of intergenic GWAS SNPs in immune-related lncRNAs. Adv. Exp. Med. Biol. 1363, 147–160 (2022).
Castellanos-Rubio, A. et al. A long noncoding RNA associated with susceptibility to celiac disease. Science 352, 91–95 (2016).
Gonzalez-Moro, I. et al. The T1D-associated lncRNA Lnc13 modulates human pancreatic β cell inflammation by allele-specific stabilization of STAT1 mRNA. Proc. Natl Acad. Sci. USA 117, 9022–9031 (2020).
Wapinski, O. & Chang, H. Y. Long noncoding RNAs and human disease. Trends Cell Biol. 21, 354–361 (2011).
Liu, L. et al. LncRNA IL21-AS1 interacts with hnRNPU protein to promote IL21 overexpression and aberrant differentiation of Tfh cells in systemic lupus erythematosus. Clin. Transl. Med. 12, e1117 (2022).
Fernandez-Jimenez, N. et al. Coregulation and modulation of NFκB-related genes in celiac disease: uncovered aspects of gut mucosal inflammation. Hum. Mol. Genet. 23, 1298–1310 (2014).
Guo, Q. et al. NF-κB in biology and targeted therapy: new insights and translational implications. Sig. Transduct. Target Ther. 9, 53 (2024).
Abadie, V. et al. IL-15, gluten and HLA-DQ8 drive tissue destruction in coeliac disease. Nature 578, 600–604 (2020).
Black, K. E., Murray, J. A. & David, C. S. HLA-DQ determines the response to exogenous wheat proteins: a model of gluten sensitivity in transgenic knockout mice1. J. Immunol. 169, 5595–5600 (2002).
Abadie, V., Khosla, C. & Jabri, B. A mouse model of celiac disease. Curr. Protoc. 2, e515 (2022).
Atlasy, N. et al. Single cell transcriptomic analysis of the immune cell compartment in the human small intestine and in celiac disease. Nat. Commun. 13, 4920 (2022).
Björck, S., Lindehammer, S. R., Fex, M. & Agardh, D. Serum cytokine pattern in young children with screening detected coeliac disease. Clin. Exp. Immunol. 179, 230–235 (2015).
DePaolo, R. W. et al. Co-adjuvant effects of retinoic acid and IL-15 induce inflammatory immunity to dietary antigens. Nature 471, 220–224 (2011).
Kornberg, A. et al. Gluten induces rapid reprogramming of natural memory αβ and γδ intraepithelial T cells to induce cytotoxicity in celiac disease. Sci. Immunol. 8, eadf4312 (2023).
Camarca, A., Rotondi Aufiero, V. & Mazzarella, G. Role of regulatory T cells and their potential therapeutic applications in celiac disease. Int. J. Mol. Sci. 24, 14434 (2023).
Bouziat, R. et al. Reovirus infection triggers inflammatory responses to dietary antigens and development of celiac disease. Science 356, 44–50 (2017).
Campos Canesso, M. C. et al. Identification of antigen-presenting cell-T cell interactions driving immune responses to food. Science 387, eado5088 (2025).
Du Pré, M. F. et al. Tolerance to ingested deamidated gliadin in mice is maintained by splenic, type 1 regulatory T cells. Gastroenterology 141, 610–620 (2011).
Meresse, B. et al. Coordinated induction by IL15 of a TCR-independent NKG2D signaling pathway converts CTL into lymphokine-activated killer cells in celiac disease. Immunity 21, 357–366 (2004).
Catassi, C. et al. Risk of non-Hodgkin lymphoma in celiac disease. JAMA 287, 1413–1419 (2002).
Gass, J., Vora, H., Bethune, M. T., Gray, G. M. & Khosla, C. Effect of barley endoprotease EP-B2 on gluten digestion in the intact rat. J. Pharmacol. Exp. Ther. 318, 1178–1186 (2006).
Dickson, B. C., Streutker, C. J. & Chetty, R. Coeliac disease: an update for pathologists. J. Clin. Pathol. 59, 1008–1016 (2006).
Bergamaschi, C. et al. Intracellular interaction of interleukin-15 with its receptor α during production leads to mutual stabilization and increased bioactivity. J. Biol. Chem. 283, 4189–4199 (2008).
Chenoweth, M. J. et al. IL-15 Can signal via IL-15Rα, JNK, and NF-κB to drive RANTES production by myeloid cells. J. Immunol. 188, 4149–4157 (2012).
Stone, K. P., Kastin, A. J. & Pan, W. NF-κB is an unexpected major mediator of interleukin-15 signaling in cerebral endothelia. Cell. Physiol. Biochem. 28, 115–124 (2011).
McDonald, P. P., Russo, M. P., Ferrini, S. & Cassatella, M. A. Interleukin-15 (IL-15) induces NF-kappaB activation and IL-8 production in human neutrophils. Blood 92, 4828–4835 (1998).
Mishra, A. et al. Aberrant overexpression of IL-15 initiates large granular lymphocyte leukemia through chromosomal instability and DNA hypermethylation. Cancer Cell 22, 645–655 (2012).
Becker-Hapak, M. K. et al. A fusion protein complex that combines IL-12, IL-15, and IL-18 signaling to induce memory-like NK cells for cancer immunotherapy. Cancer Immunol. Res. 9, 1071–1087 (2021).
Cimpean, M. et al. IL-15 priming alters IFN-γ regulation in murine NK cells. J. Immunol. 211, 1481–1493 (2023).
Fuchs, A. et al. Intraepithelial type 1 innate lymphoid cells are a unique subset of IL-12- and IL-15-responsive IFN-γ-producing cells. Immunity 38, 769–781 (2013).
Engreitz, J. M. et al. The Xist lncRNA exploits three-dimensional genome architecture to spread across the X-chromosome. Science 341, 1237973 (2013).
Quinodoz, S. A. et al. RNA promotes the formation of spatial compartments in the nucleus. Cell 184, 5775–5790.e30 (2021).
Jostins, L. et al. Host–microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 491, 119–124 (2012).
Huang, Y. et al. S1P-dependent interorgan trafficking of group 2 innate lymphoid cells supports host defense. Science. 359, 114–119 (2018).
Rao, P. et al. IκBβ acts to inhibit and activate gene expression during the inflammatory response. Nature 466, 1115–1119 (2010).
Grinberg-Bleyer, Y. et al. NF-κB c-Rel is crucial for the regulatory T cell immune checkpoint in cancer. Cell 170, 1096–1108.e13 (2017).
Ramírez, F., Dündar, F., Diehl, S., Grüning, B. A. & Manke, T. deepTools: a flexible platform for exploring deep-sequencing data. Nucleic Acids Res. 42, W187–W191 (2014).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842 (2010).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Kolberg, L. et al. g:Profiler-interoperable web service for functional enrichment analysis and gene identifier mapping (2023 update). Nucleic Acids Res. 51, W207–W212 (2023).
Kuleshov, M. V. et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 44, W90–W97 (2016).
Szklarczyk, D. et al. The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 51, D638–D646 (2023).
Acknowledgements
We thank A. Kornberg for help with psuedobulk single-cell RNA-seq analysis and C. Wu for help with CD8+ T cell-stimulation experiments. We thank I. Ivanov and Y. Huang for helpful feedback on the papert. We thank B. Jabri for sharing the HLA-DQ8 transgenic mouse (originally created by C. David). Funding was provided from a National Institutes of Health grant (R01 DK102180) to R.Y.-F., A.S., F.B.L. and S.G.
Author information
Authors and Affiliations
Contributions
Conceptualization: R.Y.-F., A.C.-R. and S.G. Formal analysis: R.Y.-F. and A.S. Methodology: R.Y.-F., G.B., A.H. and S.G. Investigation: R.Y.-F., A.S. and I.P.-G. Visualization: R.Y.-F. Funding acquisition: A.C.-R. and S.G. Project administration: R.Y.-F. and S.G. Writing: R.Y.-F. Writing review and editing: F.B.L., G.B. and S.G.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Temporal expression of T cell activation genes.
(a) integrated genome viewer (IGV)16 tracks of stranded mapped reads at the lnc13 locus in unstimulated or TCR stimulated human CD4 T cells (samples SRR11245144 and SRR11245173 from GSE146441). Due to library synthesis method, forward stranded RNA reads are mapped in the inverse orientation due to cDNA synthesis. (b) Expression of Dcp2 in naïve mouse CD4 T cells stimulated with TCR over various time points by RT-qPCR normalizing to Rpl32 housekeeping gene. ΔΔCt was calculated by normalizing to the average of unstimulated (“0”) samples. Data represent mean ± s.e.m.; unpaired, two-tailed, Student’s t-test. N=4. * p <0.05,** p<0.01, *** p<0.001, **** p<0.0001. (c–e) Time-course RNA expression in human CD4+ T cells stimulated via TCR (GSE96538): (c) FOXP3 — delayed induction peaking at 72–96h, resembling lnc13 kinetics; (d) TBX21 — mid-phase induction; (e) IFNG — early induction. Patterns illustrate phased activation of inflammatory vs. regulatory programs during T cell activation. N=3. (f) expression of normalized lnc13 in gluten-specific CD4 T cells (from GSE146441) and general peripheral naïve CD4 T cells (from GSE96538) activated via TCR stimulation. DESeq2 normalized lnc13 reads were further normalized within each dataset to the average of unstimulated (“0”). Data represent mean ± s.e.m.; statistical comparison was calculated through DESeq2. * p <0.05,** p<0.01, *** p<0.001, **** p<0.0001.
Extended Data Fig. 2 lnc13 gene body does not contain regulatory elements.
(a) Schematic of lnc13 knockout. Scissors represent CRISPR guide RNA cut sites. 3 gRNA target each cut site (Supplementary Table 2). Around 2.4kb of mouse lnc13 was cut out without disrupting Il18rap. (b) Cap analysis of gene expression (CAGE-seq) signal (FANTOM5) across the lnc13 locus, visualized in the UCSC genome browser. Signal in red (positive strand) aligns with lnc13 transcription. (c) ENCODE cis-regulatory element (cCRE) annotations at the lnc13 genomic locus. No predicted enhancer activity observed within the lnc13 gene body. (d) Chromatin marks in relevant mouse tissues (kidney, small intestine, spleen, thymus) show absence of canonical enhancer (H3K4me1, H3K27Ac) signatures within the lnc13 gene body, marked in blue (ENCODE database). For small intestines and thymus H3K27Ac and H3K4me1, gray boxes mark statistically significant called peaks. These data confirm that the lnc13 gene body does not contain an active regulatory element or overlapping enhancer.
Extended Data Fig. 3 lnc13−/−-DQ8 mice exhibit transcriptional and humoral features shared with human celiac disease.
(a) Cross-species correlation of top DEGs between lnc13−/−-DQ8 mice and human CeD patients. Mouse DEGs were mapped to human homologs and compared to human biopsy data (GSE134900). Strong correlation observed for proinflammatory genes (Cxcl5, Saa2, Oas2). Correlation was calculated using Pearson’s correlation. (b) Per-gene fold change of differentially expressed immunoglobulin (Ig) transcripts in mouse LPL RNA-seq and human intestinal biopsies (GSE134900). Each dot represents one Ig gene. For the mouse dataset, values are calculated as the fold change of the group-mean expression in lnc13 KO-DQ8 relative to WT-DQ8 LPL RNA-seq. For the human dataset, values are calculated as the fold change of the group-mean expression in celiac disease (CeD) relative to healthy controls (HC). A horizontal line indicates no change (fold change = 1). (c) Serum antibody responses following gluten challenge. lnc13−/−-DQ8 mice exhibit elevated levels of: anti-gliadin IgG, anti-deamidated gliadin peptide (DGP) IgG, anti-tissue transglutaminase (tTG) IgG. Raw optical density (OD) values are shown. Results indicate systemic autoimmune reactivity to gluten-associated antigens (n=7 mice, across 5 independent experiments). Data are shown as mean ± s.e.m.; statistical significance calculated by two-tailed, unpaired student’s t-test. * p <0.05,** p<0.01, *** p<0.001, **** p<0.0001.
Extended Data Fig. 4 Gluten challenged lnc13−/−DQ8 exhibit minimal systemic inflammation.
(a)-(d) ELISA results on the serum levels of TNF-α (A), IL-6 (B), IFN-γ (C), and CRP (D) in gluten-challenged WT-DQ8 and lnc13−/−DQ8 mice. N=5 biological replicates, across 2 independent experiments. (e)-(m) show representative flow cytometry plots for splenic (E)-(G), peripheral lymph node (H)-(J), and blood (K)-(M) frequencies of IFN-γ+ cells of live CD45+ TCRβ+ B220- CD4+ CD8α- cells (E, H, K), IFN-γ+ cells of live CD45+ TCRβ+ B220- CD4- CD8α+ cells (F, I, L), and FOXP3+ cells of live CD45+ TCRβ+ B220- CD4+ CD8α- cells (G, J, M). Quantifications of these data are shown in (q)-(s). N=4 biological replicates, across 2 experiments. (n) Histogram of GZMB expression in blood live CD45+ TCRβ+ B220- CD4- CD8α+ cells. Dotted line shows isotype control while red and blue show lnc13−/−DQ8 and WT-DQ8 samples, respectively. Quantifications of GZMB+ cells and GZMB MFI are shown in (o) and (p), respectively. N=4 biological replicates, across 2 experiments. (t) Representative flow cytometry plot of thymic live CD45+ TCRβ+ B220- cells. (u) Quantification of the frequency of CD4+ CD8α- (left) and CD8α+ CD4- cells (right) of live CD45+ TCRβ+ B220- cells across blood, spleen and peripheral lymph nodes. N=4 biological replicates, across 2 experiments. In scatter-plots, red dots represent samples from lnc13−/−DQ8 mice while blue dots represent samples from WT-DQ8 mice. Error bars represent mean ± s.e.m.; unpaired, two-tailed Student’s t-test. * p <0.05,** p<0.01, *** p<0.001, **** p<0.0001.
Extended Data Fig. 5 Lnc13−/−DQ8 maintained on gluten-free diet does not exhibit cellular markers of inflammation.
(a) Schematic of experiment. WT-DQ8 and lnc13−/−DQ8 mice at 10 weeks old are continuously maintained on gluten-free diet supplemented with sham treatment (PBS) every other day. (b-c, and f-k) Representative flow plots on the left and quantifications across replicates on the right (n=4 mice across 2 experiments). (d and e) Histogram plots on the left, where dotted line denotes isotype control, blue shows cells from WT-DQ8 mice and red shows cells from lnc13−/−DQ8 mice. Middle panels show quantification of frequency of positive cells. Right panel shows MFI quantifications of the shift in population. (b) Frequency of FOXP3+ cells of lamina propria live CD45+ TCRβ+ B220- CD4+ CD8α- cells. (c) Frequency of IFN-γ+ cells of lamina propria live CD45+ TCRβ+ B220- cells, where quantifications are further gated on CD4+ CD8- (“CD4+ T cells”) or CD8+ and CD4- (“CD8+ T cells”). (d) GZMB expression of live CD45+ TCRβ+ TCRγδ- CD8a+ CD8b+ IELs. (e) GZMB expression of live intraepithelial CD45+ TCRβ- TCRγδ- NKG2D+ cells. (f) CD107a+ and CD107b+ double-positive cells of live CD45+ TCRβ+ TCRγδ- CD8α+ CD8β+ IELs. (g) CD107a+ and CD107b+ double-positive cells of live CD45+ TCRβ+ TCRγδ- CD8α+ CD8β- IELs. (h) Show NKG2D+ cells of live CD45+ TCRβ+ TCRγδ- CD8α+ CD8β+ IELs. (i) Show frequency of IFN-γ+ cells of peripheral (inguinal) lymph nodes live CD45+ TCRβ+ B220- CD8α+ CD4- cells. (j) Show frequency of IFN-γ+ cells of peripheral (inguinal) lymph nodes live CD45+ TCRβ+ B220- CD8α- CD4+ cells. (k) Show frequency of FOXP3+ cells of peripheral (inguinal) lymph nodes live CD45+ TCRβ+ B220- CD8α- CD4+ cells. Overall, mice maintained on gluten-free diet does not exhibit cellular markers of inflammation, consistent with the lack of pathology shown in Fig. 4. In scatter-plots, red dots represent samples from lnc13−/−DQ8 mice while blue dots represent samples from WT-DQ8 mice. Error bars represent mean ± s.e.m.; unpaired, two-tailed Student’s t-test. * p <0.05,** p<0.01, *** p<0.001, **** p<0.0001. Image of mouse in a generated by BioRender; Yang-Fischer, R. https://biorender.com/hz505m6 (2026).
Extended Data Fig. 6 Gluten withdrawal reverses cellular markers of inflammation in lnc13−/−DQ8 mice.
(a) schematic of experiment. WT-DQ8 and lnc13−/−DQ8 mice at 10 weeks old are first put on a gluten-containing diet for 45 days along with 20mg of gavaged gluten every other day, then reverted back to a gluten-free diet for 45 days along with sham challenge (PBS) every other day. (b-c, and f-k) Representative flow plots on the left and quantifications across replicates on the right (n=4 mice across 2 experiments). (d, e) Histogram plots on the left, where dotted line denotes isotype control, blue shows cells from WT-DQ8 mice and red shows cells from lnc13−/−DQ8 mice. Middle panels show quantification of frequency of positive cells. Right panel shows MFI quantifications of the shift in population. (b) Frequency of FOXP3+ cells of lamina propria live CD45+ TCRβ+ B220- CD4+ CD8α- cells. (c) Frequency of IFN-γ+ cells of lamina propria live CD45+ TCRβ+ B220- cells, where quantifications are further gated on CD4+ CD8- (“CD4+ T cells”) or CD8+ and CD4- (“CD8+ T cells”). (d) GZMB expression of live CD45+ TCRβ+ TCRγδ- CD8a+ CD8b+ IELs. (e) GZMB expression of intraepithelial live CD45+ TCRβ- TCRγδ- NKG2D+ cells. (f) CD107a+ and CD107b+ double-positive cells of live CD45+ TCRβ+ TCRγδ- CD8α+ CD8β+ IELs. (g) CD107a+ and CD107b+ double-positive cells of live CD45+ TCRβ+ TCRγδ- CD8α+ CD8β- IELs. (h) NKG2D+ cells of live CD45+ TCRβ+ TCRγδ- CD8α+ CD8β+ IELs. (i) frequency of IFN-γ+ cells of peripheral (inguinal) lymph nodes live CD45+ TCRβ+ B220- CD8α+ CD4- cells. (j) frequency of IFN-γ+ cells of peripheral (inguinal) lymph nodes live CD45+ TCRβ+ B220- CD8α- CD4+ cells. (k) frequency of FOXP3+ cells of peripheral (inguinal) lymph nodes live CD45+ TCRβ+ B220- CD8α- CD4+ cells. (l) Number of CD8α+ T IELs in mice maintained on gluten diet (“GF”; with sham challenge, see Fig S10A), challenged with gluten (“gluten”; see Fig. 2a), or transiently challenged with gluten (“G>GF”; see panel A), calculated as previously17. In brief, using histology, the number of IELs per 100 IECs were counted in the terminal ileum. The rest of the small intestine of that sample was processed for IEL extraction, whereby flow cytometry was used to determine percentage of CD8α+ TCRβ+ cells out of live CD45+ live cells. This percentage was multiplied by the # of IELs per 100 IECs to obtain # of CD8α+ T IELs per 100 IECs. Overall, the inflammatory markers of gluten-challenged mice reverted when mice were put back on gluten-free diet, consistent with a rescue of pathology shown in Fig. 4. In scatter-plots, red dots represent samples from lnc13−/−DQ8 mice while blue dots represent samples from WT-DQ8 mice. Error bars represent mean ± s.e.m.; unpaired, two-tailed Student’s t-test. * p <0.05,** p<0.01, *** p<0.001, **** p<0.0001. Image of mouse in a generated by BioRender; Yang-Fischer, R. https://biorender.com/hz505m6 (2026).
Extended Data Fig. 7 Lnc13 expression across cells and cytokine regulation of lnc13 expression.
(a) DESeq2 normalized lnc13 counts across selected immune populations form the Immgen database. (b) Relative lnc13 levels, by RT-qPCR, in splenic CD8+ T cells (live CD45+ TCRβ+ B220- CD8+ CD4-) and small intestinal epithelial cells (upper fraction of percoll gradient in IEL preparation). Levels were normalized by Rpl32 housekeeping gene across samples, then ΔΔCt was calculated relative to average of CD8 T cells. N=4 mice. (c) Time-course RT-qPCR of lnc13 expression in RAW 264.7 macrophages following stimulation with various cytokines: TNF-α: no significant effect, IFN-γ and IL-6: increase lnc13 expression, IL-15: suppresses lnc13 levels. Levels were normalized by Rpl32 housekeeping gene within samples, then ΔΔCt was calculated relative to untreated (0hr) sample for each experiment. N=3 experiments. Induction and suppression were time-dependent and reflect distinct upstream signaling pathways. (d) ReMap Atlas tracks showing public ChIP-seq data for STAT1 and STAT3 binding upstream of lnc13 in T cells, supporting direct transcriptional regulation by IFN-γ and IL-6 pathways. Error bars represent mean ± s.e.m.; comparisons assessed by unpaired, two-tailed Student’s t-test. * p <0.05,** p<0.01, *** p<0.001, **** p<0.0001.
Extended Data Fig. 8 IL-15 downregulates lnc13 and induces Dcp2 expression through STAT5.
(a) RT-qPCR of Il15 expression in whole ileal tissue from WT-DQ8 and lnc13−/−-DQ8 mice. No significant differences observed, confirming that lnc13 does not regulate IL-15 expression at the transcript level (n=3-6). Relative RNA levels, ΔCt were calculated relative to Rpl32 housekeeping gene. (b-c) RT-qPCR of Il15 expression in sorted lamina propria CD11b+ myeloid cells (B; live CD45+ TCRβ- B220- CD11b+) and epithelial cells (C; upper fraction of percoll gradient in IEL preparation) in gluten-challenged mice. Levels were normalized by Rpl32 housekeeping gene within samples, then ΔΔCt was calculated relative to average WT-DQ8 samples. N=4 mice, across two independent experiments. (d) lnc13 and Dcp2 expression in naïve splenic CD8+ T cells 4 hours after soluble IL-15 stimulation. IL-15 induces Dcp2 and suppresses lnc13, consistent with a degradation-based regulatory mechanism. Levels were normalized by Rpl32 housekeeping gene within samples, then ΔΔCt was calculated relative to “no stim” samples in each mice/experiment. N=3 mice, across three independent experiments. (e) ReMap Atlas tracks showing STAT5 ChIP-seq binding peaks at the Dcp2 promoter. (f) STAT5a ChIP-seq or control IgG ChIP-seq at the Dcp2 locus (GSE213507). (g) Normalized expression of DCP2 (counts per million) in control vs STAT5 inhibitor treated cells (GSE286270). These findings support a model in which IL-15 signaling activates STAT5, which transcriptionally induce DCP2 and trigger lnc13 degradation. (h)-(i) Expression of lnc13, by RT-qPCR, in mouse naïve CD4 T cells (H) or naïve CD8 T cells (I) that were unstimulated or stimulated with IL-15SA, TCR (aCD3/aCD28), TCR+IL-15SA or IL-12 for 48h. Levels were normalized by Rpl32 housekeeping gene across samples, then ΔΔCt was calculated relative to unstimulated cells for each mouse. N=4 mice across 2 experiments. (j) Expression of Dcp2 in naïve CD4 and CD8 T cells that were unstimulated or stimulated with IL-15SA for 48h. Levels were normalized by Rpl32 housekeeping gene across samples, then ΔΔCt was calculated relative to average unstimulated samples for both CD4 and CD8 T cells. N=4 mice across 2 experiments. (k-n) Expression of Ifng (K and N), Il2 (L and M) in naïve CD4 T cells (K-L), naïve CD8 T cells (M-N), that were unstimulated or stimulated with IL-15SA, TCR, or TCR+IL-15SA for 48h. Levels were normalized by Rpl32 housekeeping gene across samples, then ΔΔCt was calculated relative to unstimulated cells for each mouse. N=4 mice across 2 experiments. Error bars represent mean ± s.e.m.; comparisons via unpaired, two-tailed Student’s t-test. * p <0.05,** p<0.01, *** p<0.001, **** p<0.0001.
Extended Data Fig. 9 IL-15 suppresses lnc13 in human CD8 T cell subsets and knockout of lnc13 does not affect Il18rap in CD8 T cells.
(a) lnc13 expression in human peripheral pan CD8 T cells after 48-hour stimulation with IL-15. Levels were normalized by RPLP0 housekeeping gene across samples, then ΔΔCt was calculated relative to average unstimulated cells. N=4 donors across 2 experiments. lnc13 is significantly downregulated, mirroring findings in murine CD8 T cells. (b) Expression of lnc13, by RT-qPCR, in human CD8 naïve (CD45RA+ CCR7+), effector memory CD8 (EM; CD45RA− CCR7−), and central memory CD8 T cells (CM; CD45RA− CCR7+). Levels were normalized by RPLP0 housekeeping gene across samples, then ΔΔCt was calculated relative to average naïve CD8 T cells. N=4 biological replicates. (c-e) Expression of lnc13, by RT-qPCR, in human naïve (c), EM (d), or CM CD8+ T cells (e) that were unstimulated or stimulated with IL-15, TCR (aCD3/aCD28), or TCR+IL-15 for 48h. Levels were normalized by RPLP0 housekeeping gene across samples, then ΔΔCt was calculated relative to unstimulated cells. N=4 biological replicates across 2 experiments. (f) CD8α+ T IELs were cultured ex vivo in the presence of IL-15SA treatment. Representative flow plots of IFN-γ expression of live cells are shown on the left. Quantifications are shown on the right. N=4 mice across 3 independent experiments. (g) Expression of Il18rap in mouse naïve CD8 T cells that were unstimulated, or stimulated for 48h with IL-15SA, anti-CD3/CD28 (TCR), or TCR + IL-15SA. Relative Il18rap levels were calculated by normalizing to Rpl32 housekeeping gene within each sample and ΔΔCt was calculated by normalizing to the average of unstimulated WT cells. Red dots denote KO cells while blue dots denote WT cells. In all scatter-plots, error bars represent mean ± s.e.m.; comparisons via unpaired, two-tailed Student’s t-test. * p <0.05,** p<0.01, *** p<0.001, **** p<0.0001.
Extended Data Fig. 10 Overlap of lnc13 RNA-seq DEGs and RAP-seq significantly called peaks and conserved function of lnc13 in human CD8 T cells.
(a) Venn diagram showing overlap of lnc13 DEGs from in vivo isolated CD8αβ T IELs post-gluten challenge (left circle) vs IL-15 inducible genes in CD8α T IELs from different genotypes (right circle). Overlap shows shared genes, where the percentage indicates percentage of in vivo DEGs. Upper Venn diagram shows comparison with IL-15 inducible genes in lnc13−/− CD8α+ T IELs, and lower Venn diagram shows comparison with those in WT CD8α+ T IELs. (b-c) Human peripheral blood CD8 T cells were nucleofected with siRNA against lnc13 or control siRNA (siNeg) that were unstimulated (B) or stimulated with IL-15 (C). Then, levels of various lnc13-target genes were examined by RT-qPCR. RNA levels were normalized by RPLP0 housekeeping gene across samples, then ΔΔCt was calculated relative to the siNeg sample (control siRNA) for each donor. N=4 donors across 2 experiments. Error bars represent mean ± s.e.m.; statistical comparisons by unpaired t-test after correcting for multiple testing. * p <0.05, ** p<0.01, *** p<0.001, **** p<0.0001. (d) Venn diagrams showing overlap of lnc13 significantly called RAP peaks from primary mouse peripheral CD8 T cells (on the left, numbers in red) and DEGs of lnc13−/− vs WT CD8α T IELs post IL-15 stimulation (on the right, numbers in black). The overlap shows the numbers of lnc13 RAP peaks (red numbers) that overlaps with various distances from lnc13 DEGs (black numbers). Top, middle, and lower Venn diagram shows comparison of RAP peaks with +/- 5 kb, +/- 50kb, or +/- 500kb of lnc13 DEG gene bodies, respectively.
Supplementary information
Supplementary Information (download PDF )
Supplementary Figs. 1–6 and Table 2.
Supplementary Table 1 (download XLSX )
Table of differentially expressed lncRNAs in gluten-specific lamina propria CD4+ T cells activated via TCR stimulation.
Supplementary Table 3 (download XLSX )
Table of DEGs in lnc13−/−-DQ8 versus WT-DQ8 CD8ab T IELs after gluten challenge RNA-seq.
Supplementary Table 4 (download XLSX )
Table of DEGs in lnc13−/− versus WT CD8a T IELs after ex vivo IL-15SA stimulation.
Supplementary Table 5 (download XLSX )
Table of IL-15 inducible genes in WT CD8a T IELs.
Supplementary Table 6 (download XLSX )
Table of IL-15 inducible genes in lnc13−/− CD8a T IELs.
Supplementary Data 1 (download XLSX )
Source data for Supplementary Fig. 1.
Supplementary Data 2 (download XLSX )
Source data for Supplementary Fig. 2.
Source data
Source Data Fig. 1 (download XLSX )
Graph raw values source data.
Source Data Fig. 2 (download XLSX )
Graph raw values source data.
Source Data Fig. 3 (download XLSX )
Graph raw values source data.
Source Data Fig. 4 (download XLSX )
Graph raw values source data.
Source Data Fig. 5 (download XLSX )
Graph raw values source data.
Source Data Fig. 6 (download XLSX )
Graph raw values source data.
Source Data Extended Data Fig. 1 (download XLSX )
Graph raw values source data.
Source Data Extended Data Fig. 3 (download XLSX )
Graph raw values source data.
Source Data Extended Data Fig. 4 (download XLSX )
Graph raw values source data.
Source Data Extended Data Fig. 5 (download XLSX )
Graph raw values source data.
Source Data Extended Data Fig. 6 (download XLSX )
Graph raw values source data.
Source Data Extended Data Fig. 7 (download XLSX )
Graph raw values source data.
Source Data Extended Data Fig. 8 (download XLSX )
Graph raw values source data.
Source Data Extended Data Fig. 9 (download XLSX )
Graph raw values source data.
Source Data Extended Data Fig. 10 (download XLSX )
Graph raw values source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yang-Fischer, R., Shearer, A., Leão, F.B. et al. The long noncoding RNA lnc13 restrains inflammatory responses to maintain oral tolerance to gluten. Nat Immunol (2026). https://doi.org/10.1038/s41590-026-02506-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41590-026-02506-6
