
Abstract
Seizures are made up of the coordinated activity of networks of neurons, suggesting that control of neurons in the pathologic circuits of epilepsy could allow for control of the disease. Optogenetics has been effective at stopping seizure-like activity in non-human disease models by increasing inhibitory tone or decreasing excitation, although this effect has not been shown in human brain tissue. Many of the genetic means for achieving channelrhodopsin expression in non-human models are not possible in humans, and vector-mediated methods are susceptible to species-specific tropism that may affect translational potential. Here we demonstrate adeno-associated virus–mediated, optogenetic reductions in network firing rates of human hippocampal slices recorded on high-density microelectrode arrays under several hyperactivity-provoking conditions. This platform can serve to bridge the gap between human and animal studies by exploring genetic interventions on network activity in human brain tissue.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

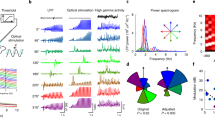
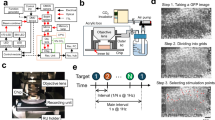
Data availability
Electrophysiological data will be made available on a DANDI public server at: https://dandiarchive.org/dandiset/001132
Other data that support the findings of this study are available in the GitHub repository: https://github.com/braingeneers/AndrewsGengVoitiukEtAl_SourceCode. Any additional data will be made available by the authors upon reasonable request.
Code availability
An annotated summary of custom code used for this study is available at https://github.com/braingeneers/AndrewsGengVoitiukEtAl_SourceCode.
MaxWell software, purchased from MaxWell Biosystems, was used for data collection.
Kilosort2 (ref. 98) was used for electrophysiologic data analysis.
All code will be made available upon reasonable request.
References
Staley, K. Molecular mechanisms of epilepsy. Nat. Neurosci. 18, 367–372 (2015).
Staley, K. J. & Dudek, F. E. Interictal spikes and epileptogenesis. Epilepsy Curr. 6, 199–202 (2006).
Paz, J. T. & Huguenard, J. R. Microcircuits and their interactions in epilepsy: is the focus out of focus? Nat. Neurosci. 18, 351–359 (2015).
Crick, F. H. Thinking about the brain. Sci. Am. 241, 219–233 (1979).
Boyden, E. S. Optogenetics and the future of neuroscience. Nat. Neurosci. 18, 1200–1201 (2015).
Lin, S.-C., Deisseroth, K. & Henderson, J. M. Optogenetics: background and concepts for neurosurgery. Neurosurgery 69, 1–3 (2011).
Boyden, E. S., Zhang, F., Bamberg, E., Nagel, G. & Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 8, 1263–1268 (2005).
Gradinaru, V. et al. Molecular and cellular approaches for diversifying and extending optogenetics. Cell 141, 154–165 (2010).
Zhang, F. et al. Optogenetic interrogation of neural circuits: technology for probing mammalian brain structures. Nat. Protoc. 5, 439–456 (2010).
Krook-Magnuson, E., Armstrong, C., Oijala, M. & Soltesz, I. On-demand optogenetic control of spontaneous seizures in temporal lobe epilepsy. Nat. Commun. 4, 1376 (2013).
Krook‐Magnuson, E. et al. In vivo evaluation of the dentate gate theory in epilepsy. J. Physiol. 593, 2379–2388 (2015).
Fradot, M. et al. Gene therapy in ophthalmology: validation on cultured retinal cells and explants from postmortem human eyes. Hum. Gene Ther. 22, 587–593 (2011).
Sahel, J.-A. et al. Partial recovery of visual function in a blind patient after optogenetic therapy. Nat. Med. 27, 1223–1229 (2021).
Avoli, M. et al. Epileptiform activity induced by low extracellular magnesium in the human cortex maintained in vitro. Ann. Neurol. 30, 589–596 (1991).
Masukawa, L. M., Higashima, M., Kim, J. H. & Spencer, D. D. Epileptiform discharges evoked in hippocampal brain slices from epileptic patients. Brain Res. 493, 168–174 (1989).
Cohen, I., Navarro, V., Clemenceau, S., Baulac, M. & Miles, R. On the origin of interictal activity in human temporal lobe epilepsy in vitro. Science 298, 1418–1421 (2002).
Köhling, R. & Avoli, M. Methodological approaches to exploring epileptic disorders in the human brain in vitro. J. Neurosci. Methods 155, 1–19 (2006).
Andersson, M. et al. Optogenetic control of human neurons in organotypic brain cultures. Sci. Rep. 6, 24818 (2016).
Ledri, M., Andersson, M., Wickham, J. & Kokaia, M. Optogenetics for controlling seizure circuits for translational approaches. Neurobiol. Dis. 184, 106234 (2023).
Wykes, R. C. et al. Optogenetic and potassium channel gene therapy in a rodent model of focal neocortical epilepsy. Sci. Transl. Med. 4, 161ra152 (2012).
Wykes, R. C., Kullmann, D. M., Pavlov, I. & Magloire, V. Optogenetic approaches to treat epilepsy. J. Neurosci. Methods 260, 215–220 (2016).
Jones, R. S., da Silva, A. B., Whittaker, R. G., Woodhall, G. L. & Cunningham, M. O. Human brain slices for epilepsy research: pitfalls, solutions and future challenges. J. Neurosci. Methods 260, 221–232 (2016).
Grone, B. P. & Baraban, S. C. Animal models in epilepsy research: legacies and new directions. Nat. Neurosci. 18, 339–343 (2015).
Bosque, J. R. et al. Molecular tools for the characterization of seizure susceptibility in genetic rodent models of epilepsy. Epilepsy Behav. 121, 106594 (2021).
Lévesque, M., Avoli, M. & Bernard, C. Animal models of temporal lobe epilepsy following systemic chemoconvulsant administration. J. Neurosci. Methods 260, 45–52 (2016).
Armstrong, C., Krook-Magnuson, E., Oijala, M. & Soltesz, I. Closed-loop optogenetic intervention in mice. Nat. Protoc. 8, 1475–1493 (2013).
Matsuzaki, Y. et al. Intravenous administration of the adeno-associated virus-PHP.B capsid fails to upregulate transduction efficiency in the marmoset brain. Neurosci. Lett. 665, 182–188 (2018).
Hordeaux, J. et al. The neurotropic properties of AAV-PHP.B are limited to C57BL/6J mice. Mol. Ther. 26, 664–668 (2018).
Zhu, D. et al. Optimal trade-off control in machine learning based library design, with application to adeno-associated virus (AAV) for gene therapy. Sci. Adv. 10, eadj3786 (2024).
Zhu, D., Schieferecke, A. J., Lopez, P. A. & Schaffer, D. V. Adeno-associated virus vector for central nervous system gene therapy. Trends Mol. Med. 27, 524–537 (2021).
Chen, X. et al. Engineered AAVs for non-invasive gene delivery to rodent and non-human primate nervous systems. Neuron 110, 2242–2257 (2022).
Goertsen, D. et al. AAV capsid variants with brain-wide transgene expression and decreased liver targeting after intravenous delivery in mouse and marmoset. Nat. Neurosci. 25, 106–115 (2022).
Chan, K. Y. et al. Engineered AAVs for efficient noninvasive gene delivery to the central and peripheral nervous systems. Nat. Neurosci. 20, 1172–1179 (2017).
Deverman, B. E., Ravina, B. M., Bankiewicz, K. S., Paul, S. M. & Sah, D. W. Gene therapy for neurological disorders: progress and prospects. Nat. Rev. Drug Discov. 17, 641–659 (2018).
Andrews, M. R. Gene therapy in the CNS—one size does not fit all. Gene Ther. 28, 393–395 (2021).
Ting, J. T. et al. A robust ex vivo experimental platform for molecular-genetic dissection of adult human neocortical cell types and circuits. Sci. Rep. 8, 8407 (2018).
Le Duigou, C. et al. Imaging pathological activities of human brain tissue in organotypic culture. J. Neurosci. Methods 298, 33–44 (2018).
Schwarz, N. et al. Long-term adult human brain slice cultures as a model system to study human CNS circuitry and disease. eLife 8, e48417 (2019).
Blümcke, I. et al. International consensus classification of hippocampal sclerosis in temporal lobe epilepsy: a Task Force report from the ILAE Commission on Diagnostic Methods. Epilepsia 54, 1315–1329 (2013).
Govorunova, E. G. et al. Kalium channelrhodopsins are natural light-gated potassium channels that mediate optogenetic inhibition. Nat. Neurosci. 25, 967–974 (2022).
Vierock, J. et al. WiChR, a highly potassium-selective channelrhodopsin for low-light one-and two-photon inhibition of excitable cells. Sci. Adv. 8, eadd7729 (2022).
Buchin, A. et al. Multi-modal characterization and simulation of human epileptic circuitry. Cell Rep. 41, 111873 (2022).
Ma, Z., Turrigiano, G. G., Wessel, R. & Hengen, K. B. Cortical circuit dynamics are homeostatically tuned to criticality in vivo. Neuron 104, 655–664 (2019).
Mele, M. et al. Transient incubation of cultured hippocampal neurons in the absence of magnesium induces rhythmic and synchronized epileptiform-like activity. Sci. Rep. 11, 11374 (2021).
Mody, I., Lambert, J. & Heinemann, U. Low extracellular magnesium induces epileptiform activity and spreading depression in rat hippocampal slices. J. Neurophysiol. 57, 869–888 (1987).
Keller, C. J. et al. Heterogeneous neuronal firing patterns during interictal epileptiform discharges in the human cortex. Brain 133, 1668–1681 (2010).
Lévesque, M. & Avoli, M. The kainic acid model of temporal lobe epilepsy. Neurosci. Biobehav. Rev. 37, 2887–2899 (2013).
Vezzani, A. et al. Interleukin-1β immunoreactivity and microglia are enhanced in the rat hippocampus by focal kainate application: functional evidence for enhancement of electrographic seizures. J. Neurosci. 19, 5054–5065 (1999).
Jimenez-Mateos, E. M. et al. Silencing microRNA-134 produces neuroprotective and prolonged seizure-suppressive effects. Nat. Med. 18, 1087–1094 (2012).
Huberfeld, G. et al. Glutamatergic pre-ictal discharges emerge at the transition to seizure in human epilepsy. Nat. Neurosci. 14, 627–634 (2011).
Herman, A. M., Huang, L., Murphey, D. K., Garcia, I. & Arenkiel, B. R. Cell type-specific and time-dependent light exposure contribute to silencing in neurons expressing Channelrhodopsin-2. eLife 3, e01481 (2014).
Lee, E. K. et al. Non-linear dimensionality reduction on extracellular waveforms reveals cell type diversity in premotor cortex. eLife 10, e67490 (2021).
Lee, K., Carr, N., Perliss, A. & Chandrasekaran, C. WaveMAP for identifying putative cell types from in vivo electrophysiology. STAR Protoc. 4, 102320 (2023).
Masuda, F. K., Aery Jones, E. A., Sun, Y. & Giocomo, L. M. Ketamine evoked disruption of entorhinal and hippocampal spatial maps. Nat. Commun. 14, 6285 (2023).
McInnes, L. & Healy, J. Accelerated hierarchical density based clustering. In Proc. of the 2017 IEEE International Conference On Data Mining Workshops (ICDMW) (eds Gottumukkala, R. et al.) https://doi.org/10.1109/ICDMW.2017.12 (IEEE, 2017).
Tønnesen, J., Sørensen, A. T., Deisseroth, K., Lundberg, C. & Kokaia, M. Optogenetic control of epileptiform activity. Proc. Natl Acad. Sci. USA 106, 12162–12167 (2009).
Castillo, P. E., Malenka, R. C. & Nicoll, R. A. Kainate receptors mediate a slow postsynaptic current in hippocampal CA3 neurons. Nature 388, 182–186 (1997).
Savtchenko, L. P. et al. Disentangling astroglial physiology with a realistic cell model in silico. Nat. Commun. 9, 3554 (2018).
Ecker, A. et al. Data‐driven integration of hippocampal CA1 synaptic physiology in silico. Hippocampus 30, 1129–1145 (2020).
Chiang, C. C., Shivacharan, R. S., Wei, X., Gonzalez‐Reyes, L. E. & Durand, D. M. Slow periodic activity in the longitudinal hippocampal slice can self‐propagate non‐synaptically by a mechanism consistent with ephaptic coupling. J. Physiol. 597, 249–269 (2019).
Schmalz, J., Quinarez, R. V., Kothare, M. V. & Kumar, G. Controlling neocortical epileptic seizures using forced temporal spike-time stimulation: an in silico computational study. Front. Comput. Neurosci. 17, 1084080 (2023).
Buzsáki, G. Theta oscillations in the hippocampus. Neuron 33, 325–340 (2002).
Kleen, J. K. et al. Bidirectional propagation of low frequency oscillations over the human hippocampal surface. Nat. Commun. 12, 2764 (2021).
Zhang, H. & Jacobs, J. Traveling theta waves in the human hippocampus. J. Neurosci. 35, 12477–12487 (2015).
Goyal, A. et al. Functionally distinct high and low theta oscillations in the human hippocampus. Nat. Commun. 11, 2469 (2020).
Lubenov, E. V. & Siapas, A. G. Hippocampal theta oscillations are travelling waves. Nature 459, 534 (2009).
Goutagny, R., Jackson, J. & Williams, S. Self-generated theta oscillations in the hippocampus. Nat. Neurosci. 12, 1491–1493 (2009).
Etter, G., van der Veldt, S., Choi, J. & Williams, S. Optogenetic frequency scrambling of hippocampal theta oscillations dissociates working memory retrieval from hippocampal spatiotemporal codes. Nat. Commun. 14, 410 (2023).
Pernía-Andrade, A. J. & Jonas, P. Theta-gamma-modulated synaptic currents in hippocampal granule cells in vivo define a mechanism for network oscillations. Neuron 81, 140–152 (2014).
Diamond, J. M. et al. Interictal discharges in the human brain are travelling waves arising from an epileptogenic source. Brain 146, 1903–1915 (2023).
Cadwell, C. R. et al. Electrophysiological, transcriptomic and morphologic profiling of single neurons using Patch-seq. Nat. Biotechnol. 34, 199–203 (2016).
Rahmati, N. et al. Unique actions of GABA arising from cytoplasmic chloride microdomains. J. Neurosci. 41, 4957–4975 (2021).
Yao, Z. et al. A taxonomy of transcriptomic cell types across the isocortex and hippocampal formation. Cell 184, 3222–3241 (2021).
Mott, D. D., Turner, D. A., Okazaki, M. M. & Lewis, D. V. Interneurons of the dentate–hilus border of the rat dentate gyrus: morphological and electrophysiological heterogeneity. J. Neurosci. 17, 3990–4005 (1997).
Jaffe, A. E. et al. Profiling gene expression in the human dentate gyrus granule cell layer reveals insights into schizophrenia and its genetic risk. Nat. Neurosci. 23, 510–519 (2020).
Murphy, B. L. et al. Heterogeneous integration of adult-generated granule cells into the epileptic brain. J. Neurosci. 31, 105–117 (2011).
Grossberger, L., Battaglia, F. P. & Vinck, M. Unsupervised clustering of temporal patterns in high-dimensional neuronal ensembles using a novel dissimilarity measure. PLoS Comput. Biol. 14, e1006283 (2018).
Caldwell, D. et al. Signal recovery from stimulation artifacts in intracranial recordings with dictionary learning. J. Neural Eng. 17, 026023 (2020).
Ayhan, F. et al. Resolving cellular and molecular diversity along the hippocampal anterior-to-posterior axis in humans. Neuron 109, 2091–2105 (2021).
Mich, J. K. et al. Functional enhancer elements drive subclass-selective expression from mouse to primate neocortex. Cell Rep. 34, 108754 (2021).
Deverman, B. E. et al. Cre-dependent selection yields AAV variants for widespread gene transfer to the adult brain. Nat. Biotechnol. 34, 204–209 (2016).
Dimidschstein, J. et al. A viral strategy for targeting and manipulating interneurons across vertebrate species. Nat. Neurosci. 19, 1743–1749 (2016).
Chuapoco, M. R. et al. Adeno-associated viral vectors for functional intravenous gene transfer throughout the non-human primate brain. Nat. Nanotechnol. 18, 1241–1251 (2023).
Wiebe, S., Blume, W. T., Girvin, J. P. & Eliasziw, M. A randomized, controlled trial of surgery for temporal-lobe epilepsy. N. Engl. J. Med. 345, 311–318 (2001).
Ma, B. B. & Rao, V. R. Responsive neurostimulation: candidates and considerations. Epilepsy Behav. 88, 388–395 (2018).
Razavi, B. et al. Real‐world experience with direct brain‐responsive neurostimulation for focal onset seizures. Epilepsia 61, 1749–1757 (2020).
Khambhati, A. N., Shafi, A., Rao, V. R. & Chang, E. F. Long-term brain network reorganization predicts responsive neurostimulation outcomes for focal epilepsy. Sci. Transl. Med. 13, eabf6588 (2021).
Bansal, A., Shikha, S. & Zhang, Y. Towards translational optogenetics. Nat. Biomed. Eng. 7, 349–369 (2023).
Priya, R. et al. Activity regulates cell death within cortical interneurons through a calcineurin-dependent mechanism. Cell Rep. 22, 1695–1709 (2018).
Nair, D. R. et al. Nine-year prospective efficacy and safety of brain-responsive neurostimulation for focal epilepsy. Neurology 95, e1244–e1256 (2020).
Ellens, N. R., Elisevich, K., Burdette, D. E. & Patra, S. E. A comparison of vagal nerve stimulation and responsive neurostimulation for the treatment of medically refractory complex partial epilepsy. Stereotact. Funct. Neurosurg. 96, 259–263 (2018).
Haneef, Z. & Skrehot, H. C. Neurostimulation in generalized epilepsy: a systematic review and meta‐analysis. Epilepsia 64, 811–820 (2023).
Skrehot, H. C., Englot, D. J. & Haneef, Z. Neuro-stimulation in focal epilepsy: a systematic review and meta-analysis. Epilepsy Behav. 142, 109182 (2023).
Ma, B. B. et al. Responsive neurostimulation for regional neocortical epilepsy. Epilepsia 61, 96–106 (2020).
Panov, F. et al. Safety of responsive neurostimulation in pediatric patients with medically refractory epilepsy. J. Neurosurg. Pediatr. 26, 525–532 (2020).
Ryvlin, P. & Jehi, L. E. Neuromodulation for refractory epilepsy. Epilepsy Curr. 22, 11–17 (2022).
Ryvlin, P., Rheims, S., Hirsch, L. J., Sokolov, A. & Jehi, L. Neuromodulation in epilepsy: state-of-the-art approved therapies. Lancet Neurol. 20, 1038–1047 (2021).
Pachitariu, M., Steinmetz, N. A., Kadir, S. N., Carandini, M. & Harris, K. D. Fast and accurate spike sorting of high-channel count probes with KiloSort. In Proc. 30th International Conference on Neural Information Processing Systems (NIPS 16) (eds Lee, D. D. et al.) 4455–4463 (Curran Associates, 2016).
Rossant, C. et al. Spike sorting for large, dense electrode arrays. Nat. Neurosci. 19, 634–641 (2016).
Siegle, J. H. et al. Survey of spiking in the mouse visual system reveals functional hierarchy. Nature 592, 86–92 (2021).
Hill, D. N., Mehta, S. B. & Kleinfeld, D. Quality metrics to accompany spike sorting of extracellular signals. J. Neurosci. 31, 8699–8705 (2011).
Someck, S. et al. Positive and biphasic extracellular waveforms correspond to return currents and axonal spikes. Commun. Biol. 6, 950 (2023).
Leland, M., John, H. & James, M. Uniform manifold approximation and projection for dimension reduction. Preprint at https://arxiv.org/abs/1802.03426 (2018).
Izhikevich, E. M. Simple model of spiking neurons. IEEE Trans. Neural Netw. 14, 1569–1572 (2003).
Spreizer, S. et al. NEST 3.3 (2022); https://juser.fz-juelich.de/record/1007653
Linssen, C. et al. NESTML 5.2.0 (2023); https://juser.fz-juelich.de/record/1005203
Acknowledgements
This project was supported by the National Institute of Neurological Disorders and Stroke, National Institutes of Health (NIH), through UCSF grant 5R25NS070680-13 (J.P.A.). Its contents are solely the responsibility of the authors and do not necessarily represent the views of the NIH. This study was also supported by the following NIH awards: Brain Initiative award UF1MH130700 (D.V.S., V.S. and T.J.N.); R01NS123263 (T.J.N.); R01MH120295 (S.R.S.); T32HG012344 (K.V.); and K08NS126573 (C.R.C.). In addition, this study was supported by the National Science Foundation (NSF) under award NSF 2034037 (M.T.), the National Human Genome Research Institute under award RM1HG011543 (M.A.T.E., A.R., A.S., D.S., D.E., D.H., S.S. and M.T.) and grants from the Schmidt Futures Foundation (SF 857) (D.H.), the Weill Neurohub (C.R.C.), U01NS132353 (C.R.C), the Esther A. & Joseph Klingenstein Fund (T.J.N.), the Shurl and Kay Curci Foundation (T.J.N. and C.R.C.) and the Sontag Foundation (T.J.N.) as well as a gift from the William K. Bowes Jr. Foundation. T.J.N. is a New York Stem Cell Foundation Robertson Neuroscience Investigator. J.L.S. was supported by the University of California, Santa Cruz (UCSC) Chancellor’s Postdoctoral Fellowship, NIH K12GM139185 (through the National Institute of General Medical Sciences to the UCSC Institute for the Biology of Stem Cells) and LRP0000018281 (National Institute of Child Health and Human Development). Support was also provided by the Citizens United for Research in Epilepsy (CURE) Taking Flight Award (C.R.C). We thank Flaticon for images used to design Fig. 2. We thank the Pacific Research Platform, supported by the NSF under awards CNS-1730158, ACI-1540112, ACI-1541349 and OAC-1826967; the University of California Office of the President; the University of California, San Diego’s California Institute for Telecommunications and Information Technology/Qualcomm Institute; as well as the Corporation for Education Network Initiatives in California. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript. Materials were obtained and supplied by the authors. We would like to acknowledge K. Hengen for feedback in manuscript preparation. We would like to acknowledge the courageous patients who agreed to have their treatments contribute to the advancement of knowledge and science.
Author information
Authors and Affiliations
Contributions
J.P.A., J.G. and K.V. contributed equally. J.P.A. conceived the project, obtained tissue, designed experiments, conducted experiments, analyzed the data, interpreted the data, performed histology and wrote the manuscript. J.G. constructed the analysis pipeline, analyzed data, interpreted results, created figures and drafted the manuscript. K.V. built and fabricated the optogenetic slice recording system, wrote the code for integration of the MEA system with optogenetic recordings, conducted experiments, wrote experimental code, created figures and drafted the manuscript. M.A.T.E., D. Shin, A.R. and A.S. also contributed equally. M.A.T.E. conducted experiments, wrote experimental code, designed the closed-loop experiment, analyzed theta wave phase propagation, created theta oscillation videos and created figures. D. Shin designed, tested and optimized the tissue culture pipeline, designed AAV plasmids, assisted in experimental design and provided feedback on manuscript drafting. A.R. designed code for MEA analysis and designed figures. A.S. designed the in silico modeling of hippocampal epileptiform activity, created the in silico modeling figure, designed and implemented the HDBSCAN analyses and created the HDBSCAN analysis figure. Other co-author contributions are as follows. A.W. cultured hippocampal slices and performed and analyzed histology. L.L. consulted on AAV design and edited the manuscript. D. Solis designed theta coherence k-distance analysis, conducted local field potential analyses and assisted with figure design. M.G.K. and J.L.S. contributed to MEA recording and optogenetic experimental design. J.A.R.d.J. transduced tissue and edited the manuscript. K.C.D. performed whole-cell patch clamping and analysis. H.H.L. cultured and processed tissue. D.E. contributed to hardware design and MEA recording techniques. K.I.A. provided mentorship and experimental tissue. S.S., V.S., T.S., D.H., C.R.C. and D.V.S. provided mentorship, intellectual consultation, experimental design input and analytic methods advice and edited the manuscript. E.F.C. provided mentorship, intellectual consultation and human tissue samples and edited the manuscript. M.T. provided mentorship and intellectual consultation, oversaw data analysis and rig fabrication, contributed to experimental design and edited the manuscript. T.J.N. conceived the project, designed experiments, interpreted data and wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Neuroscience thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Slice histology.
Representative Immunohistochemistry overlay from 7 of the 12 slices listed in Table 1. a-f, hippocampal slices transduced with AAV9-CaMKIIa-HcKCR1-eYFP. g-i, 1 of 2 slices transduced with AAV9-CaMKIIa-ChR2-eYFP. Magenta represents NeuN and cyan represents eYFP expression, correlating to neurons that express the HcKCR1-eYFP or ChR2-eYFP construct. a-c, Examples of slices where predominately CA regions were recorded. a, Slice 3C, b, slice 8D, c, slice 12G. d-f, Examples of slices where predominately granule cell layer (GCL) of the dentate gyrus areas were recorded. d, Slice 9E, e, slice 10F, and f, slice 11G. Scale bars all represent 500 microns. g-i, Slice 4C, transduced with AAV9-CaMKIIa-ChR2-eYFP.
Extended Data Fig. 2 Patch clamp of HcKCR1.
a, DIC microscopic image of a neuron in a hippocampal slice transduced with AAV9-CaMKIIa-HcKCR1-eYFP. b, Green fluorescence of the same cell as in a, confirming viral transduction. c, Voltage clamp recording of the cell above showing hyperpolarizing currents activated with brief pulses of 540nm light illumination, represented by green rectangles above the hyperpolarizing currents elicited by illumination. This was repeated in 3 cells over 2 separate hippocampal slices.
Extended Data Fig. 3 In silico model.
Raster plots of simulated activity of an in silico model of granule and basket cells of the human hippocampus. Each black dot represents a single-unit firing event, so dark areas represent clusters of high firing rates consistent with epileptiform activity. a, Examples of spontaneous seizure-like events in presence of no light-responsive cells. A simulated ‘light-on’ phase is indicated in green. b-d, Raster plots of the simulation data with b, 10%, c, 25% and d, 50% of excitatory granule cells harboring an inhibitory, light-sensitive channel.
Extended Data Fig. 4 Optogenetic inhibition in physiologic and bicuculline media.
a, Stacked raster plots of hippocampal slice 3C (n = 64 units) expressing HcKCR1 illuminated with 10s of continuous 530nm LED light. Average firing rate (Hz) is overlaid in red, with shaded red representing standard error of the mean, for multiple trials of 10s of light illumination (green rectangle from 10–20s on stacked raster plots), and paired comparisons of b, firing rate and c, spike amplitude (uV) for the 10s prior to illumination (pre), during illumination (light ON) and following illumination (OFF). On the y-axis are 5 trials (t1 – t5) of continuous 10s illumination, stacked align the light ON phase (green). d-f) Same parameters as shown in a-c, are shown for slice 5C (n = 65 units), recorded with bicuculline. g-i) same parameters for slice 7D (n = 44 units), recorded with bicuculline. j-l, for slice 8D (n = 10 units), recorded with bicuculline. Firing rate and amplitude comparisons were made with a two-tailed paired t-test comparing average firing rates and amplitudes under each condition. See Supplementary Table 4 for P-values for each comparison. For all box and whisker plots, the box extends from the first quartile to the third quartile of the data, with a line at the median. The whiskers extend from the box to the farthest data point lying within 1.5x the inter-quartile range from the box. For stacked raster plots (a, d, g, j) the translucent pink color surrounding the firing rate red line represents the standard error of the mean (SEM) for the firing rate. *P-values were judged to be significant if adjusted P-values were less than 0.05. Exact P-values reported in Supplementary Table 4.
Extended Data Fig. 5 Optogenetic inhibition in 0-mg media.
a, Stacked raster plots from Slice 9E (n = 17 units) with average firing rate (Hz) overlaid in red, with shaded red representing standard error of the mean, for multiple trials of 10s of light illumination, and paired comparisons of b, firing rate and c, spike amplitude for the 10s prior to illumination (pre), during illumination (light ON) and following illumination (OFF). Data shown from 4 slices recorded in 0-mg media. d, Raster plot with firing-rate overlay showing isolated bursts of activity in trials T8 and T10 in slice 10F (n = 228 units). e-f, firing rate and amplitude data from slice 10F (n = 228 units). g-i, Slice 11G (n = 23 units), showing isolated bursts of activity in trials T1 and T2. j-l, Slice 12G (n = 9 units). Firing rate and amplitude comparisons were made with a two-tailed paired t-test comparing average firing rates and amplitudes under each condition. See Supplementary Table 5 for P-values for each comparison. For all box and whisker plots, the box extends from the first quartile to the third quartile of the data, with a line at the median. The whiskers extend from the box to the farthest data point lying within 1.5x the inter-quartile range from the box. * P-values were judged to be significant if adjusted P-values were less than 0.05. Exact P-values reported in Supplementary Table 5.
Extended Data Fig. 6 Optogenetic inhibition in 0-mg + KA media.
a, Stacked raster plots of Slice 9E (n = 57 units) with average firing rate (Hz) overlaid in red, with shaded red representing standard error of the mean, for multiple trials of 10s of light illumination, and paired comparisons of b, firing rate (Hz) and c, spike amplitude (uV) for the 10s prior to illumination (pre), during illumination (light ON) and following illumination (OFF). Data shown for 4 slices recorded under 0-mg+KA conditions. d-f, data from Slice 10F (n = 212 units). d, slice 10F exhibits rhythmic burst activity, with persistent bursts of during trials T6-T8. e-f, Neuronal firing rate and amplitude. g-i, data from slice 11G (n = 55 units). g, Slice 11G exhibiting clusters of rhythmic burst activity that does not persist during light-ON conditions. h-i, Neuronal firing rate and amplitude. j-l, data from slice 12G (n = 17 units). All slices show reduced firing rates during light-ON conditions. See Supplementary Table 6 for P-values for each comparison. For all box and whisker plots, the box extends from the first quartile to the third quartile of the data, with a line at the median. The whiskers extend from the box to the farthest data point lying within 1.5x the inter-quartile range from the box. Firing rate and amplitude comparisons were made with a two-tailed paired t-test comparing average firing rates and amplitudes under each condition. * P-values were judged to be significant if adjusted P-values were less than 0.05, exact P-values reported in Supplementary Table 6.
Extended Data Fig. 7 Intensity sweeps.
Unit activity with firing rate overlaid during the intensity sweeps for 3 separate slices. a,b, Intensity scans of 2 slices (a = slice 3C, b = slice 5C) recorded in physiologic media, c, intensity scan of a slice (c = slice 10F) recorded in 0-mg media.
Extended Data Fig. 8 ChR2 activation.
a, Stacked raster plots from slice 4C (n = 30 units) with average unit firing rate (Hz) overlaid in red, with shaded red representing standard error of the mean, showing ChR2-mediated increases in firing rate during light-ON conditions. b, intensity sweep, c, firing rate comparisons and d, spike amplitude comparisons for the same slice. Firing rate statistics for a two-tailed paired t-test comparing average firing rates and amplitudes under each condition, n = 30 units (c): Pre vs light-ON raw P-value = 3.22E-04, holm-bonferroni adjusted P-value = 0.001. Light-ON vs OFF raw P-value = 7.34E-04, holm-bonferroni adjusted P-value = 0.001. Pre vs OFF raw P-value = 0.106, holm-bonferroni adjusted P-value = 0.106. Amplitude statistics, n = 30 units (d): Pre vs light-ON raw P-value = 0.127, holm-bonferroni adjusted P-value = 0.254. Light-ON vs OFF raw P-value = 0.027, holm-bonferroni adjusted P-value = 0.082. Pre vs OFF raw P-value = 0.106, holm-bonferroni adjusted P-value = 0.255. For all box and whisker plots, the box extends from the first quartile to the third quartile of the data, with a line at the median. The whiskers extend from the box to the farthest data point lying within 1.5x the inter-quartile range from the box.*comparisons were deemed statistically significant if adjusted P-values were less than P = 0.05.
Extended Data Fig. 9 Zero-Mg spikes.
a, Slice 10F and b, 9E showing isolated <1s bursts of activity during 5 minutes of continuous recording.
Supplementary information
Supplementary Information (download PDF )
Supplementary Figs. 1–3 and Supplementary Tables 1–6.
Supplementary Video 1 (download MOV )
Video of real-time spike activity raster and heatmap with approximate overlay on histology of slice 3C in physiologic media during LED light illumination.
Supplementary Video 2 (download MOV )
Video of real-time spike activity raster and heatmap with approximate overlay on histology of slice 5C in physiologic media with addition of bicuculline during LED light illumination.
Supplementary Video 3 (download MP4 )
Video of theta wave phase propagation from local field potential recordings in slice 10F with 0-Mg media after addition of KA. Red and blue represent opposite phases of the theta wave, with size of dots representing the magnitude of the phase relative to zero.
Supplementary Video 4 (download MP4 )
Video of theta wave phase propagation from local field potential recordings in slice 11GF with 0-Mg media after addition of KA. Red and blue represent opposite phases of the theta wave, with size of dots representing the magnitude of the phase relative to zero.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Andrews, J.P., Geng, J., Voitiuk, K. et al. Multimodal evaluation of network activity and optogenetic interventions in human hippocampal slices. Nat Neurosci 27, 2487–2499 (2024). https://doi.org/10.1038/s41593-024-01782-5
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41593-024-01782-5
