
Abstract
Local protein synthesis is vital for neuronal function, but its dysregulation in neurodegenerative diseases remains poorly defined. Here we applied spatial transcriptomics to adult mouse motor nerve axons and cell bodies to enable subcellular mapping. Among transcripts found in mature axons, the most enriched biological process is protein translation, and localization of translation machinery was confirmed using multiplexed single-molecule spatial transcriptomics combined with immunofluorescence. Amyotrophic lateral sclerosis (ALS)-associated mutations in the RNA-binding protein fused in sarcoma (FUS), which suppress local translation, disrupt the compartment-specific RNA signatures, including components of the translation machinery. In particular, eukaryotic initiation factor 5a (Eif5a), a translation factor involved in elongation and termination, is found to be locally impaired in mutant FUS axons with reduced levels of its active hypusinated form. Axon-specific treatment with polyamine spermidine restores Eif5a hypusination and ameliorates mutant FUS-dependent neuronal defects, including suppression of local protein synthesis. Finally, in vivo spermidine treatment reduces ALS-related toxicity in mutant FUS and TDP-43 Drosophila models, which may have implications for therapy development.
Similar content being viewed by others

Main
Local translation is a fundamental mechanism sustaining axonal maintenance, growth, branching and regeneration1. In vitro transcriptomic studies have revealed a few thousand transcripts in axons and dendrites, with components of the cytoskeleton, mitochondrion, ribosome and translation machinery highly localized in distal neuronal compartments2. The complex architecture of motor neurons extending their long axonal processes ending in specialized synapses, called neuromuscular junctions (NMJs), makes distal maintenance highly dependent on efficient local protein translation and degradation independent of the soma1. Therefore, it is not surprising that several motor neuron diseases and hereditary neuropathies, including spinal muscular atrophy3,4, Charcot–Marie–Tooth disease5,6,7 and amyotrophic lateral sclerosis (ALS)8, show local translation defects.
ALS is a neurodegenerative disorder caused by the death of motor neurons, which is proposed to be preceded by NMJ denervation and axonal degeneration9. Mutations in fused in sarcoma (FUS) cause familial ALS10,11 and are associated with rare forms of frontotemporal lobar degeneration (FTLD)12,13, whereby its aggregation is a major hallmark in patients14,15. FUS protein is located mainly in the nucleus, where it regulates DNA repair and RNA metabolism, but it also exerts fundamental roles in the cytoplasm related to mRNA stability, transport and translation, miRNA processing, liquid–liquid phase separation, mitochondrial biology and autophagy16. FUS also localizes in dendrites, spines, in proximity of presynaptic and postsynaptic vesicles, mature axons and at the NMJs8,17,18,19,20,21,22,23. Notably, ALS-linked mutations in the nuclear localization signal of FUS24, that is, R521H and R521C, cause aberrant accumulation of the mutant protein in axons, which leads to decreased local axonal translation and increased axon-specific stress activation through the integrated stress response pathway in motor neurons8. The mechanisms underlying mutant FUS-dependent aberrant local translation impacting early axon integrity in disease are not yet known.
Here through compartment-specific spatial transcriptomics, we unveil the mRNA composition of axonal subtypes in adult mouse nerves. Focusing on spinal motor axons, we investigated the local transcriptome during FUS-mediated neurodegeneration, which is associated with reduced intra-axonal protein translation. We further identified axon-specific alterations in the composition of the translation machinery. We find reduced eukaryotic initiation factor 5a (Eif5a) hypusination, specifically in axons of cultured neurons as well as adult sciatic nerves of mutant FUS mice before disease development8. Finally, spermidine treatment in cultured axons rescues the reduction in local translation caused by decreased hypusination, which alleviates FUS-mediated impairments, including reduced axonal network and disrupted neuronal activity. Notably, spermidine further reduces toxicity caused by expression of disease-causing FUS and TDP-43 in fly motor neurons. These findings uncover new mechanistic insights with relevance for developing therapeutic strategies to treat ALS and other neuromuscular disorders.
Results
Transcriptomic signature of spinal cord motor neuron somas uncovered through integrative spatial transcriptomics and single-nuclei RNA sequencing
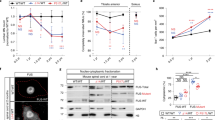
To spatially resolve the transcriptomic composition of adult mouse spinal cord lower motor neuron cell bodies and their respective axons in a compartment-specific manner, we applied spatial transcriptomics compatible with immunostaining to distinguish the neuronal and axonal populations of interest. First, we performed comparative unbiased transcriptomic analysis of digitally spatially profiled, pooled motor neuron cell bodies (<20 per section) and nonmotor soma in 12-month-old lumbar spinal cord tissue sections (choline acetyltransferase (ChAT)-positive/neurofilament H (NFH)-negative motor neurons (orange) versus ChAT-negative/NFH-negative nonmotor neurons (purple); Fig. 1a), using the GeoMx digital spatial profiling (DSP) technology from NanoString25. Ultraviolet (UV)-photocleavable probes against the whole-mouse transcriptome were collected from the two distinct pooled neuronal types, and libraries were further prepared for sequencing (Fig. 1a).

a, Experimental schematic of the workflow applied to identify motor neuron-specific transcripts using UV-photocleavable RNA probes against the whole-mouse genome in combination with cell-specific antibodies using the NanoString GeoMx DSP spatial transcriptomic platform. Top, representative immunofluorescence image of a mouse spinal cord section using ChAT and NFH antibodies to identify motor neuron cell bodies (left inset) and nonmotor neurons (right inset). Middle, representative image from the digital segmentation applied using GeoMX DSP software. Bottom, next-generation sequencing of UV light-laser-released RNAs extracted from pooled motor neurons or non-motor neurons. Scale bars, 500 µm and 50 µm (inset). Schematics in a were created with BioRender.com. b, Volcano plot of the DEGs between motor neurons (orange) and nonmotor neurons (purple). c, Heatmap of the top 20 DEGs in motor and nonmotor neurons. Colored scale, scaled log2-normalized counts. d, Multiplexed smFISH of 15 known cell markers in the mouse spinal cord. Immunofluorescence signal of NFH is reported in white. Scale bars, 500 µm; insets, 50 µm. e, Spatial integration of the transcriptomic analyses of mouse spinal cord using unbiased spatial transcriptomics (10x Genomics Visium) and the GeoMx DSP DEGs. Each dot represents a spatial transcriptional signature of the 10x Genomics Visium capture area, which is color-coded based on the normalized enrichment score (NES) for the motor neuron signature (left) and nonmotor neuron signature (center). On the right, the motor (orange) and nonmotor neuron (purple) dots derived from the analysis are reported. f,g, Spatial transcriptomic profiling (10x Genomics Visium) of mouse spinal cord motor neuron-expressing Kcnj14 (f) and Tmsb10 (g) transcripts. Scale bar of colored dots, log2 (gene expression + 1). Scale bars, 500 µm (left), 100 µm (right). h, Representative immunofluorescence images of the respective Kcnj14 and Tmsb10 proteins in the spinal cord ventral horns. Motor neurons are labeled with ChAT antibody (from two independent experiments). Scale bar, 100 µm. Bottom, magnification of the dashed white square in h. Scale bar, 50 µm. NS, not significant; MN, motor neurons; non-MN, nonmotor neurons.
To evaluate the purity of the UV light-guided selection of the two regions of interest (ROIs) corresponding to the pooled motor and nonmotor neurons, we selected a subset of cell-specific genes known to be present in the mouse spinal cord26 (Extended Data Fig. 1a). While both ChAT-positive and ChAT-negative neurons were enriched for pan-neuronal markers (Nefl, Syp, Grin1 and Kcna1) compared to astrocytic (S100b) and other glial markers (Mog, Mag, Aif1, Itgam, Itgax, Ptprc and Cx3cr1) as expected26, a distinct transcriptomic signature was identified between the two neuronal subtypes by differential expression (DE) analysis driven by the enrichment (log2 fold change (FC) >2) of cholinergic markers in ChAT-positive compared to ChAT-negative neurons. The genes found to be most significantly enriched in the ChAT-positive motor neurons are known motor neuron markers27,28,29,30 (including Chat, Slc18a3 (ref. 31), Slc5a7 (ref. 32) and Chodl 33; Fig. 1b,c), further validating the sensitivity and specificity of the approach. Through the use of multiplexed combinatorial single-molecule fluorescence in situ hybridization (smFISH; Molecular Cartography), cell-type-specific transcripts, including motor neuron subtype markers Chat and Chodl, were confirmed at a single-cell resolution in adult mouse spinal cords (Fig. 1d and Extended Data Fig. 1b–j).
To gain spatial information on our transcriptomic dataset, we applied unbiased barcoded Visium spatial transcriptomics (10x Genomics) to 12-month-old mouse spinal cord sections. We found that (1) the ChAT-positive RNA signature obtained by digital spatial segmentation mapped to transcriptomic areas localized in the ventral horn, as expected, (2) while the ChAT-negative RNA signature shown in Fig. 1b,c mapped to medial gray and dorsal horn areas (Fig. 1e; top four genes found to be most enriched in the ChAT-positive (Extended Data Fig. 1k) or ChAT-negative ROIs (Extended Data Fig. 1l)). To validate our findings from unbiased spatial transcriptomics, we conducted multiplexed smFISH targeting a highly expressed neuronal transcript (including in motor neurons (stathmin 2 (Stmn2)34) and a low abundant motor neuron transcript (galectin 1 (Lgals1)30). We found consistent results in the expression patterns with both approaches (Extended Data Figs. 1m–o).
To identify potential new transcripts that are expressed in motor neurons, we compared publicly available single-nuclei RNA-sequencing (snRNA-seq) datasets27,28,29,30 with our DSP data (Extended Data Fig. 1p). ChAT-positive and ChAT-negative nuclei were selected among the neuronal cell population from each published dataset, and the differentially expressed genes (DEGs) across the four studies and ours were compared (Supplementary Table 1). Sixty-three genes were commonly found in all five datasets (Extended Data Fig. 1p). Overall, we observe a concordance (R2 between 0.66 and 0.45) in fold expression changes in our DSP data compared to previously published snRNA-seq studies (Extended Data Fig. 1q–t), with 206 genes identified in common between our dataset and the most concordant analyzed study28. Additionally, 201 genes were uniquely identified in our DSP analysis (Extended Data Fig. 1p). Validation through unbiased barcoded spatial transcriptomics (Fig. 1f,g) and immunofluorescence (Fig. 1h) confirmed that Kcnj14 (uniquely found expressed in motor neurons in the comparison between our dataset and the others) and Tmsb10 (one of the genes that was most significantly enriched in our dataset and not previously validated at the protein level) are expressed in the ventral horn. This expression was notably prominent in motor neurons and observed, to a lesser extent, in other spinal cord neurons. Altogether, our multi-omic integrative approach identifies a robust motor neuron-enriched gene signature.
Distinct transcriptomic signatures in adult motor and nonmotor axons using spatial transcriptomics
To map the transcriptome of adult motor axons in an unbiased manner, GeoMx DSP was performed in 12-month-old mouse sciatic nerve sections using the same approach as outlined above (Fig. 2a). Through ChAT and NFH immunofluorescence costaining (Fig. 2a), motor (ChAT-positive/NFH-positive (green)) and nonmotor axons (ChAT-negative/NFH-positive (purple)) were successfully distinguished by digital segmentation, which further allowed for performing whole-transcriptomic sequencing of the pooled motor and nonmotor axons. Gene enrichment analysis of the most highly expressed genes in mature adult axons revealed as top hit transcripts related to translation, synapse and ribonuclear complex (Fig. 2b and Supplementary Table 2). Interestingly, 82% of the transcripts known to encode ribosomal proteins were detected in our dataset. To validate their axonal localization, we sought to apply multiplexed smFISH combined with immunostaining to label axons in the same tissue section (Fig. 2c), given the high resolution achieved in spinal cord sections (Fig. 1d). To establish feasibility, we included probes for key resident cell types (Fig. 2c and Extended Data Fig. 2), such as Schwann cells (Pmp22), endoneurial fibroblasts (Cspg4) and macrophages (Cd68), and demonstrated subcellular spatial resolution. We then analyzed and further confirmed the intra-axonal localization of newly identified axonal transcripts, including Eef2, Rpl4 and Rpl15, which are linked to translation (Extended Data Fig. 3a; with 5.4%, 6.7% and 8.1%, respectively, of the total transcripts localizing inside NFH-positive axons; Extended Data Fig. 3f), Hnrnpa2b1 (which belongs to the ‘Ribonucleoprotein complex’ Gene Ontology (GO) term, with 24.5% of Hnrnpa2b1 transcripts being intra-axonal; Extended Data Fig. 3b,f), Sparc (which is part of the ‘Synapse’ GO term, 13.6% of which localized in axons; Extended Data Fig. 3c), as well as transcripts previously described to localize in axons including Cox4i1, a mitochondrial respiratory chain component with 13.2% being axonal transcripts (Extended Data Fig. 3d,f), and Nefh and Nefl, two components of the neuronal cytoskeleton (Extended Data Fig. 3e; with 35.1% of Nefl and 52.3% of Nefh transcripts being detected in axons; Extended Data Fig. 3F). Altogether, the multiplexed smFISH analysis further validates the spatial axonal transcriptomic analysis.

a, Experimental schematic of the workflow applied to profile mouse adult sciatic nerve axons as detailed in Fig. 1a using the NanoString GeoMX DSP spatial transcriptomic platform. Scale bars, 100 µm; inset, 10 µm. Schematics in a were created with BioRender.com. b, GO analysis of the axonal transcripts identified in both motor and nonmotor axons. c, Multiplexed smFISH of 12 known cell markers in transversal (top) and longitudinal (bottom) sections of mouse sciatic nerve. Scale bars, 100 µm; insets, 10 µm. d, Heatmap of the top 20 DEGs in motor axons and nonmotor axons. Colored scale, scaled log2-normalized counts. e, Volcano plot of the DEGs between motor axons (blue) and nonmotor axons (purple). f,h, Lao1 (f) and Art3 (h) mRNA levels in motor (blue) and nonmotor axons (purple) from the GeoMx DSP transcriptomic analysis. Data are represented as mean ± s.e.m. (n = 4). Each data point corresponds to the measurement in one section. P = 0.002 (f) and 0.005 (h) using an unpaired two-tailed t test. g,i, Quantifications (left) and representative immunofluorescence images (right) of the respective Lao1 (g) and Art3 (i) proteins in longitudinal mouse sciatic nerve sections. Axons are labeled with NFH antibody and motor axons with anti-ChAT antibody. ROIs were selected based on NFH signal and assigned to motor or nonmotor classifications based on ChAT signal. Scale bar, 5 µm. Data are represented as mean ± s.e.m. (n = 4). P = 0.017 (g), 0.049 (i) using unpaired two-tailed t test. *P < 0.05; **P < 0.01.
Next, we characterized the motor and nonmotor axonal transcriptomes obtained from the sequencing of pooled motor (ChAT-positive and NFH-positive) and nonmotor (NFH-positive only) axons (Fig. 2d,e). DEG analysis revealed distinct RNA signatures, where Lao1, Or10a4, Sh2d2a, St14 and Etnppl were the most enriched genes in motor axons, whereas Duxf3, Art3, Csk and Apoe were the most enriched genes in the nonmotor axons (Fig. 2d). Despite the low total counts of Lao1 and Art3 (around 10–20 normalized counts; Fig. 2f,h, respectively), the axonal localization of their respective protein products was confirmed by immunofluorescence, with a clear enrichment of Lao1 in ChAT-positive motor axons (Fig. 2g) and Art3 in nonmotor axons of mouse sciatic nerves (Fig. 2i). These data uncover the transcriptomic diversity of motor and nonmotor axons in adult sciatic nerves using unbiased spatial transcriptomics.
Impaired accumulation of translation machinery components in mutant FUS motor axons
ALS-linked mutations in FUS compromise local axonal mRNA translation before disease development in mutant FUS mice8, suggesting that these axonal defects may contribute to ALS-linked phenotypes; however, the underlying molecular mechanisms are unknown. To map the mutant FUS-dependent RNA composition in motor axons, which are most vulnerable in ALS, we performed GeoMx DSP in sciatic nerves of humanized mutant FUS mice expressing the ALS-linked FUSR521H mutation (mFUSKO/hFUSR521H), which develop ALS-like motor phenotypes8 (Fig. 3a). DE analysis of 12-month-old motor axons from mFUSKO/hFUSR521H mice compared to nontransgenic animals revealed a distinct mutant-dependent RNA signature with the most upregulated transcripts being Duxf3, Acsf2, Gm52436 and B2m, while the most downregulated transcripts being Zbtb12, Dohh, Cpb2 and Micall1 (Extended Data Fig. 4a). Gene enrichment analysis of the DEGs between nontransgenic and mFUSKO/hFUSR521H motor axons (Extended Data Fig. 4b,c) revealed a mutant FUS-dependent upregulation of transcripts involved in translation. Interestingly, several transcripts encoding translation factors (such as Eif5a, Eif4a1, Eif1 and Eif4g3) and ribosomal proteins (such as Rps2, Rps15a, Rps17, Rpl17, Rpl35a and Rpl13a) were collectively dysregulated, the majority being upregulated in mFUSKO/hFUSR521H sciatic nerve motor axons (Fig. 3b,c). In contrast, their transcript levels remained unchanged in their respective spinal cord motor neuron cell bodies of mFUSKO/hFUSR521H mice compared to nontransgenic animals, with the exception of Eif1 transcript, whose levels were decreased in mutant FUS motor cell bodies (Extended Data Fig. 4d). This was also confirmed in spinal cord sections analyzed with the 10x Visium spatial transcriptomics platform, revealing that these translation-related transcripts are largely unchanged in motor neuron somas expressing mutant FUS (Extended Data Fig. 4e,f).

a, Representative images of a transversal section of a 12-month-old mFUSKO/hFUSR521H mouse sciatic nerve analyzed using the NanoString GeoMX DSP spatial transcriptomic platform as detailed in Figs. 1a and 2a. Scale bars, 100 µm; insets, 10 µm. b, Volcano plot of the DEGs between motor axons from 12-month-old nontransgenic (purple) and mFUSKO/hFUSR521H (pink) sciatic nerve animals. c, Dysregulated mRNA levels of transcripts encoding protein translation machinery components in nontransgenic (purple) and mutant FUS (pink) axons from the GeoMx DSP analysis. Data are represented as mean ± s.e.m. (n = 4). Each data point corresponds to the measurement in one section. P = 0.032 (Eif1), 0.016 (Eif4a1), 0.008 (Eif4g3), 0.015 (Rpl17), 0.021 (Rpl13a), 0.006 (Rpl35a), 0.018 (Rps15a), 0.036 (Rps17) and 0.008 (Rps2) using an unpaired two-tailed t test. d–k, Representative immunofluorescence images, their quantifications and mRNA levels of Eif4g3 (d,e), Rpl24 (f,g), Rps3a (h,i) and Rps4x (j,k) in longitudinal sections of mouse sciatic nerves. All axons are labeled with NFH antibody, and motor axons with ChAT antibody. Data are represented as mean ± s.e.m. (n = 8–15 for immunofluorescence; n = 4 for RNA quantifications). P = 0.034 (Eif4g3—protein), 0.038 (Rpl24—protein), 0.011 (Rpl24—RNA), 0.008 (Rps3a—protein), 0.121 (Rps3a—RNA), 0.015 (Rps4x—protein) and 0.861 (Rps4x—RNA) using an unpaired two-tailed t test. Scale bar, 5 µm. l–q, Representative immunofluorescence images (left) and quantifications (right) in transversal sections of mouse spinal cords of Rpl24 (l,m), Rps3a (n,o) and Rps4x (p,q). Motor axons are labeled with ChAT antibody. Scale bar, 100 µm. Data points represent the average of at least three ROIs in one section. Data are represented as mean ± s.e.m. (three to five sections per animal from three to six independent biological replicates). P = NS using an unpaired two-tailed t test. *P < 0.05; **P < 0.01.
To determine whether the mutant FUS-dependent axonal transcriptomic dysregulation observed for components of the translation machinery is recapitulated at the protein level, we performed immunofluorescence imaging of elongation factor Eif4g3 as well as ribosomal protein subunits (including Rpl24, Rps3a and Rps4x, previously found to be present in Xenopus laevis axons35,36). Eif4g3 protein levels were decreased in motor axons (Fig. 3d,e). A 2.5-fold increased accumulation of Rpl24 (Fig. 3f,g), along with a 1.5-fold reduction of Rps3a and Rps4x levels (Fig. 3h–k), was also observed in mutant FUS motor axons, but their respective levels remained unchanged in motor neuron cell bodies of adult mutant FUS mouse spinal cords, both at the RNA (Extended Data Fig. 4g) and protein level (Fig. 3l–q). Altogether, these findings validate the mutant FUS-dependent axon-specific transcriptomic dysregulation identified by GeoMx DSP of adult sciatic nerve axons.
Unique post-translational hypusination of Eif5a is decreased in mutant FUS motor axons
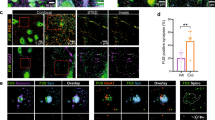
One of the most downregulated transcripts in the mutant FUS motor axons when compared to nontransgenic mice was Dohh, which encodes for deoxyhypusine hydroxylase/monooxygenase protein (Fig. 3b). Dohh is an enzyme involved in hypusination, a post-translational modification that uniquely occurs in Eif5a, a translation factor regulating key steps of protein synthesis37,38,39. Hypusine is a modified amino acid derived from the arginine/ornithine/putrescine/spermidine polyamine pathway. It is exclusively present in Eif5a in all eukaryotes and has an essential role in its activity40,41,42,43,44,45. We investigated the polyamine biosynthesis and metabolism pathway in motor axons of mFUSKO/hFUSR521H mice, including the Eif5a hypusination pathway involving deoxyhypusine synthase (Dhps)–Dohh axis (Fig. 4a). Interestingly, all transcripts encoding the enzymes involved in the production and catabolism of spermidine/spermine (including arginase 1, ornithine decarboxylase, spermidine synthase, spermine synthase and spermidine/spermine N1-acetyltransferase 1) were detected in adult motor axons, with similar mRNA counts in both nontransgenic and mutant FUS mice (Fig. 4b). Dhps and Dohh mRNAs, which encode the two enzymes that catalyze the first and last step in the conversion of lysine 50 of Eif5a into hypusine, respectively (Fig. 4a), were also detected in motor axons of control sciatic nerves. While the Dhps mRNA and protein levels remained unaffected by the FUS mutation (Fig. 4c,d), Dohh mRNA and protein levels were significantly reduced (Fig. 4e,f) in mutant FUS motor axons compared to age-matched nontransgenic controls, but not in spinal cord motor neuron cell bodies (Extended Data Fig. 5a). Similarly, Eif5a protein levels were decreased by 62% in 12-month-old motor axons of mFUSKO/hFUSR521H mouse sciatic nerves (Fig. 4h), although an increase in Eif5a mRNA levels was found by DSP (Fig. 4g), with a similar nonstatistically significant trend observed by multiplexed smFISH in the mutant motor axons (Fig. 4i–m). Eif5a mRNA and protein levels remained unchanged in spinal cord motor neuron cell bodies (Extended Data Fig. 5b,c). Hypusine levels (evaluated by immunofluorescence using an antibody specifically recognizing hypusinated Eif5a only (Extended Data Fig. 5d)) were significantly decreased in mutant FUS motor axons by 23% (Fig. 4n), and an increase, although not significant, was detected in motor neuron cell bodies (Extended Data Fig. 5e) of mFUSKO/hFUSR521H mice compared to nontransgenic animals, further suggesting a mutant FUS-dependent depletion of active Eif5a at the periphery. Consistently, no significant differences in Eif5a, Dohh and Dhps mRNA levels were detected in the spinal cord motor cell bodies using 10x Visium spatial transcriptomics (Extended Data Fig. 5f,g). Eif5a was shown to be inactivated through the acetylation of a residue located near the hypusinated lysine 50 (ref. 46). We found that in mFUSKO/hFUSR521H motor axons, acetylated Eif5a levels were increased by 2.5-fold compared to nontransgenic motor axons (Fig. 4o), in line with the decreased levels of the active form of hypusinated Eif5a (Fig. 4n). Taken together, these data support an axon-specific impairment of Eif5a activation linked to its unique post-translational hypusination modification, which may underlie local translation defects caused by ALS-linked FUS mutations8.

a, Schematic representation of the polyamine biosynthetic and metabolic pathway. The active form of Eif5a requires a unique post-translational modification, which is the incorporation of a hypusine amino acid at lysine 50, using spermidine as a substrate through a two-step enzymatic reaction involving Dhps and Dohh. b, Levels of transcripts encoding the enzymes involved in the conversion of arginine to spermine from the GeoMx DSP transcriptomic analysis of motor axons. Data are represented as mean ± s.e.m. (n = 4). P = NS, using an unpaired two-tailed t test. Each data point corresponds to the measurement in one section. c,e,g, mRNA quantification of the targets of interest in sciatic nerve motor axons in nontransgenic (purple) and mFUSKO/hFUSR521H (pink) animals. Data are represented as mean ± s.e.m. (n = 4). P = NS (c), 0.004 (e) and 0.015 (g) using an unpaired two-tailed t test. Each data point corresponds to the measurement in one section. d,f,h,n,o, Representative immunofluorescence images (left panels) and respective protein levels of the indicated target (immunofluorescence, right in sciatic nerve motor axons in nontransgenic (purple) and mFUSKO/hFUSR521H (pink) animals. Scale bar, 5 µm. Data are represented as mean ± s.e.m. (n = 3–8 biological replicates for immunofluorescence). P = NS (d), 0.005 (f), 0.017 (h), 0.023 (n) and 0.018 (o) using an unpaired two-tailed t test. i, Multiplexed smFISH of Eif5a transcript in nontransgenic sciatic nerve transversal section combined with NFH immunofluorescence to label axons. The indicated squares in i are insets shown in j. Scale bar, 100 µm (i). j, Insets indicated in i. Light blue arrows point to detected Eif5a transcripts. Scale bar, 5 µm (j). k, Multiplexed smFISH of nontransgenic (left) and mFUSKO/hFUSR521H (right) sciatic nerve longitudinal sections. The indicated blue squares represent the insets shown in l. Scale bar, 500 µm (k). l, Insets indicated in k. Light blue arrows point to detected Eif5a transcripts. Scale bar, 5 µm (l). m, Quantification of the Eif5a mRNAs detected inside NFH-positive axons using multiplexed smFISH. Data are represented as mean ± s.e.m. P = NS using an unpaired t test with four sections per animal was applied. In k,l,m, the dots are manually enlarged for visualization purposes. *P < 0.05; **P < 0.01.
Spermidine treatment rescues local translation defects in mutant FUS axons through Dohh-mediated hypusination
To determine whether mutant FUS-induced axonal translation suppression is caused by the reduced active form of Eif5a, we first treated primary cortical neurons derived from hFUSR521H and nontransgenic littermates with spermidine (100 µM for 48 h) to evaluate its impact on Eif5a hypusination and mutant FUS-dependent deficits8 (Fig. 5a,b). While hypusine levels were decreased by 25% in mutant FUS cortical axons (Fig. 5c), spermidine treatment restored axonal hypusinated Eif5a to normal levels. The defects in hypusinated Eif5a were accompanied by a 47% decrease in axonal translation (as assessed after a short pulse of puromycin incorporation), as previously reported in hippocampal neurons8 (Fig. 5d). When spermidine was added, puromycin incorporation in hFUSR521H cortical neurons reached 87% compared to control neurons, nearly restoring the amount of protein synthesis to the levels measured in nontreated control axons. Spermidine treatment also restored the mutant FUS-dependent reduction in the proximal axonal area to normal levels (Fig. 5e). Furthermore, FUS mutation decreased total Eif5a levels by 30% and increased inactive acetylated Eif5a levels in cortical axons. Spermidine treatment restored both total Eif5a (Extended Data Fig. 6a,c) and acetylated Eif5a (Extended Data Fig. 6b,d) axonal levels in hFUSR521H cortical neurons. These results are in line with a spermidine-dependent axonal accumulation of active hypusinated Eif5a. Notably, mutant FUS-expressing cell bodies (Extended Data Fig. 7a) did not show significant alterations in hypusine levels (Extended Data Fig. 7b) or puromycin incorporation (Extended Data Fig. 7c) as previously reported in hippocampal neurons8, suggesting axon-specific mechanisms. To further evaluate the functionally beneficial effects of spermidine in more mature neuronal cultures, we measured spontaneous neuronal activity using multielectrode arrays (MEA) of 1-month-old mutant FUS cortical neurons (Fig. 5f–h), as neuronal activity was previously shown to be reduced in mutant FUS neurons8. In agreement with previous findings, a mutant FUS-dependent reduction in burst duration and number of spikes per burst was measured in 1-month-old cortical neurons (Fig. 5g,h) by 65% and 28%, respectively. Treatment with spermidine significantly restored these functional deficits, close to those observed in nontransgenic neurons. Taken together, spermidine-dependent rescue of mutant FUS-induced axonal translation defects leads to improved functional activity of mutant FUS cortical neurons.

a, Experimental schematic illustrating the workflow used in primary cortical neurons cultured in multiwell dishes. b, Representative immunofluorescence images of primary cortical neurons derived from nontransgenic and hFUSR521H animals, treated with vehicle or SPMD 100 µM for 48 h, labeled with hypusine (active Eif5a), puromycin (protein synthesis) and NFH (axons) antibodies. Scale bars, 20 µm; inset, 5 µm. c,d, Quantification of the respective immunofluorescence signal in proximal axons of primary cortical neurons treated with vehicle or SPMD 100 µM for 48 h from b. Data are represented as mean ± s.e.m. (n = 3 biological replicates). P = 0.046 (c, non-tg versus hFUSR521H), 0.014 (c, hFUSR521H versus hFUSR521H + SPMD), 0.004 (d, non-tg versus hFUSR521H) and 0.024 (d, hFUSR521H versus hFUSR521H + SPMD) using one-way ANOVA with Bonferroni multiple comparisons test. e, Quantification of the axonal network (proximal NFH area). Data are represented as mean ± s.e.m. (n = 3 biological replicates). P = 0.043 (non-tg versus hFUSR521H) and 0.045 (hFUSR521H versus hFUSR521H + SPMD) using one-way ANOVA with Bonferroni multiple comparisons test. f, Representative network activity and bursts of nontransgenic and hFUSR521H cortical neurons cultured in MEA and treated with SPMD (10 µM for 48 h). The orange boxes above the representative burst recordings illustrate the duration of a burst. g,h, Quantification of burst duration (g) and spikes per burst (h) from the respective recordings reported in f. Data are represented as mean ± s.e.m. (n = 3 biological replicates). P = 0.004 (g, non-tg versus hFUSR521H), 0.045 (g, hFUSR521H versus hFUSR521H + SPMD) and 0.026 (h, non-tg versus hFUSR521H) using one-way ANOVA with Bonferroni multiple comparisons test. i, Experimental schematic illustrating the workflow used in primary cortical neurons cultured in multiwell dishes treated with siRNAs (control or against Dohh) in combination with SPMD. j, Representative immunofluorescence images of primary cortical neurons derived from nontransgenic and hFUSR521H animals, treated with or without SPMD 75 µM for 48 h and labeled with Dohh, puromycin (protein synthesis) and NFH (axons) antibodies. Scale bar, 20 µm. k, Insets of j. Scale bar, 5 µm. l,m, Quantification of the respective immunofluorescence signals from j. Data are represented as mean ± s.e.m. (n = 3 biological replicates). P = 0.002 (l, non-tg siControl versus non-tg siDOHH), NS (l, non-tg siControl versus hFUSR521H siControl), NS (l, hFUSR521H siControl versus hFUSR521H siDOHH), NS (l, hFUSR521H siDOHH versus hFUSR521H siDOHH + SPMD), <0.0001 (m, non-tg siControl versus non-tg siDOHH), <0.0001 (m, non-tg siControl versus hFUSR521H siControl), 0.0004 (m, hFUSR521H siControl versus hFUSR521H siControl + SPMD), NS (m, hFUSR521H siDOHH versus hFUSR521H siDOHH + SPMD) using one-way ANOVA with Bonferroni multiple comparisons test. Schematics in a and i were created with BioRender.com. SPMD, spermidine; ANOVA, analysis of variance. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
To gain further mechanistic insight and determine whether the improved levels of local translation by spermidine were mediated by Dohh-induced Eif5a hypusination, we silenced Dohh (using siRNAs) in control and mutant FUS cortical neurons in the presence or absence of spermidine (Fig. 5i). Dohh silencing (Fig. 5l) significantly reduced local translation in nontransgenic axons by more than half (Fig. 5m), suggesting that Dohh is critical for axonal translation. Dohh silencing also abrogated spermidine-dependent rescue of axonal translation defects caused by mutant FUS (Fig. 5j–m). These findings suggest that spermidine rescue acts at least in part through Dohh-dependent hypusination of Eif5a.
Axon-specific spermidine treatment ameliorates Eif5a hypusination, axonal translation and length defects in mutant FUS motor neurons
To test the effects of compartment-specific spermidine treatment, we then cultured primary motor neurons, which are most vulnerable in ALS, in microfluidic devices that allow separation of cell bodies from axons (Fig. 6a). Dose-dependent spermidine treatment (that is, 100 and 200 µM for 48 h) of the axonal compartment only (Fig. 6b–f), revealed a 29% and 57% increase, respectively, in hypusine levels in mutant FUS axons compared to vehicle-treated hFUSR521H axons, while hypusine levels were reduced by 41% in hFUSR521H motor axons compared to nontransgenic controls (Fig. 6c). This was accompanied by improved axonal translation reduction compared to nontreated hFUSR521H motor neurons (Fig. 6d). As observed in cortical neurons (Extended Data Fig. 7a–c), hypusine/puromycin levels were not altered in hFUSR521H motor cell bodies (Extended Data Fig. 7d–f).

a, Experimental schematic illustrating the workflow used in primary motor neurons cultured in compartmented microfluidic chambers with treatment of spermidine in the axonal compartment only. Schematics in a were created with BioRender.com. b, Representative immunofluorescence images of primary motor axons derived from nontransgenic and hFUSR521H animals in the axonal compartment, treated with or without spermidine 100 and 200 µM for 48 h and labeled with hypusine (active Eif5a), puromycin (protein synthesis) and NFH (axons) antibodies. Scale bar, 10 µm. c,d, Quantification of the respective immunofluorescence signals from b. Data are represented as mean ± s.e.m. (n = 4 biological replicates). P = 0.001 (c, non-tg versus hFUSR521H), 0.036 (c, hFUSR521H versus hFUSR521H + SPMD100), <0.0001 (c, hFUSR521H versus hFUSR521H + SPMD200), 0.0001 (d, non-tg versus hFUSR521H), NS (d, hFUSR521H versus hFUSR521H + SPMD100) and 0.017 (d, hFUSR521H versus hFUSR521H + SPMD200) using one-way ANOVA with Bonferroni’s multiple comparisons test. e, Representative immunofluorescence images of primary motor axons derived from nontransgenic and hFUSR521H animals in the axonal compartment, treated with or without spermidine 100 and 200 µM for 48 h and labeled with NFH (axons) antibody. Scale bar, 250 µm. f, Quantification of the axonal length from full axonal compartment images reported in e. Data are represented as mean ± s.e.m. (n = 3–5 biological replicates). P = 0.0001 (non-tg vehicle versus hFUSR521H vehicle), 0.016 (hFUSR521H vehicle versus hFUSR521H SPMD100) and 0.005 (hFUSR521H vehicle versus hFUSR521H SPMD200), using one-way ANOVA with Bonferroni’s multiple comparisons test. g,h, Eclosion assay in Drosophila melanogaster model of ALS induced by motor neuron-specific expression of WT and mutant FUS (g) and TDP-43 (h). Flies were fed with spermidine (a dose response of 0–5 mM). Data are represented as mean ± s.e.m. (n = 15–16 crosses/condition from three independent experiments). P = 0.001 (FUSWT, 0 versus 0.1), 0.0006 (FUSWT, 0 versus 0.5), 0.0002 (FUSWT, 0 versus 1), 0.0005 (FUSWT, 0 versus 2.5), 0.0006 (FUSWT, 0 versus 5), <0.0001 (FUSR521G, 0 versus 0.1, 0.5, 1, 2.5, 5), 0.001 (TDP-43WT, 0 versus 0.1), <0.0001 (TDP-43WT, 0 versus 0.5, 1, 2.5), 0.003 (TDP-43WT, 0 versus 5), 0.0001 (TDP-43Q331K, 0 versus 0.1) and <0.0001 (TDP-43Q331K, 0 versus 0.5, 1, 2.5, 5) using one-way ANOVA with Holm–Šídák’s multiple comparisons test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
To determine whether spermidine also improves motor axonal health, we measured axonal length and branching in the axonal compartment of cultured motor neurons (Fig. 6e,f and Extended Data Fig. 7g). Mutant FUS motor axons show reduced axonal length compared to nontransgenic axons (Fig. 6f), although the number of branches being unchanged (Extended Data Fig. 7g). Interestingly, spermidine treatment ameliorated the axonal length deficits by nearly threefold.
Taken together, these results demonstrate that axon-specific treatment with spermidine increases hypusinated Eif5a and rescues the mutant FUS-dependent translation decrease and alleviates axonal defects caused by mutant FUS.
Spermidine treatment ameliorates eclosion phenotypes in FUS and TDP-43 ALS fly models
To investigate whether spermidine also improves ALS-linked mutant-dependent defects in vivo, we dosed ALS Drosophila, whose motor neuron-specific overexpression of wild-type (WT) or mutant (R521G) human FUS leads to a severe eclosion phenotype47,48, with only 12.5% and 8.0% flies eclosing after 14 days, respectively (Fig. 6g). A dose response of spermidine was first evaluated in a control fly line that ecloses normally, revealing that spermidine is well-tolerated (Extended Data Fig. 7h). WT and mutant FUS flies were fed increasing doses of spermidine and a significant amelioration of the eclosion phenotype was observed (Fig. 6g).
Finally, to evaluate whether spermidine rescues exclusively FUS-dependent deficits or whether its potential benefit could be more widely applicable to other ALS forms, we also dosed flies that overexpress specifically in motor neurons ALS-causing RNA-binding protein human TDP-43 (either WT or mutant Q331K)49, whose aggregation is a major feature of nearly all ALS and half of FTLD cases50. Similar to FUS flies, a severe eclosion phenotype is also observed, especially in the mutant TDP-43 line with only 0.94% flies eclosing (Fig. 6h). Notably, a dose-dependent improved phenotype is observed in the spermidine-treated animals. Altogether, spermidine treatment represents a potential therapeutic strategy to improve ALS deficits induced by mutant FUS and TDP-43.
Discussion
Leveraging spatial transcriptomics technologies, we performed immuno-driven spatial profiling of specific cell populations in mouse spinal cords, integrating transcriptomic profiles of motor and nonmotor neurons within ventral and intermediate/dorsal horn areas, respectively, with snRNA-seq available datasets27,28,29,30. Additionally, to ensure the spatial specificity of the identified transcriptomic signatures, we applied unbiased spatial transcriptomic profiling of the mouse spinal cord, improving the spatial transcriptomic resolution already available on healthy and ALS spinal cords51,52, and validating the expression of poorly characterized genes in motor neurons.
Using whole-transcriptome profiling technology, we explored the subcellular spatial transcriptome of mature axons and identified distinct transcriptomic signatures in both motor and nonmotor axons. We discovered an enrichment of a substantial subset of specific transcripts that had not been previously identified in adult sciatic nerve axons. The variety of the identified axonal transcripts further supports local translation in mature axons as a physiological mechanism1. Moreover, this suggests specialized functions in distinct axonal subtypes. Art3, which was found to be enriched in nonmotor axons, is an arginine-specific ADP-ribosyltransferase previously detected in rat cortical axons after traumatic brain injury53, but its specific axonal role is not yet fully characterized. It would be of interest to assess whether Art3 can ADP-ribosylate axonal ribosomal proteins to modulate translation levels, as demonstrated in cancer54. Lao1 is a secreted L-amino acid oxidase which catabolizes L-phenylalanine, L-tyrosine, L-tryptophan and moderately L-arginine, with a recognized important role in immunomodulation55. Interestingly, a secreted protein similar to Lao1 (interleukin-4-induced 1 (Il4i1)) was found to facilitate myelin repair, effectively mitigating axonal injury and remyelination impairments in a model of multiple sclerosis56. Moreover, the Il4i1 transcript was detected in the axoplasm of rat ventral roots57, which is in line with our data. Interestingly, the presence of Lao1 in mature axons further supports the need for its local translation to maintain axons.
Mutant FUS causes the dysregulation of mRNAs encoding for ribosomal proteins and translation factors in the axons, which is reflected by their protein levels (Fig. 3). This is an expected outcome in a system where local translation is impaired8. Indeed, locally synthesized ribosomal proteins are incorporated into pre-existing axonal ribosomes, providing evidence that discards the role of nucleoli as the unique compartment for ribosome assembly and further supports the idea of plasticity of local translation35. We anticipate that translation factors, such as Eif5a, Eif4g3 and Eif4a1, are also locally translated in axons and can form active translation hubs in response to local cues.
Recognizing that Dohh mRNA is directly bound by the FUS protein58, it is possible that the mutant FUS-dependent reduction in Dohh mRNA levels observed in mutant FUS axons is also due to reduced stability or impaired transport of Dohh mRNAs. Indeed, mutations in FUS have been shown to compromise both RNA stability of specific targets16 and axonal transport of organelles59. It would be mechanistically relevant to test whether the interaction between Dohh transcript and FUS is thus modified by ALS-linked mutations. Nonetheless, our findings suggest that defects in local translation may not be uniquely caused by a direct action of Dohh, given that Dohh silencing did not further exacerbate these deficits. In contrast, we demonstrate that Dohh protein is an important player in axonal translation (Fig. 5j–m), further strengthening its involvement in the spermidine-dependent rescue of the mutant FUS-induced local translation deficits. Additionally, mutant FUS might perturb directly the spermidine pathway, altering the local translation/availability of additional enzymes involved in its production. Both mechanisms are nonmutually exclusive, and further research is therefore needed to fully understand how ALS-causing FUS mutations alter hypusine/Eif5a in axons.
To reverse the effect of mutant FUS on Eif5a specifically in axons, we applied spermidine compartment-specific treatment (Fig. 6) to serve as a substrate for the incorporation of hypusine in Eif5a. Interestingly, modulation of the polyamine pathway, including through spermidine boosting, has been reported to be beneficial not only during aging60 but also in neurodegenerative disorders, including Huntington’s disease61 and Parkinson’s disease (PD)62, and other genetic forms of ALS63,64,65,66. Hence, muscles of SOD1G93A mice display polyamine defects67.
Spermidine-mediated effects have not only been linked to Eif5a hypusination60,68 but also to autophagy activation in aging models60, TDP-43 overexpressing mice65 and models of PD69, possibly by acting as a mammalian target of rapamycin (mTOR)-independent autophagy enhancer70. In light of these previous reports and considering that FUS neurotoxicity has been associated with compromised autophagy71, this mechanism cannot be excluded as a possible synergistic effect in the spermidine-mediated amelioration of axonal defects. Our in vivo findings showing spermidine-mediated reduced toxicity in WT/mutant FUS-, as well as TDP-43-overexpressing fly models, support the rationale to test this approach in more complex in vivo systems for potential future therapy development.
Eif5a is critically involved in the release of ribosome stalling events at very-difficult-to-translate polyproline tracts72. Moreover, FUS was already shown to bind to stalled polyribosomes during translational repression by mTOR inhibition, and FUS variants bearing ALS-linked mutations R521G and P525L bind more abundantly to polyribosomes, probably increasing the affinity of this RNA-binding protein to target mRNAs73. Therefore, it is likely that disease-causing mutations in FUS lead to ribosome stalling, as recently demonstrated for mutant huntingtin61 and that the hypusinated Eif5a-dependent increase in local translation mediated by axon-specific treatment of spermidine resolves pausing of ribosomes. It would be of interest in the future to determine whether there are increased ribosome stalling events induced by FUS mutation, specifically in motor axons, and whether spermidine treatment alleviates the ribosomal stress.
In sum, our application of spatial transcriptomics in a compartment-specific manner identifies targetable pathways in neurodegenerative disorders, pointing to a possible compartment-specific treatment that can be achieved through intervention at the periphery.
Methods
Mouse strains
The mouse strains used in this study are reported in ref. 8. All mice used in this report were maintained on a pure C57BL/6 background. All experimental procedures were conducted according to the KU Leuven ethical guidelines and approved by the KU Leuven Ethical Committee for Animal Experimentation. Mice were maintained in standard housing conditions, on a diurnal 12-h light/12-h dark cycle and with continuous access to food and water. We used female mice for spatial transcriptomics experiments. For all the other mouse experiments, gender was balanced. Adult 12-month-old mice were analyzed in all the experiments.
Eclosion phenotype with spermidine
Spermidine trihydrochloride (0–10 mM; Thermo Fisher Scientific, 10177463) was dissolved in milliQ H2O and added to standard food at the indicated concentrations. For the 0 condition, H2O was added. For FUS, w-;;UAS-FUS (WT) and w-;;UAS-FUS (R521G) males were crossed to D42–Gal4 virgins. For TDP-43, w-;;UAS-TDP-43 (WT) and w-;;UAS-TDP-43 (Q331K) males were crossed to D42–Gal4 virgins. As a control, w1118 males were crossed to D42–Gal4 virgins. Females laid the eggs on drug-containing food for 48 h. Crosses were left to develop at 25 °C, and eclosion was scored 14 days after setup.
Primary neuronal cultures
Primary cortical neurons and motor neurons were prepared from embryonic day 15–16 (E15–E16) and 12–13 (E12–13), respectively, originating from positive and negative hFUSR521H littermates. Cortical and spinal cord tissues were treated with 0.25% trypsin + EDTA for 15 min at 37 °C. Trypsin activity was inhibited using Dulbecco’s modified Eagle’s medium (DMEM)/F-12 media supplemented with 10% of FBS and 1% of penicillin–streptomycin and tissues were then mechanically disaggregated using 1000 μl pipette tips. A 4% bovine serum albumin dissolved in 1× PBS solution was further added to the cell suspension as a cushion and centrifuged for 10 min at 300g. The cell pellet was resuspended in neurobasal media supplemented with 2% B27, 200 mM of L-glutamine and 1% of penicillin–streptomycin. Cells were seeded in culture plates, microfluidic devices or MEA precoated with poly-L-lysine or poly-D-lysine.
N2A cell cultures
N2A cells purchased from American Type Culture Collection were maintained in DMEM 1× supplemented with 10% FBS, 1% penicillin–streptomycin and 1% L-glutamine. Cells were seeded and further processed for western blot analysis.
Spatial transcriptomics of spinal cord neurons and sciatic nerve axons by NanoString GeoMx profiling
Spatial transcriptomics was performed on four consecutive sections of one nontransgenic and one mFUSKO/hFUSR521H mouse. In total, 5 μm-paraffin-embedded sections of spinal cord and sciatic nerve tissues were layered on normal glass microscope slides and were frozen on dry ice. Slides were analyzed on the NanoString GeoMx digital spatial profiler platform using the whole-mouse transcriptome atlas panel composed of 19,960 gene probes (NanoString). ROIs were drawn on spinal cord ventral horns and full sciatic nerve sections and further segmented into ChAT-positive and ChAT-negative areas of interest. UV-photocleavable barcodes attached to probes were collected from selected ROIs using UV light directed to the sample. Processing of the samples was performed according to available protocols from the manufacturer (NanoString). Raw Nanostring.dcc and.pkc counts and configuration files were read using the R packages NanoStringNCTools (v1.6.1)74, GeomxTools (v3.2.0)75 and GeoMxWorkflows (v1.4.0)76 within an R v4.2.0 environment to produce a non-normalized count matrix with target names, which were converted to GENEIDs. The following targets were not included in the annotation file and were excluded from the final count matrix: Gm10406, LOC118568634, D830030K20Rik and NegProbe-WTX. DE analysis was carried out using the DESeq2 package (v1.42.0)77 in R (v4.3.2). The filtered, collated count matrix and experimental metadata were imported into R, and data normalization and variance stabilization were performed on the gene counts using the DESeq2 functions. DE analysis was conducted by fitting a negative binomial generalized linear model to the normalized counts. This linear model integrated the following parameters ‘~ segment + class’ for global comparisons and a reduced model ‘~ class’ for within-class comparisons. Genes with a false discovery rate Benjamin–Hochberg-adjusted P value below the 0.05 significance threshold were considered differentially expressed. Lists of DE genes were produced for the following comparisons described in the Supplementary Information. GO of DE genes was computed with Database for Annotation, Visualization and Integrated Discovery (DAVID)78.
Comparison of the NanoString GeoMx dataset with publicly available snRNA-seq datasets
We compared our dataset obtained using NanoString GeoMx with publicly available spinal cord snRNA-seq datasets (accessions GSE161621 (ref. 27), GSE167597 (ref.28), GSE228778 (ref. 29) and GSE190442 (ref. 30)). The processed count matrices and author nuclei annotations, or Seurat RDS objects (provided by the authors of these studies when this information was not already available in the original publication), for these datasets were downloaded from GEO and analyzed using the R Seurat package (v5.0.1)79,80,81. Where necessary, we removed all nuclei annotated as non-neuronal nuclei before the DEG comparison. The author’s annotations labeled as ‘Skeletal MNs’, ‘Motoneurons’, ‘Skeletal Motor Neurons’ and ‘Motoneurons’ were used to define the ChAT-positive cluster for the comparison in GSE161621, GSE228778, GSE167597 and GSE190442, respectively. The rest of the nuclei were assigned as ChAT-negative clusters in all datasets. DEG analysis was performed between the annotated ChAT-positive and ChAT-negative groups described above using the Seurat FindAllMarkers function, applying the Wilcoxon rank-sum test, generating lists of DE genes.
Lists (filtered on adjusted P < 0.05) of DE genes from the NanoString GeoMx analysis were correlated with the DE genes from the analysis of the ChAT-positive and ChAT-negative nuclei in the publicly available datasets, based on the log2(FC) of genes present in both datasets being correlated. For datasets containing human neuronal nuclei GSE190442 and GSE228778, human gene names were annotated with the Ensembl IDs and mapped to mouse gene analogs using the getLDS function in the biomaRt R package (v2.58.0)82, and duplicate-mapped mouse Ensembl IDs were removed to compare the genes across datasets. Spearman correlation was performed using the ggpubr R package (v0.6.0) and visualized using ggplot2. Inclusion of DE genes in each list was performed and visualized using the VennDiagram R package (v1.7.3).
Unbiased spatial transcriptomics analysis of spinal cords using 10x Visium profiling
Samples were processed for spatial transcriptomics using the 10x Genomics Visium platform. Spinal cords were dissected, washed with 1× Dulbecco’s phosphate buffered saline, embedded in 5% carboxymethylcellulose and snap-frozen in chilled isopentane. Tissue blocks were stored at −70 °C in a sealed container. Both the tissue block and the proprietary Visium Spatial Gene Expression Slide (10x Genomics, PN-2000233) were equilibrated inside the cryostat for at least 30 min at −20 °C before sectioning. Sections were cut at a thickness of 10 μm and immediately placed onto the slide. Slides containing sections were stored at −70 °C for a maximum of 24 h before use. Two sections per animal per genotype were used, resulting in a total of eight sections.
Fixation, staining, imaging and construction of cDNA libraries were done according to the manufacturer’s instructions (Visium Spatial Gene Expression User Guide_Rev D; 10x Genomics, CG000239) using the Visium Spatial Gene Expression Slide & Reagent Kit (10x Genomics, PN-1000187). Briefly, sections were fixed in chilled methanol for 30 min at −20 °C and stained with haematoxylin and eosin (H&E). Imaging was performed on a Nikon-Marzhauser Slide Express 2 whole-slide scanner at ×10 magnification. After imaging, sections were permeabilized at 37 °C for 18 min. Permeabilization time was determined using the Visium Spatial Tissue Optimization Slide & Reagent Kit (10x Genomics, PN-1000193) following the Visium Spatial Tissue Optimization User Guide_RevA (10x Genomics, CG000238). After permeabilization, the on-slide reverse transcription reaction was performed at 53 °C for 45 min. Second-strand synthesis was subsequently performed on-slide for 15 min at 65 °C. All on-slide reactions were performed in a thermocycler with a metal slide adapter plate (10x Genomics). Following second-strand synthesis, samples were transferred to tubes for cDNA amplification, clean-up and library preparation. Library quality was assessed using an Agilent Technologies Bioanalyzer High Sensitivity kit (Agilent Technologies, 5067-4626).
Visium libraries were sequenced on Illumina NextSeq 2000. The sequencing depth was chosen by determining the number of 55 μm spots that were covered by tissue, and this was multiplied by 50,000 reads. Raw reads were mapped to the mouse genome reference (mm10-2020-A provided by 10x Genomics) with SpaceRanger (v1.1.0; 10x Genomics) to generate spatial gene expression matrices. SpaceRanger output files were then further processed with SCANPY (1.8.1)83. H&E images were manually inspected to identify spots overlapping spinal cord tissue and to remove spots overlapping with artifacts due to the embedding reagent. Genes that were expressed in less than three spots per capture area were filtered out. Spots were removed if they expressed fewer than 200 genes, more than 4,000 genes or if more than 40% of expressed genes were mitochondrial genes. Gene expression was then normalized to match a total sum of 104 counts per spot and logarithmized.
Motor and nonmotor neuron signature enrichment analysis of 10x Visium spinal cord data
Gene set enrichment analysis was performed on 10x Visium spinal cord data to localize ChAT-positive and ChAT-negative areas in tissues. Specifically, ChAT-positive and ChAT-negative marker genes identified from DE analysis of NanoString data were selected based on adjusted P value < 0.05 and log2(FC) > 1. Then, for each of the two sets of marker genes, we computed the normalized enrichment scores (NES) using the GSEApy Python package on scaled log-normalized Visium data subsetted to highly variable genes84 (with ssgsea function and parameters sample_norm_method = ‘rank’, min_size=5). Finally, Visium spots with NES > 0.1 were marked, respectively, as ChAT-positive and ChAT-negative.
Multiplexed single-molecule RNA fluorescence in situ hybridization
Tissue sections
Freshly dissected spinal cord and sciatic nerve samples from four controls and three mFUSKO/hFUSR521H mice were frozen in 2-methylbutane, sectioned with a cryostat and 10-µm-thick sections were placed within the capture areas of cold equilibrated Resolve Biosciences slides. Tissue sections were thawed and fixed with 4% vol/vol formaldehyde (Sigma-Aldrich, F8775) in 1× PBS for 5 min at 4 °C. After fixation, sections were washed thrice in 1× PBS for 1 min, followed by 1 min washes in 70% ethanol, isopropanol, 95% ethanol and 70% ethanol at room temperature. Fixed samples were used for Molecular Cartography (100-plex combinatorial smFISH) according to the manufacturer’s instructions (protocol 1.4; available for registered users), starting with the aspiration of ethanol and the addition of buffer DST1, followed by tissue priming and hybridization. Briefly, tissues were primed for 30 min at 37 °C, followed by 24 h hybridization of all probes specific for the target genes (see next section and Table 1 for probe design details and target list). After the hybridization step, samples were washed to remove excess probes and fluorescently tagged in a two-step color development process. ROIs were imaged as described below, and fluorescence signals were removed during decolorization. Color development, imaging and decolorization were repeated for multiple cycles to build a unique combinatorial code for every target gene that was derived from raw images as described below.
Probe design
The probes for the 30 genes were designed using Resolve’s proprietary design algorithm. Briefly, the probe design was performed at the gene level. For every targeted gene, all full-length protein-coding transcript sequences from the ENSEMBL database were used as design targets if the isoform had the GENCODE annotation tag ‘basic’85,86. To speed up the process, the calculation of computationally expensive parts, especially the off-target searches, was not performed randomly. Instead, the selection of probe sequences was limited to those with high success rates. To filter highly repetitive regions, the abundance of k-mers was obtained from the background transcriptome using jellyfish87. Every target sequence was scanned once for all k-mers, and those regions with rare k-mers were preferred as seeds for full probe design. A probe candidate was generated by extending a seed sequence until a certain target stability was reached. A set of simple proprietary rules was applied to discard sequences that were found experimentally to cause problems. After these fast screens, every kept probe candidate was mapped to the background transcriptome using ThermonucleotideBLAST88, and probes with stable off-target hits were discarded. Specific probes were then scored based on the number of on-target matches (isoforms), which were weighted by their associated Annotation of PRincipal and Alternative splice ISoforms (APPRIS) level89, favoring principal isoforms over others. A bonus was added if the binding site was inside the protein-coding region. From the pool of accepted probes, the final set was composed by greedily picking the highest-scoring probes. Table 1 highlights the gene names and catalog numbers for the specific probes designed by Resolve BioSciences.
Imaging
Samples were imaged on a Zeiss Celldiscoverer 7, using the ×50 Plan Apochromat water immersion objective with a numerical aperture (NA) of 1.2 and the ×0.5 magnification changer, resulting in a ×25 final magnification. Standard CD7 LED excitation light source, filters and dichroic mirrors were used together with customized emission filters optimized for detecting specific signals. Excitation time per image was 1,000 ms for each channel (DAPI was 20 ms). A z-stack was taken at each region with a distance per z-slice according to the Nyquist–Shannon sampling theorem. The custom CD7 CMOS camera (Zeiss Axiocam Mono 712, 3.45 µm pixel size) was used. For each region, a z-stack per fluorescent color (two colors) was imaged per imaging round. A total of eight imaging rounds were done for each position, resulting in 16 z-stacks per region. The completely automated imaging process per round (including water immersion generation and precise relocation of regions to image in all three dimensions) was realized by a custom Python script using the scripting API of the Zeiss ZEN software (Open Application Development).
Spot segmentation
The algorithms for spot segmentation were written in Java and are based on the ImageJ library functionalities. Only the iterative closest point algorithm is written in C++ based on the libpointmatcher library (https://github.com/ethz-asl/libpointmatcher).
Preprocessing
As a first step, all images were corrected for background fluorescence. A target value for the allowed number of maxima was determined based on the area of the slice in µm² multiplied by the factor 0.5. This factor was empirically optimized. The brightest maxima per plane were determined based on an empirically optimized threshold. The number and location of the respective maxima were stored. This procedure was done for every image slice independently. Maxima that did not have a neighboring maximum in an adjacent slice (called a z-group) were excluded. The resulting maxima list was further filtered in an iterative loop by adjusting the allowed thresholds for (Babs–Bback) and (Bperi–Bback) to reach a feature target value (Babs, absolute brightness; Bback, local background; Bperi, background of periphery within 1 pixel). The target values for this feature were based on the volume of the three-dimensional (3D) image. Only maxima still in a z-group of at least 2 after filtering were passing the filter step. Each z-group was counted as one hit. The members of the z-groups with the highest absolute brightness were used as features and written to a file. They resemble a 3D point cloud.
Final signal segmentation and decoding
To align the raw data images from different imaging rounds, the images had to be corrected. To do so, the extracted feature point clouds were used to find the transformation matrices. For this purpose, an iterative closest point cloud algorithm was used to minimize the error between two point clouds. The point clouds of each round were aligned to the point cloud of round one (reference point cloud). The corresponding point clouds were stored for downstream processes. Based upon the transformation matrices, the corresponding images were processed by a rigid transformation using trilinear interpolation. The aligned images were used to create a profile for each pixel consisting of 16 values (16 images from two color channels in eight imaging rounds). The pixel profiles were filtered for variance from zero, normalized by the total brightness of all pixels in the profile. Matched pixel profiles with the highest score were assigned an ID to the pixel. Pixels with neighbors having the same ID were grouped. The pixel groups were filtered by group size, number of direct adjacent pixels in the group and number of dimensions with a size of two pixels. The local 3D maxima of the groups were determined as potential final transcript locations. Maxima were filtered by the number of maxima in the raw data images where a maximum was expected. Remaining maxima were further evaluated by the fit to the corresponding code. The remaining maxima were written to the results file and considered to resemble transcripts of the corresponding gene. The ratio of signals matching codes used in the experiment to those not used was used as an estimation for specificity (false positives).
Downstream analysis
Transcript signals were inspected using the ImageJ Polylux tool plugin from Resolve BioSciences. For the quantification of the transcripts inside axons, the NFH signal was segmented using QuPath’s standard watershed-based segmentation90. Objects were exported to segmentation masks using a custom macro. These masks were then used to split the gene expression into NFH-positive and NFH-negative count matrices using an in-house script. For the quantification of the transcripts inside motor neurons, areas were manually segmented based on the expression of Chat and Chodl motor neuron marker genes. Segmentation masks were used to extract motor neuron-specific gene expression and respective areas using an in-house script, based on the skimage.measure.regionprops function (scikit-image = 0.20.0 (ref. 91)).
Immunofluorescence
Mice were perfused intracardially and fixed with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.2). The entire spinal cord and sciatic nerves were dissected, postfixed for 3 h in the same fixative and transferred to a 30% sucrose phosphate buffer for at least 2 days. The lumbar spinal cord and proximal sciatic nerve region were embedded in optimal cutting temperature compound (Sakura) and snap-frozen in isopentane (2-methylbutane) cooled at −40 °C on dry ice. Floating lumbar spinal cord cryosections (30 μm) or mounted sciatic nerve sections (10 μm) were incubated in blocking solution (1× PBS, 0.5% Tween-20 and 1.5% bovine serum albumin) for 1 h at room temperature and then in 1× PBS and 0.3% Triton X-100 overnight (ON) at room temperature with the primary antibodies (listed in the antibodies section). Primary antibodies were washed thrice with 1× PBS and then detected using AlexaFluor-488, Cy3 or AlexaFluor647 (1:500) coupled secondary antibodies (Jackson ImmunoResearch). The sections were washed thrice with 1× PBS, incubated with DAPI for 10 min, further washed and mounted in Prolong Gold mounting medium (Invitrogen). The same protocol was applied to fixed cells grown on coverslips or in microfluidic devices. Analysis was performed on a Nikon C2 laser scanning confocal microscope. Fluorescence intensity from unsaturated images captured with identical confocal settings was quantified using ImageJ (National Institutes of Health (NIH)). ROIs based on ChAT immunofluorescence signal were thresholded and chosen for quantification. Images from microfluidic devices were analyzed using a GA3 analysis script in NIS-Elements (6.02.01 and 6.10.01; Nikon Instruments Europe B.V.). For Fig. 6e,f, the images were semi-automatically segmented (step 1) and consequently skeletonized (step 2). From these binary layers, the neuron length, area and branching points were analysed. For Fig. 6b–d, binary layers were created, and the intensities of different markers at different distances from the start of the grooves were measured. For this, we used spaced lines 6.6 µm wide and 6.6 µm apart.
Compartmentalized microfluidic devices
The microfluidic devices were prepared as previously described92. Master molds were fabricated by photolithography by the Bioengineering Department of the University of KU Leuven (Belgium). Devices were molded by the addition of a liquid silicone base (Sylgard 182) on the soft-lithography-based mold and cured ON at 55 °C (Ellsworth Adhesives) as previously described93. After being cured and cut, the microfluidic devices were washed in 70% ethanol, UV-irradiated for 1 h and dried under the hood. The sterilized devices were mounted onto glass coverslips. The devices were coated with 1× poly-D-lysine for at least 2 h at 37 °C and washed sequentially in water and Neurobasal Complete Medium before plating the cells.
Puromycin incorporation assay
Primary cortical and motor neurons after 5 or 8 days in culture (day in vitro (DIV) 5−8) were incubated for 10 min with puromycin at 1 μg ml−1. After washing with 1× PBS, cells were fixed with 4% paraformaldehyde, blocked and stained with the anti-puromycin antibody (1:100) ON at 4 °C. Fluorescence was visualized using a Nikon C2 laser scanning confocal microscope.
MEA
Primary cortical neurons from nontransgenic and hFUSR521H mouse embryos were seeded on poly-L-lysine-coated MaxWell two six-well plate (MaxWell Biosystems), each containing 26,400 electrodes. Before each recording, neurons were incubated with BrainPhys media (STEMCELL Technologies) to increase neuronal excitability. Recordings of spontaneous activities during a 5-min period were performed. Analysis of all MEA activity and data processing (activity scan and network activity) was performed using MaxTwo software. Biological triplicates for each sample were included.
Immunoblotting
N2A cells were homogenized in cold radioimmunoprecipitation assay buffer supplemented with protease and phosphatase inhibitors (Thermo Fisher Scientific). The lysates were centrifuged at 4 °C for 15 min at 13,000g. The supernatants were quantified by bicinchoninic acid kit (Thermo Fisher Scientific), and equal protein amounts were separated on sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred to PVDF membranes and probed with the indicated antibodies, followed by horseradish peroxidase-conjugated secondary antibodies (Jackson ImmunoResearch). Immunoreactivity was detected with Pico or Femto enhanced chemiluminescence reagents (Pierce), and quantifications were performed using ImageJ 1.51j8 software (NIH).
Statistics and reproducibility
All data are presented as means ± s.e.m., unless otherwise stated. The number of experimental replicates and tests used is reported in the legends of Figs. 1–6 for each experiment. The datasets met the assumptions of the statistical tests used. Random mice of the same age and belonging to the indicated genotypes were selected for the described experiments. All mice were age-matched and housed under identical environmental and handling conditions to minimize potential covariates. Blinding was not performed in the present study because all the procedures, data acquisition and analysis involved objective measurements that are not influenced by operator bias. For all tests, the significance threshold was set at P < 0.05. Data were analyzed, and graphs were generated using GraphPad Analysis software. Spatial transcriptomics experiments were performed once. Data not passing the quality control step were excluded from the analysis. No statistical methods were used to predetermine sample sizes, but our sample sizes are similar to those reported in previous publications8,94.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Gene Expression Omnibus (GEO) accession for NanoString GeoMx dataset: GSE269707. GEO accession for 10x Genomics Visium dataset: GSE269377. Resolve Biosciences data are deposited in the Zenodo repository accessible through the following DOI: 10.5281/zenodo.15851243. Source data are provided with this paper.
Code availability
The generated code is available at the following link: https://github.com/sifrimlab/Piol_motor_neuron.
References
Piol, D., Robberechts, T. & Da Cruz, S. Lost in local translation: TDP-43 and FUS in axonal/neuromuscular junction maintenance and dysregulation in amyotrophic lateral sclerosis. Neuron 111, 1355–1380 (2023).
von Kügelgen, N. & Chekulaeva, M. Conservation of a core neurite transcriptome across neuronal types and species. Wiley Interdiscip. Rev. RNA 11, e1590 (2020).
Kye, M. J. et al. SMN regulates axonal local translation via miR-183/mTOR pathway. Hum. Mol. Genet. 23, 6318–6331 (2014).
Saal, L., Briese, M., Kneitz, S., Glinka, M. & Sendtner, M. Subcellular transcriptome alterations in a cell culture model of spinal muscular atrophy point to widespread defects in axonal growth and presynaptic differentiation. RNA 20, 1789–1802 (2014).
Zuko, A. et al. tRNA overexpression rescues peripheral neuropathy caused by mutations in tRNA synthetase. Science 373, 1161–1166 (2021).
Spaulding, E. L. et al. The integrated stress response contributes to tRNA synthetase-associated peripheral neuropathy. Science 373, 1156–1161 (2021).
Mendonsa, S., von Kuegelgen, N., Bujanic, L. & Chekulaeva, M. Charcot–Marie–Tooth mutation in glycyl-tRNA synthetase stalls ribosomes in a pre-accommodation state and activates integrated stress response. Nucleic Acids Res. 49, 10007–10017 (2021).
López-Erauskin, J. et al. ALS/FTD-linked mutation in FUS suppresses intra-axonal protein synthesis and drives disease without nuclear loss-of-function of FUS. Neuron 100, 816–830 (2018).
Ionescu, A., Altman, T. & Perlson, E. Looking for answers far away from the soma—the (un)known axonal functions of TDP-43, and their contribution to early NMJ disruption in ALS. Mol. Neurodegener. 18, 35 (2023).
Kwiatkowski, T. J. et al. Mutations in the FUS/TLS gene on chromosome 16 cause familial amyotrophic lateral sclerosis. Science 323, 1205–1208 (2009).
Vance, C. et al. Mutations in FUS, an RNA processing protein, cause familial amyotrophic lateral sclerosis type 6. Science 323, 1208–1211 (2009).
Broustal, O. et al. FUS mutations in frontotemporal lobar degeneration with amyotrophic lateral sclerosis. J. Alzheimers Dis. 22, 765–769 (2010).
Yan, J. et al. Frameshift and novel mutations in FUS in familial amyotrophic lateral sclerosis and ALS/dementia. Neurology 75, 807–814 (2010).
Mackenzie, I. R., Rademakers, R. & Neumann, M. TDP-43 and FUS in amyotrophic lateral sclerosis and frontotemporal dementia. Lancet Neurol. 9, 995–1007 (2010).
Moens, T. G. et al. Amyotrophic lateral sclerosis caused by FUS mutations: advances with broad implications. Lancet Neurol. 24, 166–178 (2025).
Birsa, N., Bentham, M. P. & Fratta, P. Cytoplasmic functions of TDP-43 and FUS and their role in ALS. Semin. Cell Dev. Biol. 99, 193–201 (2020).
Belly, A., Moreau-Gachelin, F., Sadoul, R. & Goldberg, Y. Delocalization of the multifunctional RNA splicing factor TLS/FUS in hippocampal neurones: exclusion from the nucleus and accumulation in dendritic granules and spine heads. Neurosci. Lett. 379, 152–157 (2005).
Yasuda, K. et al. The RNA-binding protein Fus directs translation of localized mRNAs in APC–RNP granules. J. Cell Biol. 203, 737–746 (2013).
Fujii, R. et al. The RNA binding protein TLS is translocated to dendritic spines by mGluR5 activation and regulates spine morphology. Curr. Biol. 15, 587–593 (2005).
Schoen, M. et al. Super-resolution microscopy reveals presynaptic localization of the ALS/FTD related protein FUS in hippocampal neurons. Front. Cell. Neurosci. 9, 496 (2016).
Aoki, N. et al. Localization of fused in sarcoma (FUS) protein to the post-synaptic density in the brain. Acta Neuropathol. 124, 383–394 (2012).
Picchiarelli, G. et al. FUS-mediated regulation of acetylcholine receptor transcription at neuromuscular junctions is compromised in amyotrophic lateral sclerosis. Nat. Neurosci. 22, 1793–1805 (2019).
So, E. et al. Mitochondrial abnormalities and disruption of the neuromuscular junction precede the clinical phenotype and motor neuron loss in hFUSWT transgenic mice. Hum. Mol. Genet. 27, 463–474 (2018).
Da Cruz, S. & Cleveland, D. W. Understanding the role of TDP-43 and FUS/TLS in ALS and beyond. Curr. Opin. Neurobiol. 21, 904–919 (2011).
Merritt, C. R. et al. Multiplex digital spatial profiling of proteins and RNA in fixed tissue. Nat. Biotechnol. 38, 586–599 (2020).
Krach, F. et al. Transcriptome–pathology correlation identifies interplay between TDP-43 and the expression of its kinase CK1E in sporadic ALS. Acta Neuropathol. 136, 405–423 (2018).
Blum, J. A. et al. Single-cell transcriptomic analysis of the adult mouse spinal cord reveals molecular diversity of autonomic and skeletal motor neurons. Nat. Neurosci. 24, 572–583 (2021).
Alkaslasi, M. R. et al. Single nucleus RNA-sequencing defines unexpected diversity of cholinergic neuron types in the adult mouse spinal cord. Nat. Commun. 12, 2471 (2021).
Gautier, O. et al. Challenges of profiling motor neuron transcriptomes from human spinal cord. Neuron 111, 3739–3741 (2023).
Yadav, A. et al. A cellular taxonomy of the adult human spinal cord. Neuron 111, 328–344 (2023).
Arvidsson, U., Riedl, M., Elde, R. & Meister, B. Vesicular acetylcholine transporter (VAChT) protein: a novel and unique marker for cholinergic neurons in the central and peripheral nervous systems. J. Comp. Neurol. 378, 454–467 (1997).
Kobayashi, Y. et al. Distribution of the high-affinity choline transporter in the human and macaque monkey spinal cord. Neurosci. Lett. 317, 25–28 (2002).
Enjin, A. et al. Identification of novel spinal cholinergic genetic subtypes disclose Chodl and Pitx2 as markers for fast motor neurons and partition cells. J. Comp. Neurol. 518, 2284–2304 (2010).
López-Erauskin, J. et al. Stathmin-2 loss leads to neurofilament-dependent axonal collapse driving motor and sensory denervation. Nat. Neurosci. 27, 34–47 (2024).
Shigeoka, T. et al. On-site ribosome remodeling by locally synthesized ribosomal proteins in axons. Cell Rep. 29, 3605–3619 (2019).
Cioni, J.-M. et al. Late endosomes act as mRNA translation platforms and sustain mitochondria in axons. Cell 176, 56–72 (2019).
Saini, P., Eyler, D. E., Green, R. & Dever, T. E. Hypusine-containing protein eIF5A promotes translation elongation. Nature 459, 118–121 (2009).
Schuller, A. P., Wu, C. C.-C., Dever, T. E., Buskirk, A. R. & Green, R. eIF5A functions globally in translation elongation and termination. Mol. Cell 66, 194–205 (2017).
Park, M. H., Nishimura, K., Zanelli, C. F. & Valentini, S. R. Functional significance of eIF5A and its hypusine modification in eukaryotes. Amino Acids 38, 491–500 (2010).
Park, M. H., Cooper, H. L. & Folk, J. E. Identification of hypusine, an unusual amino acid, in a protein from human lymphocytes and of spermidine as its biosynthetic precursor. Proc. Natl Acad. Sci. USA 78, 2869–2873 (1981).
Nishimura, K., Lee, S. B., Park, J. H. & Park, M. H. Essential role of eIF5A-1 and deoxyhypusine synthase in mouse embryonic development. Amino Acids 42, 703–710 (2012).
Sievert, H. et al. A novel mouse model for inhibition of DOHH-mediated hypusine modification reveals a crucial function in embryonic development, proliferation and oncogenic transformation. Dis. Model. Mech. 7, 963–976 (2014).
Kar, R. K. et al. Neuron-specific ablation of eIF5A or deoxyhypusine synthase leads to impairments in growth, viability, neurodevelopment, and cognitive functions in mice. J. Biol. Chem. 297, 101333 (2021).
Faundes, V. et al. Impaired eIF5A function causes a Mendelian disorder that is partially rescued in model systems by spermidine. Nat. Commun. 12, 833 (2021).
Park, M. H., Kar, R. K., Banka, S., Ziegler, A. & Chung, W. K. Post-translational formation of hypusine in eIF5A: implications in human neurodevelopment. Amino Acids 54, 485–499 (2022).
Cano, V. S. P. et al. Mutational analyses of human eIF5A-1—identification of amino acid residues critical for eIF5A activity and hypusine modification. FEBS J. 275, 44–58 (2008).
Bogaert, E. et al. Molecular dissection of FUS points at synergistic effect of low-complexity domains in toxicity. Cell Rep. 24, 529–537 (2018).
Tziortzouda, P. et al. PP2A and GSK3 act as modifiers of FUS-ALS by modulating mitochondrial transport. Acta Neuropathol. 147, 41 (2024).
Sreedharan, J., Neukomm, L. J., Brown, R. H. & Freeman, M. R. Age-dependent TDP-43-mediated motor neuron degeneration requires GSK3, hat-trick, and xmas-2. Curr. Biol. 25, 2130–2136 (2015).
Suk, T. R. & Rousseaux, M. W. C. The role of TDP-43 mislocalization in amyotrophic lateral sclerosis. Mol. Neurodegener. 15, 45 (2020).
Ståhl, P. L. et al. Visualization and analysis of gene expression in tissue sections by spatial transcriptomics. Science 353, 78–82 (2016).
Maniatis, S. et al. Spatiotemporal dynamics of molecular pathology in amyotrophic lateral sclerosis. Science 364, 89–93 (2019).
Shi, W. et al. The expression pattern of ADP-ribosyltransferase 3 in rat traumatic brain injury. J. Mol. Histol. 43, 37–47 (2012).
Challa, S. et al. Ribosome ADP-ribosylation inhibits translation and maintains proteostasis in cancers. Cell 184, 4531–4546 (2021).
Boulland, M.-L. et al. Human IL4I1 is a secreted L-phenylalanine oxidase expressed by mature dendritic cells that inhibits T-lymphocyte proliferation. Blood 110, 220–227 (2007).
Psachoulia, K. et al. IL4I1 augments CNS remyelination and axonal protection by modulating T cell driven inflammation. Brain 139, 3121–3136 (2016).
Farias, J., Holt, C. E., Sotelo, J. R. & Sotelo-Silveira, J. R. Axon microdissection and transcriptome profiling reveals the in vivo RNA content of fully differentiated myelinated motor axons. RNA 26, 595–612 (2020).
Lagier-Tourenne, C. et al. Divergent roles of ALS-linked proteins FUS/TLS and TDP-43 intersect in processing long pre-mRNAs. Nat. Neurosci. 15, 1488–1497 (2012).
Guo, W. et al. HDAC6 inhibition reverses axonal transport defects in motor neurons derived from FUS-ALS patients. Nat. Commun. 8, 861 (2017).
Hofer, S. J. et al. Spermidine is essential for fasting-mediated autophagy and longevity. Nat. Cell Biol. 26, 1571–1584 (2024).
Aviner, R. et al. Polyglutamine-mediated ribotoxicity disrupts proteostasis and stress responses in Huntington’s disease. Nat. Cell Biol. 26, 892–902 (2024).
Vrijsen, S., Houdou, M., Cascalho, A., Eggermont, J. & Vangheluwe, P. Polyamines in Parkinson’s disease: balancing between neurotoxicity and neuroprotection. Annu. Rev. Biochem. 92, 435–464 (2023).
Lee, J., Ryu, H. & Kowall, N. W. Motor neuronal protection by L-arginine prolongs survival of mutant SOD1 (G93A) ALS mice. Biochem. Biophys. Res. Commun. 384, 524–529 (2009).
Choi, S.-H. et al. Modulation of histone H3K4 dimethylation by spermidine ameliorates motor neuron survival and neuropathology in a mouse model of ALS. J. Biomed. Sci. 29, 106 (2022).
Wang, I.-F. et al. Autophagy activators rescue and alleviate pathogenesis of a mouse model with proteinopathies of the TAR DNA-binding protein 43. Proc. Natl Acad. Sci. USA 109, 15024–15029 (2012).
Smeltzer, S. et al. Hypusination of Eif5a regulates cytoplasmic TDP-43 aggregation and accumulation in a stress-induced cellular model. Biochim. Biophys. Acta, Mol. Basis Dis. 1867, 165939 (2021).
Ruggieri, V. et al. Polyamine metabolism dysregulation contributes to muscle fiber vulnerability in ALS. Cell Rep. 44, 115123 (2025).
Dever, T. E. & Ivanov, I. P. Roles of polyamines in translation. J. Biol. Chem. 293, 18719–18729 (2018).
Büttner, S. et al. Spermidine protects against α-synuclein neurotoxicity. Cell Cycle 13, 3903–3908 (2014).
Madeo, F., Eisenberg, T., Pietrocola, F. & Kroemer, G. Spermidine in health and disease. Science 359, eaan2788 (2018).
Ling, S.-C. et al. Overriding FUS autoregulation in mice triggers gain-of-toxic dysfunctions in RNA metabolism and autophagy-lysosome axis. eLife 8, e40811 (2019).
Gutierrez, E. et al. eIF5A promotes translation of polyproline motifs. Mol. Cell 51, 35–45 (2013).
Sévigny, M. et al. FUS contributes to mTOR-dependent inhibition of translation. J. Biol. Chem. 295, 18459–18473 (2020).
Aboyoun, P., Ortogero, N. & Yang, Z. NanoStringNCTools: NanoString nCounter Tools. Bioconductor version: release (3.18) https://doi.org/10.18129/B9.bioc.NanoStringNCTools (2023).
Ortogero, N., Yang, Z., Vitancol, R., Griswold, M. & Henderson, D. GeomxTools: NanoString GeoMx Tools. Bioconductor version: release (3.18) https://doi.org/10.18129/B9.bioc.GeomxTools (2023).
Griswold, M. et al. GeoMxWorkflows: GeoMx Digital Spatial Profiler (DSP) data analysis workflows. Bioconductor version: release (3.21). bioconductor.org/packages/GeoMxWorkflows/ (2025).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Huang, D. W., Sherman, B. T. & Lempicki, R. A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44–57 (2009).
Hao, Y. et al. Dictionary learning for integrative, multimodal and scalable single-cell analysis. Nat. Biotechnol. 42, 293–304 (2024).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587 (2021).
Stuart, T. et al. Comprehensive integration of single-cell data. Cell 177, 1888–1902 (2019).
Durinck, S., Spellman, P. T., Birney, E. & Huber, W. Mapping identifiers for the integration of genomic datasets with the R/Bioconductor package biomaRt. Nat. Protoc. 4, 1184–1191 (2009).
Wolf, F. A., Angerer, P. & Theis, F. J. SCANPY: large-scale single-cell gene expression data analysis. Genome Biol. 19, 15 (2018).
Fang, Z., Liu, X. & Peltz, G. GSEApy: a comprehensive package for performing gene set enrichment analysis in Python. Bioinformatics 39, btac757 (2023).
Frankish, A. et al. GENCODE reference annotation for the human and mouse genomes. Nucleic Acids Res. 47, D766–D773 (2019).
Yates, A. D. et al. Ensembl 2020. Nucleic Acids Res. 48, D682–D688 (2020).
Marçais, G. & Kingsford, C. A fast, lock-free approach for efficient parallel counting of occurrences of k-mers. Bioinformatics 27, 764–770 (2011).
Gans, J. D. & Wolinsky, M. Improved assay-dependent searching of nucleic acid sequence databases. Nucleic Acids Res. 36, e74 (2008).
Rodriguez, J. M. et al. APPRIS 2017: principal isoforms for multiple gene sets. Nucleic Acids Res. 46, D213–D217 (2018).
Bankhead, P. et al. QuPath: open source software for digital pathology image analysis. Sci. Rep. 7, 16878 (2017).
Van der Walt, S. et al. scikit-image: image processing in Python. PeerJ 2, e453 (2014.
Niederst, E. D., Reyna, S. M. & Goldstein, L. S. Axonal amyloid precursor protein and its fragments undergo somatodendritic endocytosis and processing. Mol. Biol. Cell 26, 205–217 (2015).
Taylor, A. M. et al. A microfluidic culture platform for CNS axonal injury, regeneration and transport. Nat. Methods 2, 599–605 (2005).
Khan, M. et al. Visualizing in deceased COVID-19 patients how SARS-CoV-2 attacks the respiratory and olfactory mucosae but spares the olfactory bulb. Cell 184, 5932–5949 (2021).
Acknowledgements
We would like to thank the LIMONE Bioimaging Core of VIB-KU Leuven for their support in the imaging experiments, in particular A.E. Ayala and A. Kerstens; the VIB Nucleomics Core; the histology facility headed by S. Minhas; the mouse expertise unit headed by L. Serneels, A. Vanschoiack (NanoString) and R. Das (Resolve Biosciences) for their critical expertise; B. Tilkin and V. Ferrari for the technical support. This work was supported by the FWO (Research Foundation-Flanders, G064721N), the Muscular Dystrophy Association (MDA; 962700), the Alzheimer Research Foundation (SAO; grant SAO-FRA 20230035 to S.D.C.), VIB-Tech Watch (to D.P. and S.D.C.), KU Leuven Opening the Future (to S.K., K.V., T.V. and S.D.C.) and the ALS Canada and Brain Canada (Discovery Grant). D.P. was a recipient of the FWO postdoctoral junior fellowship (award 1229621N) and a SAO-FRA (Alzheimer Research Foundation) pilot grant (award 20200020) and is currently supported by the MDA Development Grant (1060285). B.K. is supported by an SAO-FRA pilot grant (award 20230042) and the Alzheimer’s Association Research Fellowship to Promote Diversity (AARF-D-24-1313422). The funders had no role in study design, data collection and analysis, decision to publish or preparation of the paper.
Author information
Authors and Affiliations
Contributions
D.P., A.S. and S.D.C. conceptualized the study. D.P., B.K., T.R., T.K., M.G., G.P., D.W., P.T., Y.V., N.H., S.K., K.V. and K.D. conducted the investigation. D.P., B.K., T.R., T.K., M.G., G.P., D.W., P.T., Y.V., N.H., N.C., S.K., K.V. and K.D. developed the methodology. D.P., B.K., T.R., T.K., M.G., G.P., D.W., P.T., Y.V., N.H., S.K., K.V. and K.D. curated the data. D.P., B.K., T.R., T.K., M.G., G.P., P.T. and K.D. carried out formal analysis. D.P., B.K., T.R., T.K., M.G. and G.P. handled visualization. D.P., T.V., K.D., S.P., P.V.D., L.V.D.B., S.A., A.S. and S.D.C. provided supervision. D.P. and S.D.C. wrote the original draft of the paper. D.P., A.S. and S.D.C. did the writing, reviewing and editing of the paper. D.P., B.K., P.V.D., L.V.D.B., S.A., T.V., A.S. and S.D.C. secured funding.
Corresponding author
Ethics declarations
Competing interests
L.V.D.B. is head of the Scientific Advisory Board of Augustine Therapeutics and is part of the Investment Advisory Board of Droia Ventures. The other authors declare no competing interests.
Peer review
Peer review information
Nature Neuroscience thanks the anonymous reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Analysis of the expression of known marker genes in specific spinal cord cell types through untargeted/targeted spatial transcriptomics and comparison with publicly available snRNA-seq datasets.
a, Heatmap expression of selected spinal cord cell-type marker genes in motor and nonmotor neurons from the NanoString GeoMx DSP analysis. Colored scale, scaled log2-normalized counts. b–j, Multiplexed smFISH images of motor neuron (b), microglia (c), macrophage (d), oligodendrocyte (e), dorsal horn (f), oligodendrocyte progenitor cell (g), astrocyte (h), excitatory neuron (i) and slow-firing motor neuron (j) markers in 12-month-old mouse spinal cords. Scale bars, 500 µm and 100 µm for inset. k, 10x Genomics Visium spatial visualization of selected genes found to be most enriched in motor neurons from the NanoString GeoMx DSP dataset in 12-month-old animals. l, 10x Genomics Visium spatial visualization of selected genes found to be most enriched in nonmotor neurons from the NanoString GeoMx DSP dataset. m, 10x Genomics Visium spatial visualization of Stmn2 and Lgals1 mRNAs. For k,l,m, tissues are stained with the H&E technique, and gray/white dots represent the capture areas of the 10x Genomics Visium Chip. Scale bars, 500 µm and 100 µm for insets. n,o, Representative multiplexed smFISH images of Chat with Stmn2 (n) or Lgals1 (o) transcripts in 12-month-old mouse spinal cords. Scale bars, 500 µm and 100 µm for inset. p, Venn diagram showing the overlap of DEGs between the motor neuron cluster identified in the present study and in the indicated publicly available datasets27,28,29,30. q–t, Correlation plots between our NanoString GeoMx DSP dataset and the indicated published snRNA-seq study. Data are represented as a regression line and relative s.e.m. P values = 2.2 × 10−16 for all comparisons.
Extended Data Fig. 2 Spatial expression pattern of axonal mRNAs and cell-specific markers in transversal and longitudinal mouse sciatic nerve sections.
Multiplexed smFISH images of (a) axonal, (b) microglia/macrophage, (c) endoneurial fibroblasts, (d) macrophage, (e) nonmyelinating Schwann cell, (f) peri/endo/epineurial cell, (g,h) myelinating Schwann cell and (i) general Schwann cell markers in 12-month-old mouse sciatic nerve transversal sections. j, Quantifications of the indicated transcripts inside axons marked with NFH. Data are represented as mean ± s.e.m. (n = 6–8 biological replicates). Scale bars, 100 µm; 10 µm and 5 µm for inset.
Extended Data Fig. 3 Newly identified transcripts localized in mature axons of adult mouse sciatic nerves using axonal spatial transcriptomics.
a–e, Representative images of multiplexed smFISH combined with immunofluorescence of (a) Eef2, Rpl4 and Rpl15; (b) Hnrnpa2b1; (c) Sparc; (d) Cox4i1; (e) Nefh and Nefl transcripts (pink) together with immunolabeled axons using NFH antibody (white) in 12-month-old mouse sciatic nerve transversal sections. (f) Quantifications of the indicated transcripts inside axons marked with NFH. Data are represented as mean ± s.e.m. (n = 3–8 biological replicates). Scale bars, 100 µm and 5 µm for insets.
Extended Data Fig. 4 Mutant FUS-specific dysregulated RNA signature in adult motor axons of mFUSKO/hFUSR521H sciatic nerves identified using axonal spatial transcriptomics.
a, Heatmap of the top 20 DEGs between 12-month-old nontransgenic and mFUSKO/hFUSR521H motor axons. Colored scale, scaled log2-normalized counts. b, Gene ontology enrichment analysis of the most upregulated genes in mFUSKO/hFUSR521H motor axons obtained with DAVID. c, Gene ontology enrichment analysis of the most downregulated genes in mFUSKO/hFUSR521H motor axons obtained with DAVID. d, GeoMx DSP normalized counts of the indicated genes belonging to the translation machinery. Data are represented as mean ± s.e.m. (n = 4). P value = 0.031 (Eif1), not significant (other genes) using an unpaired two-tailed t test. Each data point corresponds to the measurement in one section. e, Expression pattern of the indicated genes in the motor neuron cluster obtained by applying 10x Genomics Visium spatial transcriptomics. Scales represent the log2-normalized expression. f, Violin plot of the log-normalized expression for the indicated genes in the motor neuron cluster originating from d. Data represent four biological replicates. The embedded box plots indicate the median (center line), interquartile range (25th and 75th percentiles; box edges) and whiskers extending to 1.5 times the interquartile range. g, GeoMx DSP normalized counts of the indicated genes. Data are represented as mean ± s.e.m. (n = 4). P values = not significant using unpaired two-way t test. Each data point corresponds to the measurement in one section.
Extended Data Fig. 5 The Eif5a hypusination pathway is not altered in motor neuron cell bodies of mutant FUS spinal cords.
a, GeoMx DSP normalized counts of the indicated genes belonging to the hypusination pathway. Data are represented as mean ± s.e.m. (n = 4). P values = not significant using unpaired two-tailed t test. Each data point corresponds to the measurement in one section. b, Representative images (left) and quantification (right) of the analysis of Eif5a transcripts in motor neurons (marked with Chat and Chodl transcripts) using multiplexed smFISH. Scale bars, 500 µm and 100 µm for insets. Data are represented as mean ± s.e.m. (n = 3–4 biological replicates). P value = not significant using unpaired two-tailed t test. c, Immunofluorescence analysis of Eif5a in spinal cord motor neurons in nontransgenic and mFUSKO/hFUSR521H animals (left) and relative quantification (right). Data are represented as mean ± s.e.m. (n = 3 biological replicates). P value = not significant using unpaired two-tailed t test. Scale bar, 50 µm. d, Western blot analysis of N2A cell lysate run in a 4–15% polyacrylamide gel and probed with antibodies specific for hypusine and Eif5a. On the left, the membrane image is juxtaposed to the enhanced chemiluminescent (ECL) signal. The experiment was performed once. e, Immunofluorescence analysis of hypusinated Eif5a in spinal cord motor neurons in nontransgenic and mFUSKO/hFUSR521H animals (left) and relative quantification (right). Scale bar, 50 µm. Data are represented as mean ± s.e.m. (n = 3 biological replicates). P value = not significant using unpaired two-tailed t test. f, Expression pattern of Eif5a, Dohh and Dhps, in the motor neuron cluster obtained by applying 10x Genomics Visium spatial transcriptomics. Scales represent the log-normalized expression. g, Violin plot of the log-normalized expression for the indicated genes in the motor neuron cluster originating from f. Data represent four biological replicates. P values are calculated using one-way ANOVA with Bonferroni’s multiple comparisons test. The embedded box plots indicate the median (center line), interquartile range (25th and 75th percentiles; box edges) and whiskers extending to 1.5 times the interquartile range.
Extended Data Fig. 6 Decreased total Eif5a and increased inactive acetylated Eif5a levels are rescued by spermidine treatment in mutant FUS cortical axons.
a,b, Immunofluorescence analysis of puromycin, total (a) or acetylated (b) Eif5a and NFH in nontransgenic or hFUSR521H-derived primary cortical neurons treated with or without spermidine 100 µM for 48 h, and then pulse-labeled with puromycin (experiment described in the schematic in Fig. 5a). Scale bars, 25 µm; 10 µm for insets. c,d, Quantifications of total (c) and acetylated Eif5a (d) signals in the indicated conditions. Data are represented as mean ± s.e.m. (n = 3 biological replicates, each dot corresponding to one axonal region of interest). P values = 0.011 (c, non-tg versus hFUSR521H), 0.004 (c, FUSR521H versus FUSR521H SPMD), 0.003 (d, non-tg versus hFUSR521H) and 0.001 (d, FUSR521H versus FUSR521H SPMD) using one-way ANOVA with Bonferroni’s multiple comparisons test.
Extended Data Fig. 7 ALS-linked mutation in FUS does not impair hypusine levels nor total protein synthesis in cell bodies.
a, Immunofluorescence analysis of puromycin, hypusine and 4′,6-diamidino-2-phenylindole (DAPI) in nontransgenic or hFUSR521H-derived primary cortical neuron cell bodies. Scale bar, 10 µm. b,c, Quantifications of hypusine (b) and puromycin (c) signals in the indicated conditions. Data are represented as mean ± s.e.m. (n = 3 biological replicates). P values are calculated using one-way ANOVA with the Bonferroni multiple comparisons test. d, Immunofluorescence analysis of puromycin, hypusine and DAPI staining in nontransgenic or hFUSR521H-derived primary motor neuron cell bodies, treated in the axonal compartment with spermidine 100 µM for 48 h and pulse-labeled with puromycin. Scale bar, 10 µm. e,f, Quantifications of hypusine (e) and puromycin (f) signals in the indicated conditions. Data are represented as mean ± s.e.m. (n = 3 biological replicates). P values are calculated using one-way ANOVA with the Bonferroni multiple comparisons test. g, Axonal branching quantification from Fig. 6e. Data are represented as mean ± s.e.m. (n = 3–5 biological replicates). P values are calculated using one-way ANOVA with the Bonferroni multiple comparisons test. h, Effect of spermidine treatment ranging between 0 and 5 mM on the eclosion of the control fly line. Data are represented as mean ± s.e.m. (n = 15–16 crosses/condition from three independent experiments). P values are calculated using one-way ANOVA with Holm–Šídák’s multiple comparisons test.
Supplementary information
Supplementary Table 1 (download XLSX )
Differentially expressed gene lists used for the comparative analysis between the GeoMx DSP dataset and publicly available snRNA-seq datasets.
Supplementary Table 2 (download XLSX )
Gene Ontology analysis of the axonal transcripts identified through GeoMx DSP.
Source data
Source Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 7 (download XLSX )
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Piol, D., Khalil, B., Robberechts, T. et al. Axonal Eif5a hypusination controls local translation and mitigates defects in FUS-ALS. Nat Neurosci 29, 53–66 (2026). https://doi.org/10.1038/s41593-025-02101-2
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41593-025-02101-2
