
Abstract
Animal germ cells deploy a specialized small RNA-based silencing system, called the PIWI-interacting RNA (piRNA) pathway, to prevent unwanted expression of transposable elements (TEs) and maintain genome integrity. In Drosophila melanogaster germ cells, the majority of piRNA populations originate from dual-strand piRNA clusters, genomic regions highly enriched in TE fragments, via an elaborate machinery centered on the Heterochromatin Protein 1 homolog, Rhino. Although Rhino binds to peptides carrying tri-methylated H3K9 in vitro, it is not fully understood why in vivo only a fraction of H3K9me3-decorated heterochromatin is occupied by Rhino. Recent work revealed that Rhino is recruited to a subset of piRNA clusters by Kipferl. Here we identify a Kipferl-independent mode of Rhino recruitment that, in addition to the previously established role of H3K9me3, also depends on the histone H3 lysine 27 methyltransferase Enhancer of Zeste. At Kipferl-independent sites, we find that Rhino specifically binds to loci marked by both H3K9me3 and H3K27me3 via its chromodomain. Although the exact mechanism of how Rhino binding is influenced by dual histone modifications remains unclear from a structural and biochemical perspective, our work suggests that combinatorial modifications may regulate the specificity of chromatin-binding protein interactions. These findings provide an enhanced understanding of how Rhino targets piRNA source loci, highlighting the sophisticated epigenetic landscape governing TE silencing in Drosophila germ cells.
Similar content being viewed by others

Main
Transposable elements (TEs) are genetic sequences, present in nearly all organisms, that are able to move and insert themselves into different positions within a genome. While TEs can contribute to genetic diversity and evolution, their uncontrolled activity in gonadal cells can have detrimental effects, including the potential to dramatically reduce fertility. In animal gonads, a mechanism known as the PIWI-interacting RNA (piRNA) pathway plays a crucial role in silencing and controlling active TEs to safeguard germline integrity1,2,3. The piRNA machinery utilizes 23–30-nt small RNAs, which associate with PIWI-family proteins, directing recognition of target TEs for post-transcriptional degradation or cotranscriptional silencing. While some piRNAs are derived from active TEs, the majority originate from dedicated piRNA source loci (piSL). Of those, a subset, called piRNA clusters, are enriched in TE content. All reported piRNA clusters in somatic follicle cells resemble canonical genes, give rise to piRNAs from one genomic strand and are referred to as unistrand clusters4. In contrast, piRNA clusters in the germline are typically expressed non-canonically, producing piRNAs from both genomic strands and are referred to as dual-strand clusters5,6,7,8,9,10.
Rhino (Rhi), a homolog of Heterochromatin Protein 1a (HP1a), is a germline-specific piRNA pathway factor essential for piRNA production, TE repression and fertility9. Rhi and HP1a belong to a distinct group of chromatin-binding proteins containing chromodomains (CDs), which display specific binding properties. For instance, Drosophila HP1a is associated with H3K9me2/3-enriched regions such as constitutive heterochromatin, whereas Polycomb (Pc), another CD protein, specifically binds to H3K27me3 often found in facultative heterochromatin11,12,13. In line with Rhi being evolutionary related to HP1a, several studies have shown that in vivo Rhi specifically localizes to a subset of piSL, including dual-strand piRNA clusters, which are typically characterized by the presence of H3K9me3 marks6,8,9,14. In vitro, the Rhi CD was shown to recognize di-/tri-methylated H3K9 peptides6,14,15.
At the genomic regions to which it binds, Rhi acts as nucleator for a multiprotein complex that triggers non-canonical transcription from heterochromatic regions and facilitates the export of the resulting piRNA precursors from the nucleus6,7,8,9,10,14,16,17,18,19,20,21. Although Rhi binds H3K9me3 in vitro6,14,15, Rhi immunofluorescence staining and previously published chromatin immunoprecipitation followed by sequencing (ChIP–seq) data indicate that in vivo Rhi associates only with a fraction of genomic regions enriched in H3K9me3 (ref. 6), hence raising questions about how its specificity is determined in vivo. A recent study identified the zinc finger domain protein Kipferl (Kipf) as a Rhi co-factor, with their interaction mediated by the CD of Rhi22,23. Kipf recruits Rhi to dedicated heterochromatic loci that exhibit enrichment in guanosine-rich motifs, suggesting that sequence content within piSL could contribute to the binding specificity of Rhi. However, while the interaction between Kipf and Rhi is critical for the recruitment of Rhi to numerous genomic loci, prominent Rhi-dependent piSL, such as piRNA clusters 42AB and 38C, show little to no dependence on Kipf22. These observations suggest that additional cues contribute to Rhi binding. Here, we uncover an unexpected, Kipf-independent mechanism for Rhi targeting that relies on the H3K27me3 methyltransferase Enhancer of Zeste (E(z)). We propose a model in which the combined recognition of H3K9me3 and H3K27me3 guides Rhino to a subset of piSL, including the most prominent cluster 42AB.
E(z) is required for TE silencing in Drosophila germ cells
Understanding how Rhi is recruited to and interacts with chromatin at piSL is crucial for unraveling the complexities of TE silencing and genome integrity. We therefore searched for factors affecting TE control in germ cells through a focused reverse-genetic screen in Drosophila melanogaster ovaries (Fig. 1a). We specifically targeted chromatin proteins and histone modifiers and probed the effects of their depletion on germline-specific TEs by quantitative RT–PCR (RT–qPCR). Knockdown of 32 genes resulted in TE derepression and highlighted potential roles of different protein complexes in TE regulation (for example, COMPASS complex and NSL complex) (Extended Data Fig. 1).

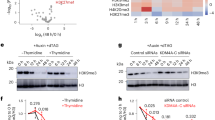
a, A schematic showing the in vivo screen workflow. Male flies carrying dsRNA or shRNA constructs under the inducible UAS promoter were crossed to females expressing the germline-specific nos-GAL4 driver and a Dcr2 transgene. F1 offspring were transferred to new vials and fertility was determined by counting eggs laid and hatched. RNA was extracted from ovaries of F1 offspring and TE levels were measured by RT–qPCR. b, An MA plot showing counts per gene (gray) and TEs (black) in polyA-selected RNA-seq libraries from ovaries depleted of w (control) and E(z) using the nos-Gal4 driver. TEs with Padj <0.05 and >4× fold change (FC) are shown in red, E(z) is indicated in purple. c, Confocal images showing RNA-FISH signal for the indicated TEs (burdock, copia and gypsy12) upon nos-Gal4 mediated knockdown or w or E(z). Scale bar, 10 µm. Data are representative of n ≥ 3 independent experiments. d, Confocal images showing RNA-FISH signal for transcripts derived from piRNA clusters 42AB, 80F and 20A in control and E(z)-depleted ovaries using the nos-Gal4 driver. Scale bar, 5 μm. Data are representative of n ≥ 3 independent experiments. e, A scatter plot depicting normalized ribo-depleted RNA levels (FPKM) of uniquely mapping reads in 1 kb bins in ovaries with nos-Gal4-driven E(z) knockdown versus control (average of three replicate experiments each). f, A scatter plot depicting normalized piRNA levels of uniquely mapping piRNAs (FPKM) in 1 kb bins (118,294 in total) in ovaries with nos-Gal4-driven E(z) knockdown versus control (two replicates each). g, UCSC genome browser tracks displaying the dual-strand piRNA cluster 42AB. Levels of uniquely mapping piRNAs and ribo-depleted RNAs from ovaries with the indicated knockdowns using the pTOsk-Gal4 driver (average of three replicate experiments each) are shown along with tracks displaying genes, repeats and mappability. The dashed line indicates approximate piRNA cluster boundaries. h, As in g but showing the dual-strand piRNA cluster 80F.
As expected from previous reports24, depletion of the H3K9 methyltransferase Eggless (Egg) resulted in a strong deregulation of germline TEs and a loss of Rhi foci by immunofluorescence (Extended Data Figs. 1 and 2a). Unexpectedly, however, depletion of the histone methyltransferase E(z) resulted in TE upregulation (Extended Data Fig. 1). Of note, we also found that the depletion of other subunits of the Polycomb repressive complex 2 (PRC2), namely Su(z)12 and Caf1-55, resulted in the derepression of some TEs (Supplementary Note 1). In Drosophila, E(z) is responsible for H3K27me3 and is essential for Polycomb-mediated chromatin silencing25,26. Depletion of E(z) using a strong Gal4 driver (UAS-Dcr2 transgene combined with nos-Gal4) caused rudimentary ovaries. To exclude the possibility that the observed TE derepression was due to pleiotropic oogenesis defects, we used different Gal4 drivers, namely nos-Gal4, which is active from the germarium through stage 2, then inactive between stages 3 and 6 of oogenesis but reactivated at later stages27, and pTOsk-Gal4, which is expressed immediately after germline cyst formation28, and two independent E(z) RNAi lines, all of which resulted in nearly wild-type ovarian morphology despite reduced H3K27me3 levels in nurse cell nuclei (Extended Data Fig. 2b), and unchanged Rhi localization as observed by immunofluorescence (Extended Data Fig. 2a). Sequencing of polyA-selected RNAs from ovaries depleted of E(z) using the nos-Gal4 driver revealed a strong upregulation of 13 germline TE families, including gypsy12, burdock and copia (Fig. 1b). This was validated by RNA-fluorescence in situ hybridization (FISH), with burdock, gypsy12 and copia expression detected in germ cells depleted of E(z) (Fig. 1c). Overall, however, the effects on TEs that we observed for E(z) knockdown were less severe compared with ovaries depleted of Rhi, which affected 53 TE families, suggesting partial contributions of E(z) to Rhi function (Extended Data Fig. 2c; data from ref. 22).
To distinguish between compromised global chromatin silencing and a specific role in the piRNA pathway, we performed RNA-FISH for precursor transcripts derived from the dual-strand piRNA clusters 42AB (Kipf independent) and 80F (Kipf dependent), and the unistrand cluster 20A (germline enriched but Rhi independent). Knockdown of E(z) revealed a strong loss in piRNA precursors derived from 42AB, but not from 80F and 20A, suggesting a potential role of E(z) in transcription of Kipf-independent dual-strand piRNA clusters (Fig. 1d and Extended Data Fig. 2d).
Next, we analyzed the effects of E(z) knockdown on piRNA precursors globally using ribo-depleted RNA-sequencing (RNA-seq). To measure transcription across all piSL, we divided the genome into 1 kb bins and quantified the signal per bin, as previously described6. Upon germline-specific E(z) depletion, we observed a strong reduction in precursor transcripts across 42AB, and moderately reduced levels for 38C, whereas 20A or flam (soma-expressed and Rhi-independent) were essentially unchanged (Fig. 1e and Extended Data Fig. 2e). Strikingly, precursor levels for Kipf-dependent piRNA cluster 80F were increased. RT–qPCR confirmed the RNA-seq results (Extended Data Fig. 2f). Of note, and as reported previously6,9,14, Rhi-depleted ovaries showed a specific and global reduction of precursor transcripts from dual-strand clusters (Extended Data Fig. 2g). These data suggest a more nuanced role of E(z) in TE silencing compared with Rhi.
Next, we sequenced piRNAs from ovaries in which E(z) or Rhi were depleted specifically in germ cells and compared these with control knockdowns (Supplementary Table 1). Consistent with the reduction of precursor transcripts from 38C and 42AB upon E(z) knockdown (Fig. 1e), we detected a severe reduction in piRNA levels (Fig. 1f). As expected from their precursor expression, piRNAs from 80F increased, while piRNAs from the unistrand clusters 20A and flam were unaffected (Fig. 1f and Extended Data Fig. 2h). The reduction in precursor and piRNA levels observed at cluster 42AB was similar for ovaries depleted for E(z) and Rhi (Fig. 1g). In contrast, precursors and piRNAs were increased at 80F, whereas there was a pronounced reduction of these RNA populations in the rhi knockdown (Fig. 1h).
E(z)-depleted ovaries also showed fewer antisense piRNAs targeting individual germline-specific TEs, which could explain their increased mRNA levels (Extended Data Fig. 2i). As expected, piRNAs against TEs that are mainly active in the somatic compartment of the ovary (for example, Tabor, gypsy and ZAM), were not affected, probably due to their origin from the soma-specific flam cluster (Extended Data Fig. 2i,j). Altogether, our results reveal that the histone methyltransferase E(z) is required for TE silencing and that its depletion affects piRNA production in germ cells from several dual-strand clusters.
H3K27me3 and H3K9me3 marks coexist on E(z)-dependent piSL
To elucidate why certain piSL require E(z), we examined the genome-wide distribution of H3K27me3, which is deposited by E(z), and H3K9me3, which has been associated with piRNA clusters and Rhi recruitment, by performing ChIP–seq and CUT&RUN (Extended Data Fig. 3a). We found that a number of genomic regions, including dual-strand piRNA cluster 42AB, are decorated by both H3K9me3 and H3K27me3 (Fig. 2a,b). A similar, albeit less strong, pattern was observed at other piSL, including the ey/Sox102F region on chromosome 4 (Extended Data Fig. 3b). Notably, Rhi preferentially occupied regions decorated both by H3K9me3 and H3K27me3 over those only carrying H3K9me3 (Fig. 2c). Additionally, the majority of H3K27me3 peaks (76%) did not overlap with Kipf (Extended Data Fig. 3c). To determine whether one of the two marks, or both, could best predict piRNA clusters, we divided the genome into 1 kb bins and ranked these based on their histone mark enrichment. To compare different marks, we calculated their classification performance as area under the receiver operating characteristic (ROC) curve (AUC) (Extended Data Fig. 3d). We used published HP1a ChIP–seq29 as a proxy for H3K9me3 as the H3K9me3 antibody appeared to capture some H3K27me3 signal (Supplementary Note 2). We found that H3K27me3 is an accurate predictor of dual-strand piRNA cluster location (AUC 0.867), similar to the predictive power of HP1a binding (AUC 0.842), but that combining both marks improved classification further (AUC 0.927) (Extended Data Fig. 3e). Of note, we observed that H3K27me3 enrichment correlated with E(z) dependence in piRNA production: clusters 42AB and 38C showed strong enrichment in H3K27me3 (Fig. 2d,e), whereas 20A and 80F whose piRNA production was E(z) independent displayed lower levels of H3K27me3 (Fig. 2d,f). This points toward a role for both H3K27me3 and H3K9me3 in the specification of certain piSL.

a, UCSC genome browser tracks of the region comprising dual-strand piRNA cluster 42AB displaying ChIP–seq signal (depicted as coverage per million reads) of Rhi (blue), H3K27me3 (red), H3K9me3 (orange), uniquely mapping piRNAs (purple, coverage normalized to miRNA reads) and tracks indicating genes and repeats in nos-Gal4>sh-w ovaries. b, A box plot showing the average log2-fold ChIP–seq enrichment over input of H3K27me3 (red), H3K9me3 (orange) and Rhi (blue) at the indicated piSL (n = 1 kb bins) in control ovaries. The box plots show the median (center line), with interquartile range (box) and whiskers indicating 1.5× the interquartile range at most). c, A Venn diagram showing the overlap between Rhi, H3K9me3 and H3K27me3 ChIP–seq signal across genomic 1 kb bins. Signal was considered to be present if the bin overlapped a MACS2 broad peak (q < 0.05, broad cutoff <0.1) across two pooled biological replicates. Enrichment was calculated as an OR between Rhi binding in the presence of indicated factor or without. d, A heat map displaying predictive performance measured as the AUC across different chromatin marks (top) and target regions (left). The regions are ordered according to their AUC for HP1a + H3K27me3. e, A line graph comparing the ability to identify dual-strand piRNA cluster 42AB based on the strength of individual features (H3K27me3 or HP1a) and combinations (mean signal for H3K27me3 + HP1a). f, As in e but showing unistrand piRNA cluster 20A. FPR, false positive rate; TPR, true positive rate.
Rhi binding to a subset of piSL depends on E(z)
The dual-strand cluster 42AB is the major source of Rhi-dependent piRNAs in germ cells (Fig. 3a), and our data indicate that its piRNA production is E(z) dependent. Prior work implicates Kipf in guiding Rhi to a subset of piSL, including 80F, which we find is E(z) independent (Fig. 1d–f,h and Extended Data Fig. 2d–f,h), but not 42AB22. We therefore hypothesized that Rhi binding to piSL that are Kipf independent may depend on E(z). To test this, we compared piRNA production from Rhi-dependent loci (n = 632, ≥2-fold piRNA reduction after Rhi loss) in Kipf- or E(z)-depleted ovaries. We observed that E(z), but not Kipf, is required for efficient piRNA production from 42AB and piSL on chr4, whereas 80F and most heterochromatic regions are E(z) independent but require Kipf (Fig. 3b,c). Other piRNA clusters, such as 38C, depend weakly on both Kipf and E(z) (Fig. 3b). Together, this suggests that E(z) and Kipf may provide distinct modes of Rhi recruitment to separate piSL.

a, The genomic origin of Rhi-dependent piRNAs. The analysis was done across 1 kb bins (n = 632) that displayed Rhi dependency (>2-fold mean piRNA loss) across several depletion strategies (pTOsk-Gal4 or MTD-Gal4-driven knockdown, and rhi knockout; one to two replicates each). b, Scatter plots showing log2-fold mean changes in uniquely mapping piRNAs in 1 kb bins relative to control (Ctrl) across several depletion strategies (nos-Gal4 or pTOsk-Gal4-driven knockdown of E(z), two replicates each; MTD-Gal4-driven knockdown of kipf or kipf knockout, one replicate each). Bins were categorized into E(z) dependent and/or Kipf dependent if displaying ≥4-fold mean loss of piRNAs. c, Genomic origin for 1 kb bins per E(z)/Kipf-dependency category as defined in b. d, A scatter plot showing log2-fold change in Rhi ChIP–seq enrichment following depletion of E(z) using nos-Gal4 or Kipf using MTD-Gal4. The dashed lines indicate a twofold mean loss of Rhi. e, Violin plots (left) and box plots (right) showing the average log2-fold Rhi enrichment by ChIP–seq from ovaries with nos-Gal4-driven E(z) knockdown versus control (average of two replicates each) in heterochromatin (HC) and euchromatic chromosome arms (EC), quantified across 1 kb bins (excluding piRNA clusters). Rhi occupancy at indicated piRNA clusters is shown as box plots (n depicts 1 kb bins analyzed). ****P < 0.0001 based on a two-sided Wilcoxon signed-rank test. Box plots show the median (center line), with interquartile range (box) and whiskers indicating 1.5× the interquartile range at most). f, A scatter plot of genomic 1 kb bins contrasting average log2-fold ChIP–seq enrichment of Rhi in ovaries with nos-Gal4-driven E(z) knockdown versus control (average of two replicates). g, A metaplot showing the mean difference in H3K27me3 and Rhi ChIP–seq signal upon nos-Gal4 driven E(z) or kipf knockdown across Rhi peaks categorized as either Kipf dependent or not (Methods). h, Genome browser tracks of the dual-strand piRNA cluster 42AB displaying uniquely mapping piRNAs upon the indicated (double-)knockdowns. i, Confocal images showing RNA-FISH signal for transcripts derived from 42AB in control, E(z) and kipf knockdown as well as in E(z)-kipf dKD, using the nos-Gal4 driver. Scale bar, 5 μm. Data are representative of n ≥ 3 independent experiments.
The production of piRNAs from dual-strand clusters depends on Rhi and we therefore expect piRNA production patterns to closely reflect Rhi binding6. To determine whether E(z) directs Rhi binding to selected piSL, we performed Rhi ChIP–seq in E(z)-depleted ovaries and compared this with Rhi ChIP–seq in Kipf-depleted ovaries22. We observed that Rhi binding closely mirrored piRNA production (Fig. 3d). Consequently, upon E(z) depletion, Rhi enrichment increased at Kipf-dependent piSL and decreased at piSL that depend on E(z) (Extended Data Fig. 4a). We next asked whether depletion of E(z) affects the global distribution of Rhi. In agreement with largely unchanged localization by immunofluorescence (Extended Data Fig. 2a), we found that Rhi was not globally redistributed between euchromatin and heterochromatin (Fig. 3e). Instead, we detected a strong reduction of Rhi at specific regions, including the ey/Sox102F region and cluster 42AB (Fig. 3e and Extended Data Fig. 4b,c), whereas other loci, such as the 80F, accumulated more Rhi (Fig. 3e). Those observations agreed with changes in H3K27me3 (Extended Data Fig. 4d). Notably, the observed changes in Rhi occupancy (Fig. 3f) followed piRNA precursor levels, which showed reduced levels for cluster 42AB and an enrichment at 80F (Fig. 1d,e). Moreover, the observed differences in Rhi binding were not explained by changes in H3K9me3, which remains essentially unaltered (Extended Data Fig. 4b,c,e).
We next analyzed published ChIP–seq data22 for genome-wide Rhi and Kipf occupancy. We identified 1,629 high-confidence Rhi binding sites using MACS2 (q < 0.05, >4-fold enrichment). Out of these, 1,166 peaks were lost in kipf knockdown and were denoted as Kipf dependent, while 463 peaks were retained and denoted as Kipf independent (Extended Data Fig. 4f). Supporting separate modes of Rhi recruitment, we observed no reduction in H3K27me3 or Rhi occupancy at Kipf-dependent Rhi peaks following E(z) depletion, in line with these loci lacking H3K27me3 marks and suggesting that Rhi is recruited by Kipf in an H3K27me3-independent manner. In contrast, we observed a broad loss of both H3K27me3 and Rhi occupancy at Kipf-independent Rhi peaks (Fig. 3g), while H3K27me3 was not affected by Kipf depletion (Fig. 3g). This suggests a strong correlation between reduced Rhi binding at Kipf-independent sites and the loss of H3K27me3 following E(z) knockdown.
In E(z)-depleted ovaries, our polyA-selected RNA-seq data indicated an up to sixfold reduction in mRNA levels of Moonshiner (Moon), a paralog of TFIIA required for piRNA production from the majority of Rhi-dependent piSL7 (Supplementary Table 2). We also observed reduced Rhi binding at Kipf-independent peaks in published ChIP–seq from moon knockdown7 (Extended Data Fig. 4g). It is thus possible that Rhi loss upon E(z) depletion may in part be driven by reduced Moon levels. However, we consider secondary effects driven by Moon unlikely since its loss was previously reported to lead to a drastic reduction of piRNAs from cluster 80F and an increase in 38C-derived piRNAs7, opposite to what we observe.
Overall, our results provide evidence that H3K9me3 and H3K27me3 coexist on a subset of piSL and appear to be required for Rhi recruitment at these regions. Thus, two non-redundant mechanisms influence Rhi binding to different piSL, namely E(z)-dependent and Kipf-dependent recruitment.
Kipf and E(z) independently guide Rhi to its targets
To further investigate the two Rhi recruitment mechanisms, we next determined the effect of simultaneous knockdown of E(z) and kipf. We sequenced small RNAs from E(z)-kipf double-knockdown (dKD) and control ovaries. When compared with knockdown of w, TE-derived piRNAs were reduced upon E(z) knockdown (Extended Data Fig. 5a). Notably, co-depletion of Kipf and E(z) resulted in more severe effects on piRNA levels (Extended Data Fig. 5b). However, while the combined depletion of E(z) and Kipf has mild effects on piRNA populations relative to E(z) knockdown alone (Extended Data Fig. 5c), it leads to pronounced effects when compared with only kipf knockdown (Extended Data Fig. 5d).
Next, analyzing piRNA levels at piSL revealed a strong reduction of piRNAs from the cluster 42AB in E(z)-kipf dKD and E(z) knockdown compared with kipf or control knockdowns (Fig. 3h). To quantify these observations, we compared each (double)knockdown to respective controls across 1 kb bins (Extended Data Fig. 5e). As expected, piRNA production from the unistrand clusters flam and 20A remained unchanged. Upon E(z)-kipf dKD we observed a 3.7-fold stronger reduction of piRNAs derived from 42AB compared with E(z) depletion alone (Extended Data Fig. 5e–g). As observed for TE-derived piRNAs, the combined depletion of E(z) and kipf had a mild effect on cluster-derived piRNAs compared with E(z) knockdown alone, but a stronger effect compared to kipf knockdown alone (Extended Data Fig. 5e,h,i). Unexpectedly, while kipf knockdown alone resulted in reduced piRNA levels from 80F, we observed a recovery of piRNA production to near-normal levels in E(z)-kipf dKD ovaries (Extended Data Fig. 5e,g). These results suggest that in the absence of both E(z) and Kipf, Rhi can nevertheless bind to piSL 80F.
Next, using RNA-FISH, we found that, similar to E(z) knockdown, no signals for 42AB were detected in the E(z)-kipf dKD ovaries (Fig. 3i). These results confirmed that transcription of 42AB piRNA precursors relies on E(z) and is Kipf independent. Notably, Rhi, which typically accumulates in large, continuous structures near the nuclear envelope at satellite regions in kipf knockdowns22, becomes redistributed throughout the nucleus in the E(z)-kipf dKD, resembling control and E(z)-depleted ovaries (Extended Data Fig. 5j). Overall, these results highlight distinct, non-redundant mechanisms of Rhi recruitment at different piSL with some relying on E(z) and others on Kipf, but no evidence of interdependence between both recruitment mechanisms.
Rhi binds H3K9me3/H3K27me3 in a Kipf-independent context
To investigate whether Rhi binding is driven by H3K9me3/H3K27me3 dual-marked chromatin domains, we used S2 cells that lack an endogenous piRNA pathway30. Recent work showed that synthetic dual-chromatin reader domains for specific histone marks can be used to detect chromatin modifications, including combinatorial histone codes, through the use of different binders linked in tandem31. Rhi, like other HP1-family proteins, contains a CD that binds H3K9me3-modified histone tails6,14,15,32. We generated constructs to recognize H3K9me3 (CD2xCBX1), H3K27me3 (CD2xPc) or H3K9me3/H3K27me3-decorated domains (a bivalent CDPc-CBX1) (Fig. 4a and Extended Data Fig. 6a), and characterized the binding patterns of dual-CD constructs in S2 cells using CUT&RUN. We confirmed that CD2xCBX1 bound H3K9me3 (Fig. 4b,c), CD2xPc bound H3K27me3 (Fig. 4b,d and Extended Data Fig. 6b) and CDPc-CBX1 bound regions with both H3K9me3 and H3K27me3 (Fig. 4b,e).

a, A schematic showing the experimental workflow for the CD-binding assay. b, UCSC genome browser tracks of a 72 kb region on chromosome 4 displaying the binding profiles of the indicated histone modifications and CD constructs measured by CUT&RUN (cpm across pooled replicates). The dashed boxes indicate representative areas with either both H3K9me3 and H3K27me3, only H3K27me3 or only H3K9me3, respectively. The mappability of 100 nt reads is indicated below the tracks. c, A Venn diagram showing overlap between CD2xCBX1 binding, H3K9me3, H3K27me3 and CD2xRhi binding across 125,499 genomic 1 kb bins. Signal was considered to be present if the bin overlapped a MACS2 broad peak (q < 0.05, broad cutoff <0.1) present in at least two biological replicates. d, The same as c, but showing CD2xRhi binding, H2AK118ub, H3K27me3 and CD2xPc binding. e, The same as c, but showing CDPc-CBX1 and CD2xRhi binding, H3K27me3 and H3K9me3. f, A heat map and hierarchical clustering (Euclidean distance) of CUT&RUN signal detected for the indicated CD constructs and histone modifications (log10(FPKM)). The signal is shown as the mean across biological replicates. Individual replicates shown in Extended Data Fig. 6d. g, The same as f, but across a subset of the genome classified as heterochromatin (10,180 out of 125,499 1 kb bins). Individual replicates shown in Extended Data Fig. 6e. h, The same as c, but showing H3K9me3, CD2xRhi binding and H3K27me3. ORs for CD2xRhi binding to H3K9me3, H3K27me3, both or neither were calculated using Fisher’s exact test. See Extended Data Fig. 6f for all major overlaps. i, An overview of differentially bound regions (called using DiffBind, two-sided Wald test, Padj <0.05) following kipf or E(z) knockdown. Consensus peaks (n = 11,053) were called using DiffBind with MACS2 broad peaks from the indicated samples. j, Heat maps showing the log2-fold change of E(z) knockdown compared to control knockdown for the indicated constructs and histone modifications across Pc-CBX1 differentially bound regions.
Next, we tested whether the Rhi CD (CD2xRhi) recapitulated the bivalent H3K9me3/H3K27me3 reader CDPc-CBX1 (Fig. 4b,e). Strikingly, while CD2xRhi binding strongly resembled both that of CD2xCBX1 and the bivalent CDPc-CBX1 genome wide (Fig. 4f and Extended Data Fig. 6c,d), within heterochromatic regions only, CD2xRhi binding strongly correlated with H3K27me3 levels (Fig. 4g and Extended Data Fig. 6e). In contrast, tandem Rhi CDs containing F48A mutations in the aromatic cage (CD2xRhiF48A) essentially recapitulated the IgG control (Fig. 4b,f,g). Although CD2xRhi binding was strong already at loci with only H3K9me3 (odds ratio (OR) 4.2, Fisher’s exact test), the strongest CD2xRhi binding was observed at loci marked with both H3K9me3 and H3K27me3 (OR 9.3), whereas binding was depleted at loci only marked with H3K27me3 (OR 0.13) (Fig. 4h). Of note, regions marked by both H3K9me3 and H3K27me3 showed no presence of the PRC1 deposited mark H2AK118ub generally associated with the presence of H3K27me3 (ref. 25) (Fig. 4b). These results indicate that Rhi recruitment depends on H3K27me3 but is independent of H2AK118ub (Fig. 4b,d). Indeed, a binning analysis revealed that CD2xRhi binding clearly differs from that of CD2xCBX1 and CD2xPc through its ability to bind both clusters 80F and 42AB, which are strong CD2xCBX1 targets, and the CD2xPc-targeted 38C region (Extended Data Fig. 6g,h). Taken together, these results show that, similar to CDPc-CBX1, CD2xRhi preferentially binds to regions decorated with both H3K9me3 and H3K27me3.
To test whether CD2xRhi binding in S2 cells was mediated by Kipf, we repeated the CD experiments upon depletion of Kipf. Despite robust depletion of kipf (Extended Data Fig. 6i) and construct expression (Extended Data Fig. 6j), we did not observe differences in the binding patterns of any tested CD construct (Fig. 4i and Extended Data Fig. 6k–n). Importantly, upon kipf knockdown, CD2xRhi binding was indistinguishable from the control knockdown (Extended Data Fig. 6n), hence suggesting that binding by the Rhi CD in this assay is Kipf independent.
Kipf-independent Rhi binding depends on E(z)
To directly test the role of H3K27me3 in specifying Rhi CD binding, we performed CUT&RUN in S2 cells for CD2xCBX1, CD2xPc, CDPc-CBX1 and CD2xRhi upon control (siRen) or E(z) knockdown (siE(z)). Principle component analysis showed strong similarities between genome-wide CD2xPc binding and H3K27me3 distribution, and between H3K9me3 distribution and CD2xCBX1 binding (Fig. 4i and Extended Data Fig. 7a). Principle component analysis also confirmed, as we observed in earlier experiments (Fig. 4b and Extended Data Fig. 6c), that binding of CD2xRhi in control cells more closely resembles that of CDPc-CBX1 over CD2xCBX1 or CD2xPc (Extended Data Fig. 7a). Following E(z) knockdown, CD2xPc distribution was strongly affected, while CD2xCBX1 and H3K9me3 remained mostly unchanged (Fig. 4i). Notably, both CD2xRhi and CDPc-CBX1 distribution become more similar to that of CD2xCBX1 and H3K9me3 in the absence of E(z) (Extended Data Fig. 7a).
To further determine the impact of H3K27me3 on Rhi binding, we divided the genome into 1 kb bins, categorized into six groups based on the degree of H3K27me3 loss upon E(z) depletion. We then plotted for each group the change in binding of the different CDs as assessed by CUT&RUN in siE(z) compared with siRen. While H3K27me3 loss strongly reduced CD2xPc binding, no major genome-wide effects were observed for CD2xCBX1, CDPc-CBX1 or CD2xRhi (Extended Data Fig. 7b, top). This reflects the ability of CD2xPc to bind H3K27me3 genome wide, whereas only a subset also marked with H3K9me3 can be bound by CDPc-CBX1 and CD2xRhi. To identify H3K9me3 and H3K27me3 dual-marked regions affected by the E(z) loss, we focused on CD2xCBX1-bound regions (reflecting H3K9me3 levels) and observed that E(z) depletion resulted in reduced binding of both CD2xRhi and CDPc-CBX1. (Extended Data Fig. 7b, bottom). As expected, we observed no differences when performing the same analysis for siKipf (Extended Data Fig. 7c).
We next identified 460 CDPc-CBX1 binding sites where binding was significantly reduced (DiffBind, Padj <0.05) upon E(z) depletion. Notably, these regions displayed a similar loss of CD2xRhi binding, despite retaining comparable H3K9me3 levels (Fig. 4j and Extended Data Fig. 7d). These results indicate that binding of CDPc-CBX1 and CD2xRhi at these loci require both H3K27me3 and H3K9me3, and that H3K9me3 alone is insufficient. Taken together, these findings suggest that a combinatorial H3K9me3/H3K27me3 histone code, that is independent of Kipf but recognized by the Rhi CD, underlies Rhi binding at a subset of piSL.
The H3K9me3/H3K27me3 histone code at major piSL is conserved
The D. melanogaster and D. simulans Rhi CDs have similar binding specificities when expressed in D. melanogaster ovaries33. This is further supported by the high sequence conservation of the Rhi CD across Drosophila species32. In particular, aromatic residues in the CD, important for binding to methylated lysine, show high conservation across Drosophilids (Extended Data Fig. 8a). On the basis of this, we hypothesized that Rhi association with H3K9me3/H3K27me3 domains could be conserved.
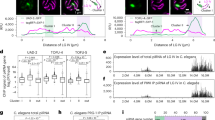
To test this, we assessed the presence of H3K9me3 and H3K27me3 at piSL in ovaries from D. simulans (most recent common ancestor (MRCA) 5.6 million years ago (Ma)), D. erecta (MRCA 8.2 Ma), D. yakuba (MRCA 11.4 Ma) and D. ananassae (MRCA 33.9 Ma) (Fig. 5a). We compared CUT&RUN data for H3K9me3 and H3K27me3 marks with available small RNA-seq of ovaries from these species4. We divided the genomes into 10 kb bins that were then classified into 15 groups according to their piRNA levels. We observed that at least for D. melanogaster and D. yakuba, bins with high levels of piRNAs also showed strong signals of H3K9me3 and H3K27me3, whereas the absence or low levels of piRNAs correlated mostly with the presence of H3K27me3 and absence of H3K9me3 (Fig. 5b,c and Extended Data Fig. 8b). Genome-wide analyses showed that piRNA producing loci are preferentially decorated with both H3K9me3 and H3K27me3 in D. simulans, D. yakuba, D. erecta and D. ananassae (Fig. 5d). We also detected both histone marks at the region of the D. simulans genome that is syntenic to 42AB in D. melanogaster (Fig. 5e). These results suggest that Rhi binding to H3K9me3/H3K27me3 regions is conserved in Drosophilids.

a, The phylogenetic tree of the five studied Drosophila species. b, Violin plots illustrating the distribution of piRNAs (purple), H3K9me3 (orange) and H3K27me3 (red) CUT&RUN signal, and Rhi (blue) ChIP–seq levels (log10(RPKM + 1)) in D. melanogaster ovaries: 10 kb bins across the genome were divided into 15 groups. Group 1 contains all 10 kb bins with no piRNA expression. The 14 remaining groups contain an equal number of 10 kb bins. Groups are ranked according to piRNA expression levels with the lowest level of the two strands shown to focus on dual-strand regions. The box plots show median (central line), interquartile range (box) and minimum and maximum values (whiskers, 1.5× the interquartile range at most). c, The same as b, but showing piRNA, H3K9me3 and H3K27me3 levels (log10(RPKM + 1)] for Drosophila yakuba ovaries. d, Line graphs showing mean piRNA abundance (n = 3 replicates; signal shown as the lowest of the two strands, log10(RPKM + 1)) plotted against H3K9me3 signal (orange, RPKM) and H3K27me3 signal (red) for all 10 kb bins of the indicated Drosophila species (D. melanogaster, D. simulans, D. yakuba, D. erecta and D. ananassae). e, UCSC genome browser tracks from D. simulans displaying ribo-depleted RNA-seq (light blue), piRNAs (purple), and CUT&RUN for H3K27me3 (red) and H3K9me3 (orange) as cpm at the 42AB-syntenic region (dashed box). The mappability of 100 nt reads is indicated below the tracks.
In conclusion, our study identifies the H3K27 methyltransferase E(z) as an important regulator of TE expression in Drosophila germ cells. We show that, in addition to the well-described H3K9me3 mark, H3K27me3 is important for guiding Rhi binding in a Kipf-independent manner, thus contributing to the definition of piSL.
Discussion
In this study, we provide evidence that co-occurrence of H3K9me3 and H3K27me3, usually thought to occupy distinct domains, plays an important role in specifying Rhi binding to piSL. By performing a screen for factors important for TE control, we identified the H3K27me3 methyltransferase E(z) and demonstrated that several Rhi-bound loci that produce vast amounts of piRNAs in germ cells are decorated with both H3K9me3 and H3K27me3 marks. We demonstrate that at these loci, Rhi recruitment is independent of Kipf and instead requires E(z), probably via its CD. These data demonstrate the existence of two independent modes of Rhi recruitment at distinct subsets of piSL: one that is reliant on the Kipf-recognized DNA sequence and one that is dependent on the chromatin context as established by E(z). Our results also point to the involvement of additional complexes in TE regulation, such as the COMPASS and NSL complex. Of note, a recent study has shown that the germline-specific depletion of subunits of the NSL complex (NSL1, NSL2 and NSL3) resulted in reduced piRNA production from telomeric, thus emphasizing the role of the NSL complex in piRNA precursor transcription within these loci34.
While previously suggested to be mutually exclusive, our work and that of others demonstrates that H3K9me3 and H3K27me3 coexist in a number of contexts35,36,37,38,39,40. This dual histone code has been reported in the ciliate Paramecium where it plays a role in TE regulation41. Furthermore, in plants and mammals, H3K27me3 serves as a backup for TE silencing, especially in contexts of low DNA methylation39,42,43,44,45,46. Interestingly, regions that give rise to 21U-RNAs in Caenorhabditis elegans are located within H3K27me3-rich genomic regions47 and recruitment of the Upstream Sequence Transcription Complex, required for 21U-RNA precursor transcription, is guided by the CD-containing protein UAD-2, which can bind H3K27me3 (ref. 48). The absence of H3K27me3 decouples UAD-2 from piSL, resembling the effect observed for Rhi upon germline depletion of E(z) in Drosophila.
Although E(z)-catalyzed H3K27me3 appears to be required for Rhi recruitment to a subset of piSL, the mechanisms by which E(z) is recruited and when H3K27me3 is deposited at specific Drosophila piSL during development remains to be determined. In Paramecium, deposition of H3K9me3 and H3K27me3 at TE insertions is mediated by the polycomb protein Ezl1 through interaction with a PIWI protein49. Notably, Drosophila E(z) has been shown to co-immunoprecipitate with Piwi in ovaries50, hinting at a possible role of PIWI proteins in guiding E(z) to piSL. Moreover, the presence of H3K27me3 at these loci in both germ cells and in S2 cells that are derived from the somatic lineage suggests that H3K27me3 is deposited maternally51 or established during early development, and retained as development progresses. A previous study demonstrated divergence in the spatiotemporal expression patterns of Kipf and Rhi, with Kipf exhibiting very low levels in ovarian germline stem cells and cystoblasts22. In light of this, we suggest that H3K27me3 may collaborate with H3K9me3 during the early stages of oogenesis, where Rhi functions independently of Kipf. Understanding how and when histone methyltransferases are guided to piSL is critical to better understand the piRNA system and its interplay with chromatin biology.
Our study shows that tandem Rhi CDs have a similar binding pattern as bivalent CD fusion proteins expected to bind H3K9me3/H3K27me3 domains, suggesting that the Rhi CD binds simultaneously to both H3K9me3/H3K27me3. Unlike the HP1a CD, the Rhi CD has been reported to form a homodimer in two crystallographic studies14,52. To investigate whether Rhi has the capacity to bind a histone H3 tail carrying dual-modified K9me3/K27me3, we successfully modeled the binding of a Rhi CD dimer to a 40 amino acid histone H3 tail peptide carrying both marks using molecular dynamics (MD) simulation (Extended Data Fig. 8c). However, recent size-exclusion chromatography studies failed to detect Rhi dimerization in solution23. Moreover, we were unable to obtain specific binding patterns in S2 cells using a construct carrying a single Rhi CD (CD1xRhi) (Extended Data Fig. 8d), possibly arguing against the ability of this construct to dimerize in vivo. However, the binding affinities of individual CD constructs were reported to be insufficient for efficient chromatin profiling31, hence complicating the interpretation of these results. While our work suggests a role for a dual histone code in determining Rhi binding, probably in a CD-dependent manner, we cannot exclude alternative mechanisms involving an additional co-factor that associates with H3K27me3 or E(z). Considering that previous work found that the Rhi CD does not bind H3K27me3 alone in vitro6,52, one interesting hypothesis is the presence of a yet-undiscovered Rhi co-factor that links it to H3K27me3. Interestingly, E(z) was recently identified as potential interactor of Deadlock in immunoprecipitation followed by mass spectrometry data from D. simulans, D. erecta and D. virilis53, another hypothesis could be that in D. melanogaster, this interaction is mediated by the Rhi CD directly. Alternatively, the presence of both H3K9me3 and H3K27me3 could provide a chromatin context (for example, chromatin compaction and/or repression of canonical transcription) more favorable to Rhi binding than H3K9me3 alone.
While bivalent chromatin, characterized by the presence of both H3K4me3 and H3K27me3, or dual domains have been previously suggested to play important roles in regulating gene expression, the diversity of chromatin reader domains able to interpret either each histone post-translational modification or exhibiting dual recognition of both modifications in vitro brings additional complexity to the understanding of chromatin biology54,55,56,57,58,59,60,61. In summary, our study provides an unexpected example highlighting in vivo the importance of a combinatorial histone code in the binding specificity of a chromatin-binding protein.
Methods
Fly husbandry and stocks
All flies were kept at 18 °C or 25 °C on standard cornmeal or propionic food. Flies were obtained from the Vienna Drosophila Resource Center or from the Bloomington Drosophila Stock Center (BDSC). All used fly stocks are listed in Supplementary Table 3. For germline-specific knockdowns, we used a fly line containing a UAS-Dcr2 transgene and a nos-GAL4 driver62, a pTOsk-GAL4 driver28 or a nos-GAL4 driver (BDSC:4937), each crossed to stocks expressing short hairpin (sh)RNAs63 or double-stranded (ds)RNAs64 under the UAS promoter. After mating at 27 °C for 5 days, parental flies were removed from the vials. Hatching F1 offspring were collected and aged with yeast for 2–3 days before use for follow-up experiments.
RNA isolation and qRT–PCR
RNA was extracted using TRIzol reagent (Thermo Fisher Scientific) according to the manufacturer’s instructions. One microgram of total RNA was treated with DNaseI (Thermo Fisher Scientific), and reverse transcribed using Superscript III (Thermo Fisher Scientific) and oligo(dT)20 primers. Primer sequences are listed in Supplementary Table 4.
TE screen
One to two different fly lines per gene (one replicate per line) expressing shRNAs or dsRNAs under the UAS promoter63,64 were crossed with a fly line containing a UAS-Dcr2 transgene and a nos-GAL4 driver, as previously described62. RNA was isolated from five to ten ovaries per cross and reverse transcribed as described above. Multiplexed qPCRs were carried out using TaqMan Universal Master Mix II, no UNG (Applied Biosystems) as described62. Experiments were performed on a CFX96 Real-Time System C1000 Touch Thermal Cycler (Bio-Rad). Z scores for TE expression were calculated on ΔCT values (CT(TE) – CT(rp49 control))65. The primers and probes used are listed in Supplementary Table 4. Positive hits were confirmed by qPCRs using SYBR green Master mix (Thermo Fisher Scientific). RT–qPCRs were performed on a QuantStudio Real-Time PCR Light Cycler (Thermo Fisher Scientific).
RNA-FISH
Single-molecule RNA-FISH for transcripts derived from piSL 42AB, 80F, 20A and TEs Burdock, Gypsy12 and copia was performed using Stellaris probes (Biosearch Technologies). Probe sequences are listed in Supplementary Table 4. Ovaries from 3–6-day-old flies were dissected in Schneider’s Drosophila medium and fixed in fixing buffer (4% formaldehyde, 0.3% Triton X-100 and 1× PBS) for 20 min at room temperature (RT), rinsed three times in 0.3% Triton X-100, once in PBS and permeabilized in 70% ethanol at 4 °C overnight. Permeabilized ovaries were rehydrated in wash buffer (10% formamide in 2× SSC) for 10 min. Ovaries were resuspended in 50 μl hybridization buffer (10% dextran sulfate and 10% formamide in 2× SSC) supplemented with 1.5 μl of probes. Hybridization was performed with rotation at 37 °C overnight. Ovaries were then washed twice with wash buffer at 37 °C for 30 min and twice with 2× SSC solution. Then, DNA was stained with 4,6-diamidino-2-phenylindole (DAPI) (1/500 dilution in 2× SSC) at RT for 20 min. Ovaries were mounted in 30 μl Vectashield mounting medium and imaged on a Zeiss LSM-800 confocal microscope. The resulting images were processed using Fiji/ImageJ (v1.54p).
Immunofluorescence
Fly ovaries were dissected in ice-cold PBS, fixed for 14 min in 4% PFA (Alfa Aesar) at RT and permeabilized with 3× 10 min washes in PBS with 0.3% Triton (PBS-Tr). Samples were blocked in PBS-Tr with 1% BSA for 2 h at RT and incubated overnight at 4 °C with primary antibodies in PBS-Tr and 1% BSA. After 3× 10 min washes at RT in PBS-Tr, secondary antibodies were incubated overnight at 4 °C in PBS-Tr and 1% BSA. After 4× 10min washes in PBS-Tr at RT with DAPI (Invitrogen) added during the third wash and 2× 5 min washes in PBS, samples were mounted with ProLong Diamond Antifade Mountant (Thermo Fisher Scientific) and imaged on a Leica SP8 or Zeiss LSM-800 confocal microscope. Images were deconvoluted using Huygens Professional or using Fiji/ImageJ (v1.54p). All used antibodies are listed in Supplementary Table 5.
Molecular cloning and constructs
CD sequences were amplified from cDNA prepared from ovaries or ordered as gBlock fragments from Integrated DNA Technologies and assembled using the NEBuilder HiFi DNA Assembly kit (New England Biolabs E2621) according to the manufacturer’s instructions. The final constructs expressed the CDs of interest tagged amino-terminally with an NLS-3xFlag-EGFP cassette under the control of the D. simulans ubiquitin promoter. A list and link to the sequence of all constructs used in this study are provided in Supplementary Table 6.
mRNA-seq (polyA selected)
Total RNA was extracted from 30 ovaries from 3–6-day-old flies using TRIzol (Thermo Fisher Scientific) in three replicates. One microgram of total RNA was subjected to polyA selection and subsequent fragmentation, reverse transcription and library preparation according to the manufacturer’s instructions using the Illumina stranded mRNA Prep for sequencing. Sequencing was performed by Novogene on an Illumina Novaseq 6000 instrument.
Ribo-depleted RNA-seq
Total RNA was extracted from 10–20 ovaries from 3–6-day-old flies using TRizol (Thermo Fisher Scientific) following the manufacturer’s instructions. For all the experiments conducted with the pTOsk-Gal4 driver, ribosomal RNA was depleted using RiboPOOL (siTOOLs, Biotech) as described66. RNA-seq libraries were produced using NEBNext Ultra Directional Library Prep kit for Illumina, following the manufacturer’s instructions for the rRNA depleted RNA. Library size distribution was analyzed on a TapeStation instrument (Agilent Technologies) using a High Sensitivity D1000 ScreenTape. Libraries were pooled in equal molar ratio and quantified with the KAPA Library Quantification kit for Illumina (Kapa Biosystems) and sequenced (paired-end 50) on an Illumina NovaSeq 6000 instrument. For the experiments conducted using the nos-Gal4d driver, rRNA depletion was performed from 1 µg total RNA using the RNA Depletion stranded Library Prep kit (BGI). The samples were then sequenced as paired-end 100 reads on the DNBSEQ G400 sequencer, and adapter-clipped reads were provided by BGI.
Small RNA-seq
Small RNA extraction was performed as described4,67. Argonaute-bound small RNAs were isolated from 10–30 pairs of ovaries from 3–5-day-old flies using TraPR columns (Lexogen, 128.08). Small RNA libraries were generated using the Small RNA-seq Library Prep kit (Lexogen, 052.96) according to the manufacturer’s instructions. For all experiments conducted with the pTOsk-Gal4 driver, ovaries of the five Drosophila species, and dKD or kipf knockdown experiments using the nos-Gal4 driver, sequencing was performed at the CRUK CI Genomics core on an Illumina NovaSeq 6000 instrument. Sequencing for the remaining experiments conducted using the nos-Gal4 driver was performed by Fasteris SA on an Illumina NextSeq550 instrument.
ChIP–seq
Chromatin immunoprecipitation was performed as previously described68 with minor modifications. Briefly, 100 ovary pairs were manually dissected into Schneider media and crosslinked in 1% formaldehyde/PBS for 10 min at RT with agitation. The crosslinking reaction was quenched by STOP buffer (PBS 1×, Triton 0.1% and glycine 1 M) and ovaries were washed in PBS and homogenized in a glass douncer: first slightly dounced in PBST 0.1% and centrifuged for 1 min at 400g, followed by strong douncing in cell lysis buffer (KCL 85 mM, HEPES 5 mM, NP-40 0.5%, sodium butyrate 10 mM and EDTA-free protease inhibitor cocktail (Sigma)) followed by 5 min centrifugation at 2,000g. We performed two washes with cell lysis buffer. The homogenates were then lysed on ice for 30 min in nucleus lysis buffer (HEPES 50 mM, EDTA 10 mM, N lauryl sarkosyl 0.5%, sodium butyrate 10 mM and EDTA-free protease inhibitor cocktail (Sigma)). DNA was sheared using a Bioruptor pico from Diagenode for 10 cycles (30 s on, 30 s off). The sonicated lysates were cleared by centrifugation and then incubated overnight at 4 °C with 5 µl of specific antibodies (Supplementary Table 5). Then 40 µl of Protein A Dynabeads was added and allowed to bind antibody complexes by incubation for 1 h at 4 °C. Following four washing steps with high salt buffer (Tris pH 7.5, 50 mM, NaCl 500 mM, Triton 0.25%, NP-40 0.5%, BSA 0.5% and EDTA pH 7.5, 5 mM), DNA–protein complexes were eluted and de-crosslinked for 10 h at 65 °C. RNA and protein were digested by RNase A and Proteinase K treatments, respectively, before purification using phenol:chloroform:isoamyl alcohol (25:24:1) (Sigma), according to the manufacturer’s instructions. Barcoded libraries were prepared using Illumina technology, and subsequently sequenced on a NextSeq High (Illumina) by Novogene (Rhi and H3K9me3 ChIP–seq), or by the Jean Perrin facility (H3K27me3 ChIP–seq).
CUT&RUN
One million cells per sample were collected and washed three times with wash buffer (20 mM HEPES, pH 7.5, 150 mM NaCl, 0.5 mM spermidine supplemented with protease inhibitors) and resuspended in 1 ml of wash buffer. Then, 10–20 fly ovaries were dissected in ice-cold PBS per sample. Ovaries were digested in 250 µl of dissociation buffer (0.5% Trypsin and 2.5 mg ml−1 collagenase A in PBS) for 1 h with shaking at 800 rpm at 30 °C. The digestion was stopped with 250 µl of Schneider medium containing 10% FBS. Cell suspensions were filtered through 40 µm strainers, pelleted and washed three times with wash buffer and resuspended in 1 ml of wash buffer. Following sample preparation, CUT&RUN was performed according to ref. 69 with some modifications. First, 10 µl of activated concanavalin A-coated magnetic beads (Bangs Laboratories) were added to each sample and rotated for 10 min at RT. Bead-bound cells were incubated with 5 µl of antibody (Supplementary Table 5) and 95 µl of antibody buffer (20 mM HEPES pH 7.5, 150 mM NaCl, 0.5 mM spermidine and 0.05% digitonin) overnight at 4 °C. Bead-bound cells were washed twice with Dig-wash buffer (0.05% digitonin in wash buffer). Bead-bound cells were resuspended in Dig-wash buffer containing 1× CUTANA pAG-MNase (Epicypher) and rotated for 1 h at RT. Following pAG-MNase binding, bead-bound cells were washed twice with Dig-wash buffer and resuspended in 100 µl Dig-wash buffer. Chromatin digestion was performed on ice for 30 min by adding 2 μl of 100 mM CaCl2. Digestion was stopped by the addition of 2× STOP buffer (340 mM NaCl, 20 mM EDTA, 4 mM EGTA, 0.05% digitonin, 100 µg ml−1 RNase A (Thermo Fisher Scientific) and 50 µg ml−1 glycogen) and samples were incubated at 37 °C for 30 min to release DNA fragments into the solution. After centrifugation, 0.1% SDS and 0.2 μg μl−1 Proteinase K were added to the supernatant and samples were incubated for 1 h at 50 °C. DNA was extracted using phenol:chloroform:isoamyl alcohol (25:24:1) (Sigma), according to the manufacturer’s instructions. Libraries were prepared following the manufacturer’s instructions with the NEBNext Ultra II DNA Library Prep kit for Illumina. Sequencing was performed on a NovaSeq 6000 instrument (Illumina).
Tissue culture, transfection and knockdowns
Drosophila Schneider 2 (S2) cells (Thermo Fisher Scientific, R69007) were cultured at 26 °C in Schneider’s Drosophila media (Gibco) supplemented with 10% heat-inactivated FBS (Sigma). S2 cells were transfected using the TransIT-Insect Transfection Reagent, using 2 million cells per transfection and 1 µg of plasmid DNA, and collected after 48 h. For knockdown experiments, two rounds of electroporation (48 h apart) were performed using the Cell Line Nucleofector kit V (Amaxa Biosystems; program G-030), as described70. CD constructs were cotransfected with 200 pmol of siRNA duplexes (oligo sequences are listed in Supplementary Table 4) during the second nucleofection round or transfected using the TransIT-Insect Transfection Reagent 24 h following the second nucleofection round. Cells were collected 5 days after the first nucleofection.
Western blot
Cell pellets and ovaries were lysed in RIPA buffer (Pierce) supplemented with protease inhibitors (Roche) and incubated for 30 min at 4 °C. The protein concentration of cleared lysates was quantified using a Direct Detect device (Merck). First, 20 µg total protein was separated on a NuPAGE 4–12% Bis–Tris denaturing gel (Thermo Fisher Scientific) and transferred to a nitrocellulose membrane using an iBLot2 dry transfer (Invitrogen). Primary antibody incubations were performed overnight at 4 °C and secondary antibodies were incubated for 1 h at RT. All antibodies and concentrations used are listed in Supplementary Table 5. Images were acquired using an Odyssey M Imaging system (LI-COR) and processed in Image Studio Lite (LI-COR).
Processing of RNA-seq
Adapters were removed using Trim Galore! (v0.6.4, --stringency 6) with additional parameters ‘-a CTGTCTCTTATA --clip_R1 1 --clip_R2 1 --three_prime_clip_R1 1 --three_prime_clip_R2 1’ if appropriate. The resulting reads were mapped to dm6 using STAR (v2.7.3a, --outMultimapperOrder Random --outSAMmultNmax 1 --outFilterMultimapNmax 1000 --winAnchorMultimapNmax 2000 --alignSJDBoverhandMin 1 --sjdbScore 3) and a genome index built using NCBI RefSeq. Gene expression was quantified using featureCounts (v1.5.3, -s 2 -O --largestOverlap -Q 50) with gene models from Ensembl (release 97) or annotations corresponding to each TE consensus sequences (RepBase, downloaded 20 April 2022).
Processing of sRNA-seq
The analyses included nos-driven E(z), kipf, w knockdown (two to three replicates each), nos-driven E(z)+w and E(z)+kipf double-knockdown (two replicates each), pTOsk-driven E(z), rhi or w knockdown (two replicates each), and previously published22. Gal4-driven kipf, rhi or w knockdown (one replicate each) and Kipf knockout, rhi knockout and w1118 flies (one replicate each). Adapters were removed using Trim Galore! (v0.6.4, --stringency 6 --length 18 --max_length 29 -q 0). For previously published small RNA-seq (sRNA-seq), we further used ‘-a’ to specify the adapter, and ‘--clip_R1’ and ‘--three_prime_clip_R1’ to remove non-standard adapters and random nucleotides at the read ends (see Supplementary Table 7 for details). The resulting reads were mapped to dm6 using Bowtie (v1.2.3, -S -n 2 -M 1 --best --strata --nomaqround --chunkmbs 1024 --no-unal). Gene expression was quantified using featureCounts (v1.5.3, -s 1 (or -s 2) -O --largestOverlap -Q 40) with gene models from Ensembl (release 97) or annotations corresponding to each TE consensus sequences (RepBase, downloaded 20 April 2022).
Differential gene expression and piRNA abundance analysis
RNA-seq differential expression analyses were performed using the DESeq2 (v1.26.0) package from R/Bioconductor. For gene expression, we first applied the DESeq2 function with default parameters followed by fold change shrinkage using the ‘ashr’ method. To analyze TE expression, we performed a similar analysis but used the size factors previously derived based on gene expression, while dispersion and fold change shrinkage were estimated based on both gene and TE expression. Reads per kilobase per million mapped reads (RPKM) values were calculated following the robust median implementation in DESeq2’s ‘fpm’ function. P values adjusted for multiple testing (Padj) were obtained using the Benjamini–Hochberg procedure. To consider a gene or TE to be differentially expressed, we required a fourfold change and Padj <0.05 unless otherwise specified.
sRNA-seq was analyzed in a similar way, except that the analysis was restricted to reads of length 23–30 nt mapping sense or antisense, respectively, to the TE consensus sequences. Size factors were estimated separately using the estimateSizeFactors function with siRNAs mapping antisense to annotated genes as input.
Processing of CUT&RUN data
CUT&RUN data were generated in one to four replicates per condition as specified in Supplementary Table 7. Sequencing adapters were removed using Trim Galore! (v0.6.4, --paired --stringency 6). The resulting reads were mapped to the dm6 reference genome using Bowtie (v1.2.3, -S -y -M 1 --best --strata --fr --minins 10 --maxins 600 --chunkmbs 2000 --nomaqround), reporting at most one hit for each read and considering insert sizes between 10 and 600 nt. PCR duplicates were removed using MarkDuplicates from Picard tools (v2.21.2). All the following analyses used the deduplicated data, except for the genome browser visualization.
Processing of ChIP–seq data
ChIP–seq reads were aligned to the dm6 genome using Bowtie2 (v2.4.2), with the alignment process set to report at most one hit for each read. In case of alignments with the same MAPQ score, the best alignment was randomly selected from among those equally scored alignments. Peaks were called using MACS2 (v2.2.7.1) to capture narrow (-q 0.05 -g dm) and broad peaks (-q 0.05 -g dm --broad --broad-cutoff 0.1). Only uniquely mapped reads were used for the peak calling. As a control we used Input for each condition.
Processing of publicly available ChIP–seq data
Some publicly available ChIP–seq libraries used for comparison to CUT&RUN were analyzed slightly differently. Specifically, HP1a ChIP–seq data from ovaries were downloaded from GEO (accession GSE140539)29. The libraries were paired-end 2× 101 nt and performed in two replicates. Rhi ChIP–seq samples from control, Rhi and Moon knockout ovaries and two ChIP–seq input samples were downloaded from GEO (accession GSE97719)7. The Rhi libraries were paired-end 2× 50 nt and performed in one replicate per condition. The input samples were 2× 100 nt and performed in two replicates. Rhi and Kipferl ChIP–seq samples from controls and Rhi and Kipf MTD-Gal4-mediated knockdown ovaries and corresponding input samples were downloaded from GEO (accession GSE202468)22. These data had variable read length (50, 74 or 100 nt) and were processed as single-end 50 nt. An overview of all ChIP–seq samples is available in Supplementary Table 7. Sequencing adapters were removed using Trim Galore! (v0.6.4 or v0.6.6, --paired --stringency 6), or alternatively ‘--hardtrim5 50’. The resulting reads were mapped to the dm6 reference genome using either Bowtie (v1.2.3, -S -y -M 1 --best --strata --fr --maxins 500 --chunkmbs 2000 --nomaqround), reporting at most one hit for each read. PCR duplicates were removed using MarkDuplicates from Picard tools (v2.21.2). All the following analyses used the deduplicated data, except for the genome browser visualization.
Peak calling using CUT&RUN or publicly available ChIP–seq data
Peaks were called using MACS2 to capture either narrow (callpeak -f BAMPE -g dm -q 0.01) or broad (callpeak -f BAMPE -g dm -q 0.05 --broad --broad-cutoff 0.1) peaks. Only uniquely mapped reads were used for the peak calling. As controls we used the corresponding IgG libraries for CUT&RUN, or input libraries for ChIP–seq, except for when Rhi ChIP–seq were compared with CUT&RUN and we used the Rhi knockout ChIP–seq as control. For histone modifications with two or more CUT&RUN replicates and CDs with three replicates, we derived a consensus peak set by first merging all peaks and then excluding peaks that were not supported by at least two replicates. For remaining CUT&RUN samples, we used all MACS2 peaks. Area-proportional Venn diagrams were created in R using the eulerr (v7.0.2) package to provide the best fit. Some low count overlaps might not be displayed due to geometric constraints. All overlaps are listed in the corresponding Source data files.
Binning analysis in Drosophila melanogaster
The binning analysis was done using either 1 kb or 10 kb bins. For 1 kb bins, the genome was divided into 144,916 non-overlapping 1 kb bins. We excluded bins with less than 20% mappability resulting in 125,519 mappable 1 kb bins. For 10 kb bins, the genome was divided into 29,918 bins using a 10 kb sliding window that moves 5 kb at a time. Bins at the chromosome ends with a size less than 5 kb were removed. We excluded bins with 20% mappability, resulting in 25,865 mappable 10 kb bins. Bins derived from mitochondria were further removed, producing a final set of 125,499 bins of size 1 kb and 25,862 of size 10 kb for CUT&RUN. When integrating data from multiple assays, selection of mappable bins was done with respect for the CUT&RUN libraries, and the same bins were used for RNA-seq and sRNA-seq analyses. For binning analysis including only sRNA-seq or RNA-seq, we re-calculated mappability using bowtie alignment with either 100 nt or 26 nt read lengths. CUT&RUN and publicly available ChIP–seq signal was quantified per 50 nt window using the bamCompare module from deepTools (v3.3.2, –binSize 50 --ignoreForNormalization chrM -p 4 --scaleFactorsMethod SES --extendReads --centerReads --exactScaling --minMappingQuality 255 -of bedgraph --operation subtract --pseudocount 0) using pooled IgG CUT&RUN samples as background, except for Rhi ChIP–seq where we used the rhi knockout ChIP–seq as background. Next, values below zero were set to zero, before the average normalized signal per window was calculated using bedtools map. RNA-seq and sRNA-seq signal was quantified in a stand-specific manner by converting uniquely mapped reads to BED format and counting the number of reads from each strand falling into a bin with at least half of their length (bedtools intersect, -c -F 0.5).
Genome browser visualization
For CUT&RUN, RNA-seq and sRNA-seq, we first counted the number of all and uniquely mapped reads, respectively, using samtools, per strand if applicable. Uniquely mapped reads were converted into bigWig files using the deepTools bamCoverage module (v3.3.2, --binSize 1 --ignoreForNormalization chrM --normalizeUsing CPM --exactScaling --scaleFactor s --skipNonCoveredRegions --minMappingQuality 255), where the scale factor, s, was set to the number of uniquely mapped reads divided by all mapped reads. Additionally, we used ‘--extendReads --centerReads’ for CUT&RUN samples to center the reads at fragments midpoints, ‘--filterRNAstrand’ for RNA-seq and sRNA-seq to separate the two strands, and ‘--minFragmentLength 23 --maxFragmentLength 30’ to select piRNAs for sRNA-seq. Uniquely mapped reads were converted into bigwig files using deepTools bamCoverage function (v3.5.0 --binSize 1 --ignoreForNormalisation ChrM --normalizeUzing CPM --extendReads --centerReads --skipNonCoveredRegions).
Assessing classification performance using AUC
Classification performance for individual histone marks or combinations was assessed using an ROC curve. Positive instances were defined as bins overlapping the cluster(s) of interest, with the remaining bins considered negative. The cumulative distribution was calculated in R using the ‘cumsum’ function with area under the curve calculated using the trapezoidal method.
Assessing Kipf dependency of Rhi peaks
Rhi ChIP–seq peaks were derived using MACS2 as described above. To focus on the most reliable binding sites, we merged peaks with at least threefold enrichment over background in individual control knockdown replicates into a high-confidence peak set. Any peak that was located to an unplaced contig or that was also present in rhi knockdown was excluded from the analysis. Finally, the remaining peaks were subdivided into Kipf-independent (n = 463) or Kipf-dependent (n = 1,166) peaks, depending on whether they overlapped a Rhi peak in Kipf knockdown or not.
Coverage plots at Rhi peaks
To visualize read coverage over Rhi peaks (Fig. 3g) we used the deepTools computeMatrix module (v3.3.2, reference-point --bin-size 50 -b 5000 -a 5000 --missingDataAsZero --reference-point center). The signal in the peak region was shown as difference between the knockdown and the control conditions.
Contribution of E(z) and Kipf, respectively, to piRNA production
Analysis was performed per 1 kb bin across the genome. First, we identified 632 bins where piRNA production depends on Rhi (>2-fold reduction in counts per million (cpm), across pTOsk-Gal4- or MTD-Gal4-driven knockdown, and rhi knockout; one to two replicates each). Next, we calculated the change in piRNA abundance in E(z)- or Kipf-depleted ovaries (nos-Gal4- or pTOsk-Gal4-driven knockdown of E(z), two replicates each; MTD-Gal4-driven knockdown of kipf or kipf knockout, one replicate each). Bins with >4-fold reduction in ovaries depleted for Kipf, E(z) or both were considered to be Kipf dependent, E(z) dependent or dependent on both.
Correlation heat maps
Heat maps were produced using the R package pheatmap (v1.0.12) with Euclidean distance.
Analysis of CUT&RUN in E(z) knockdown samples
To study the genome-wide effect of E(z) or kipf knockdown in S2 cells, we represented the genome as 117,300 1 kb bins and divided it into six equally sized groups based on the change in H3K27me3 signal upon E(z) loss. Four groups represented variable levels of H3K27me3 loss, one group represented no change and one group displayed a relative gain in H3K27me3 signal. Next, across each group, we calculated the change in 2xPc, 2xCBX1, 2xRhi and Pc-CBX1 binding affinity. Although 2xPc was strongly responsive to H3K27me3 loss, the other chromatin binders were largely unaffected. Hence, we next restricted the analysis to 46,583 bins across the six groups with H3K9me3 (defined as CD2xCBX1 above 90th percentile of euchromatic regions).
Binding affinity analysis using CUT&RUN data
Differential binding affinity analysis was performed using DiffBind (v3.8.4) applied on the MACS2 narrow or broad peaks from siRen and siEz samples, using the corresponding IgG libraries as controls. For the analysis, we used the dba, dba.count, dba.normalize, dba.contrast and dba.analyze modules with default options. This created a consensus set of peaks present in at least two samples, and then re-quantified the signal intensity at each consensus peak, and re-centered and re-sizeed the peaks to a 401 nt region around their maxima, resulting in 32,042 narrow or 11,053 broad consensus peaks. The contrasts were specified as siE(z) against siRen for each target (H3K9me3, H3K27me3, CD2xPc, CD2xCBX1, CDPc-CBX1 and CD2xRhi).
Euchromatin and heterochromatin coordinates
We used the following coordinates from Fabry and colleagues71 to define euchromatin (chr2R: 6460000–25286936, chr2L: 1–22160000, chr3L: 1–23030000, chr3R: 4200000–32079331 and chrX: 250000–21500000) and heterochromatin (chr2R: 1–6460000, chr2L: 22160000–23513712, chr3L: 23030000–28110227 and chr3R: 1–4200000) in D. melanogaster.
Reference genomes
We used the dm6 assembly for D. melanogaster downloaded from the UCSC genome browser and the following assemblies downloaded from NCBI: GCF_003285975 for D. annanassae, GCF_003286155 for D. erecta, GCF_016746395 for D. simulans and GCF_016746365 for D. yakuba.
Genome-wide mappability
To estimate genome mappability, we divided the genome into all possible n-mers, where n is either 26 (for sRNA) or 100 (for CUT&RUN). Those sequences were then mapped back onto the genome and mappability for each position was estimated as the number of reads overlapping a position, divided by n.
Analysis of CUT&RUN in five Drosophila species
CUT&RUN was performed in D. annanassae, D. melanogaster, D. erecta, D. simulans, and D. yakuba for H3K27me3 and H3K9me3 with two replicates per condition, followed by 2×50 nt paired-end sequencing. Sequencing adapters were removed using Trim Galore! (v0.6.4, --paired --stringency 6 -a GATCGGAAGAGCACACGTCTGAACTCCAGTCAC). The resulting reads were mapped to their respective reference genome using Bowtie (v1.2.3, -S -y -M 1 --best --strata --fr --minins 10 --maxins 600 --chunkmbs 2000 --nomaqround), reporting at most one hit for each read and considering insert sizes between 10 and 600 nt. One H3K9me3 replicate for D. yakuba displayed low complexity (1.3 million uniquely mapped reads with estimated library size 1.7 million) and was therefore excluded.
Analysis of sRNA-seq in five Drosophila species
Small RNA-seq was performed in D. annanassae, D. melanogaster, D. erecta, D. simulans and D. yakuba with three replicates per species. The libraries were sequenced as 2× 50 nt paired-end sequencing, but only the first read was used for the analysis. Trim Galore! (v0.6.4) was used to remove an abundant rRNA (--stringency 30 -a TGCTTGGACTACATATGGTTGAGGGTTGTA --length 18 -q 0) and sequencing adapters (--stringency 5 -a TGGAATTCTCGG --length 18 --max_length 35 -q 0). Next, Bowtie was used to exclude reads mapping to known Drosophila viruses (v.1.2.3, -S -M 1 --best --strata --nomaqround --chunkmbs 1024) using the ‘--max’ and ‘--un’ options to extract unmapped reads. These reads were mapped to their respective reference genome (-S -M 1 --best --strata --nomaqround --chunkmbs 1024), reporting at most one hit for each read.
Binning analysis in five Drosophila species
For the binning analysis, the genome was divided into either 1 kb non-overlapping bins or 10 kb bins with 5 kb overlap. To estimate the mappability within each bin, we divided the genome into all possible 100-mers and mapped those sequences back onto the genome. Mappability was calculated as the number of 100-mers mapping uniquely within each bin divided by the bin size. Bins with size <5 kb (for 10 kb bins) or with <20% mappability were excluded, leaving between 121,081 and 178,773 1 kb or between 24,687 and 38,112 10 kb bins per species. We next quantified the number of uniquely mapped CUT&RUN or sRNA-seq reads per library mapping ≥50% within each bin, considering the forward and reverse strand separately for the sRNA-seq data. To correct for differences in sequencing depth, mappability, and bin size, the resulting bin counts were normalized by their mappability and converted to fragments per kilobase of transcript per million mapped reads (FPKM) values. Bins from mitochondria were excluded from all analyses.
Scatter plots and violin plots in five Drosophila species
The normalized RPKM values were capped at 300 (or 200 for 10 kb bins) and log10-transformed using a pseudocount of 1. To focus on bins with piRNAs derived from both strands, we represented the sRNA-seq data as minimum signal on the forward and reverse strands. For H3K9me3 and H3K27me3, we used the mean of two replicates and for sRNA-seq we used the mean of three replicates. For the violin plots, we grouped the bins based on their piRNA level and displayed the piRNA, H3K27me3 or H3K9me3 level within each group. Briefly, we first constructed one group for bins with no piRNAs. Next, we sorted all remaining bins by piRNA level and extracted 15 equidistant breakpoints, including the lowest and highest piRNA level. Each pair of consecutive non-identical breakpoints were used to construct a piRNA level interval, resulting in between 11 or 14 additional groups per species (1 kb) or between 14 and 15 additional groups (10 kb). All scatter plots and violin plots were constructed in R (v3.6.2) using base graphics, imageScale from sinkr (v0.6), gridExtra and ggplot2.
Structure modeling of Rhi dimer using AlphaFold multimer
AlphaFold v2.3.2 Colab was used to generate a prediction model72. The full-length Rhi CD (amino acids 20–90) reported in the crystal structure of 4U68 was used as input twice to generate the multimer52. The structure of the Rhi CD dimer was found to resemble the original structure of 4U68 well52.
Structure modeling of histone 3 peptide using Zdock
Zdock was used to evaluate potential binding modes of the peptide to the dimer. To generate the structure of the input histone 3 peptide, the existing crystal structure of the K9me3- and K27me3-containing peptides in 4U68 and 1PFB were used52,73. Within the structure of 4U68, information for the structure of KQTARK(Me3)S, of the K9me3 section of the histone peptide was available. The structure of LATKAAR(Me3)SAP, of the K27me3 section of the histone peptide was available via 1PFB. A structure alignment was done using 4U68 and 1PFB via PyMOL, and the connecting chain of ‘TGGKAPRKQ’ was added using the Modeller tool in UCSF Chimera 1.16 (refs. 74,75). This peptide was then docked using Zdock server, with ZDOCK 3.0.2 (ref. 76) to yield a predictive model of the binding motif.
MD simulations
The structural prediction from AlphaFold multimer was used as a basis for MD simulations. MD simulations in explicit water were performed using the GPU accelerated code (PMEMD) of the Amber 16 and AmberTools 20 packages77,78. Protonation states were calculated using the PDB2PQR server. For the protein scaffold, an evolved version of the Stony Brook modification of the Amber 99 force field (ff14SB)79 parameters were applied (see below for additional non-standard parameters). TIP3P parameters were assigned to water molecules80. To incorporate the non-canonical amino acid K(Me3), into the AlphaFold model, the structure of the K(Me3) residue was used from 4U68 and the partial charges of the K(Me3) residue were set to fit the electrostatic potential generated at the HF/6-31 G(d) level by the restrained electrostatic potential model81. The charges were calculated according to the Merz–Singh–Kollman scheme using Gaussian82. Each protein complex was immersed in a pre-equilibrated cubic box with a 12 Å buffer of TIP3P water molecules using the Leap module. The systems were neutralized by the addition of explicit counterions (Na+ or Cl−). Long-range electrostatic effects were modeled using the particle mesh Ewald method with periodic boundary conditions83. An 8 Å cutoff was applied to Lennard-Jones and electrostatic interactions.
MD simulations were performed according to the following steps: (1) minimization with a maximum cycle of 5,000 and with the steepest descent algorithm for the first 2,500 cycles, with a periodic boundary for constant volume (canonical ensemble, NVT) and without the SHAKE algorithm activated. Positional restraints of 2 kcal mol−1 Å−2 were applied on heavy atoms of the protein backbone and heavy atoms of the ligand. (2) A 1 ns heating process was performed with a periodic boundary for constant volume (NVT) with SHAKE turned on such that the angle between the hydrogen atoms was kept fixed. Temperature increased from 0 to 300 K in a period of 1 ns with heat bath coupling with a time constant of 2 ps. Positional restraints of 2 kcal mol−1 Å−2 were applied on heavy atoms of the protein backbone. (3) A 2 ns equilibrium process was performed with a periodic boundary for constant volume (NVT) with SHAKE turned on such that the angle between the hydrogen atoms was kept fixed. An Andersen-like temperature coupling scheme is used to maintain the temperature at 300 K. (4) A 2 ns equilibrium process was performed with a periodic boundary for constant pressure (isothermal–isobaric ensemble, NPT) and with a constant temperature of 300 K, maintained using Langevin dynamics with the collision frequency of 5 ps−1. (5) A 100 ns equilibrium process was performed with a periodic boundary for constant pressure (NPT) and with a constant temperature of 300 K. (6) A 1,000 ns production was performed with a periodic boundary for constant pressure (NPT) and with a constant temperature of 300 K. A representative frame was obtained from 1 µS MD simulations using Chimera’s cluster analysis tool84.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Sequencing data generated in this study has been deposited to the GEO under accession (GSE247156). The following data were retrieved from GEO: RNA-seq, sRNA-seq and ChIP–seq data for Rhi and Kipf (accession GSE202468); HP1a ChIP–seq (GSE140542); Rhi ChIP–seq from control, Rhi, and Moon knockout ovaries (GSE97719); sRNA-seq data for Drosophila species (GSE225888). The dm6 genome assembly was downloaded from the UCSC genome browser and the following assemblies were downloaded from RefSeq: GCF_003285975, GCF_003286155, GCF_016746395 and GCF_016746365. Source data are provided with this paper.
Code availability
All analyses are described in Methods. No custom code was developed as part of this study.
References
Brennecke, J. et al. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 128, 1089–1103 (2007).
Czech, B. et al. piRNA-guided genome defense: from biogenesis to silencing. Annu. Rev. Genet. 52, 131–157 (2018).
Ozata, D. M., Gainetdinov, I., Zoch, A., O’Carroll, D. & Zamore, P. D. PIWI-interacting RNAs: small RNAs with big functions. Nat. Rev. Genet. 20, 89–108 (2019).
van Lopik, J. et al. Unistrand piRNA clusters are an evolutionarily conserved mechanism to suppress endogenous retroviruses across the Drosophila genus. Nat. Commun. 14, 7337 (2023).
Goriaux, C., Desset, S., Renaud, Y., Vaury, C. & Brasset, E. Transcriptional properties and splicing of the flamenco piRNA cluster. EMBO Rep. 15, 411–418 (2014).
Mohn, F., Sienski, G., Handler, D. & Brennecke, J. The rhino-deadlock-cutoff complex licenses noncanonical transcription of dual-strand piRNA clusters in Drosophila. Cell 157, 1364–1379 (2014).
Andersen, P. R., Tirian, L., Vunjak, M. & Brennecke, J. A heterochromatin-dependent transcription machinery drives piRNA expression. Nature 549, 54–59 (2017).
Zhang, Z. et al. The HP1 homolog rhino anchors a nuclear complex that suppresses piRNA precursor splicing. Cell 157, 1353–1363 (2014).
Klattenhoff, C. et al. The Drosophila HP1 homolog Rhino is required for transposon silencing and piRNA production by dual-strand clusters. Cell 138, 1137–1149 (2009).
Chen, Y. A. et al. Cutoff suppresses RNA polymerase II termination to ensure expression of piRNA precursors. Mol. Cell 63, 97–109 (2016).
Franke, A. et al. Polycomb and polyhomeotic are constituents of a multimeric protein complex in chromatin of Drosophila melanogaster. EMBO J. 11, 2941–2950 (1992).
Fischle, W. et al. Molecular basis for the discrimination of repressive methyl-lysine marks in histone H3 by Polycomb and HP1 chromodomains. Genes Dev. 17, 1870–1881 (2003).
Bernstein, E. et al. Mouse polycomb proteins bind differentially to methylated histone H3 and RNA and are enriched in facultative heterochromatin. Mol. Cell. Biol. 26, 2560–2569 (2006).
Le Thomas, A. et al. Transgenerationally inherited piRNAs trigger piRNA biogenesis by changing the chromatin of piRNA clusters and inducing precursor processing. Genes Dev. 28, 1667–1680 (2014).
Yu, B. & Huang, Y. Rhino defines H3K9me3-marked piRNA clusters. Oncotarget 6, 20740–20741 (2015).
ElMaghraby, M. F. et al. A heterochromatin-specific RNA export pathway facilitates piRNA production. Cell 178, 964–979.e20 (2019).
Kneuss, E. et al. Specialization of the Drosophila nuclear export family protein Nxf3 for piRNA precursor export. Genes Dev. 33, 1208–1220 (2019).
Hur, J. K. et al. Splicing-independent loading of TREX on nascent RNA is required for efficient expression of dual-strand piRNA clusters in Drosophila. Genes Dev. 30, 840–855 (2016).
Zhang, G. et al. Co-dependent assembly of Drosophila piRNA precursor complexes and piRNA cluster heterochromatin. Cell Rep. 24, 3413–3422.e4 (2018).
Zhang, G. et al. piRNA-independent transposon silencing by the Drosophila THO complex. Dev. Cell 56, 2623–2635.e5 (2021).
Pane, A., Jiang, P., Zhao, D. Y., Singh, M. & Schupbach, T. The Cutoff protein regulates piRNA cluster expression and piRNA production in the Drosophila germline. EMBO J. 30, 4601–4615 (2011).
Baumgartner, L. et al. The Drosophila ZAD zinc finger protein Kipferl guides Rhino to piRNA clusters. eLife 11, e80067 (2022).
Baumgartner, L. et al. Evolutionary adaptation of an HP1-protein chromodomain integrates chromatin and DNA sequence signals. eLife 13, RP93194 (2024).
Rangan, P. et al. piRNA production requires heterochromatin formation in Drosophila. Curr. Biol. 21, 1373–1379 (2011).
Margueron, R. & Reinberg, D. The Polycomb complex PRC2 and its mark in life. Nature 469, 343–349 (2011).
DeLuca, S. Z., Ghildiyal, M., Pang, L. Y. & Spradling, A. C. Differentiating Drosophila female germ cells initiate Polycomb silencing by regulating PRC2-interacting proteins. eLife 9, e56922 (2020).
Van Doren, M., Williamson, A. L. & Lehmann, R. Regulation of zygotic gene expression in Drosophila primordial germ cells. Curr. Biol. 8, 243–246 (1998).
ElMaghraby, M. F., Tirian, L., Senti, K. A., Meixner, K. & Brennecke, J. A genetic toolkit for studying transposon control in the Drosophila melanogaster ovary. Genetics 220, iyab179 (2022).
Zenk, F. et al. HP1 drives de novo 3D genome reorganization in early Drosophila embryos. Nature 593, 289–293 (2021).
Eastwood, E. L. et al. Dimerisation of the PICTS complex via LC8/Cut-up drives co-transcriptional transposon silencing in Drosophila. eLife 10, e65557 (2021).
Villasenor, R. et al. ChromID identifies the protein interactome at chromatin marks. Nat. Biotechnol. 38, 728–736 (2020).
Vermaak, D., Henikoff, S. & Malik, H. S. Positive selection drives the evolution of rhino, a member of the heterochromatin protein 1 family in Drosophila. PLoS Genet. 1, 96–108 (2005).
Parhad, S. S., Tu, S., Weng, Z. & Theurkauf, W. E. Adaptive evolution leads to cross-species incompatibility in the piRNA transposon silencing machinery. Dev. Cell 43, 60–70.e5 (2017).
Iyer, S. S. et al. The NSL complex is required for piRNA production from telomeric clusters. Life Sci. Alliance 6, e202302194 (2023).
Liu, T. et al. Broad chromosomal domains of histone modification patterns in C. elegans. Genome Res. 21, 227–236 (2011).
Ho, J. W. et al. Comparative analysis of metazoan chromatin organization. Nature 512, 449–452 (2014).
Mozzetta, C. et al. The histone H3 lysine 9 methyltransferases G9a and GLP regulate polycomb repressive complex 2-mediated gene silencing. Mol. Cell 53, 277–289 (2014).
Evans, K. J. et al. Stable Caenorhabditis elegans chromatin domains separate broadly expressed and developmentally regulated genes. Proc. Natl Acad. Sci. USA 113, E7020–E7029 (2016).
Walter, M., Teissandier, A., Perez-Palacios, R. & Bourc’his, D. An epigenetic switch ensures transposon repression upon dynamic loss of DNA methylation in embryonic stem cells. eLife 5, e11418 (2016).
Ichihara, S., Nagao, K., Sakaguchi, T., Obuse, C. & Sado, T. SmcHD1 underlies the formation of H3K9me3 blocks on the inactive X chromosome in mice. Development 149, dev200864 (2022).
Frapporti, A. et al. The Polycomb protein Ezl1 mediates H3K9 and H3K27 methylation to repress transposable elements in Paramecium. Nat. Commun. 10, 2710 (2019).
Mathieu, O., Probst, A. V. & Paszkowski, J. Distinct regulation of histone H3 methylation at lysines 27 and 9 by CpG methylation in Arabidopsis. EMBO J. 24, 2783–2791 (2005).
Weinhofer, I., Hehenberger, E., Roszak, P., Hennig, L. & Kohler, C. H3K27me3 profiling of the endosperm implies exclusion of polycomb group protein targeting by DNA methylation. PLoS Genet. 6, e1001152 (2010).
Deleris, A. et al. Loss of the DNA methyltransferase MET1 Induces H3K9 hypermethylation at PcG target genes and redistribution of H3K27 trimethylation to transposons in Arabidopsis thaliana. PLoS Genet. 8, e1003062 (2012).
Saksouk, N. et al. Redundant mechanisms to form silent chromatin at pericentromeric regions rely on BEND3 and DNA methylation. Mol. Cell 56, 580–594 (2014).
Rougee, M. et al. Polycomb mutant partially suppresses DNA hypomethylation-associated phenotypes in Arabidopsis. Life Sci. Alliance 4, e202000848 (2021).
Beltran, T. et al. Comparative epigenomics reveals that RNA polymerase II pausing and chromatin domain organization control nematode piRNA biogenesis. Dev. Cell 48, 793–810.e6 (2019).
Huang, X. et al. A chromodomain protein mediates heterochromatin-directed piRNA expression. Proc. Natl Acad. Sci. USA 118, e2103723118 (2021).
Miro-Pina, C. et al. Paramecium Polycomb repressive complex 2 physically interacts with the small RNA-binding PIWI protein to repress transposable elements. Dev. Cell 57, 1037–1052.e8 (2022).
Peng, J. C., Valouev, A., Liu, N. & Lin, H. Piwi maintains germline stem cells and oogenesis in Drosophila through negative regulation of Polycomb group proteins. Nat. Genet. 48, 283–291 (2016).
Zenk, F. et al. Germ line-inherited H3K27me3 restricts enhancer function during maternal-to-zygotic transition. Science 357, 212–216 (2017).
Yu, B. et al. Structural insights into Rhino-mediated germline piRNA cluster formation. Cell Res. 25, 525–528 (2015).
Riedelbauch, S. et al. Recurrent innovation of protein–protein interactions in the Drosophila piRNA pathway. EMBO J. https://doi.org/10.1038/s44318-025-00439-8 (2025).
Azuara, V. et al. Chromatin signatures of pluripotent cell lines. Nat. Cell Biol. 8, 532–538 (2006).
Mikkelsen, T. S. et al. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 448, 553–560 (2007).
Gaertner, B. et al. Poised RNA polymerase II changes over developmental time and prepares genes for future expression. Cell Rep. 2, 1670–1683 (2012).
Kumar, D., Cinghu, S., Oldfield, A. J., Yang, P. & Jothi, R. Decoding the function of bivalent chromatin in development and cancer. Genome Res. 31, 2170–2184 (2021).
Barral, A. et al. SETDB1/NSD-dependent H3K9me3/H3K36me3 dual heterochromatin maintains gene expression profiles by bookmarking poised enhancers. Mol. Cell 82, 816–832.e12 (2022).
Musselman, C. A., Lalonde, M. E., Cote, J. & Kutateladze, T. G. Perceiving the epigenetic landscape through histone readers. Nat. Struct. Mol. Biol. 19, 1218–1227 (2012).
Mashtalir, N. et al. Chromatin landscape signals differentially dictate the activities of mSWI/SNF family complexes. Science 373, 306–315 (2021).
Du, J. et al. Dual binding of chromomethylase domains to H3K9me2-containing nucleosomes directs DNA methylation in plants. Cell 151, 167–180 (2012).
Czech, B., Preall, J. B., McGinn, J. & Hannon, G. J. A transcriptome-wide RNAi screen in the Drosophila ovary reveals factors of the germline piRNA pathway. Mol. Cell 50, 749–761 (2013).
Ni, J. Q. et al. A genome-scale shRNA resource for transgenic RNAi in Drosophila. Nat. Methods 8, 405–407 (2011).
Perkins, L. A. et al. The Transgenic RNAi Project at Harvard Medical School: resources and validation. Genetics 201, 843–852 (2015).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408 (2001).
Munafo, M. et al. Channel nuclear pore complex subunits are required for transposon silencing in Drosophila. eLife 10, e66321 (2021).
Grentzinger, T. et al. A universal method for the rapid isolation of all known classes of functional silencing small RNAs. Nucleic Acids Res. 48, e79 (2020).
Lee, T. I., Johnstone, S. E. & Young, R. A. Chromatin immunoprecipitation and microarray-based analysis of protein location. Nat. Protoc. 1, 729–748 (2006).
Meers, M. P., Bryson, T. D., Henikoff, J. G. & Henikoff, S. Improved CUT&RUN chromatin profiling tools. eLife 8, e46314 (2019).
Batki, J. et al. The nascent RNA binding complex SFiNX licenses piRNA-guided heterochromatin formation. Nat. Struct. Mol. Biol. 26, 720–731 (2019).
Fabry, M. H. et al. Maternally inherited piRNAs direct transient heterochromatin formation at active transposons during early Drosophila embryogenesis. eLife 10, e68573 (2021).
Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583–589 (2021).
Min, J., Zhang, Y. & Xu, R. M. Structural basis for specific binding of Polycomb chromodomain to histone H3 methylated at Lys 27. Genes Dev. 17, 1823–1828 (2003).
Fiser, A. & Sali, A. Modeller: generation and refinement of homology-based protein structure models. Methods Enzymol. 374, 461–491 (2003).
Pettersen, E. F. et al. UCSF Chimera-a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Pierce, B. G. et al. ZDOCK server: interactive docking prediction of protein-protein complexes and symmetric multimers. Bioinformatics 30, 1771–1773 (2014).
Case, D. A. et al. Amber 2020 (Univ. California, 2020).
Case, D. A. et al. AMBER 2016 Reference Manual 1–923 (Univ. California, 2016).
Maier, J. A. et al. ff14SB: improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 11, 3696–3713 (2015).
Jorgensen, W. L., Chandrasekhar, J., Madura, J. D., Impey, R. W. & Klein, M. L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 79, 926–935 (1983).
Bayly, C. I., Cieplak, P., Cornell, W. & Kollman, P. A. A well-behaved electrostatic potential based method using charge restraints for deriving atomic charges: the RESP model. J. Phys. Chem. 97, 10269–10280 (1993).
Besler, B. H., Merz, K. M. Jr & Kollman, P. A. Atomic charges derived from semiempirical methods. J. Comput. Chem. 11, 431–439 (1990).
Darden, T., York, D. & Pedersen, L. Particle mesh Ewald: an N⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 98, 10089–10092 (1993).
Kelley, L. A., Gardner, S. P. & Sutcliffe, M. J. An automated approach for clustering an ensemble of NMR-derived protein structures into conformationally related subfamilies. Protein Eng. 9, 1063–1065 (1996).
Acknowledgements
We thank members of the Brasset and Hannon groups for fruitful discussions. We thank S. Jensen for her help with preliminary RNA-seq analysis. We are grateful to L. Baumgartner and J. Brennecke for sharing antibodies, unpublished data and for scientific discussion. We thank M. Bao from the Life Science Editors for feedback and comments on the paper. We thank the BDSC and the Vienna Drosophila Resource Center for the supply of fly stocks. We thank the Scientific Computing, Genomics, RICS and microscopy core facilities at CRUK Cambridge Institute and the CLIC facility at iGReD for support. G.J.H. is a Royal Society Wolfson Research Professor (RSRP\R\200001). This research was funded in whole, or in part, by Cancer Research UK (grant no. G101107 to G.J.H.; grant nos C9685/A26398, C9685/A27415 and C9545/A29580 to P.C.) and the Wellcome Trust (grant no. 110161/Z/15/Z to G.J.H.; grant no. 226627/Z/22/Z to G.J.H., B.C.N. and S.B.). Work in the Brasset lab was funded by the Agence Nationale pour la Recherche (grant no. CHApiTRE ANR-20-CE12-0005 to E.B., grant no. BiopiC ANR-21-CE12-0022 to E.B., and grant no. EpiTET 699 ANR-17-CE12-0030-03 to E.B.), the La Fondation ARC pour la recherche sur le cancer (grant no. PJA20171206129 to E.B.). This research was financed also by the French government IDEX-ISITE initiative 16-IDEX-0001 (grant no. CAP 20-25 to E.B.). The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
A.A. and E.K. performed all experiments except those stated below. S.B. performed all computational analyses except ChIP–seq analysis, and initial RNA-seq and small RNA-seq analyses, which were performed by Y.R. and S.M.-M., and the MD simulations/modeling, which was performed by M.J. under the supervision of P.C. E.L.E. performed knockdown experiments using pTOsk-Gal4 and generated RNA-seq libraries, J.v.L. generated small RNA-seq libraries. E.K., A.S. and Y.G. performed CUT&RUN from CD constructs in S2 cells, and Y.G. performed western blot analysis. N.G. generated libraries for RNA-seq and prepared probes for RNA-FISH. The project was conceived by A.A., E.K., B.C.N., E.B. and G.J.H., and supervised by B.C.N., E.B. and G.J.H. The data were analyzed and interpret by A.A., E.K., S.B., B.C.N., E.B. and G.J.H. The paper was written by A.A., E.K., S.B., B.C.N., E.B. and G.J.H. with input from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Structural & Molecular Biology thanks Julius Brennecke and Jindan Yu for their contribution to the peer review of this work. Peer reviewer reports are available. Primary Handling Editor: Sara Osman, in collaboration with the Nature Structural & Molecular Biology team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Summary of screen results.
Heatmaps summarising gene names, fertility and ovary morphology phenotypes, as well as TE (hetA, blood, burdock, tahre) and germline marker (nos, yTub) expression, measured by RT-qPCR, upon indicated germline knockdowns (displayed as z-scores). Known piRNA factors and proteins that are part of the COMPASS, PRC2 and NSL complexes are highlighted as indicated in the legend.
Extended Data Fig. 2 E(z)-depleted ovaries display piRNA loss and TE derepression.
a, Immunofluorescence staining showing Rhi expression and localisation in nurse cell nuclei of stage 4 egg chambers upon the indicated shRNA and Gal4 driver combination (scale bar: 5 μm). Data are representative of n > =3 independent experiments. b, Ovary morphology image (left) and confocal images of egg champers stained with anti-H3K27me3 (right) upon the indicated knockdowns and drivers (scale bars: 10 µm). Data are representative of n > =3 independent experiments. c, MA plot showing counts per gene (grey) and TEs (black) in polyA-selected RNA-seq libraries from ovaries of MTD-Gal4 mediated germline knockdown of rhi versus control. TEs with padj<0.05 and >4x fold change are shown in red (DESeq2, two-sided Wald test). d, Confocal images showing RNA-FISH signal for transcripts derived from piRNA clusters 42AB, 80F and 20A in control and E(z) depleted ovaries using the pTOsk-Gal4 driver (scale bar: 5 μm). Data are representative of n > =3 independent experiments. e, Scatter plot depicting normalized ribo-depleted RNA levels (fpkm) of uniquely mapping reads in 1 kb bins in ovaries with pTOsk-Gal4 driven E(z) knockdown versus control (average of four replicate experiments each). f, Bar graphs showing levels of the indicated piRNA precursor transcripts measure using RT-qPCR upon the indicated germline knockdown. Data are presented as the mean (n = 3) with error bars representing standard deviation. Statistical significance was determined using the unpaired t-test. ** corresponds to P-value < 0.01, * corresponds to P-value < 0.05, and ns corresponds to p > 0.05. g, same as e but showing pTOsk-Gal4 driven rhi knockdown versus control (average of four replicate experiments each). h, Scatter plot depicting normalized piRNA levels of uniquely mapping piRNAs in 1 kb bins in ovaries with pTOsk-Gal4 driven E(z) knockdown versus control (two replicates each). i, Scatter plot showing TE transcript fold-change against antisense piRNA fold-change in nos-Gal4 driven E(z) knockdown versus control. Filled triangles indicate TEs that change on both mRNA and piRNA level (DESeq2, two-sided Wald test, padj<0.05). j, Profile plots of piRNA reads over selected TE consensus sequences (burdock, copia, gypsy12 and gypsy). piRNA counts (normalized to miRNAs) are displayed for indicated genotypes.
Extended Data Fig. 3 Rhi colocalises with H3K9me3 and H3K27me3 in germ cells.
a, Genome-wide view of TE content across D. melanogaster chromosomes with TE fraction (grey), HP1a (beige) ChIP-seq (Zenk et al. 2021), H3K27me3 (red) and H3K9me3 (yellow) CUT&RUN and piRNA levels (purple) in wildtype ovaries. CUT&RUN was normalized to IgG signal per 50 nt (see Methods for details). b, UCSC genome browser tracks of ChIP-seq signal (coverage per million reads) of Rhi (blue), H3K27me3 (red), and H3K9me3 (yellow), and uniquely mapping piRNAs (purple, normalized to miRNA reads) at the sox102F locus in nos-Gal4>sh-w ovaries. c, Venn diagram showing overlap between Kipf and H3K27me3 ChIP-seq signal across genomic 1 kb bins. Signal was considered to be present if the bin overlapped a MACS2 broad peak (q < 0.05, broad-cutoff<0.1) across two pooled biological replicates. d, Schematic showing how predictive performance was assessed in a threshold-independent way using an area under ROC curve metric. First, all genomic 1 kb bins were ranked according to signal strength (for example, highest to lowest H3K27me3). Second, for every possible rank threshold, true positive rate (TPR) is calculated as number of bins of interest (for example, located within a dual-strand piRNA cluster) above the threshold divided by the total number of bins of interest. Similarly, false positive rate (FPR) is calculated as the number of other bins (for example, located outside of a dual-strand piRNA cluster) above the threshold, divided by the total number of other bins. Third, TPR is plotted against FPR and the area under curve (AUC) is calculated. A higher metric indicates more predictive power and AUC at 0.5 indicate performance expected by random guessing. e, Line graphs comparing the ability to identify dual-strand piRNA clusters based on the strength of individual features (H3K27me3 and HP1a) or combinations.
Extended Data Fig. 4 Rhi binding at Kipf-independent loci depends on E(z).
a, Box plots showing Rhi ChIP-seq enrichment per E(z)/Kipf-dependency category. *** corresponds to P < 0.001, ns corresponds to p > 0.05 based on Wilcoxon signed-rank test. b, UCSC genome browser tracks of the sox102F locus displaying profiles of small RNAs uniquely mapping in control, E(z)-depleted and Rhi-depleted ovaries and ChIP-seq signal for Rhi and H3K9me3 in control and E(z)-depleted ovaries. c, UCSC genome browser tracks of ChIP-seq signal for Rhi and H3K9me3 in control and E(z)-depleted ovaries of the 42AB dual-strand piRNA cluster. Dashed line indicates approximate piRNA cluster boundaries. d, Violin plots (left) and box plots (right) showing average log2-fold H3K27me3 enrichment by ChIP-seq from ovaries with nos-Gal4 driven E(z) knockdown versus control (average of two replicate experiments each) in heterochromatin (HC) and euchromatic chromosome arms (EC), quantified across 1 kb bins (excluding piRNA clusters). H3K27me3 occupancy at indicated piRNA clusters is shown as box plot quantification (n depicts 1 kb bins analyzed for each piRNA cluster). *** corresponds to P < 0.001, ns corresponds to p > 0.05 based on Wilcoxon signed-rank test. Box plots show median (centre line), with interquartile range (box) and whiskers indicating at most 1.5x interquartile range). e, Heatmap of H3K9me3 ChIP-seq signal across wildtype H3K9me3 peaks (n = 4,785) in control and E(z) knockdown ovaries. f, Metaplot showing mean difference in ChIP-seq signal for Kipf (orange) in rhi knockdown, and Rhi (light blue) in kipf knockdown across Rhi peaks categorised as either Kipf-dependent or not (see Methods). g, Metaplot showing mean difference in ChIP-seq signal for Rhi in moon (purple) knockout or E(z) knockdown (yellowish green) across Rhi peaks categorised as either Kipf-dependent or not (see Methods).
Extended Data Fig. 5 Rhi binding upon E(z)-kipf double knockdown.
a-d, MA plot showing counts per TE (black) in sRNA-seq libraries from ovaries of germline (a) E(z) KD versus control, (b) E(z)-kipf dKD versus control, (c) E(z)-kipf dKD versus E(z) KD, and (d) E(z)-kipf dKD versus kipf KD. Sense and antisense piRNAs are shown separately and TEs with padj<0.05 and >4x fold change are shown in red (DESeq2, two-sided Wald test). e, Boxplots showing changes in piRNAs mapping uniquely to genomic 1 kb bins within heterochromatin, euchromatin or indicated piRNA clusters (n refers to the number of 1 kb bins for each category). Signal shown as a log2-ratio between the indicated KD and its respective control (average of 1-3 biological replicates). P-values were calculated using a two-sided paired Wilcoxon signed-rank test for >2-fold changes. Boxplots show median (central line), interquartile range (IQR, box), and minimum and maximum values (whiskers, at most 1.5*IQR). f, Scatter plot depicting normalized uniquely mapping piRNA levels in 1 kb bins in ovaries with nos-Gal4 driven E(z) knockdowns versus control (average of two replicates each). g, Scatter plot depicting normalized uniquely mapping piRNA levels in 1 kb bins in ovaries with nos-Gal4 E(z)-Kipf dKD compared to nos-Gal4 driven w knockdown (average of two replicates each). h, Same as d but comparing E(z)-kipf dKD to nos-Gal4 driven E(z)-w dKD (average of two replicates each). i, Same as d but comparing E(z)-kipf dKD to nos-Gal4 driven kipf knockdown (average of two or three replicates each). j, Immunofluorescence staining showing Rhi expression and localisation in nurse cell nuclei upon the indicated shRNA and nos-Gal4 driver combination (scale bar: 5 μm). Data are representative of n > =3 independent experiments.
Extended Data Fig. 6 Rhi chromodomains associate with regions co-occupied by H3K9me3 and H3K27me3.
a, Schematic showing the experimental workflow and cartoon of the dual-CD constructs used in the assay. b, Venn diagram showing overlap between CD2xPc binding, H3K27me3, H3K9me3, and Rhi across 125,499 genomic 1 kb bins. Signal was considered to be present if the bin overlapped a MACS2 broad peak (q < 0.05, broad-cutoff<0.1) present in at least two biological replicates. c, Scatter plot depicting CUT&RUN signal (fpkm) of uniquely mapping reads in 1 kb bins of CD2xRhi versus CDPc-CBX1 in S2 cells (average of 2-3 replicate experiments each). d, Heatmap and hierarchical clustering (Euclidean distance) of CUT&RUN signal detected for the indicated chromodomain constructs and histone modifications [log10(fpkm)]. e, Same as d, but across a subset of the genome classified as heterochromatin (10,180 out of 125,499 1 kb bins). f, UpSet plot showing all major intersections ( > 100 1 kb bins) across the six indicated CUT&RUN experiments. g, as in c but showing CD2xRhi versus CD2xPc (average of 2 replicate experiments each). h, as in c but showing CD2xRhi versus CD2xCBX1 (average of 2 replicate experiments each). i, Western blot analyses showing Kipf expression in S2 cells (treated as indicated) or w1118 ovaries. Tubulin is shown as loading control (experiment was repeated twice with similar results). j, Western blot analyses showing the levels of the indicated chromodomain constructs (Flag) in S2 cells that were transfected with the corresponding plasmids. Tubulin is shown as loading control (similar results were obtained in two independent experiments). k, Scatter plot depicting CUT&RUN signal (fpkm) of uniquely mapping reads in 1 kb bins of CD2xCBX1 upon control or kipf knockdown in S2 cells (1 replicate experiment).l-n, as in k but depicting CD2xPc, CDPc-CBX1, and CD2xRhi, respectively (2-3 replicate experiments each, as indicated).
Extended Data Fig. 7 CD2xRhi binding in S2 cells depends on E(z).
a, Principal component analysis (PCA) plot of the binding affinities of the indicated chromodomain fusion proteins expressed in S2 cells in control condition (circle/triangle) and E(z) knockdown (arrowhead). One CD2xRhi sample without arrowhead lacks a corresponding knockdown sample. b, Box plot showing loss of binding of the indicated chromodomain fusion proteins in S2 cell upon E(z) knockdown compared to control knockdown (siRen). Shown are log10 fold change per 1 kb bin either genome wide (top, n = 117,300 bins with a non-zero change) or at CD2xCBX1-bound regions (bottom, n = 46,583 bins, CD2xCBX1 above 90th percentile of euchromatic regions). The genome-wide bins were divided into six groups according to the severity of H3K27me3 loss (higher to lower, thresholds indicated to the right). Boxplots show median (central line), interquartile range (IQR, box), and minimum and maximum values (whiskers, at most 1.5*IQR). c, Same as a, but for kipf knockdown (siKipf) compared to control knockdown (siRen). d, Heatmaps showing the indicated chromodomain fusion protein and histone modification CUT&RUN signal in the 4 kb surrounding CDPc-CBX1 differentially bound peaks in control (siRen) and E(z) knockdowns.
Extended Data Fig. 8 Dual-strand piRNA producing loci are associated with both H3K9me3 and H3K27me3 in Drosophilids.
a, Protein sequence alignment of the chromodomains of Rhi orthologs across the indicated species. Residues forming the hydrophobic pocket for trimethyl lysine binding are indicated in red. Residues important for chromodomain dimerization are indicated in green. b, Violin plots illustrating the distribution of piRNAs, H3K9me3 and H3K27me3 CUT&RUN signal [log10(rpkm+1)] in D. simulans, D. erecta, and D. ananassae ovaries. All 10 kb bins across the genome were divided into 15 groups. Group 1 contains all 10 kb bins with no piRNA expression. The 14 remaining groups contain an equal number of 10 kb bins. Groups are ranked according to piRNA expression levels with the lowest level of the two strands shown to focus on dual-strand regions. Boxplots show median (central line), interquartile range (IQR, box), and minimum and maximum values (whiskers, at most 1.5*IQR). c, Model of the Rhi chromodomain dimer in complex with a histone 3 peptide with trimethylated K9 and K27 residues. Representative frame obtained from 1 µS molecular dynamic simulations show the potential for the dimer’s interaction with both H3K9me3 and H3K27me3 to be simultaneous. Zoom-in indicated aromatic residues that interact with the methylated lysine group. d, UCSC genome browser tracks of D. melanogaster chromosome 4 displaying the binding profiles of the indicated chromodomain constructs measured by CUT&RUN (counts per million [cpm] across pooled replicates). Gene tracks are shown above.
Supplementary information
Supplementary Information (download PDF )
Supplementary Notes 1 and 2.
Supplementary Tables 1–7 (download XLSX )
File containing all Supplementary Tables (in separate tabs) related to the figures and the methods.
Source data
Source Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 6 (download PDF )
Uncropped western blots.
Source Data Extended Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 8 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 8 (download ZIP )
Pdb files related to the model of the Rhi chromodomain dimer in complex with a histone 3 peptide with trimethylated K9 and K27 residues.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Akkouche, A., Kneuss, E., Bornelöv, S. et al. Binding of heterochromatin protein Rhino to a subset of piRNA clusters depends on a combination of two histone marks. Nat Struct Mol Biol 32, 1517–1527 (2025). https://doi.org/10.1038/s41594-025-01584-8
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41594-025-01584-8
This article is cited by
-
The H3K27me3 reader UAD-2 recruits a TAF-12-containing transcription machinery to initiate piRNA expression within heterochromatic clusters
Nature Communications (2025)
-
H3K27me3 expands the epigenetic control of piRNA transcription
Nature Structural & Molecular Biology (2025)
