
Abstract
Population sizes of invasive species are commonly characterized by boom-bust dynamics, and self-limitation via resource depletion is posited as one factor leading to these boom-bust changes in population size. Yet, while this phenomenon is well-documented in plants, few studies have demonstrated that self-limitation is possible for invasive animal species, especially those that are mobile. Here we examined the invasive Asian shore crab Hemigrapsus sanguineus, a species that reached very high abundances throughout invaded regions of North America, but has recently declined in many of these same regions. We examined the relationship between diet, energy storage, reproduction, and growth in crabs collected from the New Hampshire coast. We show that energy storage and reproduction both increase with diet quality, while growth declines with diet quality. These results suggest that self-limitation may be a contributing factor to the recent declines of H. sanguineus at sites where this invader was once much more abundant. Further, these results suggest a diet-associated tradeoff in energy allocation to different vital rates, with a focus on reproduction when high quality resources are consumed, and a focus instead on growth when poor quality resources are consumed.
Similar content being viewed by others

Introduction
The concept that no population can increase forever and that something limits all populations from growing without bounds is a fundamental maxim of ecology1. Limits to population growth can occur via numerous mechanisms, including density-dependent predation2, interspecific and intraspecific competition for limiting resources3,4, and density-dependent spread of disease5. Self-limitation occurs via a subset of regulatory mechanisms where endogenous factors limit individual performance or survival, and these individual limitations in turn restrict population growth. The concept of self-limitation has been posited as one of the fundamental “laws” of population ecology6. Self-limitation can have important ecological consequences that extend beyond prohibiting runaway population growth, including facilitating the persistence of rare species in communities7 and shaping the evolution of life-history traits8.
Self-limitation can occur across a wide range of systems and through various processes, but often has resource limitation at its roots. Resource limitation can occur because of several factors, including territorial defense that limits the sharing of resources9, environmental variation in resource availability10, and competition for resources among conspecifics11. This resource limitation in turn alters vital rates so that under food-limited conditions, growth rates12, survival rates13, and reproductive rates14 all decline. These changes in vital rates may also interact. As increased food availability can lead to increased fecundity via more rapid growth rates, because larger body sizes in females can support the production of more eggs15. In addition to the amount of food, diet quality can also alter vital rates16,17. This can occur because of life history tradeoffs that set in when resources are limited18, where energy or nutrient shortfalls limit the ability of individual organisms to simultaneously achieve optimal performance in survival, growth, and reproduction. All these examples demonstrate mechanisms through which self-limitation via food availability or quality can limit the growth of a population.
Invasive species may often be regulated by self-limitation. Invasive populations commonly exhibit periods of rapid growth shortly after invasion, followed by periods of reduced growth and even population decline (i.e., a boom-bust pattern)19,20. Boom-bust patterns are often attributed to self-limitation due to lack of resources19. However, while self-limitation is often invoked to explain these patterns in invasive species, it is rarely demonstrated. Further, the mechanistic role of life history tradeoffs in this self-limitation, triggered by reduced quality or abundance of food intake, remains unknown. A literature search using the Web of Science (search terms: “self-limitation” AND “invasive species”; or the terms “resource depletion” AND “invasive species” AND “reproduction”; or the terms “density dependent” AND “invasive species” AND “reproduction”) yielded numerous studies in plant systems, but only three studies in animal systems that have demonstrated the possibility for self-limitation, and none of them demonstrated a mechanistic link between resource availability and population vital rates21,22,23.
We examined the Asian shore crab Hemigrapsus sanguineus, which is invasive to the east coast of North America24 and to Europe25. On the North American east coast, H. sanguineus was first noted in 1988 in Cape May, New Jersey24, where it was most likely introduced via ballast water, perhaps multiple times26. Over the next few years it rapidly spread from mid-coast Maine to North Carolina26, where H. sanguineus displaced other species of crabs27,28 and became the numerically dominant crab species over much of this range29,30. High densities of 100–200 individuals m-2 have been documented throughout much of its range28,31,32,33. However, after several years of maintaining these high population densities, H. sanguineus has recently declined in some areas to approximately half its former densities33,34,35,36.
Several factors may contribute to population declines, including predation, competition, disease or parasites, etc., and each of these ecological interactions impose limits to invasive crabs in general37,38,39,40,41. However, previous work suggests that these mechanisms may have relatively little impact on Asian shore crabs in their invaded range. For instance, existing evidence suggests that common potential predators do not heavily prey on Asian shore crabs42,43,44. Additionally, competition does not seem to play a major role, as this species has the upper hand in interspecific interactions with other intertidal crabs28,40 and displays fairly weak conspecific interference competition45. Finally, the Asian shore crab appears to enjoy enemy escape in its invaded range and is infected by fewer parasite species, and at lower prevalence, than conspecifics in its native range46,47,48. Thus, while all of these factors may be contributing to the recent declines in Asian shore crab populations, none of them stands out as a dominant driving force. We therefore examine the potential role of self-limitation via food reduction as an additional mechanism contributing to the Asian shore crab decline.
H. sanguineus has a generalist diet that includes primarily algae, mussels, barnacles, gastropods, and detritus49,50,51, and while their natural diet is often mostly herbivorous, they prefer to eat animal tissue when it is readily available52. Numerous studies have demonstrated the importance of diet across a wide range of brachyuran crab species for determining the extent of energy storage and reproductive success17,53,54,55,56,57. It therefore seems reasonable that energetics and reproduction in H. sanguineus should also be influenced by diet, and that this invader should generally be more successful when consumption rates are higher and/or when higher quality food is consumed. And indeed, energy storage in H. sanguineus increases strongly when consuming an animal diet compared to an algal diet58. However, the link between diet and reproductive success in H. sanguineus has not been clearly made, and there is a lack of correlation between energy storage and reproductive success in this species59. Additionally, the role of life history tradeoffs (e.g., between growth and reproduction) remains unclear for this invader. If a clear link can be made between the quality of diet and reproductive success, then this would suggest that self-limitation due to overexploitation of food resources is a contributing factor leading to the recent localized declines in H. sanguineus population sizes. Further, if food-induced life history tradeoffs can be demonstrated, then this would provide a plausible evolutionary mechanism in this species that could functionally induce self-limitation.
We examined long-term diet quality, energy storage, reproductive effort, and growth of individual H. sanguineus collected on the New Hampshire coast where this species has been present and abundant for 20 years. We sampled crabs across two sites that differ in community abundance and thus food availability in order to capture as wide a range of individual diet qualities as possible. For each crab, we assessed the general long-term dietary strategy and physiological growth and reproductive performance. We tested the general hypothesis that individual growth and reproduction would be greatest for individuals that consume a higher quality diet. We deconstruct this broad hypothesis into a series of more focused hypotheses, described below.
Results
We found, consistent with our hypothesis, that crabs with larger stomach volumes consumed more food (t = 2.90, P = 0.004). However, while the main effect of site was not significant (t = − 0.43, P = 0.67), the interaction between CW and site was significant (t = 3.02, P = 0.003), indicating that the mass of food consumed increased faster with stomach volume for crabs at Odiorne Point than for crabs at Ft. Stark (Fig. 1). When each site was examined separately, we found that the mass of food in a crab’s stomach increased by 0.150 ± 0.027 mg and by 0.338 ± 0.042 mg for every 1-mm3 increase in stomach volume for crabs at Ft. Stark and at Odiorne Point, respectively (Fig. 1). However, we found that increasing the amount of food consumed did not translate into higher energy intake. Indeed, the energy content of food found in the stomach decreased by 8.58 ± 2.24 kJ with each additional mg of food consumed (t = − 3.83, P = 0.0002, Fig. 2).

Mass of the stomach contents compared to calculated stomach volume for Hemigrapsus sanguineus captured at Odiorne Point (red circles, dashed line) and Ft. Stark (black circles, solid line).

Energy content of food found in the stomach of Hemigrapsus sanguineus as a function of the amount of food consumed.
As we explain in the Methods, we used the residual SW, after accounting for differences in SW with CW, as a proxy for long-term diet quality, where smaller residual SW indicates consumption of a consistently higher quality diet and larger residual SW indicates consumption of a consistently lower quality diet. On average, residual SW for crabs at Odiorne Point were larger than for crabs at Ft. Stark (t = 2.51, P = 0.006, Fig. 3). We found that crabs at Ft. Stark had higher mass-specific energy stores than crabs at Odiorne Point (t = − 3.61, P = 0.0004, Fig. 3). After accounting for differences across site, there was no additional impact of long-term diet quality on mass-specific energy storage (t = − 1.52, P = 0.13, Fig. 3).

Relationship between the hepatosomatic index, or the proportion of total body weight comprised of the hepatopancreas, and residual stomach width after accounting for differences in stomach width with crab carapace width. Red circles indicate crabs from Odiorne Point; black circles indicate crabs from Ft. Stark. The horizontal boxplots at the top indicate residual SW for Odiorne Point (red) and Ft. Stark (black), where the solid line at the notch indicates the median value, the box encompasses the middle 50% of the data from the 1st to the 3rd quartile, the whiskers encompass 95% of the data, and the circles are outliers that fall outside this range.
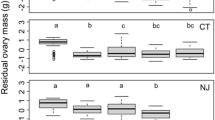
Based on the generalized linear model with a binomial error distribution, we found that the odds of a crab being gravid increased as CW increased (z = 2.49, P = 0.013, Fig. 4a), decreased as the residual SW increased (z = − 3.00, P = 0.003, Fig. 4b), and was influenced by the interaction of these two predictor variables (z = 2.74, P = 0.006). We also found that the mass of the clutch of eggs was influenced by crab CW (t = 8.51, P < 0.0001), residual SW (t = − 2.59, P = 0.011), the number of limbs that were missing (t = 1.96, P = 0.053), the site of collection (t = 1.90, P = 0.059), as well as by several interactions between these variables. Specifically, crab CW interacted with residual SW (t = 2.74, P = 0.007), with the number of missing limbs (t = − 2.32, P = 0.022), and with collection site (t = − 2.31, P = 0.023). Further, residual SW interacted with the number of missing limbs (t = 1.99, P = 0.049). Lastly, three-way interactions were significant between CW, residual SW, and the number of missing limbs (t = − 2.21, P = 0.029) and between residual SW, the number of missing limbs, and site of collection (t = 2.82, P = 0.006, Fig. 4c). While these trends are complicated due to the number of interactions, inspection of Fig. 4c,d reveals that in general, clutch mass increased for larger crabs (larger symbols found towards the top of the Fig. 4c), increased with the quality of the diet (negative trend in clutch mass with residual SW in Fig. 4c), and decreased with the number of missing limbs (Fig. 4d).

Influence of crab CW (A) and residual SW (B) on whether Hemigrapsus sanguineus are gravid. Boxplots interpreted as described in Fig. 3 caption. Relationship between the mass of the clutch of eggs and residual SW (C) and number of missing limbs (D). In (C,D): red and black symbols show Odiorne Point and Ft. Stark, respectively; symbol size shows relative crab CW, symbol type shows number of limbs missing (0: circle, 1: triangle, 2: plus sign, 3: X, 4: diamond, 5: inverted triangle). Symbols in (D) are jittered slightly in the x-direction to reduce overlap of data points for presentation purposes only.
Finally, we found that residual body mass (after accounting for CW) increased as the residual SW increased (t = 3.68, P = 0.003, Fig. 5). No other terms were included in the best-fitting model.

Residual total body mass (after accounting for changes with CW) as a function of residual SW for Hemigrapsus sanguineus for crabs collected at both sites.
Discussion
We have shown that the mass of consumed food increased with body size, but increased faster at Odiorne Point where high quality food is less available, and that there was an inverse correlation between the mass of food a crab consumed and the quality of that food. We have further demonstrated that body size and diet quality both influenced the likelihood that a crab was gravid, while the size of an egg clutch for gravid crabs increased with crab size, increased with higher diet quality (smaller residual SW), and decreased with injury, and these trends were generally seen across the two collection sites. Finally, we have shown that size-independent body mass increased as diet quality decreased, suggesting that individuals consuming a poorer quality diet have increased growth rates. These results are consistent with the concept of self-limitation in H. sanguineus, potentially driven by a tradeoff in energy allocation between reproduction and growth depending on diet quality.
While not in the context of invasive species, previous work has, however, examined self-limitation (often referred to as self-thinning) of sessile aquatic animals, such as mussels (e.g.60), barnacles61, and tunicates62. Mechanistically, such relationships can result from food competition at high densities63, analogous for these sessile animals to resource competition seen in plants that leads to a similar self-thinning phenomenon11. For these sessile plants and animals, scaling rules have been developed and debated that can be used to predict the relationship between density and biomass of individual organisms within a single age cohort60,64. Developing a similar scaling rule for mobile organisms would be challenging, as it would require density and biomass estimates across numerous replicate sites. Consequently, the concept of self-thinning or self-limitation for mobile organisms is not as thoroughly developed or demonstrated. And as highlighted in the Introduction, relatively few studies have successfully demonstrated self-limitation in mobile invasive species.
This is an observational study, and is thus open to alternative interpretations; however, our results are consistent with the concept of self-limitation in H. sanguineus. We have shown that reproduction tended to decrease in crabs that ate a lower quality diet, and in crabs that experienced higher levels of nonlethal injury (limb loss). Reduced diet quality and elevated nonlethal injury are both expected at high H. sanguineus densities due to depletion of food resources and increased conspecific aggression. These trends were apparent among individual crabs (the level of replication in this study), but also were consistent across the two sampling sites used here. Specifically, Odiorne Point has higher crab densities and lower per capita food availability, and crabs at this site generally consumed a lower quality diet and had lower reproductive rates (when comparing similarly sized crabs). Thus, reduced reproductive success, caused by depletion of high quality food, could help explain recent reductions in H. sanguineus population size in parts of its range where densities have previously been much higher.
Consumption by moderately high H. sanguineus densities (40 individuals m−2) can strongly reduce the entire algal and animal community65, and densities have historically been many times higher than this in areas where H. sanguineus has recently declined. At these high densities, H. sanguineus feeding activities greatly depress the prey community (e.g.28,51,66). If high densities of H. sanguineus depress prey abundances throughout a region, then the resulting low reproduction and regional recruitment could result in reduced population growth rates that could feasibly contribute to the population declines that have been documented.
Our results additionally suggest that self-limitation in H. sanguineus could be caused by a tradeoff in resource allocation that is driven by diet quality. Specifically, we found that there was a positive correlation between diet quality and reproductive success, but a negative correlation between diet quality and size-specific body mass (i.e., the amount of soft tissue growth since the last molt). This suggests that individuals that consume a higher quality diet allocate resources primarily to reproduction, while individuals that consume a lower quality diet may allocate resources primarily to soft tissue growth. This pattern is in direct opposition to the previously-suggested expectation that crustaceans should prioritize reproduction in resource-poor environments and growth in resource-rich environments67. Higher soft tissue mass for individuals with lower quality diet may also indicate postponement of molting, since diet quality is important for molting in crustaceans68,69. Consumption of a higher mass of lower quality food at Odiorne Point suggests that H. sanguineus is engaging in compensatory feeding to meet energy/nutrient demands at this site compared to at Ft. Stark. It is possible that this compensatory feeding triggers the tradeoff between resource allocation to growth vs. reproduction. Patterns consistent with this hypothesis have been shown in other systems, as dietary intake triggers a life history tradeoff in crickets, with increased dietary protein triggering allocation to reproduction, while increased dietary carbohydrates trigger allocation to muscle development to support migration70. Such strategies may evolve because diet composition determines the fatty acid composition of lipids that mothers deposit into eggs17, and fatty acid composition in turn influences development and survival of larvae71. Thus, when food quality is low, crabs may preferentially dedicate energy to growth rather than reproduction in order to increase size so that future reproductive attempts can be more prolific, given the allometric relationship between crab size and clutch size72.
The recent declines of H. sanguineus may have been influenced by factors other than self-limitation via food-depletion. For instance, mussel densities have declined in the Gulf of Maine over the last 40 years, possibly due to climate change73. The loss of this foundation species and the species it supports, would reduce food availability for H. sanguineus and other crabs inhabiting New England shores. Parasite infection may also play a role in limiting H. sanguineus success. This species is infected by both acanthocephalan and trematode parasites in its invaded range48,74, two groups of parasites that can impact the behavior and survival of crabs75,76. As highlighted in the Introduction, Asian shore crabs have fewer parasites in their invaded range than native species in the same area47 and fewer parasites than conspecifics in their native range46,47,48. However, while they have fewer parasites, they are more susceptible to infection46, and parasite load appears to be increasing since the initial escape from parasites at introduction, as native parasites in the invaded range expand to include this new host47. Similarly, while predation and competition have played a minor role in limiting the early invasion of this species28,40,42,43,44, it is possible that these processes may be taking on a greater role as time progresses, if native species learn to consume Asian shore crabs or more effectively interact with them. Thus, while our study does not suggest that resource limitation is the only, or even the predominant, mechanism leading to the decline of Asian shore crab populations, our results certainly do point to self-limitation because of food depletion as a contributing mechanism in the decline of this widespread invader.
Methods
Study sites
We collected crabs from Odiorne Point State Park in Rye, New Hampshire and from Fort Stark in New Castle, New Hampshire. These sites, located at the mouth of the Piscataqua River, are separated by only 2.3 km and are similar in many ways. Both are characterized by boulder fields overlying a mixed substrate of bedrock, cobble, and shell hash, and both support the same group of intertidal algal and animal species. However, Odiorne Point has more intertidal boulders than Ft. Stark, a habitat that is positively correlated with H. sanguineus abundance49. Consequently, H. sanguineus abundances averaged across all tidal heights at Odiorne Point (24.5 ± 18.8 m−2) are approximately 3X higher than those at Ft. Stark (7.5 ± 9.3 m−2)77. Further, while the two sites had nearly identical diversity and abundance of species within their intertidal communities prior to the arrival of H. sanguineus78, the higher density of H. sanguineus at Odiorne Point has had a greater impact on the prey community so that the abundance of each species is now lower at Odiorne Point than at Ft. Stark65 and references therein. The higher crab abundance and lower prey abundance at Odiorne Point means that the two sites differ considerably in the per capita amount of food available to H. sanguineus, particularly for animal prey, and therefore likely result in very different diets across the two sites. Our purpose for including crabs from both sites in our study was to capitalize on this difference in per capita prey availability in order to increase the range of diet strategies observed across crabs as much as possible. In all statistical analyses, we included site as a categorical independent variable to explicitly account for the fact that crabs were sampled across these two different sites.
Female performance
We sampled adult female crabs haphazardly from each site (n = 99 from Odiorne Point, n = 102 from Fort Stark). H. sanguineus forages most actively at night79, and so we sampled at dawn on receding tides to maximize the likelihood that there would be recently-consumed food in the stomach. We collected crabs by hand by turning over boulders to collect from intertidal regions of the shore. Upon collection, crabs were frozen in individual Ziploc bags and were returned on dry ice to Brigham Young University in Provo, UT for further analysis. H. sanguineus reaches maturity at 12.1 mm carapace width80, and only crabs exceeding this size were collected. Prior to dissection, we measured the carapace width (CW) of each crab to the nearest 0.1 mm and noted any missing or regenerating limbs. We then dissected crabs by first removing the dorsal carapace. Then we separated the hepatopancreas, cardiac stomach, egg clutch if the female had any, the ovaries, and the remainder of the body into separate pre-weighed tins that were dried to a constant weight at 65 °C and were then weighed to the nearest 0.01 mg. Prior to drying, we also measured the cardiac stomach width (SW) on the dorsal, anterior margin to the nearest 0.1 mm. All statistical tests described below were conducted using R v. 3.6.081.
We first analyzed the cardiac stomach to determine a snapshot of the amount of food consumed and how this was related to crab stomach size and collection site. We determined the mass of food consumed, independent of the mass of the stomach wall itself, as follows. We categorized the stomach as empty or not, based on visual observation through the transparent stomach wall. Using the subset of stomachs that were empty (n = 11), we determined the linear equation relating mass of the empty stomach to crab size (stomach mass = 0.0018 × CW – 0.0123, R2 = 0.71, P = 0.001) and used this to determine the mass of the stomach wall for each crab. For crabs with food in their stomachs, we then determined the mass of food consumed as the stomach total dry mass, minus the mass of the stomach wall using the relationship given above. This value inherently has error when used as a proxy of amount consumed due to variable digestibility of consumed food, time since consumption across crabs, and temporal variation in consumption amount from day to day; however, it provides a rough estimate of the relative amount of food consumed by individual crabs. We converted SW to stomach volume using the following relationship empirically determined by Griffen and Mosblack82:
We tested the hypothesis that crabs with larger stomachs would eat more using a linear model, with stomach content mass as response variable, and using stomach volume and site, and their interaction, as predictor variables, and only using crabs where stomach content mass > 0.
The mass of the stomach content indicates how much food was consumed, but says nothing about diet quality. We used energy content as a proxy for diet quality and determined the quality of the most recent meal by measuring the energy content of food found in the cardiac stomach. We combusted each stomach that contained food in a Parr 6725 semi-micro oxygen bomb calorimeter, yielding the energy content of each stomach. We ignored the energy content of the stomach wall itself because the mass of the empty stomach was below the detection limit of the calorimeter and therefore could not be combusted. The mass of food in some crab stomachs was very small, also falling below the detection limit of the calorimeter. For these, we paired nearly-empty stomachs from crabs collected at the same site and combusted the pairs together. We used a linear model to test the hypothesis that energy content (kJ) increased with the mass of food in the stomach, again only using crabs where stomach content mass > 0. We initially included sampling site in the analysis, but it was not significant (P > 0.4) and so it was removed.
The energetic content of the stomach mass just described provides an estimate of the diet quality of the last meal, but it is likely that the diet of an individual crab varies through time. The width of the cardiac stomach in crabs, and thus the stomach volume via the relationship given in the equation above, is strongly related to long-term diet. Specifically, Griffen and Mosblack82 demonstrated across 15 crab species that stomach size decreased with the amount of animal tissue included in the diet. This is likely because herbivorous crabs are generally nitrogen-limited83, and plants are often much lower in nitrogen than animal tissue84. Thus, crabs with a lower quality diet must consume a greater quantity to meet their nutritional needs (i.e., compensatory feeding). These differences in stomach size are evident throughout ontogeny, as crab species adapted to a more herbivorous diet start life with larger cardiac stomachs and experience faster allometric or isometric growth of the cardiac stomach throughout ontogeny compared to more carnivorous species85. Further, these diet-related patterns in stomach size between species are mirrored by differences between individuals within a single species. In a field experiment with H. sanguineus, Griffen and Mosblack82 showed that substantial diet variation exists between individual crabs, and that individual crabs that preferentially consumed animal material rather than algae, had smaller cardiac stomachs. Thus, intraspecific differences in long-term diet trends in this and other crab species17,86 can reliably be quantified by comparing differences in the width of the cardiac stomach, after controlling for differences in crab body size.
We assessed long-term diet quality of individual crabs using the width of the cardiac stomach. Stomach width increases linearly with CW in H. sanguineus85, but there is variation around the mean relationship. We therefore used this variation (i.e., the residual SW after accounting for CW) as a proxy for diet quality of individual crabs in the analyses that follow. We compared the general diet quality for crabs at the two sites by comparing residual SW using a t-test. We used a one-sided t-test because we expected the diet quality to be higher at Ft. Stark because, as described above, the per capita availability of animal prey is higher there77.
We used the mass of the hepatopancreas, an energy storage organ in crabs87, to test the hypothesis that energy storage differed with diet. We calculated the hepatosomatic index as the mass of the hepatopancreas divided by the mass of the crab88. We then used this index as the response variable in a linear model with residual SW and sampling site treated as predictor variables. We initially included the interaction term, but it did not explain a significant amount of the variation and so was removed.
We tested the hypothesis that reproduction varies with diet quality using the mass of eggs as our response variable. We chose to analyze eggs rather than ovaries because we reasoned that variation in ovary mass would be partially explained by time since the last egg clutch had been produced, which was unknowable for crabs not carrying a clutch of eggs when they were collected. There were a large number of crabs that were not gravid, resulting in a zero-inflated dataset. We therefore used a “hurdle” or “two-stage” modeling approach89. Specifically, we first used a generalized linear model with a binomial distribution (i.e., a logistic regression) to determine whether a crab was gravid (yes/no). Second, we used a linear model with the mass of the egg clutch as the response, including only those crabs that had an egg clutch. We included the following predictor variables and took the following approach in both of these analyses. We included CW because clutch size is expected to increase with crab size. We included the number of missing limbs because we expected that crabs that were regenerating limbs would have less energy to devote to reproduction. We included residual SW to examine our hypothesis that long-term diet strategy influences reproduction. And finally, we included site. We also included all possible interactions to create a full model and then compared the full model and all possible simpler models using AIC. In each case, we selected the best fitting model as the one with the lowest AIC and with ΔAIC for all other models being > 2.
Finally, we also examined the influence of diet quality on body mass as a proxy for growth. Crabs grow incrementally by periodically shedding their exoskeleton and growing a larger one, but in between molts, individuals grow soft tissue continually90. Thus, we examined the total body mass as the mass of the dry body, hepatopancreas, ovaries, eggs, and missing legs (i.e., all body parts with the exclusion of the full cardiac stomach so as not to include the mass of consumed food). We included the mass of missing legs to ensure that any differences between crabs were driven by differences in soft tissue mass and not differences in the number of missing legs. Thus, adding back in the mass of the missing legs reduces differences in mass between crabs and provides a conservative comparison of growth differences. We determined the mass of the missing legs by individually weighing each of the eight walking legs and the two claws from each of 30 different crabs of a range of CW. We then determined the regression equation for the mass of each of these 10 limbs as a function of CW (range of R2 values from the 10 regressions: 0.845–0.942). We then determined the mass of any missing leg for each crab using the appropriate regression equation for that specific leg. Total body mass increased allometrically with CW according to the equation: total body mass = 0.0027 × CW2.093. We therefore determined residual total body mass for each crab, after accounting for CW using this equation. Crabs with a larger residual body mass would have experienced more growth since the last molt, and we therefore used this residual body mass as the response variable in a linear model with residual SW, site, and their interaction as predictor variables; however, the interaction term and the main effect of site were not significant, and so were sequentially removed.
References
Bowman, W.D., Hacker, S.D., Cain, M.L. Ecology, 4th Edn. (Sinauer Press, 2017).
Eggleston, D. B., Lipcius, R. N. & Hines, A. H. Density-dependent predation by blue crabs upon infaunal clam species with contrasting distribution and abundance patterns. Mar. Ecol. Progr. Ser. 85, 55–68 (1992).
Boström-Einarsson, L., Bonin, M. C., Munday, P. L. & Jones, G. P. Strong intraspecific competition and habitat selectivity influence abundance of a coral-dwelling damselfish. J. Exp. Mar. Biol. Ecol. 448, 85–92 (2013).
Ruggerone, G.T., Zimmermann, M., Myers, K.W., Nielsen, J.L., & Rogers, D.E. Competition between Asian pink salmon (Oncorhynchus gorbuscha) and Alaskan sockeye salmon (O. nerka) in the North Pacific Ocean. Fish. Oceanogr. 12, 209–219 (2003).
Greer, A. L., Briggs, C. J. & Collins, J. P. Testing a key assumption of host-pathogen theory: Density and disease transmission. Oikos 117, 1667–1673 (2008).
Turchin, P. Does population ecology have general laws?. Oikos 94, 17–26 (2001).
Yenni, G., Adler, P. B. & Ernest, S. M. Strong self-limitation promotes the persistence of rare species. Ecology 93, 456–461 (2012).
Weis, A. E., Simms, E. L. & Hochberg, M. E. Will plant vigor and tolerance be genetically correlated? Effects of intrinsic growth rate and self-limitation on regrowth. Evol. Ecol. 14, 331–352 (2000).
Marino, A., Rodríguez, V. & Pazos, G. Resource-defense polygyny and self-limitation of population density in free-ranging guanacos. Behav. Ecol. 27, 757–765 (2016).
Chamaillé-Jammes, S., Fritz, H., Valeix, M., Murindagomo, F. & Clobert, J. Resource variability, aggregation and direct density dependence in an open context: The local regulation of an African elephant population. J. Anim. Ecol. 77, 135–144 (2008).
Westoby, M. The self-thinning rule. Adv. Ecol. Res. 14, 167–225 (1984).
Sedinger, J. S., Herzog, M. P., Person, B. T., Kirk, M. T., Obritchkewitch, T., Martin, P. P., & Bosque, C. Large-scale variation in growth of Black Brant goslings related to food availability. Auk 118, 1088–1095 (2001).
Marschall, E. A. & Crowder, L. B. Density-dependent survival as a function of size in juvenile salmonids in streams. Can. J. Fish. Aquat. Sci. 52, 136–140 (1995).
Zheng, X., Huang, L., Huang, B. & Lin, Y. Factors regulating population dynamics of the amphipod Ampithoe valida in a eutrophic subtropical coastal lagoon. Acta Oceanol. Sin. 32, 56–65 (2013).
Li, G. Y., & Zhang, Z. Q. Does size matter? Fecundity and longevity of spider mites (Tetranychus urticae) in relation to mating and food availability. Syst. Appl. Acarol.-UK 23, 1796–1808 (2018).
Niu, H., Zhao, L. & Sun, J. Phenotypic plasticity of reproductive traits in response to food availability in invasive and native species of nematode. Biol. Inv. 15, 1407–1415 (2013).
Cannizzo, Z. J., Lang, S. Q., Benitez-Nelson, B. & Griffen, B. D. An artificial habitat increases the reproductive fitness of a range-shifting species within a newly colonized ecosystem. Sci. Rep. 10, 1–13 (2020).
Zera, A. J. & Harshman, L. G. The physiology of life history trade-offs in animals. Annu. Rev. Ecol. Evol. Syst. 32, 95–126 (2001).
Strayer, D. L., D'Antonio, C. M., Essl, F., Fowler, M. S., Geist, J., Hilt, S., & Latzka, A. W. Boom‐bust dynamics in biological invasions: Towards an improved application of the concept. Ecol. Lett. 20, 1337–1350 (2017).
Jaćimović, M. et al. Boom-bust like dynamics of invasive black bullhead (Ameiurus melas) in Lake Sava (Serbia). Fish. Manag. Ecol. 26, 153–164 (2019).
Alcorlo, P., Geiger, W., & Otero, M. Reproductive biology and life cycle of the invasive crayfish Procambarus clarkii (Crustacea: Decapoda) in diverse aquatic habitats of South-Western Spain: Implications for population control. Fund. Appl. Limnol./Arch. Hydrobiol. 173, 197–212 (2008).
Melero, Y., Robinson, E., & Lambin, X. Density-and age-dependent reproduction partially compensates culling efforts of invasive non-native American mink. Biol. Invasions 17, 2645–2657.
Yoshida, K., Hoshikawa, K., Wada, T. & Yusa, Y. Patterns of density dependence in growth, reproduction and survival in the invasive freshwater snail Pomacea canaliculata in Japanese rice fields. Freshw. Biol. 58, 2065–2073 (2013).
Williams, A. B. & McDermott, J. J. An eastern United States record for the western Indo-Pacific crab, Hemigrapsus sanguineus (Crustacea: Decapoda: Grapsidae). Proc. Biol. Soc. Wash. 103, 108–109 (1990).
Breton, G., Faasse, M., Noël, P. & Vincent, T. A new alien crab in Europe: Hemigrapsus sanguineus (Decapoda: Brachyura: Grapsidae). J. Crustacean Biol. 22, 184–189 (2002).
Blakeslee, A. M., Kamakura, Y., Onufrey, J., Makino, W., Urabe, J., Park, S., & Miura, O. Reconstructing the invasion history of the Asian shorecrab, Hemigrapsus sanguineus (De Haan 1835) in the Western Atlantic. Mar. Biol. 164, 47 (2017).
Lohrer, A. M. & Whitlatch, R. B. Interactions among aliens: Apparent replacement of one exotic species by another. Ecology 83, 719–732 (2002).
Kraemer, G. P., Sellberg, M., Gordon, A. & Main, J. Eight-year record of Hemigrapsus sanguineus (Asian shore crab) invasion in western Long Island Sound estuary. Northeast. Nat. 14, 207–224 (2007).
Epifanio, C. E. Invasion biology of the Asian shore crab Hemigrapsus sanguineus: A review. J. Exp. Mar. Biol. Ecol. 441, 33–49 (2013).
Lord, J. P. & Williams, L. M. Increase in density of genetically diverse invasive Asian shore crab (Hemigrapsus sanguineus) populations in the Gulf of Maine. Biol. Invasions 19, 1153–1168 (2017).
Brousseau, D. J., Kriksciun, K. & Baglivo, J. A. Fiddler crab burrow usage by the Asian crab, Hemigrapsus sanguineus, in a Long Island Sound salt marsh. Northeast. Nat. 10, 415–420 (2003).
O’Connor, N. J. Invasion dynamics on a temperate rocky shore: from early invasion to establishment of a marine invader. Biol. Invasions 16, 73–87 (2014).
O’Connor, N. J. Changes in population sizes of Hemigrapsus sanguineus (Asian Shore Crab) and resident crab species in southeastern New England (2010–2016). Northeast. Nat. 25, 197–201 (2018).
Schab, C. M., Park, S., Waidner, L. A. & Epifanio, C. E. Return of the native: Historical comparison of invasive and indigenous crab populations near the mouth of Delaware Bay. J. Shellfish Res. 32, 751–758 (2013).
Bloch, C. P., Curry, K. D., Fisher-Reid, M. C. & Surasinghe, T. D. Population Decline of the Invasive Asian Shore Crab (Hemigrapsus sanguineus) and Dynamics of Associated Intertidal Invertebrates on Cape Cod, Massachusetts. Northeast. Nat. 26, 772–784 (2019).
Kraemer, G. P. Changes in population demography and reproductive output of the invasive Hemigrapsus sanguineus (Asian Shore Crab) in the Long Island Sound from 2005 to 2017. Northeast. Nat. 26, 81–94 (2019).
Stentiford, G. D., Bateman, K. S., Dubuffet, A., Chambers, E., & Stone, D. M. Hepatospora eriocheir (Wang and Chen, 2007) gen. et comb. nov. infecting invasive Chinese mitten crabs (Eriocheir sinensis) in Europe. J. Invertebr. Pathol. 108, 156–166 (2011).
Bateman, A. W., Buttenschön, A., Erickson, K. D. & Marculis, N. G. Barnacles vs bullies: Modelling biocontrol of the invasive European green crab using a castrating barnacle parasite. Theor. Ecol. 10, 305–318 (2017).
Bojko, J., Stebbing, P. D., Dunn, A. M., Bateman, K. S., Clark, F., Kerr, Stewart-Clark, S., Johannesen, Á., & Stentiford, G. D. Green crab Carcinus maenas symbiont profiles along a North Atlantic invasion route. Dis. Aquat. Organ. 128, 147–168 (2018).
Jensen, G. C., McDonald, P. S. & Armstrong, D. A. East meets west: competitive interactions between green crab Carcinus maenas, and native and introduced shore crab Hemigrapsus spp. Mar. Ecol. Progr. Ser. 225, 251–262 (2002).
DeRivera, C. E., Ruiz, G. M., Hines, A. H. & Jivoff, P. Biotic resistance to invasion: Native predator limits abundance and distribution of an introduced crab. Ecology 86, 3364–3376 (2005).
Kim, A. K. & O’Connor, N. J. Early stages of the Asian shore crab Hemigrapsus sanguineus as potential prey for the striped killifish Fundulus majalis. J. Exp. Mar. Biol. Ecol. 346, 28–35 (2007).
Brousseau, D. J., Murphy, A. E., Enriquez, N. P. & Gibbons, K. Foraging by two estuarine fishes, Fundulus heteroclitus and Fundulus majalis, on juvenile Asian shore crabs (Hemigrapsus sanguineus) in Western Long Island Sound. Estuar. Coast. 31, 144–151 (2008).
Savaria, M. C. & O’Connor, N. J. Predation of the non-native Asian shore crab Hemigrapsus sanguineus by a native fish species, the cunner (Tautogolabrus adspersus). J. Exp. Mar. Biol. Ecol. 449, 335–339 (2013).
Griffen, B. D. & Delaney, D. G. Species invasion shifts the importance of predator dependence. Ecology 88, 3012–3021 (2007).
Keogh, C. L., Miura, O., Nishimura, T. & Byers, J. E. The double edge to parasite escape: invasive host is less infected but more infectable. Ecology 98, 2241–2247 (2017).
Kroft, K. L. & Blakeslee, A. M. Comparison of parasite diversity in native panopeid mud crabs and the invasive Asian shore crab in estuaries of northeast North America. Aquat. Invasions 11, 287–301 (2016).
Blakeslee, A. M., Keogh, C. L., Byers, J. E., Lafferty, A. M. K. K. D. & Torchin, M. E. Differential escape from parasites by two competing introduced crabs. Mar. Ecol. Progr. Ser. 393, 83–96 (2009).
Lohrer, A. M., Fukui, Y., Wada, K. & Whitlatch, R. B. Structural complexity and vertical zonation of intertidal crabs, with focus on habitat requirements of the invasive Asian shore crab, Hemigrapsus sanguineus (de Haan). J. Exp. Mar. Biol. Ecol. 244, 203–217 (2000).
Ledesma, M. E. & O’Connor, N. J. Habitat and diet of the non-native crab Hemigrapsus sanguineus in southeastern New England. Northeast. Nat. 8, 63–78 (2001).
Brousseau, D. J. & Goldberg, R. Effect of predation by the invasive crab Hemigrapsus sanguineus on recruiting barnacles Semibalanus balanoides in western Long Island Sound, USA. Mar. Ecol. Progr. Ser. 339, 221–228 (2007).
Brousseau, D. J. & Baglivo, J. A. Laboratory investigations of food selection by the Asian shore crab, Hemigrapsus sanguineus: Algal versus animal preference. J. Crustacean Biol. 25, 130–134 (2005).
Griffen, B. D. Linking individual diet variation and fecundity in an omnivorous marine consumer. Oecologia 174, 121–130 (2014).
Riley, M. E., Vogel, M. & Griffen, B. D. Fitness-associated consequences of an omnivorous diet for the mangrove tree crab Aratus pisonii. Aquat. Biol. 20, 35–43 (2014).
Griffen, B. D. & Norelli, A. P. Spatially variable habitat quality contributes to within-population variation in reproductive success. Ecol. Evol. 5, 1474–1483 (2015).
Griffen, B. D. & Riley, M. E. Potential impacts of invasive crabs on one life history strategy of native rock crabs in the Gulf of Maine. Biol. Invasions 17, 2533–2544 (2015).
Belgrad, B. A. & Griffen, B. D. The influence of diet composition on fitness of the blue crab, Callinectes sapidus. PLoS ONE 11, e0145481 (2016).
Griffen, B. D., Vogel, M., Goulding, L. & Hartman, R. Energetic effects of diet choice by invasive Asian shore crabs: Implications for persistence when prey are scarce. Mar. Ecol. Progr. Ser. 522, 181–192 (2015).
Griffen, B. D. The timing of energy allocation to reproduction in an important group of marine consumers. PLoS ONE 13, e0199043 (2018).
Guiñez, R., Petraitis, P. S. & Castilla, J. C. Layering, the effective density of mussels and mass-boundary curves. Oikos 110, 186–190 (2005).
Bertness, M. D., Gaines, S. D. & Yeh, S. M. Making mountains out of barnacles: The dynamics of acorn barnacle hummocking. Ecology 79, 1382–1394 (1998).
Guiñez, R. & Castilla, J. C. An allometric tridimensional model of self-thinning for a gregarious tunicate. Ecology 82, 2331–2341 (2001).
Alunno-Bruscia, M., Petraitis, P. S., Bourget, E. & Fréchette, M. Body size–density relationship for Mytilus edulis in an experimental food-regulated situation. Oikos 90, 28–42 (2000).
Weller, D. E. A reevaluation of the‐3/2 power rule of plant self‐thinning. Ecol. Monogr. 57, 23–43 (1987).
Griffen, B. D. & Byers, J. E. Community impacts of two invasive crabs: the interactive roles of density, prey recruitment, and indirect effects. Biol. Invasions 11, 927–940 (2009).
Lohrer, A. M. & Whitlatch, R. B. Relative impacts of two exotic brachyuran species on blue mussel populations in Long Island Sound. Mar. Ecol. Progr. Ser. 227, 135–144 (2002).
Nelson, K. Scheduling of reproduction in relation to molting and growth in malacostracan crustaceans. Crustacean Egg Product. 7, 77–116 (1991).
Kibria, G. Studies on molting, molting frequency and growth of shrimp Penaeus monodon fed on natural and compounded diets. Asian Fish. Sci. 6, 203–211 (1993).
Petit, H., Nègre-Sadargues, G., Castillo, R. & Trilles, J. P. The effects of dietary astaxanthin on growth and moulting cycle of postlarval stages of the prawn, Penaeus japonicus (Crustacea, Decapoda). Comp. Biochem. Physiol. A Physiol. 117, 539–544 (1997).
Clark, R. M., Zera, A. J. & Behmer, S. T. Nutritional physiology of life-history trade-offs: How food protein–carbohydrate content influences life-history traits in the wing-polymorphic cricket Gryllus firmus. J. Exp. Biol. 218, 298–308 (2015).
Rosa, R., Calado, R., Narciso, L. & Nunes, M. L. Embryogenesis of decapod crustaceans with different life history traits, feeding ecologies and habitats: A fatty acid approach. Mar. Biol. 151, 935–947 (2007).
Hines, A. H. Allometric constraints and variables of reproductive effort in brachyuran crabs. Mar. Biol. 69, 309–320 (1982).
Sorte, C. J., Davidson, V. E., Franklin, M. C., Benes, K. M., Doellman, M. M., Etter, R. J., & Menge, B. A. Long‐term declines in an intertidal foundation species parallel shifts in community composition. Global Change Biol.23, 341–352 (2017).
Goedknegt, M. A., Havermans, J., Waser, A. M., Luttikhuizen, P. C., Velilla, E., Camphuysen, K. C., & Thieltges, D. W. Cross-species comparison of parasite richness, prevalence, and intensity in a native compared to two invasive brachyuran crabs. Aquat. Invasions12, 201–212 (2017).
Latham, A. & Poulin, R. Field evidence of the impact of two acanthocephalan parasites on the mortality of three species of New Zealand shore crabs (Brachyura). Mar. Biol. 141, 1131–1139 (2002).
Latham, A. D. M. & Poulin, R. Effect of acanthocephalan parasites on hiding behaviour in two species of shore crabs. J. Helminthol. 76, 323–326 (2002).
Griffen, B. D., van den Akker, D., NiNuzzo, E. R., Anderson, L. III, & Vernier, A. Comparing methods for predicting the impacts of invasive species (in press).
Tyrrell, M.C., & Harris, L.G. Potential impact of the introduced Asian shore crab, Hemigrapsus sanguineus, in northern New England: Diet, feeding preferences, and overlap with the green crab, Carcinus maenas. in Marine Bioinvasions: Proceedings of the First National Conference, Cambridge, MA, 24–27 January 1999 (pp. 208–220). (MIT Sea Grant College Program, 2000).
Spilmont, N., Gothland, M. & Seuront, L. Exogenous control of the feeding activity in the invasive Asian shore crab Hemigrapsus sanguineus (De Haan, 1835). Aquat. Invasions 10, 327–332 (2015).
McDermott, J. J. The western Pacific brachyuran Hemigrapsus sanguineus (Grapsidae) in its new habitat along the Atlantic coast of the United States: reproduction. J. Crustacean Biol. 18, 308–316 (1998).
R Core Team. R: A Language and Environment for Statistical Computing. (R Foundation for Statistical Computing, Vienna, 2019). https://www.R-project.org/.
Griffen, B. D. & Mosblack, H. Predicting diet and consumption rate differences between and within species using gut ecomorphology. J. Anim. Ecol. 80, 854–863 (2011).
Wolcott, D. L. & O’Connor, N. J. Herbivory in crabs: Adaptations and ecological considerations. Am. Zool. 32, 370–381 (1992).
Mattson, W. J. Jr. Herbivory in relation to plant nitrogen content. Annu. Rev. Ecol. Evol. Syst. 11, 119–161 (1980).
Griffen, B. D., Cannizzo, Z. J. & Gül, M. R. Ecological and evolutionary implications of allometric growth in stomach size of brachyuran crabs. PLoS ONE 13, e0207416 (2018).
Gül, M. R. & Griffen, B. D. Diet, energy storage, and reproductive condition in a bioindicator species across beaches with different levels of human disturbance. Ecol. Indic. 117, 106636 (2020).
Vogt, G. Functional cytology of the hepatopancreas of decapod crustaceans. J. Morphol. 280, 1405–1444 (2019).
Kyomo, J. Analysis of the relationship between gonads and hepatopancreas in males and females of the crab Sesarma intermedia, with reference to resource use and reproduction. Mar. Biol. 97, 87–93 (1988).
Zuur, A. F., Ieno, E. N., Walker, N. J., Saveliev, A. A., Smith, G. M. Zero-truncated and zero-inflated models for count data. in Mixed Effects Models and Extensions in Ecology with R 261–293. (Springer, New York, 2009).
Mente, E. Effect of ration level on individual food consumption, growth and protein synthesis in the shore crab Carcinus maenas. In Nutrition, Physiology and Metabolism of Crustaceans 53–67 (Science Publishers, Enfield, 2003).
Acknowledgements
All applicable institutional and/or national guidelines for the care and use of animals were followed. Crabs were collected under New Hampshire Fish and Game permit #MFD 1921. This work was funded by the Roger and Victoria Sant Educational Endowment for a Sustainable Environment.
Author information
Authors and Affiliations
Contributions
B.D.G. conceived and designed the study, analyzed the results, and wrote the manuscript; B.D.G., E.R.D., and L.A. collected the samples, E.R.D., L.A., J.B., J.C., A.V., M.M., and B.P. processed samples and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Griffen, B.D., Bailey, J., Carver, J. et al. Mechanisms of possible self-limitation in the invasive Asian shore crab Hemigrapsus sanguineus. Sci Rep 10, 16908 (2020). https://doi.org/10.1038/s41598-020-74053-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-020-74053-5
