
Abstract
This study investigated the cytotoxic effects of oxidative stress (OS), high mobility group box 1 (HMGB1), ADAMTS (A disintegrin and metalloproteinase with thrombospondin motifs), and neuropathology associated with coenurus cerebralis (Taenia multiceps). ADAMTS-13, HMGB1, glutathione reductase (GR), copper/zinc superoxide dismutase (Cu/Zn SOD), and 8-hydroxy-2'-deoxyguanosine (8-OHdG) expression levels were studied. The study found that ADAMTS-13 (P < 0.005), HMGB1 (P < 0.005), GR (P < 0.005), Cu/Zn SOD (P < 0.005), and 8-OHdG (P < 0.005) levels were significantly higher in T. multiceps (c. cerebralis)-infected animals compared to healthy control animals. This study's most important finding was that HMGB1 up-regulation in neurons, endothelial cells, and glial cells can directly cause brain parenchymal destruction and that HMGB1-mediated oxidative stress plays a crucial role in the neuropathogenesis of coenurosis. The results also showed that increased levels of ADAMTS-13 may play a pivotal role in regulating and protecting the blood–brain barrier integrity and neuroprotection. These findings also suggest that ADAMTS-13 and HMGB1 compete in the prevention or formation of microthrombi, which was regarded as a remarkable finding. ADAMTS-13 and HMGB1 are valuable biomarkers for disease risk assessment, estimating host neuropathy following T. multiceps (c. cerebralis) exposure, and providing a new therapeutic target. This is the first study to show that HMGB1 and ADAMTS-13 are expressed in reactive cells and are associated with neuroimmunopathology in coenurosis.
Similar content being viewed by others


Introduction
Coenurosis is a zoonotic parasitic disease that is typically seen in small ruminants. It is caused by coenurus cerebralis, the larval stage of Taenia multiceps. The parasite's final hosts are carnivores such as dogs, coyotes, and foxes1,2. This parasitic disease is significant because it can be fatal for sheep and goats and cause life-threatening human complications3,4. Clinical signs in naturally and experimentally infected sheep and goats include ataxia, incoordination, lethargy, paralysis, moving in a circle, seizures, and blindness5,6.
High mobility group box 1 (HMGB1) is a widely expressed and highly abundant protein that has been shown to play many roles in the pathogenesis of inflammation-related diseases7,8. The important point here is its close relationship with neuropathology. HMGB1 is pro-inflammatory in neurodegenerative diseases such as Alzheimer's disease9,10. However, it has been revealed that HMGB1 exacerbates inflammation, paracrine and autocrine after ischemic brain injury and increases cytokine activity by being released from necrotic neurons and glial cells11,12,13. In addition, increased HMGB1 levels are a remarkable finding, especially in patients with major inflammatory diseases such as meningitis14,15,16. It has been demonstrated that endothelial damage occurs due to increased HMGB1 expression in diabetic neuropathy and contributes significantly to brain damage17. Most importantly, HMGB1 is involved in cognitive impairment18. These studies show that HMGB1 expressions play an essential role in the neuropathogenesis of diseases and contribute to the worsening of neuropathology. HMGB1 may damage the blood–brain barrier (BBB).
HMGB1 has been found to enhance BBB permeability and damage brain endothelial cells and pericytes19. In neurological diseases such as cerebral ischemic stroke, elevated HMGB1 levels exacerbate BBB damage20,21. Diabetic neuropathy increases BBB permeability owing to HMGB1 levels17,18.
A disintegrin and metalloprotease with thrombospondin type I repeats 13 (ADAMTS-13) are mainly produced by hepatic stellate cells in the liver22. However, recent research has revealed that it is also expressed in endothelial cells, glial cells, and neurons and plays a crucial role in neuroprotection23,24,25. ADAMTS-13 has been described as protective in neuroparenchymal tissue in ischemia–reperfusion brain injury26. In mice with subarachnoid hemorrhage, recombinant ADAMTS-13 treatments have shown significant reductions in microthrombus areas and damaged brain tissue27. In addition, it has been revealed in experimental stroke models that ADAMTS-13 deficiency exacerbates neuronal damage and contributes to the recovery process by reducing neuropathology following treatment with ADAMTS-1328,29. It has been reported that von Willebrand Factor, closely related to ADAMTS-13, has important roles in BBB permeability30. In addition, it has been emphasized that ADAMTS-13 undertakes neuroprotective functions in revealing the BBB damage in the brains of small ruminants infected with Pestiviruses25 and in listeria encephalitis31, and by mitigating this damage. Likewise, BBB damage in toxoplasmic encephalitis24 and diabetic encephalopathy32 has been clearly demonstrated by ADAMTS-13 expressions, and its neuroprotective functions have been explained. These studies show that ADAMTS-13 has important roles in the BBB and neuronal parenchyma, revealing the severity of BBB damage and alleviating neuropathology.
Oxidative stress (OS) occurs when there is an imbalance between pro-oxidant and anti-oxidant mechanisms such as Glutathione reductase (GR), Copper-zinc superoxide dismutase (Cu/Zn SOD)33,34. All cellular molecules, such as lipids, proteins and especially nucleic acids, are highly susceptible to oxidative damage35,36. OS has been shown to induce BBB damage, responsible for the disruption of tight junction proteins in the BBB, and increased BBB permeability with altered blood flow37,38. As a result, OS and damaged BBB cause neurotoxicity and eventually irreversible neuropathology37,39. Neuronal cells are seen to be more sensitive to oxidative damage than other tissue cells36. OS is also very effective in triggering neuronal dysfunction and neurodegenerations and is one of the main causes of neuronal damage in many neurodegenerative diseases40.
There needs to be a comprehensive study investigating the molecular pathogenesis of Coenurus cerebralis-related neuropathology. This study aimed to determine whether there is a correlation between the previously unexplored HMGB1 expressions in T. multiceps (c. cerebralis)-infected sheep and lamb brains and the neuropathology caused by oxidative stress. In addition, the focus was on revealing the neuropathological severity of the disease and whether there is BBB damage with ADAMTS-13 expressions, which have never been investigated before. Therefore, this study aimed to investigate the molecular mechanism of neuroimmunopathogenesis associated with Coenurus cerebralis.
Results
Parasitologic results
Clusters of rosebush protoscolex typical of Taenia multiceps were visible with no magnification or special staining. The number of scolexes per cyst ranged from 23 in the small to approximately 164 in the larger one.
Clinicopathologic findings
It was found that cyst volumes and clinicopathological findings in T. multiceps (c. cerebralis)-infected lambs and sheep had a very close relationship. The clinicopathological results became more severe as cyst volumes and numbers increased. The most common neurological symptoms observed in T. multiceps (c. cerebralis)-infected animals were lateral deviation of the head and circling motion around its axis, paralysis, incoordination, and ataxia. Other findings included nystagmus, torticollis, a lack of direct light reflex in the pupil, seizures, and coma.
Gross findings
Macroscopically, white-colored protoscolex bundles ranging from 23 to 164 were found within the thin-walled, transparent, and clear fluid-filled cysts (Fig. 1B–D). The collected cysts were observed to be 2.4 × 3.3 cm–5.8 × 7.6 cm in diameter and varied in size from a ping-pong ball to a baseball (Fig. 1C,B). Sometimes, a sizeable unilateral cyst was found; in others, two to three small or large cysts were observed in different locations (Table 1). In the evaluation made according to the cyst locations in the brain, there was a more widespread distribution predominantly in the right or left cerebral hemispheres. Cysts were found in the cerebellum in only three cases (16.6%) and in the thalamus in one case (5.5%). When the cyst locations in the cerebral hemispheres were examined, it was seen that the cysts were in the right or left parietal, frontal, temporal and occipital lobes (Table 1). Because of the pressure atrophy caused by the cysts occupying the brain parenchyma (Fig. 1A–C), it was observed that the cyst area became thinner and even the skull's inner surface softened in all cases.

Gross pathology findings of T. multiceps (c. cerebralis)-infected animals. (A) C. cerebralis cyst and non-purulent meningoencephalitis area (arrow). (B) C. cerebralis cyst, scolexes and atrophy of surrounding tissue (arrowhead). (C) Cerebral hemisphere region showing cystic spaces due to c. cerebralis causing atrophy of brain tissue (arrowhead). (D) C. cerebralis cyst and scolexes (arrowhead).
Histopathologic findings
Hematoxylin and eosin-stained brain sections from healthy control animals exhibited standard architecture. Significant histopathological lesions were observed in the brain tissues of all animals. Histopathological examination revealed many multinucleated giant cells surrounding the cyst walls and lymphocytic reactions surrounded by inflammatory cells (Fig. 2A–D). Severe hyperemia and edema were detected in the meninges of T. multiceps (c. cerebralis)-infected animals. In addition, demyelination, liquefaction necrosis around cerebral cysts, degeneration of neurons, focal pressure atrophy, perivascular cell infiltrates, thrombus and fibrinopurulent meningoencephalitis were noted as the most important histopathological findings (Fig. 2A–D).

Histopathological findings of T. multiceps (c. cerebralis)-infected animals. (A) Non-purulent meningoencephalitis, areas of necrosis around the cyst membrane and numerous foreign body giant cells (arrow) against the cyst wall. (B) Foreign body giant cells (arrow) against the cyst wall and inflammatory zone area (mononuclear cell infiltration) (asterisk). (C) Numerous foreign body giant cells (arrow) and extensive areas of necrosis (arrowhead) (D) Extensive areas of necrosis (arrowhead), numerous foreign body giant cells (arrow) against the cyst wall and large inflammatory zone area (mononuclear cell infiltration) (asterisk).
Immunoperoxidase findings
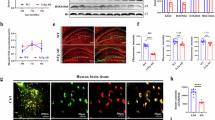
We analyzed the expression levels of ADAMTS-13, HMGB1, GR, Cu/Zn SOD, and 8-OHdG in the brain tissues from T. multiceps (c. cerebralis)-infected and healthy control animals. Immunohistochemical analysis showed significant up-regulation of ADAMTS-13, HMGB1, GR, Cu/Zn SOD, and 8-OHdG expressions in the T. multiceps (c. cerebralis)-infected lambs and sheep, unlike that in the case of the healthy control animals. Statistical data analysis on ADAMTS-13, HMGB1, GR, Cu/Zn SOD, and 8-OHdG expressions in the brain, measured by immunostaining in all the groups, are listed in Fig. 3 and Tables 2, 3, 4, 5. According to the comparison between T. multiceps (c. cerebralis)-infected lambs and sheep, ADAMTS-13 expressions were found to be statistically significantly higher in T. multiceps (c. cerebralis)-infected lambs than in T. multiceps (c. cerebralis)-infected sheep (P < 0.001) (Table 4). Conversely, HMGB1 expressions of sheep infected with T. multiceps (c. cerebralis) were statistically higher than lambs infected with T. multiceps (c. cerebralis) (P < 0.001) (Table 4). Statistically significant differences were not observed in GR, Cu/Zn SOD and 8-OHdG expressions between lambs and sheep infected with T. multiceps (c. cerebralis) (Table 5).

Significant up-regulation of ADAMTS-13, HMGB1, GR, Cu/Zn SOD and 8-OHdG expressions in brains in T. multiceps (c. cerebralis)-infected animals.
ADAMTS-13 and HMGB1 expressions
Expressions of ADAMTS-13 and HMGB1 in all brain regions of all animals in both T. multiceps (c. cerebralis)-infected and healthy control animals were carefully measured. It was noted that ADAMTS-13 and HMGB1 expressions were increased, and the difference was statistically significant and relatively high when this increase was compared with the healthy control animals and interpreted quantitatively (P < 0.005) (Fig. 3 and Table 2). We detected weak ADAMTS-13 expressions in healthy control animals, glial cells and neurons (Fig. 4A). Contrary to these findings, ADAMTS-13 expressions in brain tissues of T. multiceps (c. cerebralis)-infected animals were significantly increased, especially in neurons (Fig. 4B–F). In addition, partial ADAMTS-13 immunoreactivities were detected in glial cells (Fig. 4C). Weak or deficient HMGB1 expressions were observed in neurons, endothelial and glial cells in healthy control animals (Fig. 5A). However, it was determined that the intensity of HMGB1 expression in the brain tissues of T. multiceps (c. cerebralis)-infected animals were higher than in the healthy control animals (Table 2). Pathological levels of HMGB1 expressions were found in glial and endothelial cells, especially in necrotic and healthy neurons (Fig. 5B–D). The most striking finding of this study is that the expressions of ADAMTS-13 and HMGB1 were significantly increased in the lesioned areas and the areas close to the cyst. The most striking finding of this study is that the expressions of ADAMTS-13 and HMGB1 are significantly increased in the lesioned areas and the areas close to the cyst. T. multiceps (c. cerebralis)-infected animal brains showed enhanced ADAMTS-13 and HMGB1 levels and prolonged OS, which may contribute to neurotoxicity and parenchymal degeneration. These conditions handle severe neuropathology, such as demyelination and thrombus and permeability of the BBB.

ADAMTS-13 expressions in healthy control and T. multiceps (c. cerebralis)-infected animals. (A) Weak ADAMTS-13 expression in glial, endothelial cells and neurons of healthy control animals. ABC technique (anti-ADAMTS-13), Mayer's hematoxylin counterstain. (B) Strong expression of ADAMTS-13 in glial (arrowheads) and neuronal cells (arrow). ABC technique (anti-ADAMTS-13), Mayer's hematoxylin counterstain. (C) Strong expression of ADAMTS-13 in glial cells (arrowheads). ABC technique (anti-ADAMTS-13), Mayer's hematoxylin counterstain. (D,E,F) Strong expression of ADAMTS-13 in neuronal cells (arrows). ABC technique (anti-ADAMTS-13), Mayer's hematoxylin counterstain.

HMGB1 expressions in healthy control and T. multiceps (c. cerebralis)-infected animals. (A) Weak HMGB1 expression in glial, endothelial cells and neurons of healthy control animals. ABC technique (anti-HMGB1), Mayer's hematoxylin counterstain. (B) Strong expression of HMGB1 in healthy and necrotic neuronal cells (arrows). ABC technique (anti-HMGB1), Mayer's hematoxylin counterstain. (C) Strong expression of HMGB1 in glial (arrows) and endothelial cells (arrwheads). ABC technique (anti-HMGB1), Mayer's hematoxylin counterstain. (D) Strong expression of HMGB1 in healthy and necrotic (arrows) neuronal cells. ABC technique (anti-HMGB1), Mayer's hematoxylin counterstain.
Glutathione reductase expressions
In healthy control animals, some glial cells and neurons exhibited relatively weak or no immunoreactivity for GR (Fig. 6A). GR expression increased significantly in the vascular endothelial, glial cells, and necrotic/healthy neurons (Fig. 6B–D), which was also substantially higher than the levels in the healthy control animals (P < 0.005). We detected that an important anti-oxidant marker, GR, was up-regulated considerably in T. multiceps (c. cerebralis)-infected animals (Fig. 3 and Table 3).

GR expressions in healthy control and T. multiceps (c. cerebralis)-infected animals. (A) Weak GR expression in glial, endothelial and neuronal cells of healthy control animals. ABC technique (anti-GR), Mayer's hematoxylin counterstain. (B) Strong expression of GR in neuronal (arrows) and endothelial cells (arrwhead). ABC technique (anti-GR), Mayer's hematoxylin counterstain. (C,D) Strong expression of GR in glial (arrows) and endothelial cells (arrwheads). ABC technique (anti-GR), Mayer's hematoxylin counterstain.
Cu/Zn SOD expressions
Moderate Cu/Zn SOD expressions were observed in neuronal and glial cells in healthy control animals (Fig. 7A). Another conspicuous finding of the present study was that Cu/Zn SOD expressions were markedly increased in the endothelia, glial cells and microglia/macrophages (Fig. 7B–D), which was also significantly higher than the levels in the healthy control animals (P < 0.005). We detected that a very important anti-oxidant marker, Cu/Zn SOD, was significantly up-regulated in T. multiceps (c. cerebralis)-infected animals (Fig. 3 and Table 3).

Cu/Zn SOD expressions in healthy control and T. multiceps (C. cerebralis)-infected animals. (A) Weak Cu/Zn SOD expression in glial, endothelial and neuronal cells of healthy control animals. ABC technique (anti-Cu/Zn SOD), Mayer's hematoxylin counterstain. (B) Strong expression of Cu/Zn SOD in glial cells (arrowheads). ABC technique (anti-Cu/Zn SOD), Mayer's hematoxylin counterstain. (C,D) Strong expression of Cu/Zn SOD in glial cells (arrows) and endothelial cells (arrowheads). ABC technique (anti-Cu/Zn SOD), Mayer's hematoxylin counterstain.
8-OHdG expressions
In healthy control animals, some glial/neuronal cells showed weak immunoreactivity to 8-OHdG (Fig. 8A). 8-OHdG expression was observed in the nucleus and in the cytoplasm of neurons, endothelial cells, glial cells, and microglia/macrophages (Fig. 8B–D), being more intense in the cytoplasm. 8-OHdG expressions measurements in T. multiceps (c. cerebralis)-infected animals were significantly higher than in healthy control animals (P < 0.005) (Fig. 3 and Table 3). The most conspicuous finding of the present study was that 8-OHdG expression markedly increased in cyst walls.

8-OHdG expressions in healthy control and T. multiceps (c. cerebralis)-infected animals. (A) Weak 8-OHdG expression in glial, endothelial and neuronal cells of healthy control animals. ABC technique (anti-8-OHdG), Mayer's hematoxylin counterstain. (B) Strong expression of 8-OHdG in endothelial (arrowheads), glial and neuronal cells (arrows). ABC technique (anti-8-OHdG), Mayer's hematoxylin counterstain. (C) Strong expression of 8-OHdG in infiltrative inflammatory cells (arrowheads), neuronal and glial cells. ABC technique (anti-8-OHdG), Mayer's hematoxylin counterstain. (D) Strong expression of 8-OHdG in neuronal (arrows), endothelial and glial cells (arrowheads). ABC technique (anti-8-OHdG), Mayer's hematoxylin counterstain.
Discussion
Coenurosis is a disease caused by coenurus cerebralis, frequently seen in small ruminants, caused by the larval stage of Taenia multiceps. Its zoonotic character poses a severe public health problem that cannot be ignored1,2. There are minimal studies on the neuroimmunopathogenesis of coenurosis, a significant parasitic disease. For this reason, the findings obtained in this study are significant in elucidating the neuroimmunopathogenesis of the disease. The study's most striking first finding, a significant increase in HMGB1 expression and a HMGB1-mediated OS, has already been discussed. The second significant finding is the presence of ADAMTS-13, which is highly expressed in neurons and glial cells. It has thus been clearly demonstrated that the BBB's permeability increases, and its integrity is disrupted, contributing to the development of neuropathological findings. The third finding is that HMGB1 and ADAMTS-13 compete to form and prevent microthrombi, one of the most essential histopathological findings (Fig. 9).

Molecular pathogenesis of neuropathology in T. multiceps (c. cerebralis)-infected animals.
There is a link between oxidative stress and HMGB1 expression41. OS has been shown to regulate HMGB1 translocation, release, and activity significantly41,42. It has been found that HMGB1 functions as a critical regulator in OS-related infection, sterile inflammation, necrosis, and apoptosis42. In modeling studies of neurodegeneration in which HMGB1 inhibition is performed, it has been revealed that OS and neuronal cell death are significantly reduced and that it prevents neuropathology that may occur against BBB damage20,43,44. These studies show that neuropathology occurs in neurodegenerative diseases because of HMGB1-mediated OS. As the most important finding of this study, we showed that HMGB1 expressed in both necrotic and healthy neurons and glial cells at pathological levels are associated with OS. It is thought that a serious increase in HMGB1 expressions is among the causes of severe mitochondrial DNA (mtDNA) damage because of OS. In short, it has been clearly demonstrated that the most crucial trigger of OS-mediated neuropathology in coenurosis is HMGB1. The significantly increased levels of HMGB1 and HMGB1-associated OS shown in this study are thought to underlie the neuropathology seen in coenurosis. The development of therapeutic agents specifically targeting HMGB1 or its receptors may help to reduce the inflammatory response and OS associated with coenurosis. Inhibition of HMGB1 release or blockade of downstream signalling pathways may contribute significantly to the reduction of neuroinflammation and related pathological processes.
The blood–brain barrier is a physical and metabolic barrier that regulates and protects the microenvironment in the central nervous system (CNS) and separates the CNS from the peripheral circulation45. Considering the histopathological findings, it is seen that the permeability of the BBB increases in coenurosis. For this reason, the brain parenchyma and cells become sensitive due to the increase in BBB permeability during infection and play a critical role in triggering and exacerbating inflammation. For this reason, the brain parenchyma and cells become sensitive due to the increase in BBB permeability during infection and play a critical role in triggering and exacerbating inflammation. This study emphasises two reasons for the increase in BBB permeability. 1) The first one is BBB damage occurs due to HMGB1 expression at the pathological level; 2) HMGB1-mediated OS increases BBB permeability and damages BBB integrity. Because it has been shown that increased HMGB1 expressions cause damage to brain endothelium and pericytes, disrupting BBB integrity19,46. Similarly, it has been reported that OS increases BBB permeability and that BBB damage caused by OS causes neurotoxicity, exacerbating neuropathology37,38. Another striking finding of this study was that ADAMTS-13 expression was significantly increased, particularly in neurons.
Our previous studies demonstrated that ADAMTS-13 is expressed at severe levels in endothelial cells, glial cells and neurons in Pestivirus-related neuropathology25, listeria encephalitis31, diabetic encephalopathy32 and toxoplasmic encephalitis models24. Thus, ADAMTS-13 indicates BBB damage severity and increased BBB permeability. Similarly, we found that the severity of the existing damage can be followed up with ADAMTS-13, and it gives essential ideas for interpreting the severity in neuropathology. In addition, we revealed that ADAMTS-13 has protective effects on the BBB. We demonstrated in this study that the significantly increased ADAMTS-13 expression in the brain tissues of sheep with coenurosis has the neuroprotective effects described above and previously revealed, protects the brain from BBB damage, and provides information about the severity of the disease. In light of the studies, the role of ADAMTS-13 in investigating Coenurus cerebralis-related neuroimmunopathogenesis is significant. This and our previous studies suggest that ADAMTS-13 may play a role in modulating vascular integrity, inflammation or coagulation processes that may affect BBB permeability. Furthermore, examination of the relationship between ADAMTS-13 and BBB damage may help to elucidate the interactions between the cells that make up the BBB and its associates.
Coenurus cerebralis has been shown histopathologically to cause demyelination in the brain47,48,49. In this study, we also found demyelination in 5 sheep. Although demyelination is a significant neuropathology, no studies have been conducted to investigate the pathogenesis of demyelination in this disease. We previously investigated the role of ADAMTS-13 in preventing hydranencephaly, cerebellar hypoplasia, and, in particular, hypomyelinogenesis in Pestivirus-infected sheep. Furthermore, we discussed in the same study that it protects against the formation of myelin damage. The increase in ADAMTS-13 expression observed in this study may also play a role in demyelination prevention. It is obvious that when ADAMTS-13 expression levels are high enough, myelin damage will stop. Although demyelination is seen, the fact that it was not found in all sheep with coenurosis brains supports our theory.
The roles of ADAMTS-13 in thrombotic activity are significant. It has been revealed to prevent further deterioration of disease prognosis by cleaving ultra-large von Willebrand factor multimers into multimers with lower activity to reduce potential thrombotic activity and prevent microvascular thrombosis50,51. Mice with subarachnoid hemorrhage have shown statistically significant reductions in microthrombus areas and attenuated brain tissue damage after treatment with recombinant ADAMTS-1327. In contrast to ADAMTS-13, HMGB1 has been shown in an animal model of deep vein thrombosis to induce pro-thrombotic neutrophil extracellular trap formation and in vivo/in vitro platelet aggregation52,53,54. These studies show that increased HMGB1 activations at pathological levels may expose patients to a potential stroke. Thrombosis in coenurosis has also been linked to severe neuropathology55,56. This indicates that disturbances in thrombotic activity in coenurosis play an essential role in neuropathogenesis. At this time, it is believed that ADAMTS-13 and HMGB1 have a competitive interaction. Demyelination would not be interpreted this way if it were considered a universal neurohistopathological finding in all animals. However, the absence of demyelination in animals with coenurosis leads us to believe that ADAMTS-13 and HMGB1 compete as winners and losers. With HMGB1 promoting thrombus formation and ADAMTS-13 counteracting it, it is quite clear that an imbalance in favour of HMGB1 could lead to increased microthrombus formation and potentially contribute to tissue damage in coenurosis-related neuropathology. This demonstrated important finding suggests that targeting HMGB1 to reduce its pro-inflammatory and pro-thrombotic effects or increasing ADAMTS-13 activity to reduce microthrombus formation could be considered as potential interventions to limit disease progression.
In conclusion, this study revealed previously unknown neuroimmunopathogenesis of neuropathology in sheep with coenurosis. Damage to the BBB and increased permeability are visible. It is seen that HMGB1 alone and HMGB1-mediated OS are responsible for the damage and increased permeability of this BBB. In addition, the presence of mtDNA damage caused by OS was demonstrated by 8-OHdG expressions. In addition, it is thought that demyelination, an important neuropathological finding, is prevented with ADAMTS-13 expression. In addition, it was seen as a remarkable finding that ADAMTS-13 and HMGB1 play a competitive role in the prevention or formation of microthrombi.
Understanding the mechanisms underlying neuronal injury and inflammation in coenurosis provides important information for the development of neuroprotective interventions. With the findings obtained in this study, strategies that reduce neuronal plasticity and oxidative stress or promote tissue repair mechanisms can be investigated as potential therapeutic avenues. For example, targeting these factors, such as specific parasite proteins or antigens, may be among the first targets to reduce disease severity and promote recovery.
Materials and methods
Ethics statement
Aksaray and Atatürk University's Animal Experiments Ethics Committee did not require approval. Because this study did not include any invasive procedures for animal experiments. The study samples comprised 18 dead lambs and sheep, ranging in age from 8 to 36 months. These animals were brought to Aksaray and Atatürk University, Faculty of Veterinary Medicine, Department of Pathology for routine necropsy. No animals studied were sacrificed for this study, and all procedures were performed with the permission of the animal owners. As a healthy control animal, a total of 6 lambs and 6 sheep brains slaughtered for human consumption from Erzurum Slaughterhouse were used. Only the heads of these healthy animals sacrificed in the slaughterhouse were purchased.
Parasitological examinations
When the skulls of 18 lambs and sheep were opened, fluid-filled cysts in different regions were encountered. The cysts were carefully removed without rupture and placed in Petri dishes containing 70% ethyl alcohol. The structure and content of each cyst were examined both macroscopically and microscopically, and the advanced scolex structures were investigated. The gross and morphological appearance of cysts, their dimensions and their location in the brain were recorded and photographed.
Pathologic examination
The healthy control animals compromised the brain samples of slaughtered healthy lambs (n = 6) and sheep (n = 6). No findings were found in the brain tissues of these animals, both macroscopically and histopathologically. All steps followed the procedure described by Dincel and Kul57. The brains were removed and fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) at pH 7.4 for 48 h and then were thoroughly rinsed overnight, under tap water. After performing the routine tissue preparation procedures of dehydration using graded alcohol and xylene, the tissue samples were embedded in paraffin blocks; 4–5 μm thick paraffin sections were then cut and mounted on glass slides. Hematoyxlin-Eosin (H&E) and immunohistochemical tests were performed, and they were analyzed using a trinocular light microscope (Olympus BX51 and DP25 digital camera). The severity of coenurus cerebralis in each animal was classified according to neuropathological changes: demyelination, hyperemia, mononuclear cell infiltrates, giant cells around the cystic membrane, gliosis, astrocytosis, neuronal degeneration and malaise.
Antibodies
Commercial antibodies against ADAMTS-13 (Abcam, Cambridge, UK) diluted to 1:100, HMGB1 (Thermo Scientific, USA) diluted to 1:200, GR (Santa Cruz Biotechnology Dallas, TX, USA) diluted to 1:250, Cu/Zn SOD (Santa Cruz Biotechnology Dallas, TX, USA) diluted to 1:400, and 8-hydroxy-2-deoxyguanosine (8-OHdG) (Santa Cruz Biotechnology, Dallas, TX, USA) diluted to 1:500, were used.
Immunoperoxidase examinations
Immunohistochemistry was performed to observe ADAMTS-13, HMGB1, GR, Cu/Zn SOD and 8-OHdG in the 4–5 μm-thick paraffin sections of the tissues by using an indirect streptavidin/biotin immunoperoxidase kit (HRP, Thermo Scientific, USA), as per the manufacturer's instructions. All steps followed the procedure described by Dincel and Kul57. Briefly, the sections were placed onto adhesive slides and deparaffinized for 5 min. Each was in the 3-step xylene series, and rehydrated using a series of graded alcohol and distilled water. The antigens were retrieved by boiling the tissue sections on glass slides in citrate buffer (pH 6.0) (Thermo Scientific, USA) for 20 min. Endogenous peroxidase activity was quenched using 3% hydrogen peroxide in absolute methanol for 7 min at room temperature (RT). The tissue sections were rinsed thrice with phosphate buffer solution (pH 7.4) for 5 min, between each consecutive step. The sections were then incubated in a blocking serum for 5 min to prevent non-specific antibody binding. The sections were incubated with ADAMTS-13, HMGB1, GR, Cu/Zn SOD and 8-OHdG antibodies for 60 min in a humidity chamber at the RT. After treating the sections with biotinlabeled secondary antibody for 15 min and streptavidin-peroxidase enzyme for 15 min at RT, the colour reaction was performed using 3,3'-Diaminobenzidine chromogen for 5–10 min. Sections were counterstained with Mayer's hematoxylin for 1–2 min and suspended in water-based mounting medium (Thermo Scientific, USA). For each immunoperoxidase test, six (3 lambs and 3 sheep) negative control tissue sections were allowed as follows; as a negative control, one of the serial paraffin sections was incubated with normal mouse serum (isotype serum control) instead of primary antibody. The primary antibody step was also omitted to control non-specific endogenous peroxidase and biotin activities in each test.
Histomorphometric analysis and statistics
The density of positive staining was measured using a computerized image system composed of a Leica CCD camera DFC420 (Leica Microsystems Imaging Solutions, Ltd., Cambridge, UK) connected to a Leica DM4000 B microscope (Leica Microsystems Imaging Solutions, Ltd.). Five representative fields were selected, and consecutive pictures were captured by Leica QWin Plus v3 software under 20 × objective lens (Leica Microsystems Imaging Solutions, N Plan) at a setting identical to the imaging system for analysing. We used the same setting for all slides. The integrated optical density of all ADAMTS-13-, HMGB1-, GR-, Cu/Zn SOD-, and 8-OHdG-positive staining were measured, and the mean ADAMTS-13-, HMGB1-, GR-, Cu/Zn SOD-, and 8-OHdG-positive area/total area was calculated by Leica QWin Plus v3. All images were collected under the same lighting conditions. To avoid observer bias, a blinded investigator quantified all sections. Data were described in terms of mean and standard deviation (mean ± SD) for area %. After calculating the proportion (% pixels) of the stained area to the whole field, each slide's mean (% pixels) staining area was determined. ADAMTS-13, HMGB1, GR, Cu/Zn SOD and 8-OHdG immunohistochemical results were compared between groups using a one-way analysis of variance and Tukey's multiple comparison tests. P < 0.005 was considered statistically significant.
Data availability
All data generated or analyzed during this study are included in this published article.
References
Scott, P. R. Diagnosis and treatment of coenurosis in sheep. Vet Parasitol. 30:189(1), 75–8 (2012).
Scala, A. & Varcasia, A. Updates on morphobiology, epidemiology, and molecular characterization of coenurosis in sheep. Parassitologia 48, 61–63 (2006).
Sharma, D. K. & Chauhan, P. P. S. Coenurosis status in Afro-Asian region: A review. Small Rumin. Res. 64, 197–202 (2006).
Ing, M. B., Schantz, P. M. & Turner, J. A. Human coenurosis in North America: Case reports and review. Clin. Infect. Dis. 21, 519–523 (1998).
Mohammadi, P. et al. Clinical report of Coenurosis cerebralis outbreak in Lori sheep. Comp. Clin Path. 30, 729–733 (2021).
Oryan, A., Akbari, M., Moazeni, M. & Amrabadi, O. R. Cerebral and non-cerebral coenurosis in small ruminants. Trop. Biomed. 31, 1–16 (2014).
Andersson, U. & Tracey, K. J. HMGB1 is a therapeutic target for sterile inflammation and infection. Annu. Rev. Immunol. 29, 139–162 (2011).
Harris, H. E., Andersson, U. & Pisetsky, D.S. HMGB1: A multifunctional alarmin driving autoimmune and inflammatory disease. Nat. Rev. Rheumatol. 31:8(4), 195–202 (2012).
Paudel, Y. N. et al. Enlightening the role of high mobility group box 1 (HMGB1) in inflammation: Updates on receptor signalling. Eur. J. Pharmacol. 5(858), 172487 (2019).
Nan, K. et al. HMGB1 gene silencing inhibits neuroinflammation via down-regulation of NF-κB signaling in primary hippocampal neurons induced by Aβ25-35. Int. Immunopharmacol. 67, 294–301 (2019).
Chen, X. et al. High-mobility group box-1 translocation and release after hypoxic ischemic brain injury in neonatal rats. Exp. Neurol. 311, 1–14 (2019).
Kim, S. W. et al. Glycyrrhizic acid affords robust neuroprotection in the postischemic brain via anti-inflammatory effect by inhibiting HMGB1 phosphorylation and secretion. Neurobiol. Dis. 46(1), 147–156 (2012).
Kim, J. B., Lim, C. M., Yu, Y. M. & Lee, J. K. Induction and subcellular localization of high-mobility group box-1 (HMGB1) in the postischemic rat brain. J. Neurosci. Res. 86(5), 1125–1131 (2008).
Tang, D. et al. A pilot study to detect high mobility group box 1 and heat shock protein 72 in cerebrospinal fluid of pediatric patients with meningitis. Crit. Care Med. 36, 291–295 (2008).
Taniguchi, N. et al. High mobility group box chromosomal protein 1 plays a role in the pathogenesis of rheumatoid arthritis as a novel cytokine. Arthritis Rheum. 48, 971–981 (2003).
Wang, H. et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science. 285(5425), 248–251 (1999).
Liu, L., Wang, N., Kalionis, B., Xia, S. & He, Q. HMGB1 plays an important role in pyroptosis induced blood brain barrier breakdown in diabetes-associated cognitive decline. J. Neuroimmunol. 15(362), 577763 (2022).
Fujita, K. et al. HMGB1, a pathogenic molecule that induces neurite degeneration via TLR4- MARCKS, is a potential therapeutic target for Alzheimer’s disease. Sci. Rep. 6, 31895 (2016).
Gao, S. et al. Histidine-rich glycoprotein inhibits high-mobility group box-1-mediated pathways in vascular endothelial cells through CLEC-1A. iScience. 23(6), 101180 (2020).
Nishibori, M., Wang, D., Ousaka, D. & Wake, H. High mobility group box-1 and Blood-brain barrier disruption. Cells 9(12), 2650 (2020).
Goldstein, R. S. et al. Elevated high-mobility group box 1 levels in patients with cerebral and myocardial ischemia. Shock. 25(6), 571–574 (2006).
Zhou, W. et al. ADAMTS13 is expressed in hepatic stellate cells. Lab. Investig. 85(6), 780–788 (2005).
Shang, D., Zheng, X. W., Niiya, M. & Zheng, X. L. Apical sorting of ADAMTS13 in vascular endothelial cells and Madin-Darby canine kidney cells depends on the CUB domains and their association with lipid rafts. Blood 108(7), 2207–2215 (2006).
Dincel, G. C. & Atmaca, H. T. Increased expressions of ADAMTS-13 and apoptosis contribute to neuropathology during Toxoplasma gondii encephalitis in mice. Neuropathology 36, 211–226 (2016).
Dincel, G. C. & Kul, O. Increased expressions of ADAMTS-13, neuronal nitric oxide synthase, and neurofilament correlate with severity of neuropathology in border disease virus-infected small ruminants. PLoS ONE. https://doi.org/10.1371/journal.pone.0120005 (2015).
Khan, M. M., Motto, D. G., Lentz, S. R. & Chauhan, A. K. ADAMTS13 reduces VWF-mediated acute inflammation following focal cerebral ischemia in mice. J. Thromb. Haemost. 10(8), 1665–1671 (2012).
Vergouwen, M. D., Knaup, V. L., Roelofs, J. J., de Boer, O. J. & Meijers, J. C. Effect of recombinant ADAMTS-13 on microthrombosis and brain injury after experimental subarachnoid hemorrhage. J. Thromb. Haemost. 12(6), 943–947 (2014).
Fujioka, M. et al. ADAMTS13 gene deletion aggravates ischemic brain damage: A possible neuroprotective role of ADAMTS13 by ameliorating postischemic hypoperfusion. Blood. 115(8), 1650–1653 (2010).
Wan, H. et al. Role of von Willebrand factor and ADAMTS-13 in early brain injury after experimental subarachnoid hemorrhage. J. Thromb. Haemost. 16(7), 1413–1422 (2018).
Noubade, R. et al. von-Willebrand factor influences blood brain barrier permeability and brain inflammation in experimental allergic encephalomyelitis. Am. J. Pathol. 173(3), 892–900 (2008).
Karayigit, M. O. & Dincel, G. C. Role of ADAMTS-13 and nNOS expression in neuropathogenesis of listeric encephalitis of small ruminants. Biotech. Histochem. 95(8), 584–596 (2020).
Dincel, G. C. & Yildirim, S. Overexpression of ADAMTS-13 and neuronal nitric oxide synthase relates with neuropathology in streptozotocin-induced type 1 diabetic rats. Int. J. Clin. Exp. Pathol. 9, 4761–4778 (2016).
Flohé, L. Looking back at the early stages of redox biology. Antioxidants. 9, 1254 (2020).
Sies, H., Berndt, C. & Jones, D. P. Oxidative stress. Annu. Rev. Biochem. 86, 715–748 (2017).
Halliwell, B. Oxygen and nitrogen are pro-carcinogens. Damage to DNA by reactive oxygen, chlorine and nitrogen species: Measurement, mechanism and the effects of nutrition. Mutat. Res. 443, 37–52 (1999).
Floyd, R. A. & Carney, J. M. Free radical damage to protein and DNA: Mechanism involved and relevant observations on brain undergoing oxidative stress. Ann. Neurol. 32, 522–527 (1992).
Freeman, L. R. & Keller, J. N. Oxidative stress and cerebral endothelial cells: Regulation of the blood-brain-barrier and antioxidant based interventions. Biochim. et Biophys. Acta 1822, 822–829 (2012).
Zehendner, C. M. et al. Moderate hypoxia followed by reoxygenation results in blood-brain barrier breakdown via oxidative stress-dependent tight-junction protein disruption. PLoS ONE. 8(12), e82823 (2013).
Dincel, G. C. & Atmaca, H. T. Role of oxidative stress in the pathophysiology of Toxoplasma gondii infection. Int. J. Immunopathol. Pharmacol. 29(2), 226–240 (2016).
Li, J. O. W., Li, W., Jiang, Z. G. & Ghanbari, H. A. Oxidative stress and neurodegenerative disorders. Int. J. Mol. Sci. 14(12), 24438–75 (2013).
Yu, Y., Tang, D. & Kang R. Oxidative stress-mediated HMGB1 biology. Front. Physiol. 6, 93 (2015).
Tang, D., Kang, R., Zeh, H. J. 3rd & Lotze, M. T. High-mobility group box 1, oxidative stress, and disease. Antioxid. Redox Signal. 14(7), 1315–35 (2011).
Sasaki, T. et al. Anti-high mobility group box 1 antibody exerts neuroprotection in a rat model of Parkinson’s disease. Exp. Neurol. 275(1), 220–231 (2016).
Santoro, M. et al. In-vivo evidence that high mobility group box 1 exerts deleterious effects in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model and Parkinson’s disease which can be attenuated by glycyrrhizin. Neurobiol. Dis. 91, 59–68 (2016).
Zlokovic, B. V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 57(2), 178–201 (2008).
Festoff, B. W., Sajja, R. K., van Dreden, P. & Cucullo, L. HMGB1 and thrombin mediate the blood-brain barrier dysfunction acting as biomarkers of neuroinflammation and progression to neurodegeneration in Alzheimer's disease. J. Neuroinflamm. 13(1), 194 (2016).
Haridy, M. Coenurus cerebralis cysts in the left lateral cerebral ventricle of a ewe. J. Vet. Med. Sci. 75, 1643–1646 (2013).
Amer, S. et al. Clinical, pathological, and molecular data concerning Coenurus cerebralis in Sheep in Egypt. Data Brief. 4(16), 1–9 (2017).
Soliman, S. M. et al. Immune cell reaction associated with Coenurus cerebralis infection in sheep with particular reference to ELISA as a diagnostic tool. Life (Basel). 12(10), 1515 (2022).
Dong, J. F. et al. ADAMTS-13 rapidly cleaves newly secreted ultralarge von Willebrand factor multimers on the endothelial surface under flowing conditions. Blood 100, 4033–4039 (2002).
Chauhan, A. K. et al. Systemic antithrombotic effects of ADAMTS13. J. Exp. Med. 203, 767–776 (2006).
Yang, H. et al. Reversing established sepsis with antagonists of endogenous high-mobility group box 1. Proc. Natl. Acad. Sci. USA 101(1), 296–301 (2004).
Vogel, S. et al. Platelet-derived HMGB1 is a critical mediator of thrombosis. J. Clin. Investig. 125(12), 4638–4654 (2015).
Stark, K. et al. Disulfide HMGB1 derived from platelets coordinates venous thrombosis in mice. Blood. 128(20), 2435–2449 (2016).
Farjani Kish, G., Khodakaram-Tafti, A., Hajimohammadi, A. & Nasrollah, A. Clinical and morphopathological characteristics of an enzootic occurrence of acute coenurosis (Coenurus cerebralis) in a sheep herd. J. Parasit Dis. 39, 280–283 (2015).
Gholami, N., Dehghan Rahim Abadi, M. H., Naji Zadeh, D., Golchin, H., Ildar Abadi & Rasmi A. Clinical, pathological, and CT scan findings of multiple Coenurus cerebralis cysts in a non-native lamb in Iran. BJVM 25(4), 672–680 (2022).
Dincel, G. C. & Kul, O. First description of enhanced expression of transforming growth factor-alpha (TGF-α) and glia maturation factor-beta (GMF-β) correlate with severity of neuropathology in border disease virus-infected small ruminants. Microb. Pathog. 128, 301–310 (2019).
Acknowledgements
This study was supported by the Researchers Supporting Project (RSP2023R111), King Saud University, Riyadh, Saudi Arabia.
Funding
Researchers Supporting Project (RSP2023R111), King Saud University, Riyadh, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
G.C.D., O.Y., S.Y., E.M. A., and S.E. contributed equally to this work.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dincel, G.C., Yavuz, O., Yildirim, S. et al. ADAMTS-13 and HMGB1-induced oxidative stress in Taenia multiceps-infected animals. Sci Rep 13, 17929 (2023). https://doi.org/10.1038/s41598-023-44376-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-023-44376-0
This article is cited by
-
The first link between ADAMTS-13 and apoptosis in type 1 diabetic liver injury
Histochemistry and Cell Biology (2025)
