
Abstract
Real Christmas trees contribute ~$2.5 billion to the US economy annually. True fir (Abies spp.) are the most popular trees and account for the majority of sales. The real Christmas tree commodity board identified aroma as the primary driver for consumers to purchase a real tree. We conducted consumer aroma panels for five (Fraser, noble, Nordmann, Trojan, and concolor) of the most popular fir Christmas trees in the US. Based on aroma characteristics alone, Fraser fir was found to be most liked while noble fir ranked last amongst consumers. Gas Chromatography Mass Spectroscopy metabolite profiling of terpene (the major drivers of conifer aroma) putatively identified 132 mono- and sesquiterpene and 48 diterpene metabolites across all five species. Among these, the sesquiterpene, β-bisabolene, was strongly correlated with consumer liking. These findings provide a resource to identify compounds positively associated with consumer preference to help guide Christmas tree genetic improvement efforts.
Similar content being viewed by others
Introduction
More than 14.5 million Christmas trees are cut annually in the United States1. True fir (Abies spp.) comprises approximately 70% of the real tree market with Fraser (A. fraseri) and noble (A. procera) fir accounting for most trees sold in the US2. Other popular North American fir Christmas tree species include white (A. concolor), balsam (A. balsamea), and grand (A. grandis) fir2. In recent years, the popularity of Turkish (A. nordmanniana subsp. bornmuelleriana) and Trojan (A. nordmanniana subsp. equi trojani) fir has increased amongst US Christmas tree growers as they are more resilient to climate and pest pressures. The closely related Nordmann fir (A. nordmanniana) is the premier Christmas tree species in Europe3. The unique morphological and aroma differences amongst fir species contribute to their popularity but very little is known about the specific traits that drive consumer preferences for specific species.
Recent surveys conducted by the real Christmas tree commodity board found that consumers overwhelmingly identified scent as the primary driver for the purchase of a real tree4. Sensory attributes (i.e. aroma profiles) strongly impact consumer preferences. Aroma is an important trait for many forest, food, and horticultural crops like Cinnamomum camphora, eucalyptus, citrus, strawberries, roses, and blueberries5,6,7,9,11,12. In food crops, cultivars are evaluated for aroma and flavor to ensure marketability and consumer satisfaction. The compounds responsible for variation in sensory attributes are then evaluated and associated with consumer responses. In blueberries, preferred cultivars have an aroma profile that is described as floral and sweet while less successful cultivars lack these characteristics9. The compounds that contribute to the overall blueberry aroma profiles include small molecule metabolites such as esters and terpenes both of which have been found to be highly heritable by quantitative geneticists. For example, the terpene metabolite linalool has a significantly higher concentration in preferred blueberry cultivars. An analysis of linalool using pedigree information of preferred and less preferred blueberry cultivars found it was highly heritable with values > 97%10. Linalool is also a major aroma compound in other fruits like strawberries8. These results support the importance of including information on aroma profiles when considering breeding decisions to improve consumer outcomes.
In conifers (i.e., any member of the Pinophyta), oleoresin serves as the primary source of aroma profiles and is comprised of a diverse array of terpene metabolites13,14,15. Conifer oleoresin accumulates in specialized structures that include resin blisters, resin ducts, resin glands, resin vesicles, and resin cells13. Terpenes are responsible for the distinct aroma profiles associated with conifers and can play important ecological roles in conifer defense against herbivores including small insects and large ungulates like deer. For instance, the unique aroma profiles of certain conifers including western red cedar (Thuja plicata) and Douglas fir (Pseudotsuga menziesii) can repel deer and “warn” them of unpleasant and potentially toxic compounds16,17. However, these same volatile terpenes evaporate at ambient temperature and create the distinct, highly desirable aromatic profile of a Christmas tree.
Terpenes have been evaluated in Abies species primarily to understand conifer defenses and the composition of essential oil extracts that are commercially valuable14,15,18,19,20,21,21,. However, to date, the impact of specific conifer terpenes on consumer liking of different Christmas tree species is virtually unknown. We establish here a baseline understanding of consumer responses to five premier fir Christmas tree species selected because they are commonly grown in the largest production areas and are disease resistant2,3. We also characterize the terpene profiles of consumer preference test samples to provide putative identities and compound targets associated with consumer descriptors and liking. An improved understanding of the compounds that contribute to a consumer preferred Christmas tree aroma profile will facilitate downstream breeding program activities. Ultimately, we aim to improve the aroma profiles of real Christmas trees while simultaneously preserving the rich diversity of defensive terpenes involved in pest resilience.
Materials and methods
Plant materials
All plant samples were collected in late-October 2021 to coincide with the typical start of the Christmas tree harvest season. Therefore, aroma profiles from these plant samples would be representative of real Christmas trees that consumers purchase during the holiday season. Current year lateral shoot (CYLS) tips (Fig. 1a, b) from clonal elite mother tree genotypes of Fraser, noble, concolor, Nordmann, and Trojan fir trees were harvested on October 21st and 22nd, 2021. Fraser, concolor, and Trojan fir samples were collected from the Upper Mountain Research Station in Laurel Springs, NC, US (36°24’ N; 81°18’ W). Noble and Nordmann fir materials were collected from the North Willamette Research and Extension Center located in Aurora, OR, US (45°28’ N; 122°75’ W). CYLS were removed from branches, placed in unsterilized autoclave bags, and stored at 4 °C prior to being shipped on ice (4 °C) to the Sims lab in the Food Science & Human Nutrition Department at the University of Florida on October 26th, 2021. All CYLS were collected from the sun facing south side of the tree for all consumer preference and terpene profiling experiments. Representative individual genotypes were selected for each species and used in all experimental evaluations to standardize plant materials. Genotypes used include North Carolina Premium Fraser Fir Seed coop genotype 51; noble fir Oregon clone Riley/Fanno-15; Trojan fir North Carolina CoFirGE I accession R31C7CAN84; concolor fir Upper Mountain Research Station fir common garden; and Nordmann fir Oregon CoFirGE I accession Ambrolauri. All plant materials were stored at 4 °C until the consumer test panel was performed on October 27th, 2021.

Examples of fir species samples used in consumer aroma panel tests and terpene analyses. (a) Current year lateral shoots (CYLS; yellow circles) were removed and used for all subsequent tests and analyses. (b) CYLS from each species ready for consumer aroma test panels. (c) Fir species aroma samples ready for consumer aroma test panels. (d) Separation of needles and stem tissues prior to terpene metabolite extraction.
Consumer aroma panel participants
The consumer panel was conducted in late-October to coincide with the start of the Christmas tree harvest season to ensure aroma profiles provide a representative snapshot of a real tree. The sensory testing protocol was performed in compliance with and approved by the University of Florida Institutional Review Board (IRB). Sensory evaluation participants were recruited through emails and announcements posted throughout the University of Florida-Gainesville campus. Consumer panelists were pre-screened based on their familiarity with fresh cut Christmas trees and personal Christmas tree consumption habits. One hundred panelists were selected for the panel including both consumers who display a live tree and those who do not. The consumer aroma test panel was conducted over the course of a five-hour time-period.
Christmas tree aroma sample preparation and consumer test panel
CYLS were cut into ~ 1-inch sections, placed into single use plastic condiment containers, and provided a three-digit sample identifier label known only to panel administrators (Fig. 1c). Panelists were seated, assigned a number, and asked to sign into Compusense® (Version 21.0.24, Compusense Inc., Guelph, ON, Canada). Panelists age, gender, and Christmas tree display habits (fresh cut, artificial, or no tree displayed) were recorded. Coded samples were then distributed to panelists for evaluation of aroma liking, aroma intensity, and aroma descriptors. Panelists were asked to rate aroma liking on a nine-point hedonic scale (1 = dislike extremely, 5 = neither like nor dislike, 9 = like extremely) and aroma intensity on a scale from 0 to 100 where 100 was very strong aroma and 0 was no aroma. Finally, panelists were asked to select fall that apply from a prepared list of descriptive words that best described the aroma of each sample or encouraged to write-in words of their own choosing.
Tissue sample collection and processing
Terpene profiles of representative aroma panel test samples were evaluated over a time course to determine if participants were subjected to changes in fir aroma profiles throughout the five-hour test. Three condiment containers from each of the five fir species were collected immediately after consumer test panel samples and were prepared for downstream terpene profiling experiments. Samples were subsequently collected at the start of each hour over the course of the five-hour test yielding unique timepoints for hours 1–5. All samples were immediately flash frozen in liquid nitrogen and stored at – 80 °C to prevent further changes to aroma chemistry. Following completion of the consumer test panel, samples were shipped on dry ice to the Whitehill laboratory, Christmas Tree Genetics program at North Carolina State University in Raleigh, NC for terpene analysis. Next, needles were separated from stem (including buds, bark, and xylem) tissues for all three biological replicates, five species and five time points (Fig. 1d). Needle and stem tissues were ground to a fine powder in liquid nitrogen. A total of 100 milligrams of fresh weight (FW) tissue was weighed directly into gas chromatography-mass spectrometry (GC-MS) vials for each sample. Three replicates were included for each combination of species, time point, and tissue type. All plant tissues remained frozen throughout sample processing and were stored at – 80 °C until terpene extraction.
Terpene analysis
Terpenes were extracted according to previously published methods22. Routine chemicals used for terpene extraction and derivatization were purchased from either Sigma Aldrich St Louis, MO, USA or ThermoFisher Scientific Waltham, MA, USA unless otherwise stated. Tert-methyl butyl ether (MTBE) with an internal standard for mono/sesqui-terpenes (isobutyl benzene) and diterpenes (dichlorodehydroabietic acid) was used as the extraction solvent. The solvent and internal standard solution were added to GC-vials containing previously weighed tissue. Tissues were allowed to incubate with solvent overnight for approximately 14 h. Two separate aliquots of sample extracts were removed for (1) mono/sesqui-terpene and (2) diterpene analyses. For the mono/sesqui-terpene analyses, 0.1 M ammonium carbonate (\(\:({\text{N}\text{H}}_{4}{)}_{2}\text{C}{\text{O}}_{3}\)) pH 8 was added to the 800 µL of the monoterpene and sesquiterpene extracts23. Extracts were allowed to separate between the aqueous and non-polar fractions. The non-polar fraction was stored at – 80 °C until GC-MS analysis. Diterpene extracts were derivatized using methanol and tri-methyl-silyl-diazomethane (TMS-diazomethane) in the fume hood to minimize exposure. TMS-diazomethane was allowed to react for 20 min and then immediately evaporated under a stream of nitrogen gas. Derivatized diterpenes were resuspended in diethyl ether (inhibitor free, HPLC grade) and stored at – 80 °C until GC-MS analysis. Both mono- and sesqui-terpene extracts were shipped on dry ice to the Zerbe lab at UC Davis for GC-MS analysis. Terpenes were analyzed on an Agilent 7890B GC system with a 5977 Extractor XL MS Detector at 70 eV electron ionization and 1.2 mL min–1 He flow (Agilent, USA). Metabolite separation was performed using an Agilent DB-5MS + DG column (30 m, 250 μm i.d., 0.25 μm film; Agilent, USA) with the following GC parameters: 50 °C for 3 min, 15 °C min–1 to 300 °C, hold for 4 min with pulsed splitless injection at 250 °C. MS data from 40 to 400 m/z were collected after a 7-minute solvent delay. Compounds were quantified using total normalized peak areas against the internal standards isobutyl benzene (for mono-/sesquiterpenes) and dichlorodehydroabietic acid (for diterpenes), tissue dry weights, and solvent volumes used for extraction. Several metabolite identities were confirmed using commercially available matching standards including: α -pinene, β-pinene, camphene, β-caryophyllene, α-phellandrene, terpinolene, limonene, myrtenol, geraniol, citronellol, humulene, bornyl acetate, cis-abienol, and abietic acid (all compounds ordered from Sigma-Aldrich St Louis, MO, USA). One Nordmann fir stem diterpene sample and one noble fir stem diterpene sample were omitted from the final dataset due to errors in sample preparation. Compounds not confirmed through comparison to authentic matching standards were assigned putative mono-, sesqui- and diterpene identities through a comparison of reference mass spectra with the Wiley National Institute of Standards and Technology (NIST) 2017 database.
Statistical methods
All data were analyzed using R24. Differences between species for liking and intensity ratings were determined using analysis of variance (ANOVA) and separated post-hoc with Fisher’s least square difference (LSD) test using DescTools26. Consumer panel data was evaluated using a linear model in the lme4 package25. The goal was to evaluate if age, gender, species, and tree display impacted consumer ratings. Liking and intensity ratings were regressed against individual aroma descriptors used by panel participants to identify associations among species. Finally, consumer aroma descriptors for each species were visualized using a spider plot generated using the fmsb package27. For terpene data, normalized peak areas were log transformed prior to all analyses and a small value (10–16) was added to all observations to account for missing peaks. Log transformations were necessary to meet assumptions of normality and homogeneity of variance in most cases. Repeated measures ANOVA was conducted to evaluate compound abundance changes over time. Principal component analysis (PCA) and hierarchical clustering were used to visualize species and tissue type differences for mono-/sesqui and di-terpene datasets. Hierarchical clustering was conducted using pvclust with the ward D2 method, Euclidean distance, and 100,000 bootstraps. Approximately unbiased (AU) values were calculated to determine the significance of each cluster presented on each dendrogram. The significance level of the clusters was set to 95. A larger confidence value is indicative of a “true” cluster28. PCA analyses were performed using prcomp while ggplot2 was used to visualize results29. Standard confirmed terpenes were evaluated using two-way ANOVA when peak data were normally distributed or could be transformed to meet assumptions of normality and homogeneity of variance. Data that did not meet normality assumptions were evaluated using the nonparametric Kruskal-Wallis test. Significantly different Kruskal-Wallis tests were separated post-hoc using Dunn’s test. Kruskal-Wallis test was used as normalized peak areas for multiple terpene metabolites were not normally distributed and the overall sample size was less than 30. All library matched peaks were evaluated separately using two-way ANOVA and post-hoc separations were performed using Fisher’s LSD test (α = 0.05).
Results
Consumer test panel demographics and preference for fir Christmas trees
The objective of the study was to establish a foundational understanding of consumer preferences for the most popular fir Christmas tree species based on aroma profiles and identify potential relationships with the terpene metabolites produced by each species. To address this objective, we conducted an aroma test panel comprised of 100 participants drawn from potential consumers of Christmas trees in Gainesville, Florida. The mean age of test panel participants was 30.7 ± 1.2 years of age with 64% of attendees identifying as female and 36% as male. 55% displayed a fresh cut tree, 39% displayed an artificial tree, and 6% did not display a tree of any kind. Consumer aroma test panel participants rated each of the five fir Christmas tree species using a nine-point hedonic scale in which a score of 9 indicates extreme like and a score of 1 indicates extreme dislike. The aroma of Fraser fir (7.4 ± 0.1) was ranked significantly higher than all other species tested by consumer test panel participants while noble fir (5.7 ± 0.2) was rated the lowest. Nordmann (6.6 ± 0.1), Trojan (6.2 ± 0.2), and concolor (6.3 ± 0.2) firs were all rated intermediately to both Fraser and noble firs (Fig. 2a). Consumer aroma test panel participants also rated each species based on the aroma intensity using a scale from 0 to 100 where 100 was most intense and 0 was least intense. Consumer panelists ranked Fraser (58.2 ± 2.3), Nordmann (55.5 ± 2.3), and noble (55.4 ± 2.1) fir as intermediate for aroma intensity while concolor fir (70.8 ± 2) had the highest aroma intensity and Trojan fir (46.5 ± 2.3) had the lowest aroma intensity (Fig. 2b).

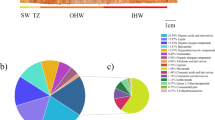
Results of consumer aroma test panels. (a) Average consumer liking scores by species. (b) Average aroma intensity ratings by species. (c) Linear regression of consumer liking scores and proportion of people who described each species as having a pleasant aroma. (d) Linear regression of consumer liking score and proportion of people who described each species as having a citrusy or fruity aroma. (e) Spider plot of aroma descriptors used to describe each species.
Correlations between participant demographics and aroma preferences
To assess how participant demographics might impact aroma panel results, a linear model was fit to explore the impact of gender, age, tree display preference (real, artificial, or no tree displayed) and species on consumer liking or intensity scores. Overall, gender did not have an impact on consumer scores (P = 0.97). Panelists aged 25, 32, 41, 57, or 61 had a significant impact in the model with P-values < 0.05 for each coefficient. While not significant overall, age trended towards a weakly positive correlation with consumer aroma liking ratings (P = 0.09). Tree display preference for those who displayed a fake tree was significant for having a negative effect on overall liking ratings (P = 0.01). The interaction between tree display preference and species was significant for a negative coefficient between displaying a fake tree and concolor fir (P = 0.02).
Consumer panel participant descriptions of christmas tree species aroma
Consumer panel participants were asked to select words from a provided list that best describe the aroma of each Christmas tree species tested. If provided words did not adequately convey the participants observations, then they were asked to provide descriptors of their own choosing. descriptors provided to test panel participants included “floral”, “citrusy/fruity”, “piney”, “woodsy”, “peppermint”, “unpleasant”, “pleasant” and “other”. Consumers who provided their own descriptors described Fraser fir as smelling “sweet/sugary”, “clean”, “Christmassy”, “fresh”, “nutmeg”, or “classic”. Those who selected other for concolor fir described it as smelling of “aged grass”, “disinfectant”, “mold and dirt”, “sweaty socks”, or “cinnamon”. Nordmann fir was described as smelling “woodsy and piney” or “plastic” while Trojan fir was described as “grassy”, “neutral”, or “leathery”. Noble fir was described as smelling of “dirt”, “astringent”, “herbal”, “asparagus”, “insecticidal”, or “delayed peppermint”. The frequency each of the eight main descriptors was regressed against aroma liking and intensity scores. The use of the word “pleasant” increased in frequency as consumer liking scores increased. Additionally, the use of the descriptor “citrusy/fruity” increased as the intensity rating increased (Fig. 2c, d). The frequency that each consumer aroma descriptor was used with each species was visualized through a spider plot which revealed that concolor fir was most frequently described as “citrusy/fruity” compared to the other species. The plot also revealed that Fraser fir was most frequently described as “pleasant” while noble fir was most frequently described as “unpleasant” (Fig. 2e).
Quantitative and qualitative differences among Fir Oleoresin mono- and sesqui-terpenes
We evaluated the composition of oleoresin to identify potential associations between terpene metabolites and consumer preferences for fir Christmas tree species based on aroma profiles. Peaks were identified as mono- and sesqui-terpenes as based on best matches to mass spectral databases and comparison to authentic standards where available. 50 metabolites were identified in Fraser fir, 66 in concolor fir, 45 in Nordmann fir, 49 in Trojan fir, and 61 in noble fir (Table S1). Fir samples were left at room temperature over the course of the five-hour aroma panel evaluation. To assess potential changes in metabolite profiles over time we compared mono- and sesqui-terpene normalized peak areas using principal component analysis (PCA). Results of the PCA demonstrated that samples from each time-point across all species consistently grouped together with no separation (Fig. S1a). To further evaluate the effect of time on sample integrity, a repeated measures ANOVA was performed and supported the PCA results (Table S2). However, abundance over time changed for several metabolites including peak 44, 83, 119, and 125. While these individual metabolite quantities differed over time the overall abundance did not decline chronologically indicating sample specific effects (Table S3). PCA was also used to compare similarities and differences among mono- and sesqui-terpene metabolite profiles between species and tissue types. For needle tissues, 20.3% of the variance was explained by principal component one (PC1) and 18.2% explained by PC2. Species largely separated within the same plane for PC1 but concolor fir separated completely in PC2 forming a distinct grouping from the other species evaluated (Fig. 3a). For stem tissues, PC1 explained 21.1% of the variance while PC2 explained 17.9% of the variance. Stem tissue metabolite profiles of Trojan, Nordmann, and Fraser fir grouped together while noble and concolor fir separated from each other in both PC1 and PC2 (Fig. 3b). The clusters observed in the PCA reflect the similarities among mono- and sesquiterpene metabolite profiles for species and tissue types while abundances of specific compounds were significantly different (Table S4). To further support PCA groupings, a hierarchical cluster analysis was performed. Overall, the hierarchical clustering dendrogram reflected the relationships observed in the PCA. A bootstrap analysis of individual needle tissue biological replicates supported the observed clusters for mono- and sesqui-terpene metabolite profiles with noble, Trojan, Fraser, and Nordmann fir samples grouping together, while concolor needle samples grouped separately (Fig. S2a). Similarly, hierarchical clustering of stem tissue mono- and sesqui-terpene metabolite profiles supported the observation that Fraser, Nordmann, and Trojan fir stem samples grouped together, while noble and concolor fir grouped independently (Fig. S2b).

Principal component analyses (PCA) of terpene metabolites in fir species by tissue type. (a) PCA of mono- and sesqui-terpene metabolites in needle tissues. (b) PCA of mono- and sesqui-terpene metabolites in stem tissues. (c) PCA of diterpene metabolites in needle tissues. (d) PCA of diterpene metabolites in stem tissues.
Mono- and sesqui-terpenes correlated with consumer preferred christmas tree species
To identify metabolites associated with consumer aroma liking preferences we performed two-way ANOVAs followed by post-hoc separations (Table S5) and Pearson correlations between consumer liking ratings and normalized abundances for each compound. Several mono- and sesqui-terpene metabolites were identified as being of interest following comparisons of the aromatic profiles of the most preferred versus least preferred species (Table 1). Peaks of interest include the most abundant metabolites identified in the highly preferred Fraser fir (peak #’s 7, 20, 21, 32, 52, 55, 77, 84, 89, 101, 112, 115, and 132). These peaks were also the least abundant in the lowest rated Christmas tree species, noble fir, suggesting roles in consumer liking preference. Putative identities for monoterpene peaks of interest include: (-) isopinocampheol (peak 7), (+)-2-bornanone (peak 20), camphor (peak 21), and δ-verbenone (peak 32). Peaks of interest putatively identified as sesquiterpenes, include α-longipinene (peak 52), longicyclene (peak 55), E-β-farnesene (peak 77), cis-α-bisabolene (peak 84), β-bisabolene (peak 89), trans-α-bisabolene (peak 101), isolongifolol (peak 112), 4-epi-α-acoradiene (peak 115), and cis-8-hydroxylinalool (peak 132).
Standard confirmed mono- and sesqui-terpenes
The identity for a subset of 15 mono- and sesqui-terpene metabolites were confirmed using exact matching standards (Table 2). Individual compounds varied in abundance between species and tissue types. Six compounds were confirmed as monoterpene hydrocarbons including α-pinene, limonene, terpinolene, camphene, α-phellandrene, and β-pinene (Fig. 4). α-Pinene was most abundant in Trojan fir and least abundant in noble and Nordmann fir (Fig. 4a). Limonene was significantly more abundant in noble fir compared to all other species (Fig. 4b). Abundance of terpinolene was similar among all species and tissue types although compound levels did vary significantly (Fig. 4c). Camphene was most abundant in concolor fir needles but was not detected in Trojan and noble fir (Fig. 4d). α-phellandrene was only detected in the stem tissue of Trojan fir (Fig. 4e). Finally, β-pinene was most abundant in concolor and Trojan fir but abundances did not differ between tissue types (Fig. 4f).

Standard confirmed monoterpene hydrocarbon metabolites. Normalized abundances are presented to highlight differences between tissue types and among species for each standard confirmed monoterpene hydrocarbon metabolite including: (a) α-pinene; (b) camphene; (c) α-pinene; (d) limonene; (e) α-phellandrene; and (f) terpinolene. Different letters indicate significant differences following post hoc separations between species and tissue types. Error bars represent the standard error of the mean.
Five standard confirmed compounds were identified as oxygenated monoterpenes including linalool, citronellol, geraniol, myrtenol, and bornyl acetate (Fig. 5). Linalool was most abundant in concolor fir with trace amounts present in the other species evaluated (Fig. 5a). Citronellol was most abundant in concolor fir with lower levels present in noble fir but not detected in any other species (Fig. 5b). Geraniol was significantly more abundant in the needles of concolor fir while least abundant in all other species and tissue types evaluated (Fig. 5c) Myrtenol was most abundant in Trojan fir, not detected in concolor fir with trace amounts present in the Fraser, Nordmann and noble fir (Fig. 5d). Bornyl acetate was most abundant in Fraser and concolor fir but trace amounts were detected in the other species (Fig. 5e).

Standard confirmed oxygenated monoterpene metabolites. Normalized abundances are presented to highlight differences between tissue types and among species for each standard confirmed oxygenated monoterpene metabolite including: (a) linalool; (b) citronellol; (c) geraniol; (d) myrtenol; and (e) bornyl acetate. Different letters indicate significant differences following post hoc separations between species and tissue types. Error bars represent the standard error of the mean.
Finally, four standard confirmed metabolites were identified as sesquiterpenes including β-caryophyllene, humulene, nerolidol, and α-bisabolol (Fig. 6). β-Caryophyllene was most abundant in Trojan and Nordmann fir (a), intermediate in concolor and noble fir (b), and least abundant in Fraser fir (c) (Fig. 6a). Humulene was mainly detected in the needles of all species except for noble fir (Fig. 6b). Nerolidol was most abundant in the stem tissues of concolor fir and least abundant in Nordmann and Fraser fir, but not detected in Trojan or noble fir (Fig. 6c). α-Bisabolol was very abundant in both noble fir stem and needle tissues with significantly lower abundances in Fraser, concolor and Nordmann fir and not detected in noble fir (Fig. 6d).

Standard confirmed sesquiterpene metabolites. Normalized abundances are presented to highlight differences between tissue types and among species for each standard confirmed sesquiterpene metabolite including: (a) humulene; (b) β-caryophyllene; (c) nerolidol; and (d) α-bisabolol. Different letters indicate significant differences following post hoc separations between species and tissue types. Error bars represent the standard error of the mean.
Quantitative and qualitative differences among fir Oleoresin diterpenes
The composition of oleoresin diterpenes were also evaluated to understand how defensive diterpenes might associate with more aromatic compounds along with aroma profiles. In total 48 compounds were identified as diterpene metabolites in the five species. The number of unique diterpene metabolite peaks identified within each species include: Fraser fir (27); concolor fir (24); Nordmann fir (21); Trojan fir (22); and noble fir (16) (Table S6). We also assessed potential changes in diterpene metabolite profiles over time and compared normalized peak areas using PCA. Results of the PCA demonstrated that samples from each time-point across all species consistently grouped together with no separation (Fig. S1b). To further evaluate the effect of time on diterpene sample integrity, a repeated measures ANOVA was performed which supported the time PCA results (Table S7). However, several individual diterpenes differed over time including peaks 154, 168, 170, and 172 but again did not decline in abundance chronologically indicating sample specific effects (Table S8). PCA was also used to compare similarities and differences among diterpene metabolite profiles among species and between tissue types. For needle tissues, PC1 explained 21.9% of the variance while PC2 explained 13.6% of the variance in diterpene profiles. Needle tissue diterpene profiles overlapped along the same plane for PC1 for all species. Noble fir needle samples separated from the other species in PC1 and further separated from all other species (except for concolor fir) in PC2. Variability in concolor fir diterpene profiles varied significantly along PC2 forming a large indistinct group from the other species (Fig. 3c). For stem tissue samples, PC1 explained 36.2% of the variance of diterpene metabolite profiles while PC2 explained 21.5% of the variance. Stem tissue metabolite profiles of Fraser, noble, concolor and Nordmann fir grouped together along PC1 while Trojan fir formed a separate cluster. Nordmann, noble, and concolor fir formed a separate cluster along PC2 while Fraser and Trojan fir clustered independently (Fig. 3d). Clusters observed in the PCA reflect similarities among diterpene profiles while individual metabolite abundances of specific compounds differed significantly with ANOVA (Table S9). A hierarchical cluster analysis was used to further explore the relationships observed in the PCA. A bootstrap analysis of individual needle tissue biological replicates supported the observed clusters for diterpene metabolite profiles. Trojan and Fraser needle samples clustered separately while Nordmann, concolor, and noble needle samples clustered together (Fig. S3a). Similarly, hierarchical clustering of stem tissue diterpene metabolite profiles supported the observation that concolor and noble extracts cluster together, while Fraser, Trojan, and Nordmann extracts grouped separately (Fig. S3b).
Diterpenes correlated with consumer preferred christmas tree species
To identify diterpene metabolites associated with consumer aroma liking preference we performed two-way ANOVA studies and post-hoc separations (Table S9) along with Pearson correlations using consumer liking ratings and normalized abundances for each compound (Table 1). Several metabolites were identified as being of interest following comparisons of the aromatic profiles of the most preferred versus least preferred species. Peaks of interest include the most abundant diterpene metabolites identified in the highly preferred Fraser fir (peak #’s 135, 145, 146, and 150). Putative identities for diterpene peaks of interest include thunbergene (peak 135), unknown diterpene #1 (peak 145), sclareol (peak 146), and levopimarinal (peak 150) (Table S10). Pearson correlations between the diterpene peaks and consumer ratings showed that these compounds were all strongly correlated with consumer liking. The diterpene metabolites most strongly correlated with consumer liking were those putatively identified as unknown diterpene #1 (peak 145), methyl levopimarate (peak 156), and thunbergene (peak 135) (Table 1).
Standard confirmed diterpenes
The identity of two diterpene metabolites [cis-abienol (143) and methyl abietate (161)] were confirmed using exact matching standards (Table 2). Both individual compounds varied in abundance between species and tissue types. Both compounds were most abundant in Fraser fir stem tissues and either absent (cis-abienol) or least abundant (abietic acid) in noble fir (Fig. 7). cis-Abienol was the most strongly correlated diterpene metabolite with consumer liking scores (Table 1).

Standard confirmed diterpene metabolites. Normalized abundances are presented to highlight differences between tissue types and among species for each standard confirmed diterpene metabolite including: (a) cis-abienol; and (b) abietic acid. Different letters indicate significant differences following post hoc separations between species and tissue types. Error bars represent the standard error of the mean.
Discussion
The results of the aroma test panel provide foundational information regarding consumer preferences for five of the most popular real fir Christmas tree species. Fraser fir was the most preferred species while noble fir was least preferred. Concolor fir was the most intense while Trojan fir was the least intense aroma. A total of 132 monoterpene and sesquiterpene metabolites were identified and 48 compounds were identified as diterpenes. The compound most strongly correlated with consumer liking was the putatively identified sesquiterpene β-bisabolene. Our study aimed to identify terpene metabolites that positively associated with consumer preferences which could help guide future breeding and selection efforts for Christmas tree genetic improvement.
Regional influences might have impacted aroma test panel results as participants derived primarily from the southeastern US. Regional variation in aroma responses is observed for other aromatic commodities such as wine30. In Christmas trees, noble fir is the major species sold on the west coast while Fraser fir is primarily found on the east coast. Aroma test panel participants had a stronger preference for Fraser fir which might be due in part to regional variation in liking responses. Future studies should incorporate consumers from both the east and west coast to account for regional Christmas tree preferences and variability amongst consumer demographics. Despite its parochial nature, the aroma panel data does provide early insight into consumer responses to Christmas tree aroma and is a useful first step for further panel evaluations. Of the demographics recorded, consumer purchasing habits (display a real, artificial, or no tree) had an unexpected impact on aroma preference. Consumers that displayed a fake tree had a significant negative effect on consumer liking preference. Further, consumers who display an artificial tree did not like concolor fir which also had the most intense aroma of all evaluated species. These results suggest that individuals that display an artificial tree dislike the aroma of a real tree. Therefore, future consumer aroma test panels should consider limiting participants to consumers that only display a real tree.
Terpene metabolite profiling demonstrated that more aromatic species (i.e. concolor fir) had a greater diversity of terpenes compared to the least intensely aromatic species (i.e. Trojan fir). Species with a more intense aroma were also not highly liked suggesting that specific compounds or compound blends are more important than compound diversity or overall abundance when assessing consumer preference. The results of PCA and hierarchical clustering analyses for terpene aromatic profiles supported these observations and led to unexpected outcomes. For instance, Fraser fir aroma profiles sometimes grouped with Trojan and Nordmann fir but not for every metabolite group or tissue type indicating its unique aromatic characteristics. However, Trojan and Nordmann fir profiles consistently grouped across tissue types which reflects their close phylogenetic relationship (Kew Gardens, 2023). The profiles of noble and concolor mono- and sesqui-terpene profiles often separated into different sections within the same clade. The close phylogenetic relationship between noble and concolor fir likely explain the observed similarities among diterpene aromatic profiles31.
The impact of a few compounds on aroma profile is also observed in other commodities like citrus where higher levels of the monoterpene myrcene is perceived as a strong earthy “green” scent while high levels of α-pinene contributes a ‘woody’ pine scent32. Previous studies have characterized the essential oil profiles of several fir species as extracts from various Abies spp. have been used as medicinal compounds, fragrances, and in aroma therapy33. The major volatile terpenes identified in the essential oils of east Asian fir species (e.g. A. firma, A. mariesii, A. homolepis, A. veitchii, and A. sachalinensis) include \(\:\delta\:\)-3-carene, myrcene, longifolene, and \(\:\alpha\:\)-pinene, camphene, bornyl acetate, and limonene34. Conversely, the essential oil profiles of fir species endemic to North America (e.g. A. fraseri, A. grandis, A. procera, and A. concolor) contain similar terpene metabolites at varying concentrations15,20,34,35,,21,35,36.
Our study expands on previous findings and identifies specific associations among metabolites and consumer preferences for specific Christmas tree species. The monoterpene bornyl acetate was identified in previous studies evaluating the composition of fir essential oil. We found that bornyl acetate was positively associated with consumer liking and is known to have a strong pine and herbaceous aroma14,37. Conversely, α-pinene – another metabolite that contributes a strong pine aroma—varied across all five species and was not positively associated with consumer liking. The sesquiterpene (E)-β-farnesene was strongly correlated with consumer liking and found primarily in highly rated species has been documented to play an important role in floral aroma profiles38. Finally, several putative isomers of bisabolene were identified as more abundant in highly rated species. Its notable that similar compounds have been documented to play an important role in consumer preference for a variety of aromatic plants including ginger39.
The role of diterpenes in aroma consumer liking are unknown as these compounds are not known to be aromatic17. However, two diterpenes including abietic acid and cis-abienol were more abundant in the most highly preferred species. The presence of high quantities of cis-abienol in the highly preferred Fraser fir is notable as it is known to have a very distinct and pleasant aroma40. Cis-abienol could also play a significant role in the persistence of overall aroma profiles which may explain its high correlation with preferred Christmas tree species. For instance, cis-abienol has been proposed to serve as a precursor for the industrial synthesis of the scent fixative AMBROX® which is widely used in the production of high-end fragrances41.
Correlating peak areas to consumer rating scores for each species limits us in our ability to make assertions about how consumers respond to each compound, but the data does allow us to pursue a more targeted approach in future evaluations where the specific compounds we identified can be further tested and their volatility better understood. The baseline identification of terpenoids enables us to have a starting place for future panels to explore aroma along with pest resistance because of the incredible diversity of terpenes present in these fir species so that energy is not spent on the many compounds with weak association to consumer preferences.
Future consumer panels should also focus on other aromatic compounds like esters, aldehydes, and sugar alcohols which might not be as dominant as terpenes in fir but are known to be aromatic in other crop plant systems and have been previously identified in fir5,42,43. Additionally, comparison of inter- and intra-specific variation within and among various fir species to assess how consumers from different regions respond to various Christmas tree commodities. Improved consumer metrics would enable genetic improvement efforts that incorporate aroma traits through selection and breeding for specific compounds. For example, aroma related compounds in other crop systems are selected with great efficacy as aromatic terpenes are frequently under the control of only a few genes that are highly heritable10. Additionally, terpene metabolites play crucial roles in plant defenses against insect pests which use aroma to identify host plants44. Olfaction is also hypothesized to be important for traits such as deer resilience16,17. Deer browse is another significant threat to fir Christmas tree production. Ultimately, knowledge of consumer preferred aroma profiles, specifically terpenes, will inform antagonistic efforts targeting the development of deer resilience in genetically improved fir Christmas tree species.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
Abbreviations
- CYLS:
-
Current year lateral shoots
- GC–MS:
-
Gas chromatography-mass spectroscopy
- MTBE:
-
Methyl-tert-butyl-ether
- TMS-diazomethane:
-
Tri-methyl-silyl-diazomethane
- ANOVA:
-
Analysis of variance
- Fisher’s LSD test:
-
Fisher’s least significant difference test
- PCA:
-
Principal component analysis
- PC1:
-
Principal component one
- PC2:
-
Principal component two
References
2022 Census of Agriculture: U.S. tree farms cut more than 14.5 million Christmas trees in 2022 | Economic Research Service. https://www.ers.usda.gov/data-products/charts-of-note/chart-detail?chartId=110530#:~:text=Home-,2022%20Census%20of%20Agriculture%3A%20U.S.%20tree%20farms%20cut%20more%20than,million%20Christmas%20trees%20in%202022&text=Christmas%20trees%20are%20grown%20commercially,than%2014.5%20million%20Christmas%20trees (2024).
USDA National Agricultural Statistics Service. 2017 Census of Agriculture. Complete data. https://www.nass.usda.gov/AgCensus/ (2017).
Cregg, B. Christmas tree species profile: Nordmann Fir Abies nordmanniana. Great Lakes Christmas Tree J. 2, 4 (2007).
Real Christmas Tree Board. Having a real Christmas tree in your home brings joy to everyone who sees it - and smells it. The Real Christmas Tree Board. https://realchristmastreeboard.com/press-releases/having-a-real-christmas-tree-in-your-home-brings-joy-to-everyone-who-sees-it-and-smells-it/ (2022).
Schweiterman, M. L. et al. Strawberry flavor: diverse chemical compositions, a seasonal influence, and effects on sensory perception. PLoS ONE 9 (2), e88446. https://doi.org/10.1371/journal.pone.0088446 (2014).
Liu, C. et al. Features of citrus terpenoid production as revealed by carotenoid, limonoid and aroma profiles of two pummelos (Citrus maxima) with different flesh color: terpenoid production of two different color pummelos. J. Sci. Food. Agric. 95 (1), 111–119. https://doi.org/10.1002/jsfa.6689 (2015).
Baudino, S. et al. Rose floral scent. Acta Hortic. 1232, 69–80. https://doi.org/10.17660/ActaHortic.2019.1232.12 (2019).
Fan, Z. et al. A multi-omics framework reveals strawberry flavor genes and their regulatory elements. New Phytol. 236 (3), 1089–1107. https://doi.org/10.1111/nph.18416 (2022).
Ferrão, L. F. V. et al. Terpene volatiles mediates the chemical basis of blueberry aroma and consumer acceptability. Food Res. Int. 158, 111468. https://doi.org/10.1016/j.foodres.2022.111468 (2022).
Ferrão, L. F. et al. Genome-Wide association of volatiles reveals candidate loci for blueberry flavor. New Phytol. 226 (6), 1725–1737. https://doi.org/10.1111/nph.16459 (2020).
Bhandari, U., Kumar, A., Lohani, H. & Chauhan, N. Chemical composition of essential oil of Camphor tree (Cinnamomum camphora) leaves grown in Doon Valley of Uttarakhand. J. Essent. Oil Bearing Plants 25 (3), 548–554. https://doi.org/10.1080/0972060X.2022.2086828 (2022).
Ahmad, R. S. et al. Eucalyptus essential oils. Essent. Oils 2023, 217–239. https://doi.org/10.1016/B978-0-323-91740-7.00005-0 (2023).
Celedon, J. M. & Bohlmann, J. Oleoresin defenses in conifers: chemical diversity, terpene synthases and limitations of Oleoresin defense under climate change. New Phytol. 224 (4), 1444–1463. https://doi.org/10.1111/nph.15984 (2019).
Yu, E. J., Kim, T. H., Kim, K. H. & Lee, H. J. Characterization of aroma-active compounds of Abies nephrolepis (Khinganfir) needles using aroma extract Dilution analysis. Flavour Fragr. J. 19 (1), 74–79. https://doi.org/10.1002/ffj.131 (2004).
Bowman, J. M. et al. Extraction method for the isolation of terpenes from plant tissue and subsequent determination by gas chromatography. Microchem. J. 56 (1), 10–18. https://doi.org/10.1006/mchj.1996.1422 (1997).
Connolly, G. et al. Deer browsing of Douglas-fir trees in relation to volatile terpene composition and in vitro fermentability. For. Sci. 26 (2), 179–193 (1980).
Vourc’h, G., Martin, J. L., Duncan, P., Escarré, J. & Clausen, T. P. Defensive adaptations of Thuja plicata to ungulate browsing: a comparative study between Mainland and Island populations. Oecologia 126 (1), 84–93 (2001). https://www.jstor.org/stable/4222823
Bohlmann, J., Phillips, M., Ramachandiran, V., Katoh, S. & Croteau, R. cDNA cloning, characterization, and functional expression of four new monoterpene synthase members of the Tpsd gene family from grand fir (Abies grandis). Arch. Biochem. Biophys. 368 (2), 232–243. https://doi.org/10.1006/abbi.1999.1332 (1999).
Bohlmann, J. & Croteau, R. Diversity and variability of terpenoid defences in conifers: molecular genetics, biochemistry and evolution of the terpene synthase gene family in grand Fir (Abies Grandis). In Novartis Foundation Symposia (eds. Chadwick, D. J. & Goode, J. A.) 132–149 (Wiley & Sons, 2007). https://doi.org/10.1002/9780470515679.ch9.
Adams, R., Kauffmann, M. & Callahan, F. The leaf essential oil of Abies grandis (Doug. Ex D. Don) Lindl. (Pinaceae): revisited 38 years later. Phytologia 97, 1 (2015).
Swor, K., Poudel, A., Rokaya, A., Satyal, P. & Setzer, W. N. Foliar essential oil of white Fir (Abies concolor (Gordon) Lindl. Ex Hildebr.): chemical composition and enantiomeric distribution. Nat. Prod. Commun. 17 (3), 1934578X2210869. https://doi.org/10.1177/1934578X221086901 (2022).
Whitehill, J. G. A. et al. Functions of stone cells and Oleoresin terpenes in the conifer defense syndrome. New Phytol. 221 (3), 1503–1517. https://doi.org/10.1111/nph.15477 (2019).
Lewinsohn, E., Savage, T. J., Gijzen, M., & Croteau, R. Simultaneous analysis of monoterpenes and diterpenoids of Conifer Oleoresin. Phytochem. Anal. 4(5), 220–225. https://doi.org/10.1002/pca.2800040506 (1993).
R Core Team. _R: A Language and Environment for Statistical Computing_. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/ (2003).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear Mixed-Effects models using lme4. J. Stat. Softw. 67 (1), 1–48. https://doi.org/10.18637/jss.v067.i01 (2015).
Signorell, A. et al. DescTools: tools for descriptive statistics. R package version 0.99.23 (2017).
Nakazawa, M. Functions for Medical Statistics Book with some Demographic Data_. R package version 0.7.5. https://CRAN.R-project.org/package=fmsb (2023).
Efron, B., Halloran, E. & Holmes, S. Bootstrap confidence levels for phylogenetic trees. Proc. Natl. Acad. Sci. U. S. A. 93, 13429–13434 (1996).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer, 2016).
Ristic, R. et al. P. Wine-related aromas for different seasons and occasions: hedonic and emotional responses of wine consumers from Australia, UK and USA. Food Qual. Prefer. 71, 250–260. https://doi.org/10.1016/j.foodqual.2018.07.011 (2019).
Xiang, Q., Wei, R., Zhu, Y., Harris, A. & Zhang, X. New infrageneric classification of Abies in light of molecular phylogeny and high diversity in Western North America. J. Syst. Evol. 56 (5), 562–572. https://doi.org/10.1111/jse.12458 (2018).
Miyazaki, T., Plotto, A., Baldwin, E. A., Reyes-De-Corcuera, J. I. & Gmitter, F. G. Jr Aroma characterization of Tangerine hybrids by gas-chromatography-olfactometry and sensory evaluation. J. Sci. Food. Agric. 92 (4), 727–735. https://doi.org/10.1002/jsfa.4663 (2012).
Yu, B. et al. Composition analysis and tyrosinase inhibitory activity of Cinnamomum cassia Presl leaf hydrosol and Cymbopogon citratus (DC.) Stapf leaf hydrosol. J. Food Process. Preserv. 46, 11. https://doi.org/10.1111/jfpp.17058 (2022).
Satou, T. et al. Components of essential oils extracted from leaves and shoots of Abies species in Japan. Chem. Biodivers. 8 (6), 1132–1138. https://doi.org/10.1002/cbdv.201000254 (2011).
Swor, K., Poudel, A., Rokaya, A., Satyal, P. & Setzer, W. N. Foliar essential oil of white Fir (Abies concolor (Gordon) Lindl. Ex Hildebr.): chemical composition and enantiomeric distribution. Nat. Prod. Commun. 17 (3), 1934578X2210869. https://doi.org/10.1177/1934578X221086901 (2022).
Ankey, E., Satyal, P. & Setzer, W. Chemical compositions and enantiomeric distributions of leaf essential oils of three conifers from Oregon. Am. J. Essent. Oils Nat. Prod. 9 (2), 7–14 (2019).
Yu, E. J., Kim, T. H., Kim, K. H. & Lee, H. J. Characterization of aroma-active compounds of Abies nephrolepis (Khingan fir) needles using aroma extract Dilution analysis. Flavour. Fragr. J. 19, 74–79. https://doi.org/10.1002/ffj.1314 (2004).
Zheng, C. H., Kim, T. H., Kim, K. H., Leem, Y. H. & Lee, H. J. Characterization of potent aroma compounds in Chrysanthemum coronarium L. (Garland) using aroma extract Dilution analysis. Flavour Fragr. J. 19 (5), 401–405. https://doi.org/10.1002/ffj.1447 (2004).
Novak, J. & Blüthner, W. D. Medicinal, Aromatic and Stimulant Plants (Springer, 2020).
Cheng, T., Zhao, G., Xian, M. & Xie, C. Improved cis-Abienol production through increasing precursor supply in Escherichia coli. Sci. Rep. 10 (1), 16791. https://doi.org/10.1038/s41598-020-73934-z (2020).
Zerbe, P. et al. Bifunctional cis-abienol synthase from Abies balsamea discovered by transcriptome sequencing and its implications for diterpenoid fragrance production. J. Biol. Chem. 287 (15), 12121-31. https://doi.org/10.1074/jbc.M111.317669 (2012).
Woosley, R. S. & Butcher, D. J. Chemical analysis of an endangered conifer: environmental laboratory experiments. J. Chem. Educ. 75 (12), 1592–1594 (1998). https://proxying.lib.ncsu.edu/index.php/login?url=https://www.proquest.com/scholarly-journals/chemical-analysis-endangered-conifer/docview/212011323/se-2.
Ferreira, V., Jarauta, I., López, R. & Cacho, J. Quantitative determination of Sotolon, maltol and free Furaneol in wine by solid-phase extraction and gas chromatography–ion-trap mass spectrometry. J. Chromatogr. A 1010 (1), 95–103. https://doi.org/10.1016/S0021-9673(03)00963-4 (2003).
Whitehill, J. G. A., Bohlmann, J. & Krokene, P. Forest insect-plant interactions. In Forest Entomology and Pathology, vol. 1 (Springer, 2023).
Acknowledgements
We thank NC State undergraduate student workers Lauren Stevens and Kaley Patton for assistance with sample process and terpene extract preparation. GC-MS work was supported by Mary Madera in the Zerbe lab at UC Davis.
Author information
Authors and Affiliations
Contributions
W.C.B., C.S., G.C., K.N., C.L., P.Z., and J.G.A.W. designed research; W.C.B., A.C., K.N., J.K. and J.G.A.W. performed experiments; W.C.B., G.W., P. Z. and J.G.A.W. analyzed data and interpreted the results. W.C.B. and J.G.A.W. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.


Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Baldwin, W.C., Chiang, A., Wyatt, G. et al. Distinct terpene metabolite blends serve as core drivers of consumer aroma preferences in fir Christmas tree species. Sci Rep 15, 18500 (2025). https://doi.org/10.1038/s41598-025-01505-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-01505-1

{kind=link}
{kind=link}