
Abstract
Insect communities serve as vital components of ecosystems and environmental indicators. To assess insect community composition and diversity across Inner Mongolia’s grassland types, a systematic survey was conducted between June and August of 2022–2023 using the sweep net method. A total of 38,200 insects were identified, belonging to 4 orders, 69 families, and 518 species. Herbivorous insects dominated the communities, accounting for 98.66% of the total individual count. Dominant species composition varied among grassland types. Significant differences in alpha diversity were observed across habitats. PCoA spatially delineated distinct community structures. Meadow and typical steppes exhibited similar insect communities, while desert and steppe desert also clustered closely. This reflected the influence of environmental gradients on insect distribution. LEfSe analysis identified specific taxa driving these macro-scale differentiations. This differentiation underscores the necessity of tailored insect management strategies. These findings establish theoretical foundations for insect diversity monitoring and contribute to the sustainable use of grassland resources.
Similar content being viewed by others
Introduction
The Inner Mongolia Autonomous Region, situated on China’s northern frontier, is a critical ecological stronghold where grasslands serve as a primary environmental and economic resource1. These grasslands constitute approximately one-fourth of China’s total grassland area, playing a pivotal role in ecological balance, agricultural productivity, and local livelihoods2,3. The region’s geographical and climatic diversity has created a distinct zonal distribution of grassland types, transitioning sequentially from meadow steppe in the east to desert in the west as precipitation gradually decreases4. This gradient significantly influences the local ecological environment, agricultural systems, and herding practices.
The grasslands of Inner Mongolia are characterized by their abundant and diverse forage species, such as Leymus chinensis, Agropyron cristatum, and Elymus dahuricus, which are well-adapted to the region’s climatic and soil conditions. These grasslands form the foundation of China’s animal husbandry industry, providing a rich base for livestock rearing. Beyond economic importance, they also fulfill critical ecological functions, including windbreak and sand fixation, soil and water conservation, water source protection, climate regulation, and biodiversity preservation5,6,7.
Insects, as one of the most species-rich groups in the animal kingdom, play a crucial role in sustaining the ecological functions of grasslands. Characterized by their small body size, high abundance, wide distribution, and sensitivity to environmental changes8,9, insects are vital components of biodiversity and essential for maintaining ecosystem functionality10. As consumers and decomposers in ecosystems, they uphold the fundamental processes of material cycling. However, the proliferation of herbivorous insects poses significant threats to ecological stability and agricultural production11. Key herbivorous insect groups, including Oedipodidae, Nymphalidae, Chrysomelidae, and Pentatomidae, exhibit diverse morphologies and behaviors, causing varied types of damage.
Insect occurrence varies by region and species, typically peaking between late spring and early summer, particularly in July and August, coinciding with their peak activity and reproduction. Understanding the composition and diversity of insect communities across different grassland types is critical for devising effective monitoring and control strategies. This study investigates the pest communities in five grassland types in Inner Mongolia: meadow steppe, typical steppe, desert steppe, steppe desert, and desert. We analyzed the species composition, community structure, and diversity indices of insects in these grassland types, aiming to provide valuable scientific insights for insect management and contribute to the sustainable development of grassland ecosystems in Inner Mongolia.
Results
Insect community composition
A total of 38,200 insects were collected during the survey, spanning 4 orders, 69 families, and 518 species. Among these, Coleoptera had the highest species count, with 195 species (37.65% of the total), and accounted for 20.06% of individuals (7663). Orthoptera, although having fewer species (148, 28.57%), dominated in individual count, comprising 74.29% of all individuals (28,377). In contrast, Hemiptera and Lepidoptera exhibited lower species and individual counts (Table 1). A core set of 28 species was found across all five grassland types. Meadow steppes and typical steppes showed the greatest species overlap, sharing 39 species, followed by deserts and steppe deserts with 37 shared species. Unique species were most abundant in typical steppes (Fig. 1).

Unique and shared insect species across five grassland types: D, desert; SD, steppe desert; DS, desert steppe; TS, typical desert; and MS, meadow steppe.
Insect community structure was dominated by herbivorous groups in terms of both species and individuals, accounting for 481 species (92.86% of the total species) and 37,689 individuals (98.66% of the total count). Non-herbivorous groups (predatory, coprophagous, and omnivorous) collectively comprised 37 species (7.14%) and 511 individuals (1.34%) (Table 2) (Supplementary Table 1).
Dominant and common insect groups in five grassland types
Heatmap analysis revealed that Leiometopon simyrides and Anatolica ningxiana exhibited higher population densities in desert habitats compared to the other four grassland ecosystems, with both species identified as common species in desert and steppe desert. The insect community in typical steppes was characterized by the dominant species Dasyhipps barbipes and Myrmeleotettix palpalis (Orthoptera), which showed high abundance. Notably, the grasshopper Oedaleus decorus demonstrated significant ecological dominance across meadow, typical and desert steppes. In desert steppe, Galeruca daurica reached a relative abundance of 18.89%, establishing it as the dominant species in this region (Fig. 2). A complete list of dominant and common species across five grassland types is provided in the Supplementary Tables 2 and 3.

Insect communities at the species level across the five grassland types. Color scale (-1.5 to 1.5) indicates standardized species abundance. Negative values (blue) indicate below-average levels.
At the family level, insects from Oedipodidae and Chrysomelidae exhibited extremely high relative abundance in all sampling sites. Following closely were Gomphoceridae and Arcypteridae, which were very abundant in desert, typical and meadow steppes. Additionally, insects from Tenebrionidae and Curculionidae had higher relative abundance in deserts and steppe deserts. Families such as Pyrgomorphidae were exclusive to meadow and typical steppes. Psyllidae, on the other hand, was found only in desert and steppe desert grasslands (Fig. 3). A complete list of dominant and common families across five grassland types is provided in the Supplementary Tables 4 and 5.

Insect communities at the family level across the five grassland types.
Diversity analysis of insect community
Significant variations in alpha diversity were observed among grassland types (Fig. 4) (Supplementary Table 6). The Shannon diversity index was highest in desert, steppe desert, and typical steppe (3.46, 3.69, and 3.57, respectively), while meadow steppe and typical steppe showed the lowest values. The Simpson diversity reached its maximum in steppe desert (0.96). Typical steppe exhibited the highest richness index (140.50) and Chao1 index (221.65). Alpha diversity analysis at the family level was detailed in the Supplementary Fig. 1.

Figures (A, B) represent the alpha diversity (Shannon index, Simpson index, Richness index, Chao1 value) of insect communities in different grassland types.
PCoA demonstrated distinct clustering of insect communities, with desert and steppe desert samples clustering in the negative PCoA1 axis, while meadow steppes, typical steppes, and desert steppes clustered in the positive axis, likely reflecting environmental gradients among grassland types (Fig. 5).

Principal component analysis of insect communities in five grassland types.
The hierarchical partitioning plot demonstrated the independent contributions of different environmental factors to insect distribution (Fig. 6) (Supplementary Tables 7–8). SMT exhibited the most significant influence on insect distribution, with an independent contribution value exceeding 30%, which was substantially higher than those of other factors. The contributions of SADP, STP, and SMRH were comparable, ranging from 15 to 20%. In contrast, SFVC showed a relatively minor impact, with an independent contribution of approximately 5%.

Effects of different environmental factors on insect distribution. Values on the y-axis represent the independent contribution percentage of each environmental variable. SMT, summer mean temperature; SADP, summer average daily precipitation; STP, summer total precipitation; SMRH, summer mean relative humidity; SFVC, summer fractional vegetation cover.
Taxonomic differentiation
From five types of grasslands, a total of 4 orders, 21 families, and 57 genera were identified, all showing significant differences among the groups classified. Desert had the highest number of differential taxa, with 14 genera from Coleoptera, including Tenebrionidae, contributing most significantly, with an LDA value of 4.68, P < 0.05. Meadow steppes were dominated by Orthoptera insects, with Arcypteridae species showing the largest effect size (LDA 4.48, P < 0.05). In typical steppes, Orthoptera insects like Dasyhipps were the key differentiators (Fig. 7) (Supplementary Fig. 2 and Supplementary Table 9). These results emphasize the distinct ecological roles and adaptations of insect taxa in each grassland type.

Linear discriminant analysis of effect size (LEfSe) analysis of the insect community. The levels of phylum, class, order, family and genus are arranged from the inside to the outside. The yellow circle represents the taxa that did not significantly differ among the grasslands.
Discussion
As a crucial terrestrial ecosystem in Inner Mongolia, grasslands not only support diverse ecological landscapes but also harbor abundant insect resources. This study, through systematic surveys of five grassland types (desert, desert steppe, steppe desert, typical steppe, and meadow steppe) in Inner Mongolia, revealed that each grassland type maintains a distinct insect community structure. Among them, the insect communities of meadow steppe and typical steppe exhibited high similarity, with herbivorous insects dominating numerically. In contrast, desert and desert steppe formed significantly different assemblages dominated by Coleoptera. This differentiation was closely correlated with environmental factors, particularly temperature, which was the most critical driver of insect community composition, diversity indices (such as the Shannon index, Simpson index, richness index, and Chao index), and spatial differentiation patterns. Given that the Inner Mongolian grasslands serve as a natural ecological security barrier in northern China, elucidating insect diversity in this region holds significant importance for biodiversity conservation and ecosystem management.
The dominant species in a community refer to the most prevalent taxa that exert the greatest influence on their habitat. Among the five grassland insect communities, the dominant species include Galeruca daurica, Oedaleus decorus, Dasyhipps barbipes, and Myrmeleotettix palpalis. Galeruca daurica predominantly inhabits arid or semi-arid steppe regions, with both adults and larvae feeding on plant leaves in destructive aggregations. The remaining three Orthoptera species, consistent with the findings reported by Chen et al., constitute severe pests in Inner Mongolia grasslands12,13. Oedaleus decorus represents a typical Poaceae feeder that exhibits minimal forage selectivity under high population densities. Dasyhipps barbipes preferentially feeds on Leymus chinensis, Agropyron cristatum, and Stipa grandis, posing significant threats to early-stage grass growth due to its early occurrence period. Myrmeleotettix palpalis additionally consumes Fabaceae, Asteraceae, and Cyperaceae plants, capable of completely denuding pastures during outbreaks13,14. This investigation revealed that herbivorous insects numerically exceeded those of predatory, coprophagous, and omnivorous groups, indicating their predominant status in Inner Mongolia grassland insect communities. This dominance may be attributed to the ecosystem’s provision of abundant food resources and diverse habitats that facilitate herbivorous insect proliferation15,16. While direct herbivory on leaves, stems, and roots structurally damages plants and impairs their growth, it simultaneously sustains higher trophic levels. Predatory insects such as Coccinellidae indirectly maintain vegetation health through Aphididae predation, demonstrating interaction mechanisms with potential applications in grassland management.
This study demonstrated that the insect communities in meadow steppe exhibited relatively low Shannon diversity index and Simpson dominance index, which may partially stem from temperature-driven ecological effects. Hierarchical partitioning further revealed that temperature was the key factor explaining variation in insect diversity. The lower environmental temperatures in northeastern regions may impose limitations on insect diversity. As ectothermic organisms, insects exhibit high sensitivity to temperature fluctuations. Within a certain range, an increase in temperature can enhance insect species diversity15,17. However, excessively high temperatures exceeding their tolerance thresholds may disrupt key physiological processes such as migration, reproduction, and development, ultimately driving insects to migrate toward higher latitudes or altitudes18,19. Conversely, extremely low temperatures can impede the completion of their life cycles, thereby reducing colonization opportunities. For instance, Kiritani et al. investigated the overwintering mortality rates of Nezara viridula and Halyomorpha halys adults, revealing a 15% decline in mortality per 1 °C temperature increase20. Due to climate warming, Ceutorhynchus obstrictus, previously unable to survive winter conditions, has now successfully established populations in northern Canada, causing damage to oilseed crops21. Additionally, increases in vegetation coverage, humidity, and precipitation typically correlate with higher primary productivity and resource availability (moisture, light, and nutrients)22,23,24. However, resource enrichment may intensify interspecific competition, resulting in the dominance of competitive species. The expansion of dominant species reduces community evenness. Although species richness increases, the high proportion of dominant species leads to declines in the Shannon and Simpson indices, as both metrics are sensitive to evenness. Furthermore, grazing is a prevalent anthropogenic disturbance in grassland ecosystems, affecting the grassland ecosystems of Inner Mongolia25,26. Research indicates that its impact intensity varies by grassland type, with meadow steppe exhibiting the most significant response, followed by typical steppe and desert steppe27. Moreover, meadow steppe possesses abundant forage resources and often faces greater anthropogenic interference, particularly from herders whose families have traditionally relied on grazing for generations, making cattle and sheep their primary, or even sole, income source. Some herders are unable to reduce stocking rates to protect the grasslands because they believe they must raise more livestock to sustain their livelihoods and repay loans26. Some herders practice year-round grazing on their contracted pastures without seasonal rotation, leading to a gradual decline in the productivity of meadow steppe and a concomitant reduction in aboveground biomass28,29. These changes undoubtedly exert both direct (physical destruction) and indirect (through altered habitats and resources) impacts on insect communities30.
Furthermore, temperature and precipitation can also indirectly drive the spatial distribution of insects by altering vegetation characteristics31. For instance, research by Gaona, F.P. et al. demonstrated that precipitation and temperature shape insect communities indirectly through their effects on vegetation32. Vegetation characteristics have been identified as key drivers of species diversity and distribution33. Research by Parker, D.M. et al. indicated that if a single vegetation management practice dominates the landscape, it may lead to a simplification of biological communities34. Research by Zhao Shulan et al. on the insect communities at Tianjin Binhai International Airport also fully demonstrated that vegetation communities significantly affect the composition and diversity of insect communities35. In grassland ecosystems, insects and plants interact and are inseparable36. Plants provide habitats and shelter for insects, and their community characteristics also influence insect community characteristics37. In the eastern meadow steppe and typical steppe, relatively abundant precipitation and lower temperatures sustain high productivity of Poaceae plants (such as Leymus chinensis and Stipa spp.), forming continuous, dense grass layers37,38. This type of vegetation provides dual habitat support for herbivorous Orthoptera insects: on the one hand, the high biomass of Poaceae plants serves as their core food source; on the other hand, the dense grass layer creates physical shelter, significantly reducing predation risk. In contrast, in the western arid regions, scarce precipitation combined with high temperatures intensifies water evaporation, suppresses the growth of herbivorous plants, and selects for super-xerophytic shrubs (such as Haloxylon ammodendron and Nitraria tangutorum)38. This desert vegetation aligns with the survival strategies of Coleoptera insects: their specialized hardened integument effectively reduces water loss, and their feeding habits adapt to consuming low-nutrient, high-secondary-metabolite shrub tissues or detritus; simultaneously, sand grains accumulating at the base of shrubs provide larvae with subterranean shelters, enabling them to avoid extreme temperature stress on the surface. LEfSe analysis validated this differentiation pattern at the macro scale. Based on this, formulating targeted insect management strategies is crucial. For example, intensified monitoring and targeted control of locust species, such as D. barbipes and O. decorus, are critical for meadow and typical steppes. Conversely, desert insect management should focus on Coleoptera, particularly Tenebrionidae and Chrysomelidae, which exhibit high reproductive capacity and potential for rapid population growth.
Insect communities serve as reliable indicators of grassland ecosystem health39. By monitoring changes in insect diversity and abundance, subtle shifts in the ecosystem can be detected in a timely manner, providing scientific guidance for ecological conservation and restoration efforts, and promoting sustainable development in the region. Although the monthly sampling times across grassland types were not fully synchronized, which could potentially affect comparisons of insect diversity among different grassland types, no extreme weather events occurred during any monthly sampling period and environmental conditions remained relatively stable. This consistency in climatic conditions may have substantially mitigated the potential impact of sampling time differences on the comparability of insect diversity data. However, given the significant annual and seasonal changes in insect composition and diversity in arid and semi—arid areas, future studies should minimize sampling time spans. Additionally, supplementary survey methods will be employed in future research to refine the study of insect diversity in Inner Mongolia, thereby providing more comprehensive and precise scientific evidence for the conservation and management of grassland ecosystems.
Conclusion
This study conducted a comprehensive investigation of insect diversity across grassland types in Inner Mongolia. Preliminary analysis of insect community composition revealed that Orthoptera dominated in abundance, with herbivorous insects representing a high proportion. Further comparisons at species and family levels demonstrated distinct compositional differences in insect communities among grassland types, with alpha diversity indices showing significant variations across habitats. Taxonomic differentiation identified key insect genera unique to specific habitats, emphasizing the need for tailored insect management strategies. For example, Orthoptera control should focus on meadow steppes and typical steppes, while Coleoptera management is critical in desert regions. These findings reflect habitat-specific relationships with insect community structure. Future research should expand taxonomic coverage and integrate multi-method long-term monitoring to strengthen scientific support for grassland ecosystem conservation and management strategies.
Methods
Study area
The Inner Mongolia Autonomous Region, located in northern China (97° 12’ E to 126° 04’ E, 37° 24’ N to 53° 23’ N), borders Mongolia and spans a vast east-to-west distance. The region crosses arid, semi-arid, and semi-humid zones, making it geographically and climatically representative of northern China14. Most of Inner Mongolia experiences a mid-temperate continental monsoon climate, with average annual temperatures ranging from 3 to 6 °C and precipitation decreasing from 400 mm in the northeast to 300 mm in the southwest. This climatic and geographic variation has created distinct vegetation zones transitioning from meadow steppe in the northeast to desert in the southwest4,14. Meadow steppes primarily consist of Leymus chinensis and Stipa baicalensis communities, while typical steppes are dominated by Stipa grandis and Stipa krylovii. Desert steppes include Stipa breviflora, Stipa glareosa, and Stipa klemenzii communities. Steppe deserts are characterized by drought-tolerant shrubs and semi-shrubs like Ammopiptanthus spp. and Nitraria spp., whereas deserts support extremely drought-resistant vegetation such as Haloxylon ammodendron and Hippophae spp.14,40,41.
Sample protocol
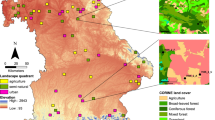
The survey was conducted from June to August in 2022 and 2023, covering the peak insect activity season. The study encompassed five grassland types, with 10 sampling points established for each type, totaling 50 sampling points across the entire study area (Supplementary Table 10). Sampling at each point was conducted by professionals from local forestry bureaus once monthly in June, July, and August each year; each sampling session was completed over a 15-day period selected at their discretion within the respective month. GPS devices recorded the latitude, longitude, and altitude of each sampling point (Fig. 8). The shapefile (SHP) data of the Inner Mongolia Region were obtained from Tianditu (https://cloudcenter.tianditu.gov.cn/administrativeDivision), with the geographic information approval number GS (2024) 0650. The study area map was generated using ArcMap 10.8 following standard cartographic procedures. Sweeping net collection was employed at a frequency of 3–4 sweeps per second, covering a 5-m radius around the transects. Collected insects from the orders Hemiptera, Lepidoptera, Coleoptera, and Orthoptera were preserved in 75% alcohol or triangular paper bags. Insect identification relied on standard entomological references, including A Guide to Insects in China42, A Guide to the Ecology of Insects in China43, and A Guide to 100 Species of Grasshoppers in China44. Specimens were classified by order, family, and species. Additionally, the collected insects were classified into herbivorous, predatory, parasitic, and omnivorous groups based on their feeding habits. The comprehensive survey and identification approach ensured accurate characterization of insect communities across different grassland types, providing robust data for ecological and biodiversity analyses.

Study area and sampling point setting: D, dessert; SD, steppe desert; MS, meadow steppe; TS, typical steppe; DS, desert steppe.
Data processing and analysis
Insect species were classified into three categories based on their relative abundance: dominant groups (species constituting more than 10% of the total catch), common groups (1%–10%), and rare groups (less than 1%). Alpha diversity indices45, including the Shannon index (effective number of common species), Richness index (total species count), Simpson diversity index (effective number of dominant species), and Chao index (predicted total species), were calculated to assess community diversity. Bray–Curtis distance is a commonly used index that takes into account the differences in species abundance. In this study, Beta diversity was evaluated using the Bray–Curtis index46 to measure dissimilarity between insect communities, with principal coordinate analysis (PCoA) employed to visualize these differences. The experimental data were organized using Microsoft Excel 2021 and analyzed for alpha diversity indices using the ‘diversity’ function and ‘specnumber’ function in the Vegan package of the R software (version 4.4.1)47,48. Based on the analysis results, figures were plotted using the ‘ggbarplot’ function, ‘ggboxplot’ function, ‘ggdotplot’ function, and ‘ggviolin’ function in the ggpubr package49. Additionally, the picante package50 was required. The ‘vegdist’ function and ‘wcmdscale’ function in the Vegan package were used to calculate Bray–Curtis distances and perform weighted principal coordinate analysis, respectively, to characterize the beta diversity of pests in different types of grasslands and test the dissimilarity of insect communities. Complex data visualization was completed using the ggplot2 package51.
The Venn diagrams were generated using the VennDiagram package and supplemented with the MicroBioInf platform (http://bioinformatics.com.cn/)52. Heatmaps were created using the pheatmap package. Raw species data were normalized using Z-score standardization. The color scale range was set from -1.5 to 1.5. Stacked plot was constructed using the ggplot2 package.
Environmental factors refer to various elements in the environment that directly or indirectly influence the growth and distribution of organisms, such as temperature, precipitation, vegetation and humidity. Data were extracted from the websites https://developers.google.com/earth-engine/datasets/catalog/MODIS_061_MCD15A3H and https://doi.org/https://doi.org/10.5067/MODIS/MOD13Q1.061. Hierarchical partitioning analysis was performed using the hier.part package to visually represent the independent contributions of environmental variables and identify the variable with the greatest independent influence on the response variables.
Linear discriminant analysis effect size (LEfSe) identified taxa with significant abundance differences across grassland types, with linear discriminant analysis (LDA) estimating the impact size of each genus using a threshold of 3.0 and a stringent one-against-one comparison for robustness53,54. This comprehensive framework enabled a detailed evaluation of insect community diversity, distribution, and taxonomic differentiation, offering valuable insights into the ecological dynamics of Inner Mongolia’s grasslands.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Han, X. et al. The grasslands of Inner Mongolia: A special feature. Rangel. Ecol. Manage. 62(4), 303–304. https://doi.org/10.2111/09-002.1 (2009).
Bork, E. W. et al. Light to moderate long-term grazing enhances ecosystem carbon across a broad climatic gradient in northern temperate grasslands. Sci. Total Environ. 894, 164978. https://doi.org/10.1016/j.scitotenv.2023.164978 (2023).
Hua, Y. C., Sa, R. L. & Wang, B. Spatial and temporal variation of grassland NPP and its driving forces in Inner Mongolia. J. Desert Res. 41(05), 130–139 (2021).
Le, R. W. et al. The spatiotemporal changes in aboveground biomass of Inner Mongolia grasslands from 2000 to 2019. J. Univ. Chin. Acad. Sci. 39(01), 21–33 (2022).
Chen, L. et al. Grassland ecosystems resilience to drought: The role of surface water ponds. Land Degrad. Dev. 34(7), 1960–1972. https://doi.org/10.1002/ldr.4581 (2023).
Wardle, D. A. & Grime, J. P. Biodiversity and stability of grassland ecosystem functioning. Oikos 100(3), 622–623. https://doi.org/10.1034/j.1600-0706.2003.12283.x (2003).
Dong, S. K. et al. Analysis of grassland zoning and functions in China under the background of new era ecological civilization. J. Nat. Resour. 37(03), 568–581 (2022).
Banks, J. E., Reyes-Gallegos, E. & Njoroge, L. Insect diversity and abundance measured two decades after farmland conversion to forest using different human-assisted restoration strategies in Gede, Kenya. Pan-Pacific Entomol. 98(1), 28–41. https://doi.org/10.3956/2022-98.1.28 (2022).
Ramalingam, R. & Dharma Rajan, P. Habitat quality and edge area of fragments determine insect diversity in a heavily used landscape: Implications for forest landscape restoration. Biotropica 54(6), 1440–1456. https://doi.org/10.1111/btp.13027 (2022).
Stork, N. E. How many species of insects and other terrestrial arthropods are there on earth?. Annu. Rev. Entomol. 63, 31–45 (2018).
Liu, S. et al. Impacts of livestock production on grassland grasshopper disasters. Agronomy-Basel 14(4), 820. https://doi.org/10.3390/agronomy14040820 (2024).
Chen, C., Zhang, G., Liang, Y. & Wang, H. Impacts of locust feeding on interspecific relationships and niche of the major plants in Inner Mongolia grasslands. Global Ecol. Conserv. 51, e02913. https://doi.org/10.1016/j.gecco.2024.e02913 (2024).
Wu, H. H. A study on the food adaptability of dominant grasshopper species in typical grasslands of Inner Mongolia. (2012).
Fu, Z. Y. A Study on the spatial differentiation characteristics and ecological stability of vegetation in Inner Mongolia grasslands. (2022).
Beirao, M. V., Neves, F. S. & Fernandes, G. W. Climate and plant structure determine the spatiotemporal butterfly distribution on a tropical mountain. Biotropica 53(1), 191–200. https://doi.org/10.1111/btp.12860 (2021).
Houghton, D. C. et al. The distinctive plant and insect assemblages of an experimental forest in Northern Lower Michigan (United States). Environ. Entomol. 51(4), 716–727. https://doi.org/10.1093/ee/nvac035 (2022).
Gimesi, L. et al. The effect of climate change on the phenology of moth abundance and diversity. Appl. Ecol. Environ. Res. 10(3), 349–363. https://doi.org/10.15666/aeer/1003_349363 (2012).
Chen, I. C. et al. Elevation increases in moth assemblages over 42 years on a tropical mountain. Proc. Natl. Acad. Sci. USA 106(5), 1479–1483. https://doi.org/10.1073/pnas.0809320106 (2009).
Hickling, R., Roy, D. B., Hill, J. K. & Thomas, C. D. A northward shift of range margins in British Odonata. Glob. Change Biol. 11(3), 502–506. https://doi.org/10.1111/j.1365-2486.2005.00904.x (2005).
Kiritani, K. The low development threshold temperature and thermal constant in insects, mites and nematodes in Japan. Miscellaneous Publications of the National Institute of Agro-Environ. Sci. 21, 1–72 (1997).
Carcamo, H., Herle, C., Otani, J. & McGinn, S. Cold hardiness and overwintering survival of the cabbage seedpodweevil, Ceutorhynchus obstrictus. Entomol. Exp. Appl. 133(3), 223–231 (2009).
Fornoff, F., Staab, M., Zhu, C. D. & Klein, A. M. Multi-trophic communities re-establish with canopy cover and microclimate in a subtropical forest biodiversity experiment. Oecologia 196(1), 289–301. https://doi.org/10.1007/s00442-021-04921-y (2021).
Schoneberg, T. et al. Pruning of small fruit crops can affect habitat suitability for Drosophila suzukii. Agric. Ecosyst. Environ. 294, 106860. https://doi.org/10.1016/j.agee.2020.106860 (2020).
Chang, X. N., Gao, H. J., Chen, F. J. & Zhai, B. P. Effects of environmental humidity and precipitation on insects. Chin. J. Ecol. 04, 619–625 (2008).
Debano, S. J. Effects of livestock grazing on aboveground insect communities in semi-arid grasslands of southeastern Arizona. Biodivers. Conserv. 15(8), 2547–2564. https://doi.org/10.1007/s10531-005-2786-9 (2006).
Fang, X. N. & Wu, J. G. Causes of overgrazing in Inner Mongolian grasslands: Searching for deepleverage points of intervention. Ecolo. Soc. 27(1), https://doi.org/10.5751/es-12878-270108 (2022).
Zhang, Y. X., Du, Y. L., Liu, X. Q. & Ma, S. K. Impacts of grazing activities on the grassland ecosystem in Inner Mongolia. Acta Ecologica Sinica. (10), 1–12. https://doi.org/10.20103/j.stxb.202408161941 (2025).
Chen, Y. et al. Response of Leymus chinensis functional traits to varying grazing intensities in temperate meadow steppe. Grassland Turf. 43(02), 143–151. https://doi.org/10.13817/j.cnki.cyycp.2023.02.017 (2023).
Wang, Y. X. & Cao, J. M. Approaches to addressing overgrazing in Inner Mongolian grasslands. Ecol. Econ. (07), 58–60. (2007).
Wang, N., Song, X., Wang, J. & Wang, L. Impacts of different fencing periods and grazing intensities on insect diversity in the desert steppe in Inner Mongolia. Front. Ecol. Evolut. 10, 1021677. https://doi.org/10.3389/fevo.2022.1021677 (2022).
Zhang, Y. & Feng, G. Distribution patterns of insect species diversity in Inner Mongolia and their mechanisms. Biodiversity Sci. 26(07), 701–706 (2018).
Gaona, F. P. et al. Climate-driven vegetation characteristics shape phytophagous and carnivorous insect diversity in South African savannahs. J. Biogeography 52(4), e15076. https://doi.org/10.1111/jbi.15076 (2025).
Tews, J. et al. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 31(1), 79–92. https://doi.org/10.1046/j.0305-0270.2003.00994.x (2004).
Parker, D. M., Stears, K., Olckers, T. & Schmitt, M. H. Vegetation management shapes arthropod and bird communities in an African savanna. Ecol. Evolut. 13(3), 9880. https://doi.org/10.1002/ece3.9880 (2023).
Zhao, S. L., Li, Z. F. & Duo, L. Effects of vegetation management on the composition and diversity of the insect community at Tianjin Binhai International Airport, China. Bull. Entomological Res. 111(5), 553–559. https://doi.org/10.1017/s0007485321000316 (2021).
Joern, A. Variation in grasshopper (Acrididae) densities in response to fire frequency and bison grazing in tallgrass prairie. Environ. Entomol. 33(6), 1617–1625. https://doi.org/10.1603/0046-225x-33.6.1617 (2004).
Li, J. L. et al. Survey of grasshopper community structure and diversity in Hongsongwa Grassland, Bashang Region, Hebei Province. J. Plant Protect. 48(01), 195–201. https://doi.org/10.13802/j.cnki.zwbhxb.2021.2021826 (2021).
Wang, Y. H., Zhao, M. F. & Kang, M. Y. Plant community distribution patterns in Inner Mongolian grasslands: Explained by dominant environmental factors. J. Beijing Normal Univ. (Nat. Sci.) 52(01), 83–90. https://doi.org/10.16360/j.cnki.jbnuns.2016.01.017 (2016).
Sollai, G. & Solari, P. An overview of “insect biodiversity”. Diversity-Basel 14(2), 134. https://doi.org/10.3390/d14020134 (2022).
Fu, Z. et al. Community differentiation and ecological influencing factors along environmental gradients: Evidence from 1200 km belt transect across Inner Mongolia grassland, China. Sustainability 14(1), 361. https://doi.org/10.3390/su14010361 (2022).
Ma, W. H. & Fang, J. Y. The relationship between species richness and productivity in typical grasslands of northern China. Biodiversity Sci. 01, 21–28 (2006).
Cai, W. Z. & Li, H. A guide to insects in China.
Zhang, W. W. & Li, Y. S. A guide to the ecology of insects in China.
Zhang, L. A guide to 100 species of grasshoppers in China.
Causey, B. D. Parametric estimation of the number of classes in a population. J. Appl. Stat. 29(6), 925–934. https://doi.org/10.1080/02664760220136221 (2002).
Bacaro, G., Gioria, M. & Ricotta, C. Testing for differences in beta diversity from plot-to-plot dissimilarities. Ecol. Res. 27(2), 285–292. https://doi.org/10.1007/s11284-011-0899-z (2012).
Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 14(6), 927–930. https://doi.org/10.1111/j.1654-1103.2003.tb02228.x (2003).
Li, F. et al. Taxonomic, functional, and phylogenetic beta diversity in the Inner Mongolia grassland. Global Ecol. Conserv. 28, e01634. https://doi.org/10.1016/j.gecco.2021.e01634 (2021).
Kassambara, A. ggpubr:‘ggplot2’based publication ready plots. 2, (2018).
Kembel, S. W. et al. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 26(11), 1463–1464. https://doi.org/10.1093/bioinformatics/btq166 (2010).
Ginestet, C. ggplot2: Elegant graphics for data analysis. J. Royal Statist. Soc. Ser. a-Statist. Soc. 174, 245–245. https://doi.org/10.1111/j.1467-985X.2010.00676_9.x (2011).
Chen, H. & Boutros, P. C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. Bmc Bioinform. 12, 1–7. https://doi.org/10.1186/1471-2105-12-35 (2011).
Chang, F., He, S. & Dang, C. Assisted selection of biomarkers by linear discriminant analysis effect size (LEfSe) in microbiome data. Jove-J. Visualiz. Exp. (183). https://doi.org/10.3791/61715 (2022).
Zhang, C. et al. Structural modulation of gut microbiota in life-long calorie-restricted mice. Nat. Commun. 4, 2163. https://doi.org/10.1038/ncomms3163 (2013).
Funding
This project is supported by the Study on the Faunal Composition and Taxonomy of Aradidae in the Hengduan Mountains Region (No. 31760630).
Author information
Authors and Affiliations
Contributions
All authors took part in the collection and identification of pests. B.X.S. provided data and reviewed the manuscript. L.Y.R. and J.L.P. analyzed the data. L.Y.R. wrote the manuscript. J.L.P. reviewed the discussion of the results, and did language revision. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, Y., Ji, L. & Bai, X. Survey and diversity assessment of key insect communities in different grassland types in Inner Mongolia. Sci Rep 15, 22307 (2025). https://doi.org/10.1038/s41598-025-07830-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-07830-9
