
Abstract
Walking is an accessible and beneficial form of physical activity for older adults. However, when performed under perturbed conditions, it can place greater demands on both neuromuscular control and metabolic efficiency, highlighting the need to better understand the underlying adaptations. We compared stable and perturbed walking on a treadmill at three different speeds in 16 healthy older adults. Muscle synergies were extracted from electromyographic signals of eight lower limb muscles, oxygen uptake (VO2), minute ventilation, heart rate, and rate of perceived exertion (RPE) were also collected. Perturbed walking at 3 km/h increased oxygen uptake (p = 0.003; d = -0.52) and energetic cost (EC; p = 0.003; d = -1.12) compared to the stable condition. VO2 (p < 0.001; \(\:{\eta\:}_{p}^{2}\) = 0.92), EC (p = 0.02; \(\:{\eta\:}_{p}^{2}\) = 0.25) and RPE (χ2 = 4.45; p = 0.04) were higher at 3 km/h than at higher speeds. Four muscle synergies were required to accurately model the neuromuscular control during both conditions. Full-width at half maximum revealed that the transition from stable to perturbed walking led to statistically significant modifications in gait synergy composition (p = 0.01; \(\:{\eta\:}_{p}^{2}\)= 0.04). Weight sparsity revealed no significant differences between conditions. These results suggest that the transition from an accurate to a more robust locomotor control for managing continuous perturbations may come at the expense of increased metabolic demand.
Similar content being viewed by others

Introduction
Regular physical activity is one of the keys to successful aging. As recommended by the World Health Organization, older adults (> 65 years) should engage in at least 150–300 min of moderate-intensity aerobic physical activity, 75–150 min of vigorous-intensity aerobic physical activity, or an equivalent combination of moderate- and vigorous-intensity activity per week to reap substantial health benefits1. Among the various physical and sports activities that can be proposed to achieve this goal, walking is a low-cost option that most older people2 can do with health benefits at various levels3,4,5,6.
Although walking in places such as shopping malls is commonly considered safe and comfortable7,8 moving over uneven terrain is a daily challenge for older adults because of the need for the Central Nervous System (CNS) to flexibly modify its control strategies to cope with varying surface conditions. These demands have an impact not only from a cognitive perspective but also from a metabolic perspective. The Energy Cost (EC) of locomotion (amount of metabolic energy required to move a given distance, typically expressed as J/kg/m) increases on irregular surfaces (e.g., sand, snow, and trails) compared to smooth surfaces (e.g., tarmac, flooring, and track)9,10,11,12,13,14,15 but the underlying biomechanical and neuromuscular mechanisms are not fully understood. Among the various parameters that may affect the EC between uneven and smooth terrains, one must consider step width and step length, as both tend to change with age16. Irregular terrains may also perturb gait step-by-step and cause greater variability. Such surfaces might also demand more mechanical work from the legs, regardless of their effects on step parameters. Kuo17 previously hypothesized that the walking economy improves by pushing with the rear leg just before the collision with the front leg. This push redirects the body’s center of mass and, if timed correctly, can reduce the amount of negative work done during the collision. Irregular terrains may alter the timing of these events so that a collision occurring earlier or later than the push would lead to increased negative mechanical work. This would then require muscles to compensate and perform more positive work elsewhere, as stable walking requires overall zero net work. It is difficult to predict how work will be distributed among the lower limb joints, but altered timing would likely require more work overall and, thus, a higher metabolic energy expenditure.
Another potential factor contributing to increased energy expenditure could be the co-activation of muscles that may occur in unpredictable or irregular terrain conditions or when there is a potential risk of falling. Indeed, on uneven surfaces, healthy adults adopt several gait modifications, such as a lowered center of mass, greater co-contractions during stance, and increased toe clearance during swing phase18. Supporting this, a study by Voloshina and colleagues13 found that walking on uneven terrain with a surface variability of only 2.5 cm increased net metabolic cost by 28%, similar to the effort required for walking up a 2% incline19. Despite only modest changes in step strategy being demonstrated, with a 4% decrease in step length and no significant change in step width, variability in both step length and width increased by 22% and 36%, respectively. This increased variability, along with a more crouched posture and greater muscle activity (especially at the hip), likely contributed to the metabolic cost. Biomechanically, the hip performed 62% more positive work, and the knee 26% more negative work. Although they observed an increase in vastus lateralis and vastus medialis muscle activities during the stance phase, it is not possible to estimate the cost of co-activations. However, these co-activations likely contributed to the increased metabolic cost on uneven terrains. In another study by Hawkins et al.18, participants increased limb work by up to 20% on uneven terrain20 with older adults relying more on proximal muscles at the hip and knee, which are more susceptible to age-related decline. Interestingly, this increase in proximal workload was partially offset by reduced ankle kinetics.
From this point of view, new methodological approaches are needed to evaluate motor control strategies during gait over different terrains in a quantitative and non-invasive way. One promising method involves analyzing muscle synergies21—low-dimensional control modules through which the CNS coordinates multiple muscles using a limited set of time-dependent neural commands22,23. These synergies generate complex electromyographic activity via descending or afferent pathways24,25 facilitating motor control (e.g., locomotor control) with some degree of supraspinal regulation26. These synergies are extracted from surface EMG recordings using algorithms such as Non-Negative Matrix Factorization (NNMF)27. Research shows that human locomotion28,29 in both healthy and pathological conditions30,31 can be described using just 4 to 5 synergies32,33 depending on factors like the muscle set, preprocessing, and algorithm used. Remarkably, these synergies tend to be preserved even under continuous external perturbations, suggesting that the CNS compensates by expanding and modulating basic motor commands to maintain stable movement serves as a compensatory strategy to deal with the perturbations34,35.
Thus, muscle synergies enable an objective and non-invasive assessment of the CNS’s motor control strategies during different tasks. In this context, muscle synergies and the analysis of EC are promising frameworks for evaluating shifts in neuromuscular and metabolic strategies in response to different conditions, such as walking on even and uneven terrains. Moreover, integrating metabolic data with muscle synergy analysis may offer valuable insights into the central correlates of muscle synergies, further elucidating the neural mechanisms underlying motor control adaptations.
This study aims to address a knowledge gap regarding the metabolic and neuromuscular adaptations required for older adults to walk successfully during unpredictable medio-lateral variations, a common challenge encountered in daily life (such as normal walking on uneven terrain, as typically found in extra-urban, rural, or mountainous areas). Accordingly with previous literature, we hypothesized that, compared to flat walking, walking on a perturbed surface at the same speed would require adaptations in motor control strategies—shifting from accurate (i.e., mature and functionally fine-tuned) to more robust ones (i.e., better suited to handle unexpected errors)35—along with an increase in energy cost (EC)13 and no change in perceived effort36. Although the advantages of physical activity for aging populations are well documented, there is a paucity of research examining the specific EC and motor control strategies employed when walking over irregular surfaces or during unpredictable stimuli. By examining the interplay between motor control strategies and energy expenditure, this study provides novel insights into the impact of unpredictable perturbations on gait performance in healthy older adults. Notably, while numerous studies have independently investigated the effects of various terrain conditions on the metabolic and neuromuscular systems, this is the first study to simultaneously explore both metabolic and neuromuscular control changes resulting from medio-lateral perturbations. These findings may potentially inform strategies for improving mobility, reducing fall risk, and promoting overall health in older adults, thereby enhancing the effectiveness of physical activity interventions targeted at this population.
Results
The metabolic and neuromuscular performance of older adults walking during Flat Condition (FC) and Unpredictable Roll Variations (URV) conditions were compared using metabolic parameters including Energy Cost (EC, metabolic energy required per unit distance), Oxygen Consumption (VO₂, volume of oxygen consumed), Heart Rate (HR), and Minute Ventilation (VE, volume of air breathed per minute), as well as neuromuscular metrics such as muscle synergy composition (optimal number of muscle synergies and description of the activation coefficients and weight vectors), Variance Accounted For (VAF, goodness of muscle synergy model in reconstructing the original EMG data), Full-Width at Half Maximum (FWHM, number of activation coefficient time-instants exceeding half of the signal’s maximum value), Weight Sparsity (WS, number of muscles mainly enrolled within each muscle synergies), and Cosine Similarity of weight vectors (CS, degree of correlation between couple of weight vectors extracted during walking conditions). Moreover, the Rated Perceived Exertion (RPE, subjective measure of effort) data were analyzed during both walking conditions.
Metabolic analysis
The results of the metabolic analysis are detailed in Table 1, with the indication of the statistically significant differences between walking conditions at the same speed (indicated through asterisks). To notice, given that at moderate exercise intensities a steady state is generally reached within the first three minutes, to identify potential duration-dependent metabolic alterations that could arise due to prolonged exposure, we conducted an additional sub-analysis of the physiological variables at the midpoint of each stage (i.e., EC3min and EC6min).
The Shapiro-Wilk test showed that all the metabolic data were normally distributed.
2-way ANOVA revealed no statistically significant difference in EC3min values between flat and unpredictable roll variations conditions based on Speed (\(\:F\)(2, 26) = 1.74, p = 0.20; \(\:{\eta\:}_{p}^{2}\) = 0.12). Significant differences were detected based on Condition (\(\:F\)(1, 13) = 9.08; p = 0.01; \(\:{\eta\:}_{p}^{2}\) = 0.41) and Speed × Condition (\(\:F\)(2, 26) = 11.1; p < 0.001; \(\:{\eta\:}_{p}^{2}\) = 0.46). Post-hoc analysis revealed a significant interaction effect between flat and perturbed conditions at 3 km/h (pbonf = 0.02; Cohen’s d = -1.11). Considering EC6min, statistically significant differences were detected between flat and perturbed conditions based on Speed (\(\:F\)(2, 26) = 4.33, p = 0.02; \(\:{\eta\:}_{p}^{2}\) = 0.25). Significant differences were detected based on Condition (\(\:F\)(1, 13) = 14.97; p = 0.002; \(\:{\eta\:}_{p}^{2}\) = 0.54) and Speed × Condition (\(\:F\)(2, 26) =13.49; p < 0.001; \(\:{\eta\:}_{p}^{2}\) = 0.51). Post-hoc analysis revealed a significant interaction effect between flat and perturbed conditions at 3 km/h (pbonf = 0.003; Cohen’s d = -1.12).
2-way ANOVA revealed a statistically significant difference in VO2 values between flat and perturbed conditions based on Speed (\(\:F\)(2, 28) = 158.1, p < 0.001; \(\:{\eta\:}_{p}^{2}\) = 0.92), Condition (\(\:F\)(1, 14) = 11.06; p = 0.005; \(\:{\eta\:}_{p}^{2}\) = 0.44), and Speed × Condition (\(\:F\)(2, 28) = 8.08; p < 0.002; \(\:{\eta\:}_{p}^{2}\) = 0.37). Post-hoc analysis revealed an interaction between flat and perturbed conditions at 3 km/h (pbonf = 0.003; Cohen’s d = -0.52).
2-way ANOVA revealed a statistically significant difference in HR values based on Speed (\(\:F\)(1.393, 28) = 73.63, p < 0.001; \(\:{\eta\:}_{p}^{2}\) = 0.84). No differences were detected for Condition (\(\:F\)(1, 14) = 3.35; p = 0.09; \(\:{\eta\:}_{p}^{2}\) = 0.19) and Speed × Condition (\(\:F\)(2, 28) = 0.95; p = 0.39; \(\:{\eta\:}_{p}^{2}\) = 0.06). The post-hoc analysis did not reveal any interaction effect between the conditions matched for speed.
Finally, no significant differences in VE for Speed (\(\:F\)(2, 30) = 95.71; p < 0.001; \(\:{\eta\:}_{p}^{2}\) = 0.87), Condition (\(\:F\)(1, 15) = 6.65; p = 0.02; \(\:{\eta\:}_{p}^{2}\) = 0.31), and Speed × Condition (\(\:F\)(2, 30) = 0.85; p = 0.44; \(\:{\eta\:}_{p}^{2}\) = 0.05) were detected.
The raincloud plots of the metabolic parameters are represented in Fig. 1S in the Supplementary Materials.
Muscle synergy composition
Figure 1 shows the average weight vectors and activation coefficients of the muscle synergies identified during the two walking conditions (flat and unpredictable roll variations) at different speeds (i.e., 3, 4, and 5 km/h).

Comparison of muscle synergies averaged across the sample population during stable (green) and perturbed (orange) walking conditions. (A) presents the muscle synergies extracted during the first three minutes of each walking condition, while (B) displays those from the final three minutes. Activation coefficients (C) and weight vectors (W) are represented as mean ± standard errors. Muscle abbreviations: VL – vastus lateralis, VM – vastus medialis, TA – tibialis anterior, PL – peroneus longus, SOL – soleus, GM – gastrocnemius medialis, ST – semitendinosus, and BF – biceps femoris.
On average, four muscle synergies were required to accurately model the original EMG signals across both walking conditions and all walking speeds. From a biomechanical standpoint, each muscle synergy can be linked to a distinct motor function, as outlined below:
-
First muscle synergy is crucial for hip stabilization during heel strike and the load acceptance phases, primarily driven by the activation of the VL and VM muscles;
-
Second muscle synergy is responsible for propulsion during the mid and late stance phases, primarily characterized by the activation of the SOL and GM muscles;
-
Third muscle synergy contributes to leg and foot deceleration during the late swing phase, preparing the limb for foot contact. This synergy is mainly characterized by the activation of the ST and BF muscles;
-
Fourth muscle synergy ensures forefoot clearance during the swing phase, primarily driven by the activation of the TA and PL muscles.
The composition of muscle synergies (i.e., activation coefficients and weight vectors) remains largely consistent across both walking conditions and all walking speeds, suggesting a well-organized and adaptable control system that effectively accommodates varying walking conditions.
Muscle synergy analysis
The average muscle synergy-derived values are detailed in Tables 2 , 3, and Table 4 with the indication of the statistically significant differences between walking conditions and speeds (indicated through asterisks). To identify potential duration-dependent alterations in motor control resulting from prolonged exposure, we analyzed muscle synergy-derived parameters across two consecutive 3-minute intervals (i.e., I Half and II Half).
The Shapiro-Wilk test showed that all the muscle synergy data were normally distributed.
3-way ANOVA revealed no statistically significant differences in VAF values based on Speed (\(\:F\)(2, 178) = 1.91, p = 0.15; \(\:{\eta\:}_{p}^{2}\) = 0.02), Condition (\(\:F\)(1, 178) = 0.37, p = 0.54; \(\:{\eta\:}_{p}^{2}\) = 0.002), and Time Window (\(\:F\)(1, 178) = 1.27, p = 0.26; \(\:{\eta\:}_{p}^{2}\) = 0.007). The post-hoc analysis did not reveal any interaction effect between factors.
No statistically significant differences in FWHM values based on Speed (\(\:F\)(2, 178) = 2.36, p = 0.10; \(\:{\eta\:}_{p}^{2}\) = 0.03) and Time Window (\(\:F\)(1, 178) = 0.42, p = 0.52; \(\:{\eta\:}_{p}^{2}\) = 0.002) were detected. However, a statistically significant difference was detected based on Condition (\(\:F\)(1, 178) = 6.80, p = 0.01; \(\:{\eta\:}_{p}^{2}\)= 0.04). The post-hoc analysis did not reveal any interaction effect between factors.
3-way ANOVA revealed no statistically significant differences in WS values based on Condition, Time Window, and Speed.
Finally, 2-way ANOVA revealed no statistically significant differences in CS values based on Time Window and Speed.
The raincloud plots of the muscle synergy-derived parameters are represented in Fig. 2S in the Supplementary Materials.
Exertion analysis
The Rated Perceived Exertion (RPE) data analyzed using the Shapiro-Wilk test revealed a deviation from normal distribution. Consequently, the Friedman test was conducted to assess the differences between the two conditions. A significant difference was found (see Table 5) at 3 km/h (χ2(1, N = 16) = 4.45; p = 0.04) and at 5 km/h (χ2(1, N = 16) = 6.23; p = 0.01), while the statistical significance was not reached for 4 km/h (χ2(1, N = 16) = 1.92; p = 0.17).
Discussions
The present study examined the metabolic and neuromuscular control changes during a walk at three different speeds (3, 4, and 5 km/h) in two conditions: stable/flat and unpredictable/perturbed in healthy older adults. The aim was to analyze modular locomotor control through muscle synergy, cardio-pulmonary, and perceived exertion measures. It was hypothesized that perturbed walking at the same speed would result in increased neuromuscular and metabolic demands, reflected by a shift from accurate (i.e., mature, functionally fine-tuned) to more robust (i.e., able to cope with unexpected errors) motor control, higher energy cost (EC), and unchanged perceived effort.
Our results showed that participants had higher VO2 values and, consequently, higher EC6min in the perturbed condition at 3 km/h compared to the flat condition (VO2 = + 18%; EC6min = + 28%). In addition, VO2 and EC values were also higher at 3 km/h than at higher speeds. This is further supported by the perceived exertion score (Borg Scale), where subjects reported greater perceived exertion in the perturbed compared to the flat condition at 3 km/h (+ 58%). These results indicate that the 3 km/h speed was uncomfortable because it was too slow compared to the subjects’ usual walking speed, and unexpected perturbations exacerbated this difference.
The difference in RPE between perturbed and flat conditions was significant at 5 km/h (+ 16%) but not at 4 km/h. Since there were no differences between conditions in terms of metabolic and ventilatory variables (VO2, EC, and VE), the results suggest that at 4 km/h, participants were in a “comfort zone” where the speed was comfortable and manageable for them, allowing them to efficiently make the necessary postural/muscular adjustments required by the perturbations. This is elegantly supported by Saibene and Minetti37 who showed that 1.1 m/s (4 km/h) is considered the optimal walking speed. The difference in perception at the higher speed could be caused by processes acting at a level different from the metabolic one.
Continuous exposure to unexpected perturbations is capable of challenging the system as it requires the use of alternative motor control strategies at the neuromuscular level. In fact, continuously variable and unexpected perturbations would exclude any short-term predictive behaviour of the system35. A previous study38 evaluated the EC on uneven surfaces during running, reporting an EC increase of + 5%. This increase was accompanied by changes in the variability of step width (+ 27%), length (+ 26%), and height (+ 125%) on uneven terrain. Additionally, step period variability on uneven terrain increased significantly by 30%38. Notable increases in mean muscle activity were observed in the vastus medialis (+ 7%), rectus femoris (+ 20%), and medial hamstring (+ 19%) muscles. However, no significant differences in mean muscle activity were observed considering the vastus lateralis muscle or any other lower-limb muscle between the conditions. Furthermore, the authors demonstrated a notable degree of EMG variability between conditions for the soleus and gastrocnemius (both + 14%), the vastus medialis (+ 15%), the rectus femoris (+ 35%), and the medial hamstring (+ 26%) muscles. An alternative approach to the previous study, which used disturbances created by the treadmill surface by placing blocks of different heights on the belt, was to examine the effects of unpredictable medio-lateral disturbances on a cohort of well-trained runners from a metabolic and electromyographic perspective at different speeds36. The authors showed that the cardio-metabolic parameters remained unaltered by the perturbations. Additionally, the amplitude and width of EMG activation peaks demonstrated no variation between conditions. However, the variability of EMG exhibited a notable impact from the conditions, exhibiting discrepancies in the coefficient of variation in peak amplitude and peak width, which were elevated in the perturbed compared to the flat condition. It can be reasonably deduced that the absence of notable cardio-metabolic variations between the conditions can be attributed to the limited contact times, which did not permit the subject to discern the alterations introduced by the treadmill and, consequently, to regulate an optimal metabolic response. This is corroborated by Voloshina’s13,38 studies on the impact of surface conditions on running and walking. Their findings indicate that the influence of surface irregularities on running performance is approximately 5%, whereas it is 27% for walking. However, the observed differences in the coefficient of variation in peak amplitude and peak width demonstrate how variations in medio-lateral inclination are detected by the neuromuscular system and addressed with different control strategies by the CNS36. Indeed, during walking, in the presence of perturbations in the medio-lateral direction, participants do not change their speed as a coping mechanism for stability, but rather they change the spatio-temporal parameters of their gait. Indeed, as demonstrated by Hak et al.39,40 subjects did not change their walking speed in response to the balance perturbations, but their steps became shorter, faster, and wider as the intensity of the perturbations increased. With regard to the EC assessed under both even and uneven terrain conditions during treadmill walking, an increase of + 27–28% was observed in the latter13,38. These changes were accompanied by alterations in gait biomechanics, including a 4% reduction in step length, a 22% increase in step length variability, and a 36% increase in step width variability13. In support of this, a study by Grimmitt and colleagues41 found that a 1% increase in stride length variability was associated with a 0.7% increase in EC. Reducing the energy expenditure associated with unstable locomotion may not be the primary objective during movement. Sometimes, compromises may be necessary to prioritize other factors, such as safety or perceived comfort35.
From a motor control perspective, our results showed that four muscle synergies were required to accurately model the neuromuscular control during both stable and perturbed walking. These findings align with previous literature on gait synergies, which suggests that four to five muscle synergies (depending on the type and number of muscles acquired) are necessary to effectively describe human locomotion28,32,42. Results revealed that perturbed walking does not affect the motor control capacity (i.e., number of muscle synergies) of elderly participants, substantiating the hypothesis behind the muscle synergy theory, which states that a consistent set of muscle synergies can be used to describe slightly different motor tasks, such as flat and perturbed walking. In other words, the CNS seems to be able to modulate the existing set of muscle synergies to adapt to varying locomotion conditions. This conclusion is further supported by prior studies demonstrating a consistent number of muscle synergies across different walking conditions32,35,43,44. A reduction in the number of muscle synergies between stable and perturbed walking was observed only in studies that applied more intense walking perturbations capable of inducing falls45,46. The recruitment of a common set of muscle synergies for executing different motor tasks is also reported as a strategy for the central nervous system to respond more rapidly to force perturbations rather than generating a new set of muscle synergies47. These findings were further reinforced by a comprehensive analysis of the muscle synergies using the Full-Width at Half Maximum (FWHM), Weight Sparsity (WS), and Cosine Similarity (CS) parameters. Results revealed that, while the total number of gait synergies remained unchanged, the transition from stable to perturbed walking led to statistically significant modifications in gait synergy modulation. Specifically, the neural activation patterns (i.e., activation coefficients) were notably altered, as indicated by a significant increase in FWHM (p = 0.01), reflecting a broader and more prolonged activation of muscle synergies throughout the gait cycle. Even if participants’ stability state was not directly measured in our experimental protocol, we can assume that these unpredictable perturbations reduced participants’ stability compared to stable walking. Thus, it can be speculated that the temporal rearrangement (i.e., widening) of the neural activation patterns can represent an enhancement in the motor control’s resilience to cope with external disturbances. In other words, these findings may suggest a transition from an accurate to a more robust locomotor control to deal with continuously variable perturbations. These findings are supported by the study by Brüll et al.33 which investigated the impact of gait perturbations on the spatiotemporal modulation of muscle synergies. Their findings indicated that the four basic gait synergies were preserved across all applied perturbations (including medio-lateral and antero-posterior, both unpredictable and predictable treadmill-induced perturbations). However, the temporal recruitment patterns within each synergy (activation coefficients) were strongly modified. The widening of activation coefficients during challenging walking tasks appears to be a common adaptive mechanism, as it has been observed in both healthy individuals34 and patients suffering from multiple sclerosis48,49. Furthermore, the accurate-to-robust transition appears to be influenced by walking speed. Specifically, the increase in FWHM (see Table 2) from stable to perturbed walking was more pronounced at 3 km/h (+ 15%) compared to 5 km/h (+ 4%). This may be attributed to the shorter stance phase at higher speeds, which limits the time available for sensory feedback processing and motor control strategy adaptation. As a result, participants may have been less able to discern and compensate for perturbations introduced by the treadmill at faster speeds13.
From a spinal-level perspective, the analysis of the WS (i.e., the number of muscles actively enrolled within each muscle synergy, see Table 3) and CS (i.e., the degree of correlation between couple of weight vectors extracted during walking conditions, see Table 4) parameters revealed no significant differences between stable and perturbed walking, suggesting that unpredictable perturbations did not substantially alter muscle recruitment patterns at the spinal level. This finding further supports the idea that adjustments to motor control strategies in response to external disturbances mainly occur at the supraspinal level, with cortical and subcortical structures likely playing a dominant role in modulating locomotor control under variable conditions.
Previous studies have shown that walking on uneven terrain increases kinematic variability and perceived instability in older adults, especially those with lower mobility function, with greater step-to-step variability and reduced walking speed compared to younger individuals50,51. Moreover, an increase in task difficulty is accompanied by a corresponding rise in cortical activity. This augmented cerebral activity may encompass augmented activation within a specific region and/or the recruitment of supplementary brain regions. To this end, the capacity to control complex walking tasks may be constrained by limited cognitive resources, particularly within the prefrontal cortex, affecting the quality of movement52. The prefrontal cortex, which is primarily responsible for cognitive processes, has been observed to undergo age-related declines, which may subsequently result in further impairments to motor control during complex walking tasks53. Therefore, the observed neuromuscular and energetic adaptations may represent a broader age-related strategy with the aim of maintaining more conservative or compensatory motor control (assessed through FWHM) at the expense of energy expenditure.
Gait activity is determined by multiple factors, both latent and contextual, which act synergistically, influencing both metabolic efficiency and neuromuscular control. The present work is positioned in this perspective. To the best of the authors’ knowledge, this is the first study to simultaneously evaluate both metabolic and motor control adaptations through a multi-modal approach in response to unpredictable medio-lateral perturbations during walking. The modifications in motor control strategies, as identified through the muscle synergy analysis, align with the metabolic findings, revealing wider neural activation patterns and increased EC during perturbed compared to flat walking, respectively. These findings suggest that coping with continuous perturbations prompts a shift toward more robust locomotor control, which may lead to higher metabolic demand. This association highlights the potential trade-off between stability and efficiency, reinforcing the idea that neuromuscular adaptations aimed at enhancing resilience against perturbations may require greater energy expenditure.
Limitations and future perspectives
In this study, we must consider the sample size as a limitation, as it should be considered that if the results did not reach statistical significance, it may have been due to the small sample size. The results presented here cannot be considered conclusive, as they should be further investigated on a larger-scale sample that includes, potentially, individuals with pathologies as well as both young and adult individuals. Moreover, we investigated eight muscles on a single limb. This setup is considered the minimum required to perform assessments using the muscle synergy technique. A larger number of analyzed muscles (e.g., spinal erectors) could potentially reveal additional modules with different behaviours during perturbed condition54. However, since no one has ever examined the mechanical–bioenergetic interaction during walking under unpredictable medio-lateral perturbations, one of the future directions should be to explore these aspects. In the future, the combination of biomechanical parameters, spatio-temporal gait parameters, metabolic data, and perceived exertion should be further investigated to better define the associations with the study of muscle synergies.
Materials and methods
Participants
A convenience sample of 16 healthy older adults (8 males and 8 females; age = 69 ± 5 years; weight = 68 ± 10 kg; height = 1.67 ± 0.08 m; BMI = 24.3 ± 2.7 kg/m2) took part in this study. Inclusion criteria were: (i) an age of 65–80 years, (ii) home dwelling, and (iii) the ability to walk independently on a treadmill for at least 20 min35. Exclusion criteria were any previous history of neuromuscular disorders or injury in the last six months. All the participants were informed about the testing procedure. They provided written informed consent before participating in this study, which was approved by the local Ethical Advisory Committee (University of Verona —approval no: 11.R1/2023) and performed in accordance with the Helsinki Declaration.
Experimental design
The study comprised two visits interspersed by 1 week. Visit 1 consisted of familiarization and anthropometric measures (approximately 1 h), while Visit 2 was the testing session (approximately 2 h). During Visit 1, the participants were familiarized with the Rated Perceived Exertion scale (RPE, Scale: 0-100) and the unstable condition for each speed used during the test. At the second visit (Visit 2), participants were tested on a treadmill using two modalities (stable and unstable, presented in random order) and at three different speeds (3, 4, and 5 km/h)37, with the slope fixed at 1%55. During this session, metabolic data and EMG signals from 8 different muscles of the dominant lower limb were collected, as well as scores from the RPE scale.
Data recordings
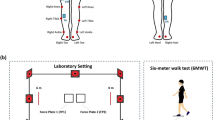
During the second visit, participants completed a 6-minute trial for each walking speed (i.e., 3, 4, and 5 km/h), both in Flat Condition (FC) and Unpredictable Roll Variations (URV) condition, in randomized order. Each trial was separated by a two-minute rest period. During the perturbed condition, a randomized instability routine36 previously used by Skroce and colleagues33 was given based on the following criteria: (i) lateral inclination ranged from − 7° to + 7° and (ii) each position was maintained for 1 to 3 s. To make the results comparable, the same routine was administered to all subjects and at all speeds. All walking tests were conducted on a motorized treadmill (ReaxRun, Reaxing, Milan, Italy; see Fig. 2).
Metabolic data were recorded through a breath-by-breath analyzer (K5, Cosmed, Rome, Italy) that was previously set and calibrated according to the manufacturer’s instructions for each testing session. Lactate samples from the ear lobe were collected and analyzed (Biosen C-line, EKF Diagnostics, Barleben, Germany) before and after every trial.
Gait data were collected through five wireless probes (one used to acquire linear accelerations and four to record EMG signals; sampling frequency: 2048 Hz; gain: 200 V/V; bandwidth: 10–500 Hz; A/D converter resolution: 16 bit; DueLite, OTBioelettronica, Turin, Italy), which allowed the positioning of 8 pairs of bipolar electrodes (CDE-C, Ø=24 mm, OTBioelettronica, Turin, Italy). The electrodes were placed, according to the SENIAM guidelines56 on the following 8 muscles of the right (dominant) lower limb: Bicep Femoris (BF), Gastrocnemius Medialis (GM), SemiTendinosus (ST), Peroneus Longus (PL), Soleus (SOL), Tibialis Anterior (TA), Vastus Lateralis (VL) and Vastus Medialis (VM). The inter-electrode distance was 20 mm. Before the placement of electrodes, the skin was prepared by shaving the hair, applying an abrasive paste (Nuprep, Weaver and Company, Aurora, CO, USA), and, in the end, cleaning with water to reduce the impedance57. To prevent movement artifacts, the electrodes and cables were secured to the subjects’ skin with adhesive tape (Hypafix, 5 × 10 cm; Leukoplast, Hull, UK). Moreover, one wireless probe (2048 Hz; DueLite, OTBioelettronica, Turin, Italy) was placed over the right tibia through adhesive tape to detect gait events through the accelerometer data.
At the beginning and end of each trial, subjects were asked to provide self-assessments regarding RPE.

Sketch of the treadmill used with the medio-lateral oscillations of + 7/-7° highlighted. Figure created from original photographs and processed using Adobe Photoshop (version 21.1.1) for illustrative purposes.
Data analysis - metabolic analysis
The variables recorded included oxygen consumption (VO2), minute Ventilation (VE), Heart Rate (HR), and blood Lactate concentration (La).
The net energy cost (\(\:EC\)) of constant walking speed58 was computed according to the following formula (1):
where \(\:{VO}_{2net}\) (expressed in mlO2·min− 1·kg− 1) represents the net oxygen uptake, and \(\:v\) is the treadmill speed (expressed in m·min− 1). \(\:ELab\) (ml·min− 1·kg[− [1) is regarded as the energy derived from anaerobic lactic energy sources. \(\:ELab\) is conventionally determined by multiplying the net lactate production by the energy equivalent and dividing it by the total exercise duration (i.e. 6 min), expressed as (mM·ml·kg− 1·mM− 1)·min−1. Since during walking the subjects did not produce a lactate buildup as no anaerobic component was utilized, the \(\:ELab\) (mlO2·min− 1·kg−1) was not considered in the \(\:EC\) formula (1). Finally, the energy cost was expressed in J/kg/m using an energy equivalent which takes into account the respiratory exchange ratio (RER)58: VO2·(4.94·RER + 16.04) [J·mlO2 − 1].
Since, at intensities within the moderate domain, the steady state is typically achieved within the first three minutes, an additional sub-analysis of the physiological variables was performed halfway through each step (EC3min and EC6min). This analysis aimed to identify potential duration-dependent metabolic alterations that could arise due to prolonged exposure to both flat and perturbed conditions. Specifically, this approach was intended to investigate whether extended exercise duration would elicit changes in oxygen uptake, lactate accumulation, or other metabolic responses as the body adapted to the varying demands of each condition.
Data analysis - muscle synergy extraction and analysis
Gait Cycles (GCs) were segmented based on the shank-mounted IMU recordings. The identification of Heel-Strike (HS) and Toe-Off (TO) events was based on angular velocity signals in the sagittal plane obtained from the gyroscope59,60. Specifically, an HS event was identified as the time instant corresponding to the minimum angular velocity occurring after the peak angular velocity of the shank, which corresponds to the mid-swing phase. A TO event, instead, was identified as the time instant corresponding to the minimum angular velocity occurring just before the peak angular velocity. To consider steady-state behaviour only, the first and the last 10 gait cycles of each task condition and speed were discarded. The walking task was divided into two time-windows (i.e., the first 3 min and the last 3 min of each trial) to assess potential duration-dependent neuromuscular alterations that could arise due to prolonged exposure to both flat and perturbed conditions61. This approach aligns with the methodology used in the Energy Cost analysis.
Before extracting muscle synergies, EMG data were pre-processed according to the following standard steps. Firstly, an envelope extraction was performed through a 4th -order high-pass Butterworth filter at 35 Hz, a full-wave rectification, and a 4th -order low-pass Butterworth filter at 10 Hz, according to Ghislieri and collegues62,63. Secondly, each GC was time-normalized to 1000 samples64 assigning 600 samples to the stance phase and 400 samples to the swing phase. This was done to avoid biases due to the absolute durations of gait sub-phases and to help assess the temporal contribution of the different muscle synergies to each gait cycle sub-phase65. Then, an amplitude normalization was performed to the global maximum of each acquired muscle66. Lastly, the Non-Negative Matrix Factorization (NNMF) was applied to the time- and amplitude-normalized EMG envelopes following the optimized version proposed by Lee and Seung67.
The NNMF algorithm enables the decomposition of the original data (M) into two low-dimensional components: (i) the activation coefficients (C), which describe the temporal modulation of motor control (i.e., the timing of muscle group activation over task duration) and (ii) the weight vectors (W), which capture the spatial information of motor control (i.e., the muscles synergistically activated). The MATLAB function “nnmf” (release R2023b; MathWorks Inc., Natick, USA) was used to extract the “time-invariant” muscle synergy model from the EMG data, setting the maximum number of iterations equal to 1000, the number of replicates equal to 15, and the function and search tolerance equal to 1e− 662,63. A sparseness constraint was applied to the first initialization of the weight vector to improve algorithm performance68. The NNMF algorithm was applied to the same EMG data changing the muscle synergy number (N) between 1 and 8 (i.e., the minimum and maximum number of recorded muscles in the study) to explore different factorization solutions. To assess the goodness of the muscle synergy model in describing the original EMG data, the R2 similarity between the original and the reconstructed EMG signals was computed for each tested number of synergies. Before muscle synergy analysis, NNMF solutions extracted from different walking conditions and subjects were sorted in the same order according to Pearson’s correlation coefficients calculated between each pair of weight vectors. The activation coefficients were then sorted consequently.
For each volunteer, each walking condition, and both time-windows, muscle synergies were quantitatively analyzed based on: (i) the required number of muscle synergies (Nsyn), (ii) the full width at half maximum (FWHM) of the activation coefficients, and (iii) the sparsity of the weight vectors (WS). Additionally, muscle synergy compositions were qualitatively compared.
Number of muscle synergies (Nsyn). To avoid setting arbitrary cut-off thresholds on model reconstruction quality, the required number of muscle synergies for modelling the original EMG data was determined using the “elbow” criterion - identifying the point of highest curvature on the R2 vs. number of muscle synergies curve. Specifically, the curvature was calculated for each set of three consecutive points along the R2 vs. number of muscle synergies curve, iteratively excluding the smallest number of synergies27. The optimal number of muscle synergies to accurately model the walking task across the entire sample population was then defined as the mode of the synergy numbers calculated for each volunteer. Nsyn ranges from 1 to 8, corresponding to the minimum and maximum number of muscles recorded in the study.
Full Width at Half Maximum (FWHM). The duration of each activation coefficient was computed cycle-by-cycle as the number of time instants exceeding half of the signal’s maximum value (i.e., full width at half maximum), after subtracting the minimum within the same gait cycle. The FWHM values for each participant were then averaged across gait cycles and muscle synergies, obtaining a single representative FWHM value per individual62,63. FWHM is expressed in percentage of the gait cycle duration, ranging from 0% (indicating a null activation duration) to 100% (indicating a long activation duration).
Weight vector Sparsity (WS). WS was defined as the number of significantly active muscles per muscle synergy. To determine this, an adaptive threshold (\(\:t{h}_{k}\)) was computed for each muscle (\(\:k\:\)) as follows (2):
where \(\:{W}_{k}\) is a vector containing the contributions of muscle \(\:k\:\) to each muscle synergy. The overall threshold (\(\:th\:\)) was then determined as the maximum of all individual \(\:t{h}_{k}\) values. A muscle was then considered significantly active in a given synergy if its contribution (i.e., weight vector value) exceeded \(\:th\). WS ranges from 1 to 8, reflecting the minimum and maximum number of recorded muscles in this study.
Cosine Similarity (CS). To quantify the degree of correlation between couple of weight vectors extracted during different walking conditions, the Cosine Similarity (CS) was computed for each muscle synergy69. The CS between two weight vectors was defined as the normalized scalar product between the vectors. CS values range between 0 (indicating a null correlation between two weight vectors) and 1 (indicating a strong correlation between two weight vectors).
Statistical analysis
To assess data distribution normality, the Shapiro-Wilk test was performed. In the case of normally distributed data, a 2-way ANOVA (Speed × Condition) and a 3-way ANOVA (Speed × Condition × Time Window) were performed to test statistically significant differences in metabolic and muscle synergy data, respectively. Otherwise, the Friedman test for non-parametric data was performed. Post-hoc tests were corrected with Bonferroni method for multiple comparisons. The effect size in ANOVAs was reported as \(\:{\eta\:}_{p}^{2}\). Threshold values70 for \(\:{\eta\:}_{p}^{2}\) were set at 0.01, 0.06, and 0.14, corresponding to small, medium, and large effect sizes, respectively. Sphericity was assessed using Mauchly’s test. In cases where the assumption of sphericity was violated (p < 0.05), degrees of freedom were corrected using the Greenhouse-Geisser adjustment to reduce the risk of Type I error. For all the statistical analyses, the significance level (α) was set equal to 0.05. The outliers were manually removed. All the statistical analyses were performed in JASP (Version 0.19).
Data availability
All data are available from the corresponding author upon reasonable request.
Abbreviations
- BF:
-
Bicep femoris
- CNS:
-
Central nervous system
- CS:
-
Cosine similarity
- EC:
-
Energetic cost
- EC3min :
-
Energetic cost assessed from zero to 3 min from the onset
- EC6min :
-
Energetic cost assessed from 3 min to 6 min
- ELab:
-
Energy derived from anaerobic lactic energy sources
- FC:
-
Flat condition
- FWHM:
-
Full width at half maximum
- GC:
-
Gait cycle
- GM:
-
Gastrocnemius medialis
- HR:
-
Heart rate
- HS:
-
Heel-strike
- La:
-
Blood lactate concentration
- NNMF:
-
Non-negative matrix factorization
- PL:
-
Peroneus longus
- RER:
-
Respiratory exchange ratio
- SOL:
-
Soleus
- ST:
-
SemiTendinosus
- TA:
-
Tibialis anterior
- TO:
-
Toe-off
- URV:
-
Unpredictable roll variations
- VAF:
-
Variance accounted for
- VE:
-
Minute ventilation
- VL:
-
Vastus lateralis
- VM:
-
Vastus medialis
- VO2 :
-
Oxygen consumption
- WS:
-
Weight sparsity
References
Organization, W. H. WHO Guidelines on Physical Activity and Sedentary Behaviour (World Health Organization, 2020).
Szanton, S. L. et al. Older adults’ favorite activities are resoundingly active: findings from the NHATS study. Geriatr. Nurs. 36, 131–135. https://doi.org/10.1016/j.gerinurse.2014.12.008 (2015).
Lee, L. L. et al. Walking for hypertension. Cochrane Database Syst. Reviews. https://doi.org/10.1002/14651858.CD008823.pub2 (2021).
King, A. C., Oman, R. F., Brassington, G. S., Bliwise, D. L. & Haskell, W. L. Moderate-intensity exercise and self-rated quality of sleep in older adults. A randomized controlled trial. Jama 277, 32–37. https://doi.org/10.1001/jama.1997.03540250040029 (1997).
Lee, I. M. & Buchner, D. M. The importance of walking to public health. Med. Sci. Sports Exerc. 40, 512–518. https://doi.org/10.1249/MSS.0b013e31817c65d0 (2008).
Brustio, P. R. et al. Indoor mobility, frailty, and disability in Community-Dwelling older adults: A mediation model. Int. J. Environ. Res. Public. Health. 19 https://doi.org/10.3390/ijerph191811386 (2022).
Ory, M. G., Towne, S. D. Jr., Won, J., Forjuoh, S. N. & Lee, C. Social and environmental predictors of walking among older adults. BMC Geriatr. 16, 155. https://doi.org/10.1186/s12877-016-0327-x (2016).
King, D. K. et al. Safe, affordable, convenient: environmental features of malls and other public spaces used by older adults for walking. J. Phys. Act. Health. 13, 289–295. https://doi.org/10.1123/jpah.2015-0118 (2016).
Zamparo, P., Perini, R., Orizio, C., Sacher, M. & Ferretti, G. The energy cost of walking or running on sand. Eur. J. Appl. Physiol. Occup. Physiol. 65, 183–187. https://doi.org/10.1007/bf00705078 (1992).
Pinnington, H. C. & Dawson, B. The energy cost of running on grass compared to soft dry beach sand. J. Sci. Med. Sport. 4, 416–430. https://doi.org/10.1016/s1440-2440(01)80051-7 (2001).
Heinonex, A., Karvones, M. & Ruosteenoja, R. The energy expenditure of walking on snow at various depths. Ergonomics 2, 389–394. https://doi.org/10.1080/00140135908930455 (1959).
Pandolf, K. B., Haisman, M. F. & Goldman, R. F. Metabolic energy expenditure and terrain coefficients for walking on snow. Ergonomics 19, 683–690. https://doi.org/10.1080/00140137608931583 (1976).
Voloshina, A. S., Kuo, A. D., Daley, M. A. & Ferris, D. P. Biomechanics and energetics of walking on uneven terrain. J. Exp. Biol. 216, 3963–3970. https://doi.org/10.1242/jeb.081711 (2013).
Jensen, K., Johansen, L. & Kärkkäinen, O. P. Economy in track runners and orienteers during path and terrain running. J. Sports Sci. 17, 945–950. https://doi.org/10.1080/026404199365335 (1999).
Bettega, S. et al. Energetic and neuromuscular impact of running on even or uneven surfaces in standardized laboratory conditions. J. Electromyogr. Kinesiol. 81, 102987. https://doi.org/10.1016/j.jelekin.2025.102987 (2025).
Murray, M. P., Kory, R. C. & Clarkson, B. H. Walking patterns in healthy old men. J. Gerontol. 24, 169–178. https://doi.org/10.1093/geronj/24.2.169 (1969).
Kuo, A. D. Energetics of actively powered locomotion using the simplest walking model. J. Biomech. Eng. 124, 113–120. https://doi.org/10.1115/1.1427703 (2002).
Hawkins, K. A., Clark, D. J., Balasubramanian, C. K. & Fox, E. J. Walking on uneven terrain in healthy adults and the implications for people after stroke. NeuroRehabilitation 41, 765–774. https://doi.org/10.3233/nre-172154 (2017).
Margaria, R. Positive and negative work performances and their efficiencies in human locomotion. Int. Z. Angew Physiol. 25, 339–351. https://doi.org/10.1007/bf00699624 (1968).
Lim, Y. P., Lin, Y. C. & Pandy, M. G. Lower-limb muscle function in healthy young and older adults across a range of walking speeds. Gait Posture. 94, 124–130. https://doi.org/10.1016/j.gaitpost.2022.03.003 (2022).
Bizzi, E., Mussa-Ivaldi, F. A. & Giszter, S. Computations underlying the execution of movement: a biological perspective. Science 253, 287–291. https://doi.org/10.1126/science.1857964 (1991).
Mussa-Ivaldi, F. A., Giszter, S. F. & Bizzi, E. Linear combinations of primitives in vertebrate motor control. Proc. Natl. Acad. Sci. U S A. 91, 7534–7538. https://doi.org/10.1073/pnas.91.16.7534 (1994).
Bizzi, E., Cheung, V. C., d’Avella, A., Saltiel, P. & Tresch, M. Combining modules for movement. Brain Res. Rev. 57, 125–133. https://doi.org/10.1016/j.brainresrev.2007.08.004 (2008).
Tresch, M. C., Saltiel, P., d’Avella, A. & Bizzi, E. Coordination and localization in spinal motor systems. Brain Res. Brain Res. Rev. 40, 66–79. https://doi.org/10.1016/s0165-0173(02)00189-3 (2002).
Bizzi, E. & Cheung, V. C. The neural origin of muscle synergies. Front. Comput. Neurosci. 7, 51. https://doi.org/10.3389/fncom.2013.00051 (2013).
Roh, J., Cheung, V. C. & Bizzi, E. Modules in the brain stem and spinal cord underlying motor behaviors. J. Neurophysiol. 106, 1363–1378. https://doi.org/10.1152/jn.00842.2010 (2011).
Tresch, M. C., Cheung, V. C. K. & d’Avella, A. Matrix factorization algorithms for the identification of muscle synergies: evaluation on simulated and experimental data sets. J. Neurophysiol. 95, 2199–2212. https://doi.org/10.1152/jn.00222.2005 (2006).
Rimini, D., Agostini, V. & Knaflitz, M. Intra-subject consistency during locomotion: similarity in shared and subject-specific muscle synergies. Front. Hum. Neurosci. 11, 586. https://doi.org/10.3389/fnhum.2017.00586 (2017).
Monaco, V., Ghionzoli, A. & Micera, S. Age-related modifications of muscle synergies and spinal cord activity during locomotion. J. Neurophysiol. 104, 2092–2102. https://doi.org/10.1152/jn.00525.2009 (2010).
Ghislieri, M., Lanotte, M., Knaflitz, M., Rizzi, L. & Agostini, V. Muscle synergies in parkinson’s disease before and after the deep brain stimulation of the bilateral subthalamic nucleus. Sci. Rep. 13, 6997. https://doi.org/10.1038/s41598-023-34151-6 (2023).
Taborri, J. et al. Feasibility of muscle synergy outcomes in clinics, robotics, and sports: a systematic review. Appl. Bionics Biomech. 2018, 3934698 (2018). https://doi.org/10.1155/2018/3934698
Ivanenko, Y. P., Poppele, R. E. & Lacquaniti, F. Five basic muscle activation patterns account for muscle activity during human locomotion. J. Physiol. 556, 267–282. https://doi.org/10.1113/jphysiol.2003.057174 (2004).
Brüll, L. et al. Spatiotemporal modulation of a common set of muscle synergies during unpredictable and predictable gait perturbations in older adults. J. Exp. Biol. 227 https://doi.org/10.1242/jeb.247271 (2024).
Martino, G. et al. Neuromuscular adjustments of gait associated with unstable conditions. J. Neurophysiol. 114, 2867–2882. https://doi.org/10.1152/jn.00029.2015 (2015).
Santuz, A., Ekizos, A., Eckardt, N., Kibele, A. & Arampatzis, A. Challenging human locomotion: stability and modular organisation in unsteady conditions. Sci. Rep. 8, 2740. https://doi.org/10.1038/s41598-018-21018-4 (2018).
Skroce, K. et al. Flat versus simulated mountain trail running: A multidisciplinary comparison in Well-Trained runners. Int. J. Environ. Res. Public. Health. 20 https://doi.org/10.3390/ijerph20065189 (2023).
Saibene, F. & Minetti, A. E. Biomechanical and physiological aspects of legged locomotion in humans. Eur. J. Appl. Physiol. 88, 297–316. https://doi.org/10.1007/s00421-002-0654-9 (2003).
Voloshina, A. S. & Ferris, D. P. Biomechanics and energetics of running on uneven terrain. J. Exp. Biol. 218, 711–719. https://doi.org/10.1242/jeb.106518 (2015).
Hak, L. et al. Speeding up or slowing down? Gait adaptations to preserve gait stability in response to balance perturbations. Gait Posture. 36, 260–264. https://doi.org/10.1016/j.gaitpost.2012.03.005 (2012).
Hak, L. et al. Stepping strategies for regulating gait adaptability and stability. J. Biomech. 46, 905–911. https://doi.org/10.1016/j.jbiomech.2012.12.017 (2013).
Grimmitt, A. B., Whelan, M. E., Martini, D. N. & Hoogkamer, W. Walking with increased step length variability increases the metabolic cost of walking. BioRxiv https://doi.org/10.1101/2024.05.28.596299 (2024).
Steele, K. M., Tresch, M. C. & Perreault, E. J. The number and choice of muscles impact the results of muscle synergy analyses. Front. Comput. Neurosci. 7, 105. https://doi.org/10.3389/fncom.2013.00105 (2013).
Cappellini, G., Ivanenko, Y. P., Poppele, R. E. & Lacquaniti, F. Motor patterns in human walking and running. J. Neurophysiol. 95, 3426–3437. https://doi.org/10.1152/jn.00081.2006 (2006).
Janshen, L., Santuz, A., Ekizos, A. & Arampatzis, A. Modular control during incline and level walking in humans. J. Exp. Biol. 220, 807–813. https://doi.org/10.1242/jeb.148957 (2017).
Sawers, A. & Bhatt, T. Neuromuscular determinants of slip-induced falls and recoveries in older adults. J. Neurophysiol. 120, 1534–1546. https://doi.org/10.1152/jn.00286.2018 (2018).
Sawers, A., Pai, Y. C., Bhatt, T. & Ting, L. H. Neuromuscular responses differ between slip-induced falls and recoveries in older adults. J. Neurophysiol. 117, 509–522. https://doi.org/10.1152/jn.00699.2016 (2017).
Berger, D. J., Gentner, R., Edmunds, T., Pai, D. K. & d’Avella, A. Differences in adaptation rates after virtual surgeries provide direct evidence for modularity. J. Neurosci. 33, 12384–12394. https://doi.org/10.1523/jneurosci.0122-13.2013 (2013).
Janshen, L., Santuz, A., Ekizos, A. & Arampatzis, A. Fuzziness of muscle synergies in patients with multiple sclerosis indicates increased robustness of motor control during walking. Sci. Rep. 10, 7249. https://doi.org/10.1038/s41598-020-63788-w (2020).
Janshen, L., Santuz, A. & Arampatzis, A. Muscle synergies in patients with multiple sclerosis reveal Demand-Specific alterations in the modular organization of locomotion. Front. Hum. Neurosci. 14, 593365. https://doi.org/10.3389/fnhum.2020.593365 (2020).
Shah, V. A. et al. Correlates of gait speed changes during uneven terrain walking in older adults: differential roles of cognitive and sensorimotor function. Exp. Brain Res. 243, 72. https://doi.org/10.1007/s00221-025-07019-6 (2025).
Downey, R. J. et al. Uneven terrain treadmill walking in younger and older adults. PLoS One. 17, e0278646. https://doi.org/10.1371/journal.pone.0278646 (2022).
Clark, D. J. Automaticity of walking: functional significance, mechanisms, measurement and rehabilitation strategies. Front. Hum. Neurosci. 9, 246. https://doi.org/10.3389/fnhum.2015.00246 (2015).
Seidler, R. D. et al. Motor control and aging: links to age-related brain structural, functional, and biochemical effects. Neurosci. Biobehav Rev. 34, 721–733. https://doi.org/10.1016/j.neubiorev.2009.10.005 (2010).
Peyré-Tartaruga, L. A. et al. Mechanical work as a (key) determinant of energy cost in human locomotion: recent findings and future directions. Exp. Physiol. 106, 1897–1908. https://doi.org/10.1113/ep089313 (2021).
Jones, A. M. & Doust, J. H. A 1% treadmill grade most accurately reflects the energetic cost of outdoor running. J. Sports Sci. 14, 321–327. https://doi.org/10.1080/02640419608727717 (1996).
Hermens, H. J., Freriks, B., Disselhorst-Klug, C. & Rau, G. Development of recommendations for SEMG sensors and sensor placement procedures. J. Electromyogr. Kinesiol. 10, 361–374. https://doi.org/10.1016/s1050-6411(00)00027-4 (2000).
Merletti, R., Cerone, G. L. & Tutorial Surface EMG detection, conditioning and pre-processing: best practices. J. Electromyogr. Kinesiol. 54, 102440. https://doi.org/10.1016/j.jelekin.2020.102440 (2020).
Zamparo, P. et al. Mechanical work and efficiency of 5 + 5 m shuttle running. Eur. J. Appl. Physiol. 116, 1911–1919. https://doi.org/10.1007/s00421-016-3443-6 (2016).
Trojaniello, D. et al. Estimation of step-by-step spatio-temporal parameters of normal and impaired gait using shank-mounted magneto-inertial sensors: application to elderly, hemiparetic, parkinsonian and choreic gait. J. Neuroeng. Rehabil. 11, 152. https://doi.org/10.1186/1743-0003-11-152 (2014).
Bertoli, M. et al. Estimation of spatio-temporal parameters of gait from magneto-inertial measurement units: multicenter validation among parkinson, mildly cognitively impaired and healthy older adults. Biomed. Eng. Online. 17, 58. https://doi.org/10.1186/s12938-018-0488-2 (2018).
Aminian, K., Najafi, B., Büla, C., Leyvraz, P. F. & Robert, P. Spatio-temporal parameters of gait measured by an ambulatory system using miniature gyroscopes. J. Biomech. 35, 689–699. https://doi.org/10.1016/S0021-9290(02)00008-8 (2002).
Ghislieri, M., Agostini, V. & Knaflitz, M. Muscle synergies extracted using principal activations: improvement of robustness and interpretability. IEEE Trans. Neural Syst. Rehabil Eng. 28, 453–460. https://doi.org/10.1109/tnsre.2020.2965179 (2020).
Ghislieri, M. et al. Muscle synergy assessment during Single-Leg stance. IEEE Trans. Neural Syst. Rehabil Eng. 28, 2914–2922. https://doi.org/10.1109/tnsre.2020.3030847 (2020).
Clark, D. J., Ting, L. H., Zajac, F. E., Neptune, R. R. & Kautz, S. A. Merging of healthy motor modules predicts reduced locomotor performance and muscle coordination complexity post-stroke. J. Neurophysiol. 103, 844–857. https://doi.org/10.1152/jn.00825.2009 (2010).
Santuz, A. et al. Modular control of human movement during running: an open access data set. Front. Physiol. 9, 1509. https://doi.org/10.3389/fphys.2018.01509 (2018).
Besomi, M. et al. Consensus for experimental design in electromyography (CEDE) project: amplitude normalization matrix. J. Electromyogr. Kinesiol. 53, 102438. https://doi.org/10.1016/j.jelekin.2020.102438 (2020).
Lee, D. D. & Seung, H. S. Learning the parts of objects by non-negative matrix factorization. Nature 401, 788–791. https://doi.org/10.1038/44565 (1999).
Soomro, M. H., Conforto, S., Giunta, G., Ranaldi, S. & De Marchis, C. Comparison of initialization techniques for the accurate extraction of muscle synergies from myoelectric signals via nonnegative matrix factorization. Appl. Bionics Biomech. 2018, 3629347 (2018). https://doi.org/10.1155/2018/3629347
d’Avella, A. & Bizzi, E. Shared and specific muscle synergies in natural motor behaviors. Proc. Natl. Acad. Sci. U S A. 102, 3076–3081. https://doi.org/10.1073/pnas.0500199102 (2005).
Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 20, 37–46. https://doi.org/10.1177/001316446002000 (1960).
Acknowledgements
This study was supported by “Interconnected Nord-Est Innovation Ecosystem (iNEST)” area “4.Digital, Industry, Aerospace” - nel quadro del Piano Nazionale di Ripresa e Resilienza (PNRR) – Ecosistemi dell’Innovazione – Missione 4 Istruzione e Ricerca - Componente 2 dalla ricerca all’impresa – Investmento 1.5, funded by the European Union –NextGenerationEU; identification code ECS00000043, CUP B43C22000450006.
Author information
Authors and Affiliations
Contributions
SD’E, GB, FS and CT conceived the study. LG, LB, FM, SB and SD’E collected the data. CT and SD’E supervised the data collection. LG, LB and MG analyzed the data. MG and SD’E performed the statistical analysis. SD’E and MG wrote the original draft, and DR, CT, GB and FS revised it. All Authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Compliance with ethical standards
The subjects involved provided written informed consent before they participated in this study, which was approved by the local Ethical Advisory Committee (Dep. Neurosciences, Biomedicine and Movement Sciences, University of Verona—approval n°11.R1/2023) and performed in accordance with the Helsinki Declaration.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.


Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
D’Emanuele, S., Ghislieri, M., Ghiotto, L. et al. Muscle synergies and metabolic adaptations during perturbed walking in older adults. Sci Rep 15, 23597 (2025). https://doi.org/10.1038/s41598-025-07835-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-07835-4

{kind=link}
{kind=link}