
Abstract
Bletilla striata, an important traditional Chinese medicine resource, holds high medicinal and ornamental value. However, unscientific habitat selection for its cultivation has led to low yields and poor quality as medicinal materials in China. The optimized MaxEnt model is a powerful tool for analyzing the potential impacts of environmental factors on species distribution and predicting habitat changes under climate change. It offers great significance for the protection and development of B. striata in China. Based on 269 B. striata distribution records in China and 15 major environmental factors, this study simulated the distribution patterns of potentially suitable B. striata habitats under four different climate change scenarios (SSP1.26, SSP2.45, SSP3.70, and SSP5.85) and three time periods (the current period, 2050s, and 2070s). The analysis was conducted using the MaxEnt model which exhibited high predictive accuracy and minimal overfitting. Solar radiation, annual temperature range and mean diurnal range were revealed as the dominant factors affecting B. striata distribution, and their thresholds were ≤ 16,265.39 kJ/m2·d−1, ≤ 39.7 ℃ and ≤ 12.6 ℃, respectively. The results showed that the total potentially suitable B. striata habitats in China were 30.07 × 105 km2 under current climate conditions, mainly distributed in 14 provinces or regions in southern China. Under future climate change conditions, the predicted potentially suitable B. striata habitats will decrease significantly over time, and the centroid of the predicted potentially suitable habitats at all levels will shift northward. The research results can guide future B. striata resource conservation, variety selection, and cultivation.
Similar content being viewed by others
Introduction
B. striata is an important traditional Chinese medicine resource found in forests, roadside grasslands, and rock crevices at 100–3200 m altitudes. B. striata is mainly cultivated in Sichuan, Guizhou, Shaanxi, Anhui, Zhejiang, Hubei, Hunan, Guangdong, and Guangxi1,2. B. striata is noted for its properties of astringency, hemostasis, swelling reduction, and muscle regeneration in ancient Chinese medical texts such as the Shennong Bencao Jing and the Compendium of Materia Medica. B. striata polysaccharides exhibit various pharmacological activities, such as hemostasis, gastric mucosal protection, anti-tumor effects, anti-fibrosis, wound healing promotion, and plasma replacement3,4. The B. striata gum extracted from the tubers of B. striata plants also serves as an auxiliary material in various medicines, including insoluble drug carriers, topical hydrogels, wound dressings, targeted drugs, and biological scaffolds5,6. In cosmetics, B. striata gum is a non-toxic and harmless plant-based additive that can moisturize the skin and delay its aging7. With the recent increase in industrial production of relevant products, the demand for B. striata has been increasing year by year. B. striata is listed in Appendix I of the Convention on Inter national Trade in Endangered Species of Wild Fauna and Flora and is under the second-class protection of the national wild plant conservation list in China (the second batch). Recently, with the increasing demand for B. striata, the market gap between supply and demand has increased yearly and is now very prominent. Therefore, the scale of cultivation of B. striata has expanded rapidly in China8. However, due to the lack of research on the suitability of white and environmental factors, there are significant differences in yield and quality among different planting bases in China9. Based on a review of relevant domestic literature and field investigations, this article conducts a suitability analysis of the white fungus grown in China, providing a reference for understanding the distribution of white fungus resources in China and guiding artificial cultivation.
The current global ecological change is a response to climate change, rising environmental temperatures, and insufficient freshwater, which has led to periodic droughts, high temperatures, or a combination of both pressures. These stresses lead to a decline in plant growth and productivity10. According to reports, in the context of current climate change, the richness of orchids in China is concentrated in the southwestern region, and the lowest richness in the Tibetan Plateau, Xinjiang Region and the western Inner Mongolia. The orchid richness is also low in the plain areas, such as the North China Plain, the Northeast China Plain and the Middle and Lower Yangtze Plain11. Several Crassulacean acid metabolism (CAM) plants have been reported to adapt to semi-arid habitats such as periodic drought and high temperature.Though with an increased ability to tolerate drought, some CAM plants still showed a negative response when facing drought stress12. As a typical CAM plant, it has been found that these environmental pressures limit B. striata growth and development and disturb its different metabolic and physiological activities13. Meanwhile, the sensitivity of B. striata to high temperatures leads to increased pests and diseases, decreased adaptability, and decreased yield14,15. It is imperative to predict and evaluate the suitable areas for its resources.
Species distribution models (SDMs) empirically quantify the environmental niches of species and predict their spatial distributions by correlating species occurrence data with a set of environmental variables16,17. Among the various SDMs, the maximum entropy (MaxEnt) model has emerged as a widely used tool, particularly for modeling species distributions with presence-only data. The inherent complexity of ecological mechanisms motivates the use of this statistical approach for theory development. Given the multitude of factors influencing organisms and their interactions, as well as the extensive array of traits that differentiate species, isolating the most influential factors for theory-building is challenging. MaxEnt addresses this challenge by eliminating the need for such selective decisions. As demonstrated in prior research, MaxEnt not only provides accurate predictions of macroecological patterns but also helps identify the most significant underlying mechanisms18. MaxEnt calculates the probability distribution of maximum entropy based on environmental constraints and remains robust even with small sample sizes19,20. Due to these advantages, it has been extensively applied in recent years to predict species distributions21,22,23. Currently, approximately 33 SDMs are utilized by researchers in China, while the global number of such models likely exceeds 40. Among these, the MaxEnt model has gained widespread popularity since its introduction in 2006. This widespread adoption is primarily attributed to its mathematical algorithm, which excels at inferring unknown distributions from limited known information. Additionally, the model’s ability to generate continuous output data enables fine-grained distinctions in habitat suitability across different regions, enhancing its adaptability and precision in ecological modeling24.
Previous studies have employed SDMs, including MaxEnt, to assess the potential impacts of climate change on species distributions, thereby supporting the conservation of rare and endangered species25,26,27. Researchers have also used these models to evaluate species invasions, test evolutionary hypotheses, and predict disease transmission risks28,29,30,31. However, studies have shown that complex species-environment relationships may accurately fit certain datasets but fail to generalize to others32,33,34. For instance, the MaxEnt model has been found to perform poorly without appropriate adjustments to model settings and optimization procedures35,36. Numerous studies have demonstrated that optimized MaxEnt significantly improves predictive performance, yielding more accurate and ecologically plausible species distribution maps29,37.
Through simulation analysis, potentially suitable B. striata habitats in China can be predicted and categorized into different suitability levels, providing scientific references for its habitat selection and protection38,39,40,41. A clear understanding of geographical distribution is foundational and instrumental for effective B. striata protection and management. Therefore, this research aims to (1) use an optimized MaxEnt to predict the distribution of currently suitable B. striata habitats, (2) determine the distribution of important environmental variables and the limitations of ecological characteristics, and (3) predict future changes in the potentially suitable B. striata habitats. By analyzing the distribution trend under future climate changes, this study provides a reasonable reference and decision-making basis for the long-term management and protection of B. striata.
Materials and methods
B. striata distribution data sources
Detailed distribution information in the form of latitude and longitude data was acquired from the literature and various sources, including the Global Biodiversity Information Facility (GBIF, https://www.gbif.org), Chinese Virtual Herbarium (CVH, http://v5.cvh.org.cn/), Chinese Flora Herbarium (CFH, https://www.cfh.ac.cn/), and National Specimen Information Infrastructure (NSII, http://www.nsii.org.cn/), and specimen coordinates were derived in Google Earth. To prevent spatial autocorrelation of species distribution data and match climate data at 30 arcsecond resolution, only one data point was retained within a diameter of 5 km16. Finally, 269 origin coordinates of B. striata were obtained to construct the maximum entropy model (Fig. 1).

Occurrence of B. Striata in China. Red circles indicate 269 species occurrences of B. Striata in China after removing highly spatially auto-correlated and duplicate records.
Environmental data sources and screening
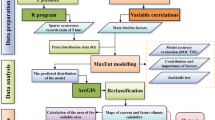
The 24 bioenvironmental climate data in this study are among the most important environmental factors affecting the potential species distribution and are often employed to construct niche models42. The current and future climate data (2021–2040) were collected from WorldClim (http://www.worldclim.org). Among them, the future climate data were based on the BCC-CSM2-MR Climate model for China’s geographical environment in CMIP6. The included sustainable development path was limited to 2 °C (SSP1.26), the moderate development path was limited to 3 °C (SSP2.45), the local development path was limited to 4.1 °C (SSP3.70), and the conventional development path was limited to 5 °C (SSP5.85)43. The slope and slope direction data were extracted from the digital elevation data of the world climate data using ArcGIS. The surface solar radiation was obtained from the National Tibetan Plateau Data Center (http://data.tpdc.ac.cn). When simulating species geographic distribution, the MaxEnt model should be used first to avoid over-fitting model prediction results caused by the multicollinearity of various environmental variables. The importance and contribution rate of the environmental variables should be tested through Jackknife, and factors with a contribution rate ≥ 0.1% should be considered. Factors with a correlation coefficient | r |≥ 0.8 and a low contribution rate are removed44. Then, the distribution data and environment variables were imported into ArcGIS, the sample tool within the extraction tool was used for sampling, and the dot and tape layers were extracted into Excel tables. Then, the correlation between B. striata distribution and climate factor variables was analyzed in SPSS, and environmental variables with a correlation coefficient above 0.95 were retained. Finally, ENMTools was used to conduct Pearson correlation analysis on the environmental variables, and Pearson correlation heat maps were plotted in R (Fig. 2). Variables with a correlation coefficient above 0.8 but a small contribution to the prediction were excluded45. First, the Jackknife method in MaxEnt was used to evaluate the contribution rate of the 24 environmental factors to the distribution of potential B. striata habitats, and Spearman correlation analysis was performed on these factors in SPSS (Fig. 1) Second, efforts were made to avoid over-fitting. Specifically, factors with a contribution rate ≥ 0.1% were selected, and those with a correlation coefficient | r |≥ 0.8 but a low contribution rate were removed (Fig. 2). Finally, 15 factors with high influence on the distribution of potential B. striata habitats were identified and selected for modeling. These include climate factors (bio1, bio2, bio3, bio7, bio8, bio10, bio12, bio15, bio17, and bio18), altitude (ALT, m), solar radiation (SRAD, J/m2·d), vaporpressure(VAPR, Pa), slope(SLOPE, °), and aspect (ASPECT) (Table 1).

Correlation analysis of the main climate factors. See Table 1 for description of environmental variables.
Maximum entropy model establishment, optimization, and evaluation
The “kuenm” package in R 3.6.3 was used to optimize the MaxEnt model parameters. The regularization multiplier (RM) was from 0.5 to 4 (increment 0.5), and the feature classes (FC) adopt the following combinations: L, LQ, H, LQH, LQHP, and LQHPT (L: linear, Q: quadratic, H: hinged, P: Product, T: threshold)46.The “kuenm” package was used to test the above parameter combinations. The model’s fit and complexity were evaluated based on the delta AICc value of the Akaike Information Criterion, and the parameter combination with the smallest delta AICc was selected for model construction47. Furthermore, the area under the Receiver Operating Characteristic (ROC) curve (AUC value) was used to evaluate and verify the accuracy of model results48. An AUC value below 0.6 indicates model failure, a 0.6–0.7 AUC is poor, a 0.7–0.8 AUC is fair, a 0.8–0.9 AUC is good, and an AUC above 0.9 is excellent39. Finally, the B. striata occurrence point data and the selected environmental factors were imported into MaxEnt, and the optimized model parameter combination was repeated 10 times to produce the prediction results of the current potential B. striata distribution. ROC test was performed, and the contribution rates of the environmental factors were obtained using the Jackknife method. The prediction results were imported into ArcGIS 10.8 for reclassification, and the threshold of training sensitivity plus specificity was considered a more robust method for species classification49. The training sensitivity plus specificity threshold of the model prediction results after 10 iterations was 0.19 ± 0.06, and the potential B. striata habitats were classified into four fitness grades: not suitable areas (0–0.2), low suitable areas (0.2–0.4), medium suitable areas (0.4–0.6), and highly suitable areas (0.6–1). Finally, ArcGIS 10.8 was used to visualize the prediction results (Fig. 3).

Performance optimization screening of candidate models.
Potential B. striata distribution changes and centroid shift based on future CMIP6
The ASCII files generated by the MaxEnt model were imported into ArcGIS 10.8 and converted into raster data to calculate the changes in B. striata and distribution areas of different fitness grades under different scenarios in the future and current times50. The SDM toolkit of ArcGIS 10.8 was used to compare the changes in the area and scope of the suitable habitats under the future and current scenarios, allowing for the identification of the expansion and contraction areas and the geographical locations in the future. Taking 0.2 as the threshold, the data was converted into binary data, and the centroid of the suitable habitats and its change direction were calculated using the SDM toolbox51.
Results and analysis
Model optimization and accuracy evaluation
The MaxEnt model based on the 269 distribution sites and the selected environmental variables was used to simulate and predict the potentially suitable B. striata habitats. According to the model optimization results, a p-value (pval_pROC) of 0 indicates statistical significance. The omission rate of 0.0333% is below the 5% threshold and the default value of the MaxEnt model. The W_AICc of 1 is below 2, and its corresponding RM and FC are 0.1 and L. By taking the mean of 10 models as the final result, with an AUC value of 0.8943,52 (Fig. 4).

The ROC test of the model prediction results.
Currently suitable B. striata habitat distribution
The potential B. striata habitats are mainly located in Southwest China and parts of South China. High suitability areas are mainly in Guizhou and most parts of Chongqing, southeast Sichuan, northeast Yunnan, Hubei, Hunan, Guangxi, and other regions. The high suitability area is about 94,200 km2, accounting for 31.33% of the total suitable habitats. The medium suitability area is about 121,300 km2, accounting for 40.34% of the total suitable habitats. The low suitability area is about 85,200 km2, accounting for 28.33% of the total suitable habitats (Fig. 5).

Distribution of B. striata suitable areas.
Major environmental factors affecting B. striata resource distribution
Among the 15 environmental factors used for modeling, the contribution rate of climate factors SRAD, bio7, bio2, and VAPR reached 91.6%. The average contribution rate of SRAD was 45.5%, the average contribution rate of bio7 was 39.4%, the average contribution rate of bio2 was 3.4%, and the average contribution rate of VAPR was 3.3%. Regarding the permutation importance of climate factors among the 15 environmental factors, SRAD, bio7, bio2, bio15, and bio18 accounted for 85.2% of the importance of species distribution. The top four in importance are ranked in order as bio7, SRAD, bio2, and bio18, Among them, bio7 contributed the most at 48.2%. Overall, SRAD, bio7, and bio2 are important factors affecting the suitable growth area of B. Striata. The average contribution rate of temperature variable factors (bio1, bio2, bio3, bio7, bio8, and bio10) (61.7%) was significantly higher than that of precipitation variable factors (bio12, bio15, bio17, and bio18) (19.5%) (Table 2). The thresholds of the dominant environmental factors were calculated. Specifically, the threshold of solar radiation was 9292.4 to 16,265.39 kJ/m−2·d−1, that of the annual temperature range (bio7) was 8.76–39.7℃, and that of the mean diurnal range (bio2) was 3.81–12.6℃ (Fig. 6).

Response curve of main environmental factors. The red line shows the average obtained from 10 model runs, and the blue boundary indicates the standard deviation (SD) of 10 operations.
The area and suitability level changes of B. striata habitats under future climate changes (CMIP6 scenario)
Under the four shared social and economic paths, the variation trends of B. striata habitats and suitability levels were basically the same, while the predicted area of high suitability habitats showed obvious declines under different climate scenarios, with a decline amplitude of over 60%. Under the climate scenarios of SSP2.45 and SSP5.85, the area of high-suitability B. striata habitats in 2050 decreased significantly, reaching 95.22% and 93.52%, respectively. Under the climate scenarios of SSP1.26 and SSP5.85, the area of high-suitability B. striata habitats decreased significantly in 2070, reaching 94.27% and 88.32%, respectively. The predicted area of medium-suitability B. striata habitats also decreased to a certain extent, and the area of medium-suitability B. striata habitats declined the most, reaching 75.93%, in 2070 under the climate scenario of SSP1.26 (Table 3, Fig. 7). These results showed that the growth of SSP5.85 was most restricted under the conventional development path with the growth in high temperatures. In addition to the SSP1.26 condition, the area of suitable B. striata habitats decreased linearly, and the other climatic conditions showed fluctuating declines.

B. Striata suitability areas in different climate situations in 2050 and 2070.
The change range and centroid transfer characteristics of B. striata habitats under future climate changes (CMIP6 scenario)
Under the four different climate scenarios, the suitable B. striata habitats in 2050 and 2070 showed significant decreasing trends compared with the current, and the decreases were mainly in Guizhou, Chongqing, northeast Yunnan, and southeast Sichuan. Over time, the centroid gradually shifted to higher latitudes (Fig. 8).

The trend of center of mass migration in B. Striata suitable area in the future.
Discussion
Model predictive performance
B. striata is a rare plant genus valued for its medicinal and ornamental purposes. Due to the extremely low germination rate of sexually propagated seeds under natural conditions, climate change, and human exploitation, wild B. striata resources are being depleted. A detailed understanding of its distribution is the prerequisite for understanding its rational distribution and utilization in the ecosystem11. This study conducts a detailed analysis of global potential B. striata habitats under current and future climate conditions based on the MaxEnt model, whose good performance ensures accurate prediction of species distribution in relation to local climate conditions. Key environmental factors affecting B. striata distribution include solar radiation, annual temperature range, mean diurnal range, and vapor pressure. Under various climate change scenarios, the model predicts the potential areas of suitable B. striata habitats in different regions during the 2050s and 2070s. The results provide a theoretical basis for formulating practical B. striata cultivation and management measures53. The accuracy of species distribution models (SDMs) is largely influenced by the occurrence record, the study area, and the choice of environmental factors54. This study only used bioclimatic variables as predictors, while species distributions may also be influenced by other factors (biological factors, soil, evolution, dispersal capacity, etc.)55. However, as with most plants worldwide, ecological information related to B. striata does not exist. Despite these inevitable uncertainties in model predictions, SDMs remain useful macro-ecological tools for exploring the dynamic relationship between species distribution and climate conditions56,57,58,59.
Potential distribution area and ecological characteristics of B. striata
Previous studies have shown that B. striata in China is mainly distributed south of the Qinling-Huaihe Line, and its endemic provinces are Yunnan, Hubei, Sichuan, Hunan, Jiangxi, and Zhejiang. The minimum temperature in April and October, the annual temperature range, and the average precipitation in November are the most important meteorological factors affecting the potential distribution of B. striata60. The results of this study are similar to those of previous research in that the most suitable habitats are in Yunnan, Sichuan, Chongqing, Guizhou, Hunan, Hubei, Shaanxi, and other places. However, the predicted distribution of contiguous suitable habitats in this study encompasses regions such as Guizhou, Shaanxi, and others, demonstrating a strong alignment with observed real-world conditions. This consistency underscores the model’s capability to effectively identify and delineate suitable habitats, thereby validating its reliability and accuracy in ecological applications. The most important meteorological factors affecting its potential distribution are SRAD, bio7, and bio2. The Solar radiation has the highest percent contribution at 45.5%, but its permutation importance is relatively low at 14%. The annual temperature range shows a percent contribution of 39.4% and a permutation importance of 48.2%, indicating its significant role in the model. The mean diurnal range has a lower percent contribution and permutation importance, at 3.4% and 8.4%, respectively. Moreover, these findings reveal that vapor pressure contributes 3.3% to the model, ranking as the fourth most influential factor. This highlights its significant impact on the distribution patterns of striped bifidobacteria, underscoring its importance in shaping the species’ habitat suitability. Vapor pressure plays an important role in the relationship between plants and water resources, and increased vapor pressure causes the closure of plant stomata, reduces water loss, and avoids adverse water tension conditions within the xylem61,62. However, little is currently known about the effect of vapor pressure on herb performance, and future controlled quantitative studies may address this issue.
Effects of climate change on future B. striata distribution
It is evident that rising temperatures significantly hinder the growth of B. striata, with the area of highly suitable habitats for this species decreasing across all scenarios by more than 67.8%. Notably, under the RCP4.5 scenario for 2050, the reduction in highly suitable habitats for B. striata is the most pronounced, reaching 95.5%. As a typical orchid species, B. striata exhibits a preference for low-light, warm, and humid conditions. However, in the context of global warming, its sensitivity to temperature and light poses significant challenges to its survival. It is well established that moderate heat stress can induce photoinhibition of Photosystem I (PSI) in heat-sensitive plants, such as wheat63,64. Additionally, heat stress can damage oxygen-evolving complexes (OECs), leading to photoinhibition of Photosystem II (PSII)65. Under natural conditions, B. striata is often affected by the dual pressures of shade and drought. For instance, under complete shade treatment, B. striata downregulates carbohydrate-related gene expression, resulting in significant reductions in net photosynthetic rate, stomatal conductance, and transpiration rate. Similarly, under drought stress, the levels of homocysteine-a biomarker for metabolic disorders in tubers-increase, along with the accumulation of organic acids66. Furthermore, B. striata experiences stress in both fully sun-exposed and fully shaded forest environments, highlighting its vulnerability to extreme light conditions. Consequently, the species’ viable habitat is increasingly restricted to the low-light forest edge zone, further narrowing its potential growth range. These findings underscore the complex interplay between environmental stressors and the physiological responses of B. striata, emphasizing the need for further research to mitigate the impacts of climate change on this species.
B. striata habitat selection and protection strategies
Effective conservation of rare and endangered plants, such as B. striata, is essential. Based on their survival status and population distribution, we believe that the following protection policies and measures should be adopted to enhance protection efforts. (1) Strengthened efforts should be devoted to constructing nature reserves and protecting mountains and forests. Based on the situation of natural and biological resources, efforts should be made to upgrade certain nature reserves. Provincial and county-level nature reserves should be expanded when conditions permit, and new nature reserves should be established to protect rich biodiversity and special biological populations. Special attention should be paid to special and minimal populations. Moreover, the wild B. striata populations have been endangered by human exploitation, and their habitats must be prioritized in the conservation plans. (2) Research on B. striata germplasm resource collection and breeding should be strengthened. Rare and endangered plants are important germplasm genetic resources, and breaking reproductive barriers helps species escape survival crises. Priority should be given to the research on B. striata germplasm resource conservation and breeding excellent varieties, and efficient B. striata breeding technologies and high-quality seeds and seedlings should be explored. (3) Manual interventions should be conducted to expand B. striata planting bases. New planting bases can be established in suitable areas under the forest, and the existing bases can be improved using shade nets and intercropping with tall crops to address adverse factors such as high temperatures. These measures could promote the sustainable development of B. striata resources.
Conclusion
Under future climate change scenarios, the area of suitable B. striata habitats is significantly reduced, and their centroids shift northward. SRAD, bio7, and bio2 are the dominant environmental factors affecting B. striata distribution in China, and SRAD and bio7 are the key factors. The predicted results are generally consistent with the current B. striata distribution in China. However, due to anthropogenic factors and habitat loss, fragmentation, degradation, and severe damage, viable B. striata habitats may be further reduced and fragmented. Conditions for B. striata are expected to be tougher, and measures should be taken to enhance its conservation, utilization, and development. These include strengthening nature reserves and forest protection, promoting B. striata breeding and germplasm resource collection research, expanding B. striata planting bases, and improving B. striata resource management.
Data availability
All data used and analyzed during the current study are available from the corresponding author on reasonable request.
References
Chen, J. et al. Integrative analyses of transcriptome and metabolome shed light on the regulation of secondary metabolites in pseudobulbs of two Bletilla striata (Thunb.) Reichb. F. varieties. J Appl Res Med Aromat Plants. 20, 100280. https://doi.org/10.1016/j.jarmap.2020.100280 (2021).
Zhang, M., Luo, D., Fang, H. L., Zhao, W. & Zheng, Y. Effect of light quality on the growth and main chemical composition of Bletilla striata. J. Plant Physiol. 272, 153690. https://doi.org/10.1016/j.jplph.2022.153690 (2022).
Kong, W. H., Xu, J. B. & Cui, Q. Research progress in chemical components and pharmacological actions of Bletilla striata and extraction technology of its polysaccharide. Inf TCM. 38, 69–78 (2021).
Li, X. L., Zhang, X. G. & Yin, S. P. Polysaccharides of radix bletillae inhibit inflammatory reaction and oxidative stress in rats with ulcerative colitis. Basic Clin Med. 40(2), 224–228 (2020).
Zhang, R. R. et al. Research progress of natural high polymer Bletilla striata polysaccharide in the medical field. Adv Clin Med. 10, 2926–2933 (2020).
Liu, X. W. et al. Experimental study of the Bletilla striata glucomanan composited scaffolds by orthogonal experiment. J Shandong Univ (Health Sci). 52(3), 40–44 (2014).
Zhu, S. M., Chen, H. Y. & Fan, Z. D. Research progress on dosage forms of Bletilla Striata gum. Chin J Mod Appl Pharm. 36, 3130–3135 (2019).
Li, W. P. et al. Rapid propagation of Bletilla striata by synthetic seeds technology. China J Chin Mater Med. 37(22), 3386–3390 (2012).
Zhang, M. et al. Bletilla striata: a review of seedling propagation and cultivation modes. Physiol Mol Biol Plants. 25, 601–609. https://doi.org/10.1007/s12298-019-00644-w (2019).
Swarts, N. D. & Dixon, K. W. Perspectives on orchid conservation in botanic gardens. Trends Plant Sci. 14(11), 590–598 (2009).
Zhang, Z. et al. Distribution and conservation of orchid species richness in China. Biol. Cons. 181, 64–72 (2015).
Taticharoen, T. et al. Response and acclimatization of a CAM orchid, Dendrobium Sonia ‘Earsakul’to drought, heat, and combined drought and heat stress. Sci Hortic. 309, 111661. https://doi.org/10.1016/j.scienta.2022.111661Get (2023).
Han, S. et al. Plastomes of Bletilla (Orchidaceae) and phylogenetic implications. Int. J. Mol. Sci. 23(17), 10151. https://doi.org/10.3390/ijms231710151 (2022).
Hasanuzzaman, M., Nahar, K., Alam, M. M. & Roychowdhury, R. M. Fujita physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 14(5), 9643–9684 (2013).
Bakhtavar, M. A., Afzal, I., Basra, S. M. A., Ahmad, A. H. & Noor, M. A. Physiological strategies to improve the performance of spring maize (Zea mays L.) planted under early and optimum sowing conditions. PLoS ONE 10, e0124441. https://doi.org/10.1371/journal.pone.0124441 (2015).
Aiello-Lammens, M. E., Boria, R. A., Radosavljevic, A., Vilela, B. & Anderson, R. P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 38(5), 541–545 (2015).
Guisan, A., Thuiller, W. & Zimmermann, N. E. Habitat suitability and distribution models: with applications in R (Cambridge University, 2017).
Harte, J. & Newman, E. A. Maximum information entropy: A foundation for ecological theory. Trends Ecol. Evol. 29(7), 384–389. https://doi.org/10.1016/j.tree.2014.04.009 (2014).
Phillips, S. J., Anderson, R. P. & Schapire, R. E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 190(3–4), 231–259 (2006).
Guisan, A., Zimmermann, N. E., Graham, C., Phillips, S. J. & Peterson, A. What matters for predicting the occurrences of trees: Techniques, data, or species’ characteristics?. Ecol. Monogr. 77(4), 615–630 (2007).
Qin, A. et al. Maxent modeling for predicting impacts of climate change on the potential distribution of thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 10, 139–146 (2017).
Alatawi, A. S., Gilbert, F. & Reader, T. Modelling terrestrial reptile species richness, distributions and habitat suitability in Saudi Arabia. J. Arid Environ. 178, 104153. https://doi.org/10.1016/j.jaridenv.2020.104153 (2020).
Holder, A. M., Markarian, A., Doyle, J. M. & Olson, J. R. Predicting geographic distributions of fishes in remote stream networks using maximum entropy modeling and landscape characterizations. Ecol. Model. 433, 109231. https://doi.org/10.1016/j.ecolmodel.2020.109231 (2020).
Liu, X. T., Yuan, Q. & Ni, J. Research advances in modelling plant species distribution in China. Chin J Plant Ecol. 43(4), 273–283. https://doi.org/10.17521/cjpe.2018.0237 (2019).
Guo, Y. et al. Prediction of the potential geographic distribution of the ectomycorrhizal mushroom tricholoma matsutake under multiple climate change scenarios. Sci. Rep. UK 7, 46221. https://doi.org/10.1038/srep46221 (2017).
Guo, Y., Li, X., Zhao, Z. & Nawaz, Z. Predicting the impacts of climate change, soils and vegetation types on the geographic distribution of polyporus umbellatus in China. Sci. Total Environ. 648, 1–11 (2019).
Nneji, L. M. et al. Species distribution modelling predicts habitat suitability and reduction of suitable habitat under future climatic scenario for Sclerophrys perreti: A critically endangered Nigerian endemic toad. Afr. J. Ecol. 58(3), 481–491 (2020).
Li, R. Protecting rare and endangered species under climate change on the Qinghai Plateau. China. Ecol. Evol. 9, 427–436 (2019).
Bowen, A. K. M. & Stevens, M. H. H. Temperature, topography, soil characteristics, and NDVI drive habitat preferences of a shade-tolerant invasive grass. Ecol. Evol. 10(19), 10785–10797 (2020).
Lin, H., Gu, K., Li, W. & Zhao, Y. Integrating coalescent-based species delimitation with ecological niche modeling delimited two species within the Stewartia sinensis complex (Theaceae). J. Syst. Evol. 60(5), 1037–1048 (2021).
Escobar, L. E. et al. Declining prevalence of disease vectors under climate change. Sci. Rep. UK 6, 39150. https://doi.org/10.1038/srep39150 (2016).
Heikkinen, R., Marmion, M. & Luoto, M. Does the interpolation accuracy of species distribution models come at the expense of transferability?. Ecography 35(3), 276–288 (2011).
Wenger, S. & Olden, J. Assessing transferability of ecological models: An underappreciated aspect of statistical validation. Methods Ecol. Evol. 3(2), 260–267 (2012).
Čengić, M. et al. On the importance of predictor choice, modelling technique, and number of pseudoabsences for bioclimatic envelope model performance. Ecol Evol 10, 12307–12317 (2020).
Radosavljevic, A. & Anderson, R. P. Making better Maxent models of species distributions: complexity, overfitting and evaluation. J. Biogeogr. 41(4), 629–643 (2014).
Morales, N. S., Fernández, I. C. & Baca-González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations?A systematic review. Peer J. 5, e3093. https://doi.org/10.7717/peerj.3093 (2017).
Freeman, B., Jiménez-García, D., Barca, B. & Grainger, M. Using remotely sensed and climate data to predict the current and potential future geographic distribution of a bird at multiple scales: The case of Agelastes meleagrides, a western African forest endemic. Avian Res. 10, 1–9 (2019).
Hu, S. P. & He, L. W. Analysis of suitable distribution areas of Fargesia denudata in Baishuijiang national nature reserve using MaxEnt model and ArcGIS. Chin. J. Ecol. 39(6), 2115–2122 (2020).
Yang, Q. J. & Li, R. Predicting the potential suitable habitats of Alsophila spinulosa and their changes. Chin. J. Appl. Ecol. 32(2), 538–548 (2021).
Che, L., Cao, B., Bai, C. K., Wang, J. J. & Zhang, L. L. Predictive distribution and habitat suitability assessment of Notholirion bulbuliferum based on MaxEnt and ArcGIS. Chin. J. Ecol. 33(6), 1623–1628 (2014).
Liu, X. et al. Study on growth suitability for Coptis chinensis based on ecological factors analysis by Maxent and ArcGIS model. China J. Chin. Mater Med. 41, 3186–3193 (2016).
Dan, L. W. & Seifert, S. N. Ecological niche modeling in Maxent: The importance of model complexitly and the performanee of model selection criteria. Ecol. Appl. 21(2), 335–342 (2011).
Walden-Schreiner, C., Leung, Y. F., Kuhn, T., Newburger, T. & Tsai, W. L. Environmental and managerial factors associated with pack stock distribution in high elevation meadows: Case study from Yosemite National Park. J. Environ. Manage. 193, 52–63 (2017).
Zhu, G. P. & Qiao, H. J. Effect of the MaxEnt model,s complexity on the prediction of species potential distributions. Biodivers Sci. 24(10), 1189–1196 (2016).
Dormann, C. F. et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 36(1), 27–46 (2013).
Zhang, H., Zhao, H. X. & Wang, H. Potential geographical distribution of populus euphratica in China under future climate change scenarios based on Maxent model. Acta Ecol. Sin. 40(18), 6552–6563 (2020).
Waren, D. L. & Seilert, S. N. Ecological niche modeling in Maxent: The importanee of model complexity and the perfomance of model selection eriteria. Ecol. Appl. 21(2), 335–342 (2011).
Ahmad, R., Khuroo, A. A., Hamid, M., Charles, B. & Rashid, I. Predicting invasion potential and niche dynamics of Parthenium hysterophorus ( Congress grass ) in India under projected climate change. Biodivers. Conserv. 28, 2319–2344 (2019).
Meyer, A. L. S., Pie, M. R. & Passos, F. C. Assessing the exposure of lion tamarins (Leontopithecus spp) to future climate change. Am J Primatol. 76(6), 551–562 (2014).
Zhao, G. H. et al. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol Indic. 132, 108256. https://doi.org/10.1016/j.ecolind.2021.108256 (2021).
Cong, M. Y., Xu, Y. Y., Tang, L. Y., Yang, W. J. & Jian, M. F. Predicting the dynamic distribution of Sphagnum bogs in China under climate change since the last interglacial period. PLoS ONE 15(4), e0230969. https://doi.org/10.1371/journal.pone.0230969 (2020).
Hosmer, D. W., Lemeshow, S. & Sturdivant, R. X. Applied logistic regression (Wiley, Hoboken, 2013).
Guillera-Arroita, G., Lahoz-Monfort, J. & Elith, J. MaxEnt is not a presence absence method: a comment on Thibaud et al. Methods Ecol. Evol. 5(11), 1192–1197 (2014).
Commander, C. J. C., Barnett, L. A. K., Ward, E. J., Anderson, S. C. & Essington, T. E. The shadow model: How and why small choices in spatially explicit species distribution models affect predictions. Peer J. 10, e12783. https://doi.org/10.7717/peerj.12783 (2022).
Urban, M. C. et al. Improving the forecast for biodiversity under climate change. Science 353(6304), aad8466. https://doi.org/10.1126/science.aad8466 (2016).
Abdelaala, M., Foisa, M., Fenua, G. & Bacchettaa, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. Egypt. Ecol. Inform. 50, 68–75 (2019).
Fois, M., Cuena-Lombraña, A., Fenu, G. & Bacchetta, G. Using species distribution models at local scale to guide the search of poorly known species: Review, methodological issues and future directions. Ecol. Model. 385, 124–132 (2018).
Kaky, E. & Gilbert, F. Assessment of the extinction risks of medicinal plants in Egypt under climate change by integrating species distribution models and IUCN Red List criteria. J. Arid Environ. 170, 103988. https://doi.org/10.1016/j.jaridenv.2019.05.016 (2019).
Vasconcelos, T. S., Rodríguez, M. Á. & Hawkins, B. A. Species distribution modelling as a macroecological tool: A case study using New World amphibians. Ecography 35(6), 539–548 (2012).
Gong, Y., Jing, P. F., Wei, Y. K., Huang, W. C. & Cui, L. J. Potential distribution of Bletilla striata (Orchidaceae) in China and its climate characteristics. Plant Divers Resourc. 36(2), 237–244 (2014).
Appleby, R. F. & Davies, W. J. A possible evaporation site in the guard cell wall and the influence of leaf structure on the humidity response by stomata of woody plants. Oecologia 56, 30–40 (1983).
Grossiord, C. et al. Plant responses to rising vapor pressure deficit. New Phytol. 226(6), 1550–1566 (2020).
Brestic, M., Zivcak, M., Kunderlikova, K. & Allakhverdiev, S. I. High temperature specifically affects the photoprotective responses of chlorophyll b-deficient wheat mutant lines. Photosynth. Res. 130, 251–266. https://doi.org/10.1007/s11120-016-0249-7 (2016).
Chovancek, E. et al. Transient heat waves may affect the photosynthetic capacity of susceptible wheat genotypes due to insufficient photosystem I photoprotection. Plants. 8, 282. https://doi.org/10.3390/plants8080282 (2019).
Wen, X., Gong, H. & Lu, C. Heat stress induces an inhibition of excitation energy transfer from phycobilisomes to photosystem II but not to photosystem I in a cyanobacterium Spirulina platensis. Plant Physiol. Biochem. 43, 389–395. https://doi.org/10.1016/j.plaphy.2005.03.001 (2005).
Yin, R., Chen, L., Deng, P., Cao, X. & Xu, X. Characterization of changes in active ingredients and mining of key metabolites in Bletilla striata under shading and drought stresses. Horticulturae. 10, 163. https://doi.org/10.3390/horticulturae10020163 (2024).
Acknowledgements
This research was supported by the project of the Central Guidance on Local Science and Technology Development Fund of Guizhou Province (Grant No.[2023]007), the National Key Research and Development Program of China (Grant No. 2021YFD1601002), the Science and Technology Program of Guizhou Province (Grant No. ZK[2024] General 554), the Guizhou Agricultural Science Doctoral Fund (Grant No. [2025] 05) and the Modern Agricultural Technology System of Traditional Chinese Medicine in Guizhou Province (GZYCCYJSTX-04). We thank Mingkai Wu for valuable advises on terminology.
Author information
Authors and Affiliations
Contributions
M. L.: Writing—original draft, Writing—review and editing. P. Y.: Methodology, Investigation. L. Y.: Methodology, Project administration. Z. Z.: Data curation, Visualization, Supervision. Y.C.: Writing—review and editing. H. L.: Writing—review and editing. M. W.: Conceptualization, Funding acquisition, Supervision.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.


Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Luo, M., Yang, P., Yang, L. et al. Predicting potentially suitable Bletilla striata habitats in China under future climate change scenarios using the optimized MaxEnt model. Sci Rep 15, 21231 (2025). https://doi.org/10.1038/s41598-025-08372-w
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-08372-w

{kind=link}
{kind=link}