
Abstract
Throughout history, humans have had a complex relationship with lions, both reverencing and fearing them. Interactions between Neanderthals and cave lions (Panthera spelaea) remain poorly documented due to the scarcity of direct evidence. This study examines the selective use of cave lion bones by Neanderthals to determine whether this behaviour was driven by practical, functional, or symbolic factors, through a detailed zooarchaeological analysis. Previous studies highlighted Neanderthals’ skinning and butchering of cave lions, yet new discoveries at Scladina Cave (Belgium) offer deeper insights into this relationship. Dated to the end of the Saalian, the faunal assemblage provides the earliest evidence of bone tools crafted from cave lion remains. A tibia was deliberately processed into multifunctional tools, initially serving as an intermediate tool before being repurposed as retouchers. Proteomic analysis applied on the remains, confirmed the specific identification. These findings reveal that Neanderthals not only competed with but actively utilized cave lions for practical purposes, indicating complex ecological and behavioral interactions. Relationships between pre-humans and large predators, rooted in the Middle Pleistocene, suggest a strategic exploitation of carnivore remains. However, opportunistic procurement cannot be entirely ruled out as a potential factor influencing animal choice in tool production. The intentional transformation of lion bones into functional tools highlights Neanderthals’ cognitive skills, adaptability, and capacity for resource utilization beyond their immediate survival needs.
Similar content being viewed by others

Introduction
Despite their documented coexistence spanning several hundred millennia1,2, direct evidence of interaction between large felids and pre-humans remains exceedingly rare. The earliest evidence of the exploitation of these carnivores predates the Neanderthals. Panthera leo fossilis bones discovered in Layer TD10-1 of Gran Dolina (Spain, MIS 9), bearing numerous cutmarks, suggest the manipulation of a fresh carcass, perhaps even hunted, for food3. Moreover, a fragmentary humerus of Homotherium latidens, found in Layer 13 II-4C (“Spear Horizon”) of Schöningen (Germany, MIS 9), shows typical features of a bone being used as a tool (retoucher)4,5. This bone was obviously fragmented while still fresh, also indicating access to a fresh carcass.
Neanderthal interactions with large carnivores have long been debated, with some studies suggesting that these hominins actively hunted or scavenged cave lions (Panthera spelaea). So far, the earliest indications of Neanderthal exploitation appeared on a third phalanx discovered in Einhornhöhle (MIS 7-6, Germany), likely resulting from the skinning of a cave lion6. Some younger European sites have yielded evidence of butchering activities on cave lions, such as those found in Bolomor Cave7 (MIS 5, Spain) and “Chez-Pinaud”8 (MIS 4, France). Excavations in Siegsdorf (MIS 3, Germany) uncovered cave lion remains, including a rib with marks interpreted as possible impacts from wooden spears, accompanied by numerous butchery marks on various anatomical segments. These findings strongly suggest that Neanderthals engaged in hunting and the exploitation of these large predators6.
At Scladina Cave (Belgium), recent discoveries of bone tools made from cave lion bones raise questions about their function and significance. This study aims to determine whether these modifications reflect purely utilitarian practices or if they suggest a deeper behavioural pattern and contributes to discussions on Neanderthal subsistence strategies and material selection. This research relies both on zooarchaeological and taphonomic evidence as well as analytical techniques, that are ZooMS and LC–MS/MS, which have been used to assert the species identification.
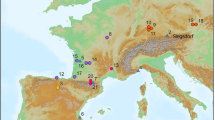
Scladina Cave, located along the right bank of the Meuse Valley between Andenne and Namur, has been under scientific investigation since 1978 (Fig. 1). The site presents an exceptionally well-preserved stratigraphic sequence, comprising at least 120 layers and spanning approximately 400,000 years (Fig. 2)9. Over the past two decades, extensive interdisciplinary studies have refined our understanding of the cave’s sedimentary dynamics, paleoenvironmental history, and chronostratigraphy. This robust framework contextualizes the archaeological findings and taphonomic processes affecting the site’s assemblages10,11,12,13.

Scladina Cave is in the Meuse Valley, in Southern Belgium, close to the limit between Paleozoic limestones (Southeastern part of Belgium) and the low plateaus (Northwestern part). The cave belongs to a network connecting different caves, where Saint-Paul and Sous-Saint-Paul caves are the two other main cavities. The map was generated in Global Mapper v.22.1 using SRTM data from the NASA (https://www.earthdata.nasa.gov/data/instruments/srtm).

Stratigraphic sequence of Scladina Cave exhibiting the archaeological richness of the site. The sedimentary Unit 5 has yielded the main archaeological assemblage containing the bone retouchers made from carnivoran remains (Ursus spelaeus and Panthera spelaea). Above Unit 2A, the system is filled by sediments from two sources: the entrance to the cave on the right and an open doline in the ceiling of the cave on the left (Modified after Bonjean et al., 2014).
Excavations at Scladina have yielded multiple Paleolithic assemblages, including limited Upper Paleolithic evidence associated with the Aurignacian and more substantial Neanderthal occupations linked to Mousterian industries14,15,16. The discovery of the Scladina Child, one of the best-preserved Neanderthal juvenile remains in western Europe, further highlights the site’s significance12,13,17,18.
The main archaeological assemblage has been retrieved within the stratigraphic Unit 5 (also referred as Vb on the terrace), which consists of a diamicton affected by solifluxion12. Uranium/Thorium dates recently obtained on speleothems led to position the thick CC4 speleothem from Sedimentary complex 4 in the Eemian9. The underlying Unit 5 is now placed at the end of the Middle Pleistocene (MIS 6, Fig. 2), consistent with the infrared stimulated luminescence on feldspar (IRSL) ages obtained from the overlying Unit 4B (153 ± 15 ka BP; Balescu in 13) and with the thermoluminescence (TL) date obtained on burnt flint directly from Unit 5 (130 ± 20 ka BP)19.
Archaeological assemblage 5 is characteristic of the Mousterian techno-complex, associated with Middle Paleolithic Neanderthal populations. The toolkit primarily comprises flint artifacts, reflecting a well-structured chaîne opératoire involving local and non-local raw material procurement, core preparation, and systematic flake production20,21,22. Retouched tools, while not abundant, include side scrapers, notches, and denticulates, often showing evidence of extended use and rejuvenation. The diversity of tool types, along with a high frequency of cortical flakes and débitage, indicates on-site tool manufacture and maintenance.
The presence of 29 bone retouchers within the archaeological assemblage 5, particularly one made from a cave bear (Ursus spelaeus) remains (17), points to a complex toolkit that integrated lithic and osseous technologies23. These tools were likely employed in retouching flint edges, as indicated by micro-chipping patterns and embedded lithic residues23,24. Overall, the lithic industry of Unit 5 reflects a flexible and adaptive technological behavior well-aligned with Neanderthal subsistence strategies. Zooarchaeological analyses of Unit 5 indicate that Neanderthals primarily hunted chamois (Rupicapra rupicapra) but also processed a wide range of species, from small mammals to megafauna25,26,27. These observations, combined with the palynological records, suggest that the Neanderthal occupation took place during a cold phase, where the environment was open; probably during a winter given the age at which the chamois have been hunted26.
Several carnivores have been identified within Unit 5, such as the wolf (Canis lupus) and the red fox (Vulpes vulpes), which are the most represented carnivores alongside with cave bears23,24,28. Among the remains, modified by Neanderthals and used as tools, are several bones belonging to the cave lion. These remains, which are quite singular, both for this period and for the animal used, are the subject of this study, which aims to determine their origin and methods of exploitation.
Results
Description of the bone retouchers
The four retouchers made from cave lion remains display typical features indicative of their use (Table 1; Fig. 3): notches, in the shape of scores and pits, are evident on the cortical surface. As per Mallye’s terminology29, these features are either concentrated on the primary use surfaces (Sc1982-348-25 (1), Sc1986-1278-160 (1), Sc1986-1270-203, Sc1983-338-17) or dispersed on secondary ones (Sc1982-348-25 (2), Sc1986-H16-160 (2)). The use surfaces, defined by the concentration of impact, are predominantly located in the distal part of the fragments and can be either centered or lateral as per Mallye’s terminology29. The observed traces on these bone tools primarily consist of rectilinear scores with smoothed-side shapes and triangular pits, suggesting they were likely used on flint29. Still embedded within the cortical bone, lithic chips are present in most of the bone tools. The chemical composition has been recorded for fragments embedded in Sc1986-1278-160 (Fig. 4) using Energy-dispersive X-ray spectroscopy (EDX). The recorded Silicon (Si) peak highlights the presence of exogenous siliceous materials. Unfortunately, this chemical signature does not allow us to distinguish between siliceous materials such as flint, quartz, or quartzite, which are all present in lithic materials collected within sedimentary Unit 520.

The four bone retouchers made on cave lion remains excavated in Scladina Cave Archaeological Assemblage 5—Sc1986-1278-160 (A), Sc1982-348-25 (B), Sc1986-1270-203 (C) and Sc1983-338-17 (D). Detailed picture of Sc1982-348-25 exhibits the typical pits and scores related to the use as a bone retoucher. A and B refit together (see also Fig. 5). As suggested by their relative positions projected onto a left tibia of a cave lion excavated in Goyet (F), the four retouchers, associated with a distal end of the tibia (E), seem to be crafted from a single and unique bone.

Energy-dispersive X-ray (EDX) spectroscopy analysis made on a lithic inclusion embedded in a score located in the main use area of Sc1986-1278-160. The spectroscopy highlights peaks of Si and O, which is consistent with the siliceous material. The mapping of Si highlights in purple the targeted lithic chips.
The frequencies of marks suggest a variable use of these tools, where one of the use surfaces (Sc1986-1278-160 (2)) appears to have not met the requested needs, as the marks are few and dispersed, unlike the other surface on the same tool that has been more intensively utilized.
The blanks exhibit typical fresh bone fracture patterns30,31, and this relative freshness is also evidenced by cut marks present on all blanks. Furthermore, scraping marks observed on the use area of Sc1983-338-17 suggest the removal of the periosteum prior to its use as a bone tool. The surfaces are well preserved, with minimal erosion and weathering. The spongiosa still present on Sc1986-1270-203 is also perfectly preserved. This good state of preservation allowed refitting between tools Sc1982-348-25 and Sc1986-1278-160 without significant material loss. No alterations linked to carnivore activity or advanced weathering stage was detected on the tools, nor on the other lion remains uncovered in this unit.
Additional insights into their use are provided by impact points visible on some supports, as well as flake scars. However, among these bone tools, Sc1986-1278-160 and Sc1982-348-25 stand out (Fig. 5). Their refitting reconstitutes a fragment of left tibia diaphysis, which appears to have been carefully prepared and used (Fig. 3). After, the bone was deliberately fractured and the fragments used separately as retouchers, as previously observed with retouchers made from cave bear remains within the same context23. The diaphyseal tibia fragment exhibits distinct features on its apical and basal ends. The basal end has been intentionally shaped by bifacial retouching, resulting in a bevel form, contrasting with what appears to be a shattered splinter from a blow to the apical part of the blank. This bone fragment could have functioned as an intermediate bone tool, such as a chisel32,33,34. The most prominent part (Fig. 5F) shows slight polishing and chipping that do not seem related to the shaping of the piece. After closer inspection under high magnification (up to 200 x), there is some polish and rounding at the edge of the tool that is connected between the two sides of the breakage of the bone. This polish is smoother and more compact than the natural surface. The distribution of this polish is also limited to the edge of the tool, spreading a bit into the background. These observations indicate that the polish is connected to the use of the tool rather than production traces or post-depositional surface modification. From the directionality of the polish and some small edge removals that also seem connected to use, the directionality of the movement is transversal. The observed polish is smooth(ening), domed, bright, somewhat fluid, with transversal and parallel directionality, and few shallow, short, wide striations in a transversal directionality in the background and parallel on the corner of the edge. Although the shape resembles chisels previously mentioned in the literature33, it is still too early to assign a precise function to it. The recurring fracturing observed on long bones, particularly from chamois, to extract marrow could be one possibility, as well as the splitting of woody material along the grain. Based on the polish attributes, for now we suggest the use on bone, but further experimental and use-wear analyses are necessary for the clearer understanding of the function of this tool. In summary, the distribution, location and sequencing of the various intentional modifications—bifacial reshaping for making a chisel-like tool followed by intentional fracturing and subsequent use of the isolated fragments as bone retouchers—impart successive utilitarian functions to these artifacts.

The refitting of Sc1982-345-25 and Sc1986-1278-160 allows for the reconstruction of a diaphysis fragment of a left tibia (A,B). The bifacial reshaping observed on the distal part (C,D) presents a polish (F) that occurred prior to the breakage of the bone as it is present on both side of the fracture. Combined with blow fractures observed on the proximal part (A,B), these are all indicators supporting the multifunctional use of this tibial fragment as an intermediate tool as suggested in the drawing by S. Lambermont (E), before the different fragments were used separately as retouchers as illustrated in Figs. 3A,B
Despite a relatively diverse faunal assemblage in the sedimentary Unit 526,28, there is limited diversity in the species used for manufacturing bone retouchers. Most of these tools were made from cave bear remains (17). While some specimens have yet to be identified to the species level, the use of other taxa—such as woolly rhinoceros (Coelodonta antiquitatis), reindeer (Rangifer tarandus), horse (Equus caballus) or Bos/Bison (Bos primigenius/Bison priscus)—is anecdotal, as only one tool could be associated with each of these species (Table 2). The second most represented species is the cave lion, with four tools attributed to it, however, these four tools could belong to a single individual.
In addition to the variety of species used, Neanderthals selected a range of anatomical elements to craft their tools, including fragments of metapodials, ribs, femurs, and tibias. These elements share a common characteristic: they were relatively fresh at the time of shaping and use. While it is possible that the mechanical properties of bones vary depending on the anatomical element and/or species selected, a comparison of the dimensions of the different tools (Fig. 6) reveals no significant size differences between the species used. Despite taxonomic variability—ranging from Ursus spelaeus to Panthera spelaea, Bos/Bison, and Rangifer tarandus—the tools exhibit a consistent dimensional range. The consistent size range across different taxa suggests that Neanderthals prioritized the size and shape of the blank over the species from which it derived. This degree of dimensional standardization implies a functional criterion in raw material selection, likely aimed at optimizing tool ergonomics and efficiency. Thus, the selection process reflects deliberate choices rooted in practical concerns rather than species-specific preferences, highlighting a sophisticated understanding of material properties and tool design.

The size distribution of the bone retouchers from Scladina Unit 5 does not exhibit major differences between the species used.
Species identification
Sc1986-1278-160 was originally thought to have been made from a cave bear tibia, based on morphological approach, and was published as such23,35. However, the popliteal line appeared to be very prominent, raising doubts about its identification. As with other bone tools (Table 2), it was decided to carry out a series of additional analyses designed to objectify the identifications (ZooMS and LC–MS/MS).
Zooarchaeology by mass spectrometry (ZooMS)
ZooMS analysis of the retouchers shows that they were made from Ursus, Panthera, Panthera/Crocuta, Rangifer, Bos/Bison and Stephanorhinus or Coelodonta bones. The Table 2 shows the list of retouchers and the ZooMS identification and the Supp. File S1 contains the ZooMS results. The identifications were based on database form Sam Presslee in University of York (https://docs.google.com/spreadsheets/d/1ipm9fFFyha8IEzRO2F5zVXIk0ldwYiWgX5pGqETzBco/edit?gid=1005946405#gid=1005946405). The distinction between Panthera and Hyaenidae was achieved using the peptide COL1A2T41 (E)36. For Panthera the mass of the peptide E was m/z 2820.354726 and m/z 2792.32343 for Hyaenidae. Only the identification of the retoucher Sc1986-1270-203 is ambiguous, and it is identified as either Panthera or Hyaenidae. The averages of deamidation values are 51.6% + /- 7.4% and 53.1% + /- 7.3% for bone fraction and acid fraction, respectively. These high deamidation values indicate that they are old proteins and correlate with the deamidation value for other analysis by paleoproteomics on Scladina cave bones37. To ensure identification of the retouchers identified as Panthera and those with Panthera/Crocuta ambiguity, LC–MS/MS analyses were carried out.
Liquid chromatography–mass spectrometry (LC–MS/MS)
The samples number Sc1982-348-25, Sc1983-338-17, Sc1986-1270-203 and Sc1986-1278-160 have been analyzed by LC–MS/MS to validate the ZooMS results (Supp. File S2). Panthera leo type 1 collagen was identified with the highest score when the database containing COL1A1 and COL1A2 (1624 sequences) was queried. Single Amino acid variations at position 1071 have been identified on COL1A2 sequences for all samples. These variations have been identified on Panthera spelaea bones by proteomics in Bray and collaborators37. These results were confirmed using the ClassiCOL software, which identified the Panthera genus for all 4 samples (Supp. File S3, Figures S1–S4). PEAKSX analysis of sample Sc1983-382-120 identified the retoucher as Bos/Bison. The COL1A1 and COL1A2 proteins are identical between Bos taurus and Bison bison or Bison bonasus. However, COL3A1 contains SAPs (single amino polymophisms) and the GEPGAPGLK sequencing peptide (positions 120–128) is specific to Bos taurus COL3A1 (Supp. File S3, Figures S5–6). this identification has been validated with ClassiCOL software (Supp. File S3, Figure S7).
Discussion and conclusions
The relationship between humans and large felids has been complex and multifaceted throughout history. In many cultures, lions, for instance, are both revered and feared: they are admired for their strength and majesty but are also considered threats to livestock and human safety. This ambivalence endures through time and space38. In the Upper Paleolithic, lions held profound symbolic value, as evidenced by iconic artworks like the Chauvet Cave paintings (France) or the Löwenmensch figurine from Hohlenstein-Stadel (Germany). Yet such symbolic associations are difficult to attribute to Neanderthal contexts, where evidence remains sparse and primarily functional. At Scladina Cave, the deliberate transformation of cave lion bones into tools reveals a markedly different dynamic—one grounded not in symbolism but in pragmatism and adaptive behavior.
The cave lion is primarily represented by limb bone fragments (Table 3), and the remains uncovered thus far do not suggest the presence of more than a single adult individual. Aside from the elements used as tools, no clear anthropogenic modifications are visible on the other preserved bones. However, the refitting of the tibia fragments emphasizes the intentional nature of the process—going beyond simple butchery waste recovery—and reflects the care invested in tool production. Alongside retouchers made from a cave bear femur, this case exemplifies the execution of a structured operational sequence (chaîne opératoire), like those commonly observed in lithic industries.
The selection of long bones for crafting bone tools aligns with broader Neanderthal technological behavior, as these elements were typically preferred across sites and species24,39,40. The findings from Unit 5 thus represent the earliest known evidence of bone tools made from cave lion remains, with the tibia-based artifacts demonstrating deliberate shaping, fracturing, and subsequent reuse as retouchers—further underscoring the complexity of Neanderthal technological practices.
Importantly, the morphometric analysis of all the retouchers found in Scladina Unit 5, spanning several species, reveals that Neanderthals employed a standardized approach to toolmaking. Despite the taxonomic diversity of the raw materials, including Ursus spelaeus, Panthera spelaea, Bos sp., Rangifer tarandus, and others, the dimensions of the retouchers remained consistent, indicating that size, shape, and perhaps structural integrity of the bone blank were the primary criteria for selection. This standardization suggests that Neanderthals were not opportunistically using any available bone but rather were deliberately selecting blanks, sometimes reshaping them, to meet specific functional requirements. The anatomical diversity of the selected bones (metapodials, ribs, femurs, tibias) and their consistent state of freshness at the time of shaping support this interpretation. In this light, the choice of species appears incidental to the practical utility of the material, reinforcing the idea of a function-driven technological tradition rather than one influenced by symbolism or cultural considerations. Moreover, the presence of woolly rhinoceros and reindeer further supports the hypothesis of site occupation during a relatively cold period, consistent with the end of the Saalian (MIS 6), as also indicated by recent chronological data9.
The deliberate exploitation of carnivore remains by Neanderthals at Scladina Cave is well-documented, particularly through anthropogenic modifications observed on cave bear bones23. While the abundance of bear remains suggests that Neanderthals may have hunted or scavenged animals that died naturally within the cave, the origin of the cave lion remains is more ambiguous. The bones of this individual show no evidence of natural alterations—such as erosion, weathering, or trampling—nor of carnivore activity. In addition, anthropogenic modifications including fractures, percussion notches, and cut marks indicate that the carcass was relatively fresh at the time of processing, a conclusion supported by the successful refitting of bone fragments. This raises the possibility that the lion was either hunted or recovered shortly after death, potentially reflecting a confrontation between Neanderthals and one of their main ecological competitors.
Although the possibility of opportunistic scavenging from a naturally deceased animal cannot be entirely excluded, the strategic use of the lion remains and the absence of taphonomic alterations strongly support a more active role. Given the extent of the excavations at Scladina, the meticulous recovery methods, and the spatial distribution of the lion bones—primarily concentrated in the first chamber of the cave, which is nearly completely excavated—the most plausible scenario is that Neanderthals deliberately brought a partial carcass into the site for processing. Whether this reflects hunting, scavenging, or defensive confrontation remains unresolved. However, if hunting was indeed involved, the act would carry considerable symbolic weight due to the high risk posed by such a formidable predator, especially compared to more commonly hunted prey like cervids or chamois16. These findings underscore Neanderthal behavioral flexibility and their capacity to exploit even dangerous carnivores for both functional and possibly symbolic purposes.
Ultimately, the discoveries at Scladina enrich our understanding of Neanderthal interaction with their environment and with other large predators. The integration of technological, zooarchaeological, and biomolecular data reveals not only the practical exploitation of carnivores but also a refined capacity for planning, selection, and reuse. These behaviors attest to Neanderthals’ advanced cognitive abilities and underscore their place as innovative, adaptive hominins capable of navigating complex ecological relationships.
The discovery at Scladina of bone retouchers crafted from cave lion remains represents an extraordinary and unparalleled finding within the Paleolithic archaeological record. It not only demonstrates the Neanderthals’ capacity to selectively exploit available resources, including those derived from large carnivores, but also reflects their ability to transform such remains into multifunctional tools following a structured operational sequence. The use of a predator as iconic and potentially dangerous as the cave lion—whether through active hunting or opportunistic recovery—reveals a high degree of behavioral flexibility and a sophisticated understanding of the technical properties of osseous materials. Beyond their rarity, these artifacts prompt a reevaluation of Neanderthal interactions with large carnivores, emphasizing that these animals were not merely ecological competitors but could also serve practical and possibly symbolic purposes in Neanderthal lifeways.
Methods
The material unearthed in Unit 5 underwent a systematic review aimed at identifying potential bone retouchers. This fruitful analysis documented a relatively intense exploitation of cave bears, although other species were also identified but are less well represented23,24,41. This study notably proposed a genuine debitage sequence on a cave bear femur, demonstrating both access to a relatively fresh carcass and a series of reasoned actions to extract several supports from the diaphysis transformed into tools23. Although refitting and morphological studies identified numerous species, several tools could not be identified at the specific level due to the significant modifications applied by Neanderthals. Consequently, we resorted to proteomic analyses (Table 2), which had previously been tested and yielded conclusive results37.
The cave lion remains excavated so far seem to belong to a single left tibia of an adult specimen, according to the degree of ossification and fusion of the epiphyses. The individual is mainly represented by limb bones (Table 3).
Most of the cave lion remains are distributed in the first part of the cave, close to the current porch (Fig. 7). This concentration is the same as that already observed for tools made from cave bear remains23.

Spatial distribution of the cave lion remains (Table 2). Cranial fragments are figured by yellow diamonds, left lower limb by blue squares, right lower limb by red squares, right upper limb in blue dots, and non-lateralized bones by green dots.
Microwear analysis
The samples Sc1986-1278-160 and Sc1982-348-25 were subjected to preliminary microwear analysis. The surface and the edges of the tool were examined and photographed with a Zeiss AxioZoom V16 macroscope with magnification range between 7-112x. Zeiss ZEN Core software (v.3.4; https://www.zeiss.com/microscopy/en/products/software/zen-core.html) was used to acquire the micrographs. Areas of interest were also assessed with an Olympus BX53M microscope under 50 ×, 100 ×, and 200 × magnifications. Micrographs were taken with the use of a Nikon D750 DSLR camera and Best Scientific 1.9X coupler and captured with Helicon Remote software (v.4.5.3; https://www.heliconsoft.com/) and processed with Helicon Focus software (v.8.3.4; https://www.heliconsoft.com/).
Nitrogen content analysis
A small fragment of the bone retoucher Sc1983-338-17 has been submitted to a nitrogen content analysis to evaluate the possibility to extract collagen. This extraction attempt was made to perform carbon and nitrogen isotopic analyses and compare them to cave bears and cave lions’ data from Scladina Cave. Following the methodology presented by Bocherens and colleagues42, 5 mg of bone powder were analyzed using an Elemental Analyzer to measure its nitrogen content, a proxy for collagen in ancient bone. The obtained results (%N = 0.23%) are below the threshold of 0.4%, equivalent to 10% of the original collagen and therefore indicate that the collagen content is insufficient to yield enough collagen for isotopic analysis. No further analysis was attempted on this specimen.
Proteins digestion on plate
Bone proteins were analyzed using methods by Bray and collaborators43. All solutions were eluted through the wells using a vacuum manifold (Merck KGaA, Darmstadt, Germany) pumped by a DS 102 rotary vane pump (Agilent, Santa Clara, USA). The wells of a 0.45 μm hydrophobic-high protein binding (MSIPS4510, Millipore, Billerica, MA, USA) were prepared by washing with 70% ethanol flowed through the filter. In each well, 1–5 mg of bone powder was deposited and 100 µL of demineralization solution (5% v/v TFA (trifluoroacetic acid)) were added. The plate was incubated at 4 °C for 16 h without shaking. The demineralization solution in TFA was recovered in a V-bottom well-collecting plate by applying vacuum. Then 6 µL of NaOH 6 M were added to neutralize the TFA and 100 µL of 100 mM ABC (ammonium bicarbonate) pH 8.8. The plate was kept at 4 °C for digestion. The bone powder in each well was washed three times with 100 µL of digestion buffer (50 mM ABC, pH 8.8). The plate was incubated at 65 °C for 1 h on a heating stirrer MHR23 (Hettich, Tuttlingen, Germany) for collagen gelatinization. The digestion of bone powder and demineralization solution was performed by adding 0.5 μg of sequencing grade trypsin (Promega, Madison, WI, USA) and incubating the mixture at 37 °C for 16 h with shaking on a heating stirrer. Peptides from bone powder were recovered in a V-bottom well-collecting plate by applying vacuum and the plate was washed one time with digestion buffer. Tryptic peptides from bone powder and demineralization solution were acidified with 1 µL of acetic acid (final concentration 0.5% of acetic acid). Tryptic peptides were desalted on 96 wells plates C18 (Affinisep, Petit-Couronne, France). Before analysis, each sample was resuspended in 10 µL of H2O, 0.1% formic acid. The concentration of peptide was measured with DS-11 + (Denovix, Wilmington, USA) at 214 nm.
MALDI analysis
MALDI analyses were conducted using the protocol described by Bray and collaborators43. Desalted peptides (1 µL) were deposited on 384 Ground steel MALDI plates (Bruker Daltonics, Bremen, Germany), then 1 µL of HCCA matrix at 10 mg/mL in ACN/H2O 80:20 v/v 0.1% formic acid was added for each sample spot and dried at ambient temperature. MALDI-FTICR experiments were carried out on a Bruker 9.4 Tesla SolariX XR FTICR mass spectrometer (Bruker Daltonics, Bremen, Germany). A Bruker Smartbeam-II Laser System was used for irradiation at a frequency of 1000 Hz and using the “Minimum” predefined shot pattern. MALDI-FTICR spectra were generated from 500 laser shots in the m/z range from 693.01 to 5000 with 2 M data points (i.e., transient length of 5.0332 s). Twenty spectra were averaged. The transfer time of the ICR cell was set to 1.2 ms and the quadrupole mass filter operating in RF-only mode was set at m/z 600.
LC–MS/MS analysis
LC–MS/MS analyses were performed on an Orbitrap Q Exactive plus mass spectrometer hyphenated to a U3000 RSLC Microfluidic HPLC System (ThermoFisher Scientific, Waltham, Massachusetts, USA) according to the method presented in Bray and collaborators43. 1 μl of the peptide mixture at a concentration of 1 µg/µL was injected with solvent A (5% acetonitrile and 0.1% formic acid v/v) for 3 min at a flow rate of 10 μl.min−1 on an Acclaim PepMap100 C18 pre-column (5 μm, 300 μm i.d. × 5 mm) from ThermoFisher Scientific. The peptides were then separated on a C18 Acclaim PepMap100 C18 reversed phase column (3 μm, 75 µm i.d. × 500 mm), using a linear gradient (5–40%) of solution B (75% acetonitrile and 0.1% formic acid) at a rate of 250 nL.min−1 in 165 min and then 100% of solution B in 5 min. The column was washed for 5 min with buffer B and then re-equilibrated with buffer A. The column and the pre-column were placed in an oven at a temperature of 45 °C. The total duration of the analysis was 185 min. The LC runs were acquired in positive ion mode with MS scans from m/z 350 to 1600 in the Orbitrap mass analyser at 70,000 resolution at m/z 400. The automatic gain control was set at 3E6. Sequentially MS/MS scans were acquired in the high-energy collision dissociation cell for the 15 most-intense ions detected in the full MS survey scan at 35,000 resolution at m/z 400. Automatic gain control was set at 5E5, and the normalized collision energy was set to 30 eV. Dynamic exclusion was set at 30 s and ions with 1 and more than 8 charges were excluded.
Bioinformatics for MALDI data
MALDI FTICR were processed using Compass DataAnalysis (V.5.0.; https://bruker-compass-dataanalysis.software.informer.com) SNAP algorithm was employed with the following parameters of S/N > 3 and quality 0.6. The procedure for the deamidation value calculation from MALDI FTICR was based on Bray and colleagues43 and the identification of taxonomic rank was realized with data from the literature and ZooMS data base from Sam Presslee in University of York (https://docs.google.com/spreadsheets/d/1ipm9fFFyha8IEzRO2F5zVXIk0ldwYiWgX5pGqETzBco/edit?gid=1005946405#gid=1005946405).
Bioinformatics for LC–MS/MS data
Proteomics data were processed with PEAKS X + (https://www.bioinfor.com/peaks-studio-x-plus) against a home-made database containing 1,765 collagen sequences extracted from NCBI database (All_Collagen, downloaded 08-2023) restricted to Mammalian, as already described by Bray and collaborators43. Precursor’s mass tolerance was fixed to 10 ppm and fragment ion mass tolerance to 0.02 Da. Semi-trypsin digestion mode was used. Cysteine carbamidomethylation was set as fixed modification. Methionine oxidation and asparagine, glutamine deamidation and hydroxylation of amino acids (RYFPNKD) were selected as variable modifications. PEAKS PTM and SPIDER ran with the same parameters. Results were filtered using the following criteria: protein score − 10logP ≥ 20, 1% peptide False Discovery Rate (FDR), PTM with Ascore = 20, mutation ion intensity = 5% and Denovo ALC ≥ 50%. Peptides with amino acids substitutions were filtered with minimal intensity set as 1E + 7.
A second bioinformatic analysis was done with Mascot against Mammalian database from NCBI containing 9,016,701 sequences. Peptide mass tolerance and fragment mass tolerance were fixed to 10 ppm. Semi-trypsin digestion mode was used. Cysteine carbamidomethylation was set as fixed modification. Proline oxidation and asparagine and glutamine deamidation were selected as variable modifications. CSV file were exported and analyzed with ClassiCOL (V.1.0.2; https://github.com/EngelsI/ClassiCOL/tree/main/ClassiCOL_version_1_0_2) with default parameters44.
Energy-dispersive X-ray (EDX) spectroscopy
Scanning electron microscope (SEM) images were acquired using a Quanta 200 ESEM, manufactured by FEI. X-ray emissions associated with imaging were collected using an energy dispersive X-ray spectrometer (EDS APOLLO 10, Silicon Drift detector by EDAX), enabling semi-quantitative chemical analysis of elements ranging from beryllium (Z = 4) to uranium (Z = 92).
The imaging was conducted under low-vacuum conditions (15 Pa) of water vapor, with a 30 kV incident electron beam and a working distance of 13 mm. The SEM images were produced by detecting backscattered electrons (BSE), allowing the detection of different phases based on their molecular weights.
For chemical analysis, EDS data can be obtained either as individual data points or through chemical mapping. This combination of techniques offers a rapid, non-destructive method for analyzing and distriminating between lithic materials.
Data availability
The mass spectrometry proteomics data (MALDI-FTICR, LC–MS/MS raw data and PEAKS X + results) have been deposited on the ProteomeXchange Consortium (http://proteomecentral.proteomexchange.org) via the PRIDE partner repository47 with the data set identifier PXD055624.
References
Diedrich, C. G. Palaeopopulations of late pleistocene top predators in Europe: Ice age spotted hyenas and steppe lions in battle and competition about prey. Paleontol. J. 2014, 106203. https://doi.org/10.1155/2014/106203 (2014).
Bocherens, H. et al. Palaeoenvironmental and palaeodietary implications of isotopic biogeochemistry of last interglacial neanderthal and mammal bones in Scladina Cave (Belgium). J. Archaeol. Sci. 26, 599–607 (1999).
Blasco, R., Rosell, J., Arsuaga, J. L., Berm dez de Castro, J. M. & Carbonell, E. The hunted hunter: The capture of a lion (Panthera leo fossilis) at the Gran Dolina site, Sierra de Atapuerca, Spain. J. Archaeol. Sci. 37, 2051–2060 (2010).
Van Kolfschoten, T., Parfitt, S. A., Serangeli, J. & Bello, S. M. Lower Paleolithic bone tools from the ‘Spear Horizon’ at Schöningen (Germany). J. Human Evol. 89, 226–263. https://doi.org/10.1016/j.jhevol.2015.09.012 (2015).
Serangeli, J., Van Kolfschoten, T., Starkovich, B. M. & Verheijen, I. The European saber-toothed cat (Homotherium latidens) found in the “Spear Horizon” at Schöningen (Germany). J. Human Evol. 89, 172–180. https://doi.org/10.1016/j.jhevol.2015.08.005 (2015).
Russo, G. et al. First direct evidence of lion hunting and the early use of a lion pelt by Neanderthals. Sci. Rep. 13, 16405. https://doi.org/10.1038/s41598-023-42764-0 (2023).
Blasco, R. et al. Environmental availability, behavioural diversity and diet: a zooarchaeological approach from the TD10–1 sublevel of Gran Dolina (Sierra de Atapuerca, Burgos, Spain) and Bolomor Cave (Valencia, Spain). Quat. Sci. Rev. 70, 124–144. https://doi.org/10.1016/j.quascirev.2013.03.008 (2013).
Beauval, C. La faune des niveaux Mousteriens de ‘Chez-Pinaud’ (Jonzac, Charente-Maritime, France). Première analyse. In Le Site Paléolithique de Chez-Pinaud à Jonzac, Charente-Maritime Vol. Supplément 8 (ed J. Airvaux) 125–156 (Préhistoire du Sud-Ouest 2004, 2004).
Vonhof, H. et al. Improving the age constraints on the archeological record in Scladina Cave (Belgium): new speleothem U-Th ages and paleoclimatological data. Clim. Past Discuss. 2024, 1–30. https://doi.org/10.5194/cp-2024-27 (2024).
Pirson, S. Contribution à l'étude des dépôts d’entrée de grotte en Belgique au Pléistocène supérieur. Stratigraphie, sédimentologie et paléoenvironnement Doctorat thesis, Université de Liège (2007).
Pirson, S., Court-Picon, M., Haesaerts, P., Bonjean, D. & Damblon, F. New data on geology, anthracology and palynology from the Scladina Cave pleistocene sequence: Preliminary results. Mem. Geol. Surv. Belg. 55, 71–93 (2008).
Pirson, S. The stratigraphic sequence of Scladina Cave. In The Scladina I-4A Juvenile Neandertal (Andenne, Belgium). Palaeoanthropology and Context Vol. 134 (eds Michel Toussaint & Dominique Bonjean) 49–68 (Etudes et Recherches Archéologiques de l’Université de Liège, 2014).
Pirson, S. et al. The palaeoenvironmental context and chronostratigraphic framework of the Scladina Cave sedimentary sequence (units 5 to 3-SUP). In The Scladina I-4A Juvenile Neandertal (Andenne, Belgium). Palaeoanthropology and Context Vol. 134 (eds Michel Toussaint & Dominique Bonjean) 69–92 (Etudes et Recherches Archéologiques de l’Université de Liège, 2014).
Abrams, G. et al. Investigating the co-occurrence of Neanderthals and modern humans in Belgium through direct radiocarbon dating of bone implements. J. Hum. Evol. 186, 103471. https://doi.org/10.1016/j.jhevol.2023.103471 (2024).
Di Modica, K. & Bonjean, D. Scladina (Sclayn, province de Namur): Ensembles lithiques moustériens méconnus. Notae Praehistoricae 24, 5–8 (2004).
Moncel, M.-H., Patou-Mathis, M. & Otte, M. Halte de chasse au chamois au Paléolithique moyen : la couche 5 de la grotte Scladina (Sclayn, Namur, Belgique). In Economie préhistorique : Les comportements de subsistance au Paléolithique. XVIIIe Rencontres Internationales d’Archéologie et d’Histoire d’Antibes 291–308 (Editions APDCA, Sophia Antipolis, 1998).
Toussaint, M. & Pirson, S. Scladina I-4A in the chronological context of the Neandertals from the Belgian Meuse Valley and northwest Europe. In The Scladina I-4A Juvenile Neandertal (Andenne, Belgium). Palaeoanthropology and Context Vol. 134 (eds Michel Toussaint & Dominique Bonjean) 395–408 (Etudes et Recherches Archéologiques de l’Université de Liège, 2014).
Toussaint, M., Bonjean, D. & Pirson, S. The Scladina I-4A juvenile Neandertal: A synthesis. In The Scladina I-4A Juvenile Neandertal (Andenne, Belgium). Palaeoanthropology and Context Vol. 134 (eds Michel Toussaint & Dominique Bonjean) 409–418 (Etudes et Recherches Archéologiques de l’Université de Liège, 2014).
Huxtable, J. & Aitken, M. J. Thermoluminescence dating of burned flint and stalagmitic calcite. In Recherches aux grottes de Sclayn. Vol 1. Le Contexte Vol. 27 Etudes et Recherches Archéologiques de l’Université de Liège (ed Marcel Otte) 175–178 (Etudes et Recherches Archéologiques de l’Université de Liège, 1992).
Di Modica, K., Toussaint, M., Abrams, G. & Pirson, S. The Middle palaeolithic from Belgium: Chronostratigraphy, territorial management and culture on a mosaic of contrasting environments. Quatern. Int. 411, 77–106. https://doi.org/10.1016/j.quaint.2015.12.072 (2016).
Di Modica, K. Les productions lithiques du Paléolithique moyen de Belgique: Variabilité des systèmes d’acquisition et des technologies en réponse à une mosaïque d’environnements contrastés Doctorat thesis, Université de Liège et Muséum d’Histoire Naturelle de Paris, (2010).
Moncel, M.-H. L'industrie lithique de la grotte Scladina (Sclayn). La couche moustérienne eemienne 5. Les comportements techniques et les objectifs de la production dans un Moustérien de type Quina. In Recherches aux grottes de Sclayn. Volume 2. L'Archéologie Vol. 79 Etudes et Recherches Archéologiques de l’Université de Liège (eds Marcel Otte, Marylène Patou-Mathis, & Dominique Bonjean) 181–247 (Université de Liège, 1998).
Abrams, G., Bello, S. M., Di Modica, K., Pirson, S. & Bonjean, D. When Neanderthals used cave bear (Ursus spelaeus) remains: Bone retouchers from unit 5 of Scladina Cave (Belgium). Quat. Int. 326–327, 274–287. https://doi.org/10.1016/j.quaint.2013.10.022 (2014).
Abrams, G. Palaeolithic bone retouchers from Belgium: a preliminary overview of the recent research through historic and modern bone collections. In The Origins of Bone Tool Technologies Vol. 35 RGZM – TAGUNGEN (eds J.M. Hutson et al.) 197–214 (Verlag des Römisch-Germanisches Zentralmuseums - Leibniz-Forschungsinstitut für Archäologie, 2018).
Patou-Mathis, M. Les espèces chassées et consommées par l’homme en couche 5. In Recherches aux grottes de Sclayn. Volume 2. L'Archéologie Vol. 79 Etudes et Recherches Archéologiques de l’Université de Liège (eds Marcel Otte, Marylène Patou-Mathis, & Dominique Bonjean) 297–310 (Université de Liège, 1998).
Patou-Mathis, M. & Bocherens, H. Comportements alimentaires des hommes et des animaux à Scladina. In Recherches aux grottes de Sclayn. Volume 2. L'Archéologie Vol. 79 Etudes et Recherches Archéologiques de l’Université de Liège (eds Marcel Otte, Marylène Patou-Mathis, & Dominique Bonjean) 329–336 (Université de Liège, 1998).
Bonjean, D., Abrams, G. & di Modica, K. Andenne/Sclayn: Grotte Scladina, une preuve de la chasse au lièvre au Paléolithique moyen. Chronique de l’Archéologie wallonne 19, 209–211 (2012).
Patou-Mathis, M. Origine et histoire de l’assemblage osseux de la couche 5. Comparaison avec la couche 4 sus-jacente, non-anthropique. In Recherches aux grottes de Sclayn. Volume 2. L'Archéologie Vol. 79 Etudes et Recherches Archéologiques de l’Université de Liège (eds Marcel Otte, Marylène Patou-Mathis, & Dominique Bonjean) 281–295 (Université de Liège, 1998).
Mallye, J.-B. et al. The Mousterian bone retouchers of Noisetier Cave: Experimentation and identification of marks. J. Archaeol. Sci. 39, 1131–1142 (2012).
Fernandez-Jalvo, Y. & Andrews, P. Atlas of Taphonomic Identifications 1001+ Images of Fossil and Recent Mammal Bone Modification. 1st edn (2016).
Lyman, R. L. Vertebrate Taphonomy (Cambridge University Press, 1994).
Kozlikin, M. B., Rendu, W., Plisson, H., Baumann, M. & Shunkov, M. V. Unshaped bone tools from denisova Cave, Altai. Archaeol. Ethnol. Anthropol. Eurasia Arkheol. Ehtnografija Antropologija Evrazii 48, 16–28. https://doi.org/10.17746/1563-0110.2020.48.1.016-028 (2020).
Tartar, E. The recognition of a new type of bone tools in Early Aurignacian assemblages: Implications for understanding the appearance of osseous technology in Europe. J. Archaeol. Sci. 39, 2348–2360. https://doi.org/10.1016/j.jas.2012.02.003 (2012).
Tejero, J.-M. et al. New insights into the Upper Palaeolithic of the Caucasus through the study of personal ornaments. Teeth and bones pendants from Satsurblia and Dzudzuana caves (Imereti, Georgia). PLoS ONE 16, e0258974. https://doi.org/10.1371/journal.pone.0258974 (2021).
Abrams, G. et al. Les retouchoirs en os de l’ensemble sédimentaire 5 de Scladina: Utilisation des restes d’ours des cavernes (Ursus spelaeus). Notae Praehistoricae 34, 5–19 (2014).
Ruebens, K., Discamps, E., Smith, G. & Hublin, J.-J. Integrating ZooMS and zooarchaeology to assess the châtelperronian and carnivore occupations at cassenade (Dordogne, France). PaleoAnthropology 2024, 263–281. https://doi.org/10.48738/2024.iss2.1101 (2024).
Bray, F. et al. Extinct species identification from late Middle Pleistocene and earlier Upper Pleistocene bone fragments and tools not recognizable from their osteomorphological study by an enhanced proteomics protocol. Archaeometry 65, 196–212. https://doi.org/10.1111/arcm.12800 (2023).
Somerville, K. Humans and Lions: Conflict, Conservation and Coexistence (2019).
Toniato, G., Münzel, S. C., Starkovich, B. M. & Conard, N. J. In The Origins of Bone Tool Technologies Vol. 35 RGZM – TAGUNGEN (eds J.M. Hutson et al.) 251–268 (Verlag des Römisch-Germanisches Zentralmuseums - Leibniz-Forschungsinstitut für Archäologie, 2018).
Sévêque, N. & Auguste, P. The Origins of Bone Tool Technologies Vol. 35 (eds Hutson, J. M. et al.) 133–164 (Verlag des Römisch-Germanisches Zentralmuseums - Leibniz-Forschungsinstitut für Archäologie, 2018).
Abrams, G. Close Encounters of the Third Kind? Neanderthal and modern humans in Belgium, a bone story. PhD thesis, Universiteit Leiden (2023).
Bocherens, H., Drucker, D., Billiou, D. & Moussa, I. Une nouvelle approche pour évaluer l’état de conservation de l’os et du collagène pour les mesures isotopiques (datation au radiocarbone, isotopes stables du carbone et de l’azote). Anthropologie 109, 557–567. https://doi.org/10.1016/j.anthro.2005.06.005 (2005).
Bray, F. et al. Robust high-throughput proteomics identification and deamidation quantitation of extinct species up to pleistocene with ultrahigh-resolution MALDI-FTICR mass spectrometry. Anal. Chem. 95, 7422–7432. https://doi.org/10.1021/acs.analchem.2c03301 (2023).
Engels, I. et al. Classification of collagens via peptide ambiguation, in a paleoproteomic LC-MS/MS-based taxonomic pipeline. J. Proteome Res. 24, 1907–1925. https://doi.org/10.1021/acs.jproteome.4c00962 (2025).
Vizcaíno, J. A. et al. The PRoteomics IDEntifications (PRIDE) database and associated tools: Status in 2013. Nucleic Acids Res. 41, D1063–D1069. https://doi.org/10.1093/nar/gks1262 (2012).
Acknowledgements
The Scladina Cave permanent excavation program is managed by the Espace muséal d’Andenne with the support of the Wallonia-Brussels Federation, the City of Andenne and the Wallonia Heritage Agency. Authors deeply thank Sylviane Lambermont for her drawing as well as Michel Toussaint who guided her throughout its elaboration. We also wanted to thank Justin Coppe for giving us his first opinion on the intermediate tool and encouraging us to go furthermore into a developed experimental archaeological program. We thank Patrick Semal and Annelise Folie of the Royal Belgian Institute of Natural Sciences for facilitating access to their imaging facility and digital data and Aurore Mathys for operating the 3D scan of the cave lion left tibia from Goyet (specimen IRSNB Vert-00-2182). The authors acknowledge the IBiSA and ProFi network for financial support of the UAR 3290 (MSAP) proteomics facility. The mass spectrometers were funded by University of Lille, CNRS, Région Hauts-de-France and the European Regional Development Fund. The authors deeply thank the financial support from the IR INFRANALYTICS FR2054 CNRS for conducting the research is gratefully and the CNRS MITI project (Mission pour les initiatives transverses et interdisciplinaires) named PBPHomo. Tristan Dedrie is a holder of a special PhD fellowship of the Research Foundation - Flanders (FWO-Vlaanderen) [grant number 11Q0H24N].
Author information
Authors and Affiliations
Contributions
G.A., P.A. and F.B. designed the study; G.A., P.A., I.D.G., C.P., T.D., C.M., V.F., H.B., F.B., É.H., S.P. and K.D.M. performed all data collection and analyzed them; G.A., I.D.G., S.P., H.B., É.H. and F.B. wrote the main manuscript text with the input from all authors; G.A., S.P., K.D.M., Y.V., É.H. and F.B. prepared all the figures; All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abrams, G., Auguste, P., Pirson, S. et al. Earliest evidence of Neanderthal multifunctional bone tool production from cave lion (Panthera spelaea) remains. Sci Rep 15, 24010 (2025). https://doi.org/10.1038/s41598-025-08588-w
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-08588-w
