
Abstract
A new dinosaur assemblage from the Lower Jurassic at Wande Town, Wuding County, Yunnan Province, China is discovered recently. Here a new sauropodomorph from this site, Wudingloong wui gen. et sp. nov., is described on the basis of a partial skeleton. Wudingloong is different from other non-sauropodiform sauropodomorphs particularly in having an ascending ramus of the maxilla excavated by triangular antorbital fossa, distinct ventral keel on the middle cervical centra, a slender humerus with a flat and low humeral head, a gracile metacarpal V with the proximal end as wide as the distal end, and a large and robust ungual of manual digit I. Wudingloong was excavated from the lowest Lower Jurassic Yubacun Formation, which is the lowest dinosaur bone bed in East Asia. Both the phylogenetic analysis and stratigraphic horizon indicate that Wudingloong represents the earliest-diverging and stratigraphically oldest sauropodomorph dinosaur discovered in East Asia so far. The discovery of this new taxon provides further evidence that the southwestern China sauropodomorph assemblage is one of the most taxonomically diverse and morphologically disparate in the pre-Toarcian Early Jurassic worldwide, represented by various taxa from near the base of the Massopoda to non-sauropodan sauropodiforms.
Similar content being viewed by others

Introduction
Chinese non-sauropodan sauropodomorphs are mainly known in the Lower Jurassic Lufeng Formation in Lufeng and its neighboring areas, Yunnan Province, and they are represented by Lufengosaurus, Yunnanosaurus, Jingshanosaurus, Xingxiulong, Yizhousaurus, and some other taxa1,2,3,4,5,6,7,8,9,10. Qianlong, which was reported from the Lower Jurassic Ziliujing Formation of neighboring Guizhou Province, represents a recent discovery expanding our understanding on the distribution of the non-sauropodan sauropodomorphs in China11.
Since September 2020, a previously unknown dinosaur assemblage has been found from the Lower Jurassic at Wande Town, Wuding County, Yunnan Province. Here, we report a new non-sauropodan sauropodomorph from the lowest Lower Jurassic Yubacun Formation12, which is the first prepared dinosaur specimen from this new assemblage in Wuding and to our knowledge represents the earliest dinosaur bone discovered in East Asia. This discovery, which provides insights into Yunnan dinosaur provincialism, improves our understanding on the early evolution of sauropodomorphs and extends the earliest record of East Asian dinosaur assemblage around the Triassic-Jurassic boundary.
Results
Systematic palaeontology
Dinosauria Owen, 1842.
Saurischia Seeley, 1887.
Sauropodomorpha Huene, 1932.
Massopoda Yates, 2007.
Wudingloong wui gen. et sp. nov.
Etymology
The genus name, Wudingloong, “Wuding” refers to Wuding County where the fossil site was found, and “loong” means Chinese “龙”. The specific name, wui, honors Dr. Xiao-Chun Wu (Canadian Museum of Nature) for his significant contribution to the research on the dinosaurs and other vertebrate fossils from Yunnan Province.
Holotype
LFGT-YW002, housed at Center for Dinosaur Research and Protection, Bureau of Natural Resources of Lufeng City, is a partial skeleton including skull and mandible, atlas-axis, third to the tenth cervical vertebrae, first to the seventh dorsal vertebrae, dorsal ribs, right scapula and coracoid, right forelimb including humerus, ulna, radius, and manus (Figs. 1 and 2). The fully co-ossified cranial elements and completely closed neurocentral sutures of the vertebrae demonstrates that the specimen was probably a mature individual13.

Reconstructed skeleton and representative bones of Wudingloong wui gen. et sp. nov. (LFGT-YW002). (A) Cranium in left lateral view. (B) Axis and third cervical vertebra in lateral view. (C) Sixth cervical vertebra in lateral view. (D) Seventh cervical vertebra in lateral view. (E) First to third dorsal vertebrae in lateral view. (F) Right coracoid and scapula in lateral view. (G) Right humerus in posterior view. (H) Right ulna in lateral view. (I) Right manus in ventral view. I–V, manual digit I–V; ai, axial intercentrum; ati, atlantal intercentrum; cf, coracoid foramen; ct, coracoid tubercle; hh, humeral head; mc III–V, metacarpal II–V; ns, neural spine; od, odontoid; ole, olecranon process; vk, ventral keel. Individual scale bars equal 5 cm; scale bar of the reconstructed skeleton equals 50 cm.

Cranium of Wudingloong wui gen. et sp. nov. (LFGT-YW002). (A) Photograph of the cranium in left lateral view. (B) Interpretative drawing of the cranium in left lateral view. (C) Photograph of the cranium in right lateral view. (D) Interpretative drawing of the cranium in right lateral view. an, angular; aof, antorbital fossa; ar, articular; d, dentary; j, jugal; l, lacrimal; lf, left frontal; lpf, left prefrontal; m, maxilla; p, parietal; pf, prefrontal; pm, premaxilla; po, postorbital; pop, paraoccipital process; q, quadrate; qj, quadratojugal; rf, right frontal; rpf, right prefrontal; sq, squamosal; sa, surangular; stf, supratemporal fenestra; n, nasal. Scale bars equal 5 cm.
Locality and horizon
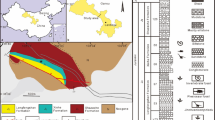
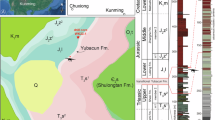
The specimen was excavated from the Lower Jurassic Yubacun Formation (Hettangian to early Sinemurian, probably 200.17 Ma14) at Wande Town, Wuding County, Yunnan Province, China (Fig. S1A). The Yubacun Formation, which was established by Pang et al.12 in Lufeng City of central Yunnan, is characterized by alternations of gray-green, light gray fine-grained sandstone and purple-red mudstone. The central Yunnan region is located southwest of the Tibetan Plateau. A series of Jurassic basins developed in central Yunnan, such as Chuxiong Basin and Lufeng Basin. These basins commonly rest on the pre-Cambrian basement across the western Yangtze craton15 and are filled with thousands of meters of terrestrial deposits, known as the Central Yunnan Jurassic red beds, which were divided into two subregions: the Chuxiong subregion and the Kunming subregion16,17. Wuding dinosaur fossil site is located in the Kunming subregion. In this subregion, the lithosequence is divided in ascending order into the Lower Jurassic Yubacun Formation and Lufeng Formation, the Middle Jurassic Chuanjie Formation and Laoluocun Formation, and the Upper Jurassic Madishan Formation and Anning Formation (Fig. S1B). The Yubacun Formation, which represents the lowest Jurassic sequence, conformably overlies the gray-green thick-bedded sandstone of the Upper Triassic Shezi Formation and is conformable with the purple-red thick-bedded sandstone and mudstone of the Lower Jurassic Lufeng Formation. Invertebrate fossils found in the Yubacun Formation, such as bivalve Psilunio, gastropods Lufengospira, and ostracods Gomphocythere, indicate an Early Jurassic in relative age12. A paleomagnetic study has constrained the Yubacun Formation to an age of Hettangian to early Sinemurian18.
Diagnosis
A small to medium-sized non-sauropodan sauropodomorph (Tables S1–S4) diagnosed by the following unique combination of characters (autapomorphies indicated by an asterisk): ascending ramus of the maxilla excavated by triangular antorbital fossa; axial intercentrum wider and completely fused to the axis; distinct ventral keel on middle cervical centra; slender shaft of the humerus with a flat and low humeral head; a large and robust ungual of manual digit I with length, proximal height and width three times those of the ungual of manual digit III *; gracile metacarpal V with proximal end as wide as distal end*.
Description
The skull is somewhat damaged caused by lateral crushing. The antorbital fenestra is sub-triangular, and the orbit is large and circular. The anteroposterior length of the antorbital fenestra (including the antorbital fossa) is less than that of the orbit based on the better-preserved right side of the skull. The anterior margin of the infratemporal fenestra is located behind the orbit. The supratemporal fenestra is sub-ovoid, and its long axis is oriented anteroposteriorly (Fig. 2).
In lateral view, the main body of the premaxilla is rectangular and longer anteroposteriorly than dorsoventrally high. No subnarial foramen can be identified, probably due to erosion. A small neurovascular foramen is present on the lateral surface of the right premaxilla behind the first premaxillary tooth.
The maxilla is triradiate and composed of an anterior process, an ascending process, and a posteriorly directed main process (Fig. 2A, B). The anterior process is anteroposteriorly longer than dorsoventrally high. The ascending process is dorsoposteriorly oriented and sets an angle of 80° with the main process of the maxilla. The posteroventral corner of the ascending process of the left maxilla is excavated by the antorbital fossa, which forms the anteromedial wall of the antorbital fenestra. This antorbital fossa is almost sub-triangular in outline, although it is less extensive than that of Plateosaurus, Unaysaurus, and Coloradisaurus19,20,21,22,23. The main process is longer and dorsoventrally lower than the anterior process. The lateral surfaces of both maxillae are damaged so that the presence of the neurovascular foramina cannot be confirmed.
Only the right nasal and prefrontal are partially preserved (Fig. 2C, D). The nasal forms a small part of the dorsal margin of the antorbital fenestra. The ventrolateral process of the nasal contacts the ascending process of the maxilla and reaches almost to the halfway of the ascending process. The ventral process of the prefrontal contacts the dorsal end of the lacrimal dorsally, and the anterior process contacts the nasal. In dorsal view, the prefrontal appears to be widened mediolaterally.
The lacrimal is inverted L-shaped in lateral view. It contributes to the posterodorsal border of the antorbital fenestra and the anteroventral border of the orbit. The length of the anterior process of the lacrimal is less than half the height of its ventral process. A web of bone spanning junction between the anterior and ventral process is present and obscures the posterodorsal corner of the antorbital fossa.
The anterior and ventral processes of the postorbital form the dorsal and the posterior edges of the orbit, respectively. The dorsal margin of the postorbital is slightly concave. The jugal consists of three processes, including the anterior, dorsal, and posterior processes. In right lateral view, the anterior process terminates posteriorly to the posteroventral corner of the antorbital fossa and makes a small contribution to the antorbital fenestra. The dorsal process sutures with the posterior margin of the ventral process of the postorbital. The posterior process, although broken at its distal end, contact the anterior process of the quadratojugal. The dorsal process of the quadratojugal contacts the ventral process of the squamosal. The angle between the dorsal and the anterior processes of the quadratojugal is less than 90°.
The symphyseal end of the dentary is dorsoventrally weakly expanded, with the ventral margin curved ventrally at the anterior tip, similar to that of Plateosaurus20. In contrast, Unaysaurus and Mussaurus possess a dentary with a more notably expanded symphyseal end22,24. The presence of the neurovascular foramina is difficult to ascertain due to abrasion of the lateral surface. The external mandibular fenestra is large and approximately 0.12 times the length of the mandible. The surangular extends anteriorly to contact the dentary and forms most of the dorsal and posterior borders of the external mandibular fenestra. The angular contacts the surangular dorsally and dentary anteriorly. It contributes to the posteroventral border of the external mandibular fenestra. The retroarticular process is finger-shaped, projecting posteriorly to the posterior margin of the skull.
The dentition of Wudingloong is consistent with that of other non-sauropodan sauropodomorphs. There are four premaxillary teeth and at least 15 maxillary teeth. The premaxillary teeth are longer and broader than the rest of the teeth. At least five maxillary teeth have a coarsely serrated crown. The serrations are present on the mesial and distal carinae of the tooth, resembling that of most non-sauropodan sauropodomorphs but differing from Yunnanosaurus in which the teeth lack the serrations5. The dentary teeth are all concealed by the maxillary teeth.
The atlas-axis complex and eight postaxial cervical vertebrae are preserved. Therefore, Wudingloong has ten cervical vertebrae including the atlas. All the cervical vertebrae are amphicoelous and lateromedially compressed at their midlength. The atlantal elements consist of a right neural arch, an odontoid, and an atlantal intercentrum. The axial intercentrum is slightly anteroposteriorly shorter and transversely narrower than the atlantal intercentrum. The axial intercentrum is completely fused to the axis and slightly mediolaterally wider than the anterior surface of the centrum of the axis (Fig. 1B). A complete fusion between the axial intercentrum and axis is rare among non-sauropodan sauropodomorphs, although this condition is also observable in Buriolestes25. The axis is approximately 0.7 times the length of the third cervical centrum, resembling the condition in most non-sauropodan sauropodomorphs, but in contrast with Adeopapposaurus in which the axis is short, and this ratio is 0.526. The posterior margin of the axial postzygapophyses extends only marginally posteriorly to the posterior margin of the axial centrum.
The third to fifth cervical centra are relatively elongate, with their anteroposterior length approximately 3.5 times the dorsoventral height of the anterior surface. This anterior cervical vertebral elongation ratio of Wudingloong is slightly higher than that of Lufengosaurus (3.3) and Xingxiulong (3.0) but contrasts with the more elongated cervical centra present in other massospondylid taxa including Adeopapposaurus (5.0), Coloradisaurus (4.5), Leyesaurus (4.9), and massopodans such as Yunnanosaurus (3.7), and Jingshanosaurus (3.9)26,27,28. As in other early sauropodomorphs, a keel is present on the ventral surface of all the cervical centra. The ventral keel of the sixth and seventh cervical vertebrae, which extend along the entire length of the centrum, is prominent, blade-like, and dorsoventrally taller and shaper than those of other non-sauropodan sauropodomorphs (e.g. Plateosaurus, Lufengosaurus, Adeopapposaurus, Massospondylus, and Xingxiulong)23,26,29. The parapophysis is placed at the anteroventral region of the lateral surface of the centrum. Cervical lamination is poorly developed. Only the posterior centrodiapophyseal and prezygodiapophyseal laminae are present in the posterior cervical vertebrae. All the cervical vertebrae lack the pneumatic foramina or fossae30,31. The epipophysis is well developed in all the cervical vertebrae. The posterior end of the epipophysis of the anterior cervical vertebrae has a free pointed tip rather than joining the postzygapophysis.
The anterior seven dorsal vertebrae are preserved. The dorsal centra are slightly amphicoelous. The blade-like ventral keel is well-developed in the first and second dorsal centra and diminishes although still present in the third dorsal centrum. It is approximately 0.5 mm in height in the first dorsal vertebra. The diapophyseal laminae of the dorsal vertebrae are well-developed. The anterior and posterior centrodiapophyseal, and postzygodiapophyseal laminae are present in all preserved dorsal vertebrae. The prezygodiapophyseal lamina is developed in the third to eighth dorsal vertebrae. Pneumatic foramina or fossae are absent in all the dorsal vertebrae30,31. The laterally expanded table is developed at the dorsal end of the anterior dorsal neural spines (Fig. 1E), as in Xingxiulong8.
The coracoid has a pronounced coracoid tubercle present at the lateral surface ventral to the glenoid. The coracoid foramen is sub-circular, deep, and developed dorsally to the coracoid tubercle. The suture between the scapula and coracoid is clear. The scapula is relatively gracile. The minimum width of the scapular blade is about 0.16 times the total length of the scapula (Fig. 1F), as also occurs in Lufengosaurus, Adeopapposaurus, Coloradisaurus, and Plateosaurus23,26,28,32 but differing from the robust scapula present in Xingxiulong and Jingshanosaurus. The dorsal end of the scapula is highly asymmetric with a posterodorsal corner, resembling that of most non-eusauropod sauropodomorphs, but unlike the almost symmetric dorsal end of the scapula in Lufengosaurus and Massospondylus29. No pneumatic foramina or fossae are present in the coracoid and scapula.
The humerus is a gracile element. In anterior or posterior view, the humeral head is low and only slightly convex proximally (Fig. 1G), similar to that of Unaysaurus and Plateosaurus22,23. However, the humeral head of other non-sauropodan sauropodomorphs, such as Lufengosaurus, Massospondylus, Adeopapposaurus, Yunnanosaurus, and Jingshanosaurus26,29, is more developed and domed. In anterior view, the lateral margin of the deltopectoral crest is almost straight, as opposed to the strongly sinuous deltopectoral crest present in Lufengosaurus, Coloradisaurus, and Plateosauravus28,33. The minimum width of the humeral shaft is 0.12 times its total length, similar to the condition present in Riojasaurus22, but in contrasts to the more robust elements of Lufengosaurus (0.14), Yunnanosaurus (0.15), Jingshanosaurus (0.19) and Xingxiulong (0.16). The ulna exhibits a triradiate proximal end with a very shallow radial fossa, as in other non-sauropodan sauropodomorphs (Fig. 1H). The humerus is about 1.6 times the length of the ulna. The radius is slender, and its proximal and distal ends are both expanded. It is about 0.61 times the length of the humerus, resembling Plateosaurus, Unaysaurus, and Adeopapposaurus in which this ratio is above 0.622,23,26, but this ratio is higher than that of most massospondylids and massopodans, e.g. Lufengosaurus, Jingshanosaurus, Yizhousaurus, and Mussaurus7,34.
The manual elements are represented by the distal ends of metacarpals II and III, metacarpals IV and V, digits I–III and V, and the proximal end of a non-terminal phalanx of the digit IV (Fig. 1I). The proximal articular surface of metacarpal IV is triangular in profile. Metacarpal V is relatively gracile and longer than it is wide at the proximal end. It has an hourglass-shaped outline, with its proximal end almost as wide transversely as the distal end, whereas other non-sauropodan sauropodomorphs (e.g. Plateosaurus, Sarahsaurus, Lufengosaurus, Yunnanosaurus, Adeopapposaurus, and Mussaurus) have a metacarpal V with a wider proximal end than the distal end23,26,32,34,35,36. The proximal articular surface of metacarpal V is slightly convex, and the distal surface is flat.
The phalangeal formula of the first three manual digits is 2:3:4. The first phalanx of digit I is the largest manual phalanx. Its distal end is twisted medially approximately 60° with respect to the proximal end. The distal condyles are asymmetrical, with the lateral condyle being larger and extending more distally, similar to that of Sefapanosaurus37, but differing from the nearly symmetrical distal condyles generally present in other non-sauropodan sauropodomorphs, e.g. Thecodontosaurus, Plateosaurus, Lufengosaurus, Yunnanosaurus, Adeopapposaurus, Massospondylus, and Mussaurus23,26,29,32,34,38. The medial collateral ligament pit is large and deep. The ungual of digit I is much larger and more robust than other unguals. Its length is almost 2.9 times the length of the ungual of digit III. Furthermore, the height and width of the proximal end of the first ungual is 3.2 times those of the third ungual, respectively. In other non-sauropodan sauropodomorphs, e.g. Plateosaurus, Lufengosaurus, Adeopapposaurus, Massospondylus, Sarahsaurus, Jingshanosaurus, Yizhousaurus, and Irisosaurus, the length of the first ungual is relatively shorter or proximally narrower when compared with the third ungual7,23,26,35,39,40. The proximal phalanx of digit V is subconical with a strongly convex distal end.
Discussion
The strict consensus tree shows a relatively good degree of resolution, although containing polytomies at the base of Sauropodomorpha, and recovers Wudingloong as an earlier diverging member of Sauropodomorpha than other taxa from China. Furthermore, Wudingloong is resolved as the sister taxon to the clade consisting of Plateosauravus and Ruehleia, which represents an early-diverging lineage of Massopoda (Fig. 3; Fig. S2). The phylogenetic positions of other Chinese sauropodomorphs, including Lufengosaurus, Yunnanosaurus, Jingshanosaurus, Xingxiulong, Yizhousaurus, Irisosaurus, and Qianlong are consistent with other recent analyses11,22,36,39,41,42.

Phylogenetic relationships of Wudingloong wui gen. et sp. nov. (LFGT-YW002). Abbreviated time-calibrated strict consensus tree depicting phylogenetic position of Wudingloong wui and the reconstructions of the manus of Plateosaurus21,39, Wudingloong, Lufengosaurus, and Jingshanosaurus for comparisons along the lineage. Scale bars equal 5 cm.
Wudingloong is clearly distinguished from other non-sauropodiform sauropodomorphs by several features. First, Wudingloong differs from the earlier-diverging taxa, e.g. Plateosauridae20,23, by having an antorbital fenestra (including the antorbital fossa) anteroposteriorly shorter than the orbit, fewer number of premaxillary and maxillary teeth, a transversely wider distal end of the humerus, and widened, shorter phalanges of the manual digits (Fig. 3). Second, in contrast to the taxa of Massospondylidae, especially Lufengosaurus also from China, Wudingloong possesses a triangular antorbital fossa, a more posteriorly located anterior margin of the infratemporal fenestra, a wider axial intercentrum, a slender humerus with a flat, low humeral head, and a straight anterolateral margin of the deltopectoral crest. Third, compared with the late members more closely related to the sauropodiforms, e.g. Yunnanosaurus, Jingshanosaurus, and Xingxiulong, Wudingloong has smooth enamel surfaces of the teeth, a more gracile scapula, a low humeral head, a high radius/humerus ratio, and a longer manus.
The antorbital fossa on the ascending process of the maxilla is normally crescentic with strongly concave posterior margins in non-sauropodan sauropodomorphs, e.g. Lufengosaurus, Jingshanosaurus, Massospondylus, and Adeopapposaurus4,9,26,43. The triangular-shaped antorbital fossa with a nearly straight posterior margin present in Wudingloong is similar to although less expansive than that of Unaysaurus, Plateosaurus, Coloradisaurus, and later-diverging taxa such as Aardonyx and Melanorosaurus NMQR 331422, suggesting a homoplastic distribution of this feature in early sauropodomorphs. However, as in Unaysaurus and Plateosaurus, the anterior rim of the antorbital fossa of Wudingloong is sharp and deep impressed, whereas in Coloradisaurus and NMQR 331421,44, this rim is rounded or just has a change in slope and only weakly impressed. Another noticeable similarity between Wudingloong, Unaysaurus, and Plateosaurus is the slender humerus with a flat and low humeral head, and a relatively long manus, although the manus of Wudingloong is slightly shorter than those of Unaysaurus and Plateosaurus. In contrast, the humerus of massospondylids and later-diverging massopodans, e.g. Jingshanosaurus, Yunnanosaurus, Antetonitrus, and Yizhousaurus7,45, is more robust and has proximally domed humeral head. Meanwhile, in sauropodiforms, the radius/humerus ratio is low, and the manus is shorter, with the phalanges being more widened. This condition may indicate that the flexible forelimb with the functions of grasping or defense were replaced slowly by the less flexible forelimb with locomotor function along the evolution of the sauropodomorphs46, supporting quadrupedality beginning from the massopodans more closely related to sauropodiforms as suggested by McPhee et al.47. Furthermore, the relatively enlarged and robust first ungual of Wudingloong implies that this taxon has a strong capability of grasping or defense48,49 and is a constant biped given that the enlarged ungual would have hampered the quadrupedal stance46. It also should be noted that the hourglass-shaped metacarpal V is uncommon for early sauropodomorphs in which the proximal end of metacarpal V is normally transversely wider than the distal end. And in some sauropodiforms, e.g. Melanorosaurus QR331450 and early sauropods, e.g. Shunosaurus51, the proximal end is narrower than the distal end. Therefore, this feature is tentatively autapomorphic for Wudingloong.
The absence of postcranial skeletal pneumaticity (PSP) in Wudingloong adds to the growing evidence that early massopodan sauropodomorphs exhibited considerable variability regarding the presence and distribution of pneumatic features. The unaysaurid Macrocollum from the Late Triassic demonstrates that invasive PSP, including pneumatic foramina and protocamerae, was present in the posterior cervical and anterior dorsal vertebrae, but absent in more anterior cervical and posterior dorsal vertebrae31. This patchy distribution of PSP in Macrocollum and other non-sauropod sauropodomorphs (e.g. Plateosaurus, Eucnemesaurus, Antenonitrus, and Aardonyx), which also shows restricted and inconsistent pneumaticity, indicates that the evolution of skeletal pneumatization before the diversification of Jurassic eusauropods was not cladistically consistent23,30,31,52. The apneumatic condition observed in Wudingloong suggests that the acquisition of extensive PSP was not a universal feature among early Massopoda and supports the hypothesis that the invasive air sac system evolved in a mosaic fashion, with repeated losses or reductions in some lineages before becoming widespread and consolidated in later Eusauropoda.
To date in East Asia, the Early Jurassic dinosaurs were only reported in southwestern China, and only one Late Triassic dinosaur, Isanosaurus, was reported from the Nam Phong Formation of Thailand53. However, the Nam Phong Formation was dated as Late Jurassic afterwards54. Wudingloong was discovered from the Lower Jurassic Yubacun Formation of Wuding, which is constrained from Hettangian to early Sinemurian18. The upper unit of the Lufeng Formation, which has also produced a large number of non-sauropodan sauropodomorphs, is then probably constrained from Sinemurian to Pliensbachian. Other Early Jurassic dinosaur bone beds from China, e.g. the Zhenzhuchong Member of the Ziliujing Formation of Pingba district of Guizhou Province11, are all constrained to Sinemurian or a younger age. Therefore, Wuding dinosaur fauna site represents so far the lowest dinosaur bone bed in China as well as in East Asia.
Based on the phylogenetic analysis and stratigraphic horizon, Wudingloong represents the earliest known occurrence of the Early Jurassic sauropodomorphs in East Asia. Wudingloong is placed as an earlier-diverging sauropodomorph than Massospondylidae and Sauropodiformes, which adds new information to the sauropodomorph assemblage of southwestern China. Therefore, the Early Jurassic southwestern China sauropodomorph assemblage is now probably represented by four unique associations of taxa, including the relatively small and gracile Wudingloong, the medium-sized massospondylid Lufengosaurus (and possibly Xixiposaurus)1,10,41, the medium-large-bodied and possibly quadrupedal massopodans, e.g. Jingshanosaurus, Xingxiulong, and Qianlong2,3,6,11,47, and the early-diverging sauropodiforms, e.g. Yizhousaurus and Irisosaurus7,39, which is similar to the Late Triassic-Early Jurassic early sauropodomorph assemblages that have a wider time range, including the Elliot Formation of South Africa55, the South America56,57, and the Zimbabwean sauropodomorph assemblages58.
The close phylogenetic relationship between Wudingloong and Plateosauravus from the Late Triassic Lower Elliot Formation of South Africa55,59 and Ruehleia from the Late Triassic of Germany32 indicates that the dispersal of early sauropodomorphs in East Asia either occurred at least in Rhaetian of the Late Triassic or just happened around the Triassic-Jurassic boundary. Further discoveries of more specimens and other analyses are necessary to test this hypothesis. Nonetheless, the discovery of Wudingloong raises the question about the association between the distribution of the East Asian non-sauropodan sauropodomorphs and the Triassic-Jurassic extinction event.
Methods
Phylogenetic analysis
In order to assess the phylogenetic position of LFGT YW002, we added it into an updated version of the data matrix of McPhee et al.22, with the addition of 19 characters from recent studies28,34,36,37,60,61,62 and one new character proposed in this study. 17 additional operational taxonomic units (OTUs) were added, including Arcusaurus, Bagualosaurus, Jaklapallisaurus, Pampadromaeus, Nambalia, Plateosaurus erlenbergensis (AMNH FARB 6810), Musankwa, Pradhania, Macrocollum, Ngwevu, Qianlong, Irisosaurus, Yizhousaurus, Kholumolumo, Sefapanosaurus, Meroktenus, and Ingentia7,11,20,37,39,58,62,63,64,65,66,67,68,69,70,71. Plateosaurus engelhardti was replaced by Plateosaurus trossingensis, which was appointed as the new type species of Plateosaurus by ICZN72,73. Scorings for the cranium of Jingshanosaurus was modified based on Zhang et al.9. The new data matrix consists of 79 OTUs and 398 characters (see Supplementary Dataset). The phylogenetic analysis was performed in TNT version 1.574 with the most parsimonious trees (MPTs) recovered via a Traditional Search of 1000 replicates of Wagner trees followed by tree bisection reconnection (TBR) swapping algorithm, saving 10 trees per replication. Characters were equally weighted, and 44 characters were treated as ordered. The analysis resulted in 168 MPTs with a length of 1647 steps (CI = 0.286, RI = 0.658). The MPTs were calibrated against geological time using the R packages Paleotree75 and Strap76. Time scale is based on the International Chronostratigraphic Chart (ICS) v. 2022/02; geological timescales are derived from Pol et al.41, Apaldetti et al.42, McPhee et al.55, and our own estimate.
Data availability
Data is provided within the manuscript or supplementary information files.
References
Young, C. C. A complete osteology of Lufengosaurus huenei Young (gen. et sp. nov.) from Lufeng, Yunnan, China. Palaeontol. Sin. 7, 1–53 (1941).
Young, C. C. Yunnanosaurus huangi Young (gen. et sp. nov.), a new Prosauropoda from the Red Beds at Lufeng, Yunnan. Bull. Geol. Soc. China. 22, 63–104 (1942).
Zhang, Y. & Yang, Z. L. A new complete osteology of Prosauropoda in the Lufeng Basin, Yunnan, China: Jingshanosaurus, 100 (Yunnan Publishing House of Science and Technology, 1995).
Barrett, P. M., Upchurch, P. & Wang, X. L. Cranial osteology of Lufengosaurus huenei Young (Dinosauria: Prosauropoda) from the Lower Jurassic of Yunnan, People’s Republic of China. J. Vertebr. Paleontol. 25, 806–822 (2005).
Barrett, P. M., Upchurch, P., Zhou, X. D. & Wang, X. L. The skull of Yunnanosaurus huangi Young, 1942 (Dinosauria: Prosauropoda) from the Lower Lufeng Formation (Lower Jurassic) of Yunnan, China. Zool. J. Linn. Soc. 150, 319–341 (2007).
Wang, Y.-M., You, H.-L. & Wang, T. A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China. Sci. Rep. 7, 41881. https://doi.org/10.1038/srep41881 (2017).
Zhang, Q.-N., You, H.-L., Wang, T. & Chatterjee, S. A new sauropodiform dinosaur with a ‘sauropodan’ skull from the Lower Jurassic Lufeng Formation of Yunnan Province, China. Sci. Rep. 8, 13464. https://doi.org/10.1038/s41598-018-31874-9 (2018).
Wang, Y.-M., Wang, T., Yang, Z.-W. & You, H.-L. Cranium and vertebral column of Xingxiulong chengi (Dinosauria: Sauropodomorpha) from the Early Jurassic of China. Anat. Rec. 303, 772–789. https://doi.org/10.1002/ar.24305 (2020).
Zhang, Q.-N., Wang, T., Yang, Z.-W. & You, H.-L. Redescription of the cranium of Jingshanosaurus xinwaensis (Dinosauria: Sauropodomorpha) from the Lower Jurassic Lufeng Formation of Yunnan Province, China. Anat. Rec. 303, 759–771. https://doi.org/10.1002/ar.24113 (2020).
Wang, Y.-M., Zhao, Q. & You, H.-L. Reassessment of ‘Gyposaurus’ sinensis Young, 1941 (Dinosauria: Sauropodomorpha) from the Early Jurassic Lufeng Basin, Yunnan Province, China. Zool. J. Linn. Soc. zlae032. https://doi.org/10.1093/zoolinnean/zlae032 (2024).
Han, F. L. et al. Exceptional Early Jurassic fossils with leathery eggs shed light on dinosaur reproductive biology. Natl. Sci. Rev. 11, nwad258. https://doi.org/10.1093/nsr/nwad258 (2024).
Pang, Q. Q. et al. Establishment of the Yubacun Formation and the Lower Boundary of the Continental Jurassic in Central Yunnan. Geol. Rev. 48, 1–8 (2002).
Brochu, C. A. Closure of neurocentral sutures during crocodilian ontogeny: implications for maturity assessment in fossil archosaurs. J. Vertebr. Paleontol. 16, 49–62. https://doi.org/10.1080/02724634.1996.10011283 (1996).
Chen, J. et al. Triassic–Jurassic environmental instability on the subtropical eastern Tethyan margin linked to the dinosaur occurrence. PREPRINT (Version 1) Available Res. Square. https://doi.org/10.21203/rs.3.rs-6720557/v1 (2025).
Burchfiel, B. C. & Chen, Z. L. Tectonics of the Southeastern Tibetan Plateau and its Adjacent Foreland, 210 (Geological Society of America, 2012).
Fang, X. S. et al. The Jurassic red bed in the Central Yunnan of China, 111 (Geological Publishing House, 2008).
Fang, X. S. et al. in Proceedings of the Third National Stratigraphical Congress of China 208–214 (Geological Publishing House, 2000).
Huang, B. et al. Magnetostratigraphy of the Jurassic in Lufeng, central Yunnan. Geol. Bull. China. 24, 322–328 (2005).
Leal, L. A., Azevedo, S. A. K., Kellner, A. W. A. & Da Rosa, Á. A. S. A new early dinosaur (Sauropodomorpha) from the Caturrita Formation (Late Triassic), Paran Basin, Brazil. Zootaxa. 690, 1–24. https://doi.org/10.11646/zootaxa.690.1.1 (2004).
Prieto-Márquez, A. & Norell, M. A. Redescription of a nearly complete skull of Plateosaurus (Dinosauria: Sauropodomorpha) from the Late Triassic of Trossingen (Germany). Am Mus. Novit 3727 (2011).
Apaldetti, C., Martinez, R. N., Pol, D. & Souter, T. Redescription of the skull of Coloradisaurus brevis (Dinosauria, Sauropodomorpha) from the Late Triassic Los Colorados Formation of the Ischigualasto-Villa Union Basin, northwestern Argentina. J. Vertebr. Paleontol. 34, 1113–1132. https://doi.org/10.1080/02724634.2014.859147 (2014).
McPhee, B. W., Bittencourt, J. S., Langer, M. C., Apaldetti, C. & Da Rosa, Á. A. S. Reassessment of Unaysaurus tolentinoi (Dinosauria: Sauropodomorpha) from the Late Triassic (early Norian) of Brazil, with a consideration of the evidence for monophyly within non-sauropodan sauropodomorphs. J. Syst. Palaeontol. 18, 259–293. https://doi.org/10.1080/14772019.2019.1602856 (2020).
Schaeffer, J. Osteological redescription of the holotype of Plateosaurus trossingensis (Dinosauria: Sauropodomorpha) from the Upper Triassic of SW Germany and its phylogenetic implications. J. Syst. Palaeontol. 22, 2335387. https://doi.org/10.1080/14772019.2024.2335387 (2024).
Pol, D. & Powell, J. E. Skull anatomy of Mussaurus patagonicus (Dinosauria: Sauropodomorpha) from the Late Triassic of Patagonia. Hist. Biol. 19, 125–144. https://doi.org/10.1080/08912960601140085 (2007).
Müller, R. T. et al. Early evolution of sauropodomorphs: anatomy and phylogenetic relationships of a remarkably well-preserved dinosaur from the Upper Triassic of southern Brazil. Zool. J. Linn. Soc. 184, 1187–1248. https://doi.org/10.1093/zoolinnean/zly009 (2018).
Martínez, R. N. Adeopapposaurus mognai, gen. et sp. nov. (Dinosauria: Sauropodomorpha), with comments on adaptations of basal Sauropodomorpha. J. Vertebr. Paleontol. 29, 142–164. https://doi.org/10.1671/039.029.0102 (2009).
Apaldetti, C., Martinez, R. N., Alcober, O. A. & Pol, D. A new basal sauropodomorph (Dinosauria: Saurischia) from Quebrada del Barro Formation (Marayes-El Carrizal Basin), northwestern Argentina. PLoS One. 6, e26964. https://doi.org/10.1371/journal.pone.0026964 (2011).
Apaldetti, C., Pol, D. & Yates, A. M. The postcranial anatomy of Coloradisaurus brevis (Dinosauria: Sauropodomorpha) from the Late Triassic of Argentina and its phylogenetic implications. Palaeontology. 56, 277–301. https://doi.org/10.1111/j.1475-4983.2012.01198.x (2013).
Barrett, P. M., Chapelle, K. E. J., Staunton, C. A., Botha, J. & Choiniere, J. N. Postcranial osteology of the neotype specimen of Massospondylus carinatus Owen, 1854 (Dinosauria: Sauropodomorpha) from the upper Elliot formation of South Africa. Palaeontol. Afr. 53, 114–178 (2019).
Yates, A. M., Wedel, M. J. & Bonnan, M. F. The early evolution of postcranial skeletal pneumaticity in sauropodomorph dinosaurs. Acta Palaeontol. Pol. 57, 85–100 (2012).
Aureliano, T. et al. The origin of an invasive air sac system in sauropodomorph dinosaurs. Anat. Rec. 307, 1084–1092. https://doi.org/10.1002/ar.25209 (2024).
Galton, P. The prosauropod dinosaur Plateosaurus MEYER, 1837 (Saurischia: Sauropodomorpha; Upper Triassic). II. Notes on the referred species. Revue De Paleobiologie. 20, 435–502 (2001).
van Heerden, J. The morphology and taxonomy of Euskelosaurus (Reptilia: Saurischia; Late Triassic) from South Africa. Navors Nas Mus. (Bloemfontein). 4, 21–84 (1979).
Otero, A. & Pol, D. Postcranial anatomy and phylogenetic relationships of Mussaurus patagonicus (Dinosauria, Sauropodomorpha). J. Vertebr. Paleontol. 33, 1138–1168 (2013).
Marsh, A. D. & Rowe, T. B. Anatomy and systematics of the sauropodomorph Sarahsaurus aurifontanalis from the Early Jurassic Kayenta Formation. PLoS One. 13, e0204007. https://doi.org/10.1371/journal.pone.0204007 (2018).
Otero, A. & Pol, D. Ontogenetic changes in the postcranial skeleton of Mussaurus patagonicus (Dinosauria, Sauropodomorpha) and their impact on the phylogenetic relationships of early sauropodomorphs. J. Syst. Palaeontol. 19, 1467–1516. https://doi.org/10.1080/14772019.2022.2039311 (2021).
Otero, A., Krupandan, E., Pol, D., Chinsamy, A. & Choiniere, J. A new basal sauropodiform from South Africa and the phylogenetic relationships of basal sauropodomorphs. Zool. J. Linn. Soc. 174, 589–634 (2015).
Ballell, A., Rayfield, E. J. & Benton, M. J. Osteological redescription of the Late Triassic sauropodomorph dinosaur Thecodontosaurus antiquus based on new material from Tytherington, southwestern England. J. Vertebr. Paleontol. 40, e1770774. https://doi.org/10.1080/02724634.2020.1770774 (2020).
Peyre de Fabrègues, C. et al. A new species of early-diverging sauropodiformes from the Lower Jurassic Fengjiahe Formation of Yunnan Province, China. Sci. Rep. 10, 10961. https://doi.org/10.1038/s41598-020-67754-4 (2020).
Cooper, M. R. The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance. Occas Pap. Natl. Mus. Rhod. B Nat. Sci. 6, 690–840 (1981).
Pol, D. et al. Earliest evidence of herd-living and age segregation amongst dinosaurs. Sci. Rep. 11, 20023. https://doi.org/10.1038/s41598-021-99176-1 (2021).
Apaldetti, C., Pol, D., Ezcurra, M. D. & Martínez, R. N. Sauropodomorph evolution across the Triassic–Jurassic boundary: body size, locomotion, and their influence on morphological disparity. Sci. Rep. 11, 22534. https://doi.org/10.1038/s41598-021-01120-w (2021).
Sues, H.-D., Reisz, R. R., Hinic, S. & Raath, M. A. On the skull of Massospondylus carinatus Owen, 1854 (Dinosauria: Sauropodomorpha) from the Elliot and Clarens Formations (Lower Jurassic) of South Africa. Ann. Carnegie Mus. 73, 239–257 (2004).
Yates, A. M. The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria). Spec. Pap. Palaeontol. 77, 9–55 (2007).
McPhee, B. W., Yates, A. M., Choiniere, J. N. & Abdala, F. The complete anatomy and phylogenetic relationships of Antetonitrus ingenipes (Sauropodiformes, Dinosauria): implications for the origins of Sauropoda. Zool. J. Linn. Soc. 171, 151–205. https://doi.org/10.1111/zoj.12127 (2014).
Otero, A. & Peyre de Fabrègues, C. in South American Sauropodomorph Dinosaurs: Record, Diversity and Evolution (eds. Otero, A., Carballido, J. L. & Pol, D.) 51–92 (Springer International Publishing, 2022).
McPhee, B. W., Benson, R. B. J., Botha-Brink, J., Bordy, E. M. & Choiniere, J. N. A giant dinosaur from the earliest Jurassic of South Africa and the transition to quadrupedality in early sauropodomorphs. Curr. Biol. 28, 1–9. https://doi.org/10.1016/j.cub.2018.07.063 (2018).
Galton, P. & Upchurch, P. in the Dinosauria (eds Weishampel, D. B., Dodson, P. & Osmólska, H.) 232–258 (University of California Press, 2004).
Otero, A., Allen, V., Pol, D. & Hutchinson, J. R. Forelimb muscle and joint actions in Archosauria: insights from Crocodylus johnstoni (Pseudosuchia) and Mussaurus patagonicus (Sauropodomorpha). PeerJ. 5, e3976. https://doi.org/10.7717/peerj.3976 (2017).
Bonnan, M. F. & Yates, A. M. A new description of the forelimb of the basal sauropodomorph Melanorosaurus: implications for the evolution of pronation, manus shape and quadrupedalism in sauropod dinosaurs. Spec. Pap. Palaeontol. 77, 157–168 (2007).
Zhang, Y. Shunosaurus lii. The Middle Jurassic Dinosaur Fauna from Dashanpu, Zigong, Sichuan: Sauropod Dinosaurs, vol. 3, 89 (Sichuan Publishing House of Science and Technology, 1988).
Regalado Fernández, O. Variability of vertebral laminae in eight specimens of Plateosaurus (Saurischia, Sauropodomorpha). Revue De Paleobiologie. 43, 85–100. https://doi.org/10.581/zenodo.10732345 (2024).
Buffetaut, E. et al. The earliest known sauropod dinosaur. Nature. 407, 72–74 (2000).
Racey, A. & Goodall, J. G. S. Palynology and stratigraphy of the Mesozoic Khorat Group red bed sequences from Thailand. Geol. Soc. Lond. Spec. Publ. 315, 69–83. https://doi.org/10.1144/SP315.6 (2009).
McPhee, B. W., Bordy, E. M., Sciscio, L. & Choiniere, J. N. The sauropodomorph biostratigraphy of the Elliot Formation of southern Africa: Tracking the evolution of sauropodomorpha across the Triassic–Jurassic boundary. Acta Palaeontol. Pol. 62, 441–465. https://doi.org/10.4202/app.00377.2017 (2017).
Pol, D., Otero, A., Apaldetti, C. & Martínez, R. N. Triassic sauropodomorph dinosaurs from South America: The origin and diversification of dinosaur dominated herbivorous faunas. J. S. Am. Earth Sci. 107, 103145. https://doi.org/10.1016/j.jsames.2020.103145 (2021).
Otero, A., Carballido, J. L. & Pol, D. South American Sauropodomorph Dinosaurs: Record, Diversity and Evolution, 582 (Springer International Publishing, 2022).
Barrett, P., Chapelle, K. E. J., Sciscio, L., Broderick, T. J. & Zondo, M. A new Late Triassic sauropodomorph dinosaur from the Mid-Zambezi Basin, Zimbabwe. Acta Palaeontol. Pol. 69, 227–241. https://doi.org/10.4202/app.01100.2023 (2024).
Haughton, S. H. The fauna and stratigraphy of the Stormberg Series. Ann. S. Afr. Mus. 12, 323–497 (1924).
Cerda, I. A. et al. Novel insight into the origin of the growth dynamics of sauropod dinosaurs. PLoS One. 12, e0179707. https://doi.org/10.1371/journal.pone.0179707 (2017).
Bronzati, M. & Rauhut, O. W. M. Braincase redescription of Efraasia minor Huene, 1908 (Dinosauria: Sauropodomorpha) from the Late Triassic of Germany, with comments on the evolution of the sauropodomorph braincase. Zool. J. Linn. Soc. 182, 173–224. https://doi.org/10.1093/zoolinnean/zlx029 (2018).
Pretto, F. A., Langer, M. C. & Schultz, C. L. A new dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Brazil provides insights on the evolution of sauropodomorph body plan. Zool. J. Linn. Soc. 185, 388–416. https://doi.org/10.1093/zoolinnean/zly028 (2019).
Kutty, T., Chatterjee, S., Galton, P. M. & Upchurch, P. Basal sauropodomorphs (Dinosauria: Saurischia) from the Lower Jurassic of India: their anatomy and relationships. J. Paleontol. 81, 1218–1240 (2007).
Novas, F. E., Ezcurra, M. D., Chatterjee, S. & Kutty, T. S. New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of Central India. Earth Env Sci. T R so. 101, 333–349. https://doi.org/10.1017/S1755691011020093 (2011).
Yates, A. M., Bonnan, M. F. & Neveling, J. A new basal sauropodomorph dinosaur from the Early Jurassic of South Africa. J. Vertebr. Paleontol. 31, 610–625. https://doi.org/10.1080/02724634.2011.560626 (2011).
Cabreira, S. F. et al. A unique Late Triassic dinosauromorph assemblage reveals dinosaur ancestral anatomy and diet. Curr. Biol. 26, 1–6. https://doi.org/10.1016/j.cub.2016.09.040 (2016).
Apaldetti, C., Martínez, R. N., Cerda, I. A., Pol, D. & Alcober, O. An early trend towards gigantism in Triassic sauropodomorph dinosaurs. Nat. Ecol. Evol. 2, 1227–1232. https://doi.org/10.1038/s41559-018-0599-y (2018).
Müller, R. T., Langer, M. C. & Dias-da-Silva, S. An exceptionally preserved association of complete dinosaur skeletons reveals the oldest long-necked sauropodomorphs. Biol. Lett. 14, 20180633. https://doi.org/10.1098/rsbl.2018.0633 (2018).
Chapelle, K. E. J., Barrett, P. M., Botha, J. & Choiniere, J. N. Ngwevu intloko: a new early sauropodomorph dinosaur from the Lower Jurassic Elliot Formation of South Africa and comments on cranial ontogeny in Massospondylus carinatus. PeerJ. 7, e7240. https://doi.org/10.7717/peerj.7240 (2019).
Peyre de Fabrègues, C. & Allain, R. New material and revision of Melanorosaurus thabanensis, a basal sauropodomorph from the Upper Triassic of Lesotho. PeerJ 4, e1639. https://doi.org/10.7717/peerj.1639 (2016).
Peyre de Fabrègues, C. & Allain, R. Kholumolumo ellenbergerorum, gen. et sp. nov., a new early sauropodomorph from the lower Elliot Formation (Upper Triassic) of Maphutseng, Lesotho. J. Vertebr. Paleontol. 39, e1732996. https://doi.org/10.1080/02724634.2019.1732996 (2019).
Fraas, E. Die neuesten Dinosaurierfunde in der schwäbischen Trias. Naturwissenschaften. 1, 1097–1100. https://doi.org/10.1007/BF01493265 (1913).
ICZN. Opinion 2435 (Case 3560)—Plateosaurus Meyer, 1837 (Dinosauria, Sauropodomorpha): new type species designated. Bull. Zoological Nomenclature. 76, 144–145. https://doi.org/10.21805/bzn.v76.a042 (2019).
Goloboff, P. A. & Catalano, S. A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics. 32, 221–238. https://doi.org/10.1111/cla.12160 (2016).
Bapst, D. W. paleotree: an R package for paleontological and phylogenetic analyses of evolution. Methods Ecol. Evol. 3, 803–807. https://doi.org/10.1111/j.2041-210X.2012.00223.x (2012).
Bell, M. A. & Lloyd, G. T. strap: an R package for plotting phylogenies against stratigraphy and assessing their stratigraphic congruence. Palaeontology. 58, 379–389. https://doi.org/10.1111/pala.12142 (2015).
Acknowledgements
We thank Shu-Yong Yue, Shu-Kai Yue, Shu-Wei Yue, Guo-Peng Li, Liu-Rong Zhu, Lin Luo, and Wen-Qing Feng for the fieldwork and specimen preparation, and Wen-Qing Feng, Wei Gao, and Xiang-Yuan Chen for helping with the photography. We are grateful to Mario Bronzati and Tito Aureliano for their thorough and constructive reviews that greatly improved the manuscript. We also thank the Willi Henning Society for the freely available TNT software. This research was supported by the National Natural Science Foundation of China (42002014, 42288201, and 42372030), the Beijing Natural Science Foundation (5224037), the State Key Laboratory of Palaeobiology and Stratigraphy of the Nanjing Institute of Geology and Palaeontology (213126), the Strategic Priority Research Programme of the Chinese Academy of Sciences (XDB26000000), and Yunnan Revitalization Talent Support Program (202305AB350006).
Author information
Authors and Affiliations
Contributions
Y-M.W., Q-N.Z., Z.F., X.X., T.W., and H-L.Y. designed the research. Y-M.W. and Y-C.W. prepared the figures. All authors wrote and reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, YM., Zhang, QN., Wang, YC. et al. A new Early Jurassic dinosaur represents the earliest-diverging and oldest sauropodomorph of East Asia. Sci Rep 15, 26749 (2025). https://doi.org/10.1038/s41598-025-12185-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-12185-2
