
Abstract
Biotic and abiotic factors influence species habitat selection across space and time. Predator habitat selection is often studied in relation to their primary prey, however, how predators shift their space use in response to secondary prey and the corresponding ecological consequences have received less attention. We used four years (2018, 2020, 2021, 2022) of wolf (Canis lupus) GPS data to examine how wolf habitat selection relates to active American beaver (Castor canadensis) lodge locations. We hypothesized that wolf selection of beaver lodges would reflect seasonal beaver behavior, intensifying through the ice-free season as beaver vulnerability increases as they spend more time away from their lodges and the vulnerability and availability of moose (Alces alces) decreases. Using a generalized linear mixed model and a mixed generalized additive model in a Bayesian framework, we analyzed how wolf habitat selection changed, especially near active beaver lodges. We used 834 unique active beaver lodges in our analysis, 395 [0.74 lodges /km2] in 2018, 386 [0.72 lodges /km2] in 2020, 344 [0.64 lodges /km2] in 2021, and 168 [0.31 lodges /km2] in 2022, a total decline of 57% since wolf restoration. We collected 18,932 wolf GPS locations (N2018 = 1200, N2020 = 7224, N2021 = 3690, N2021 = 6818) from 23 wolves (10 females and 13 males). Our results suggest that wolves shifted their habitat selection to increase encounters with beavers, supporting previous work demonstrating the importance of beavers to the wolf diet during snow-free periods. This shift supports wolf prey-switching behavior to beavers when their primary prey, moose, are more difficult and riskier to kill.
Similar content being viewed by others
Introduction
Biotic and abiotic factors influence species habitat selection across spatial and temporal scales. Predators have direct (lethal) and indirect (behavioral) effects on their prey and changes in predator habitat selection may have unexpected, cascading ecological consequences1. Often, predator habitat selection is studied in relation to their primary prey2 and how they respond to secondary prey sources receives relatively less attention.
Foragers must balance the benefits of obtaining food with the costs of locating and consuming resources. Specifically, predators often select habitats that increase encounter rates and the capturability of their primary prey3. Predators may opportunistically consume secondary prey or alter their foraging strategy to take advantage of shifts in prey availability or vulnerability. For example, gray wolves (Canis lupus) in Riding Mountain National Park, Manitoba, Canada selected habitat to increase encounters with their primary prey, moose (Alces alces), but not their secondary prey, elk (Cervus canadensis)3. In contrast, Valeix et al.4 observed that lions avoided human settlements when wild prey was abundant but shifted their habitat selection to prey on cattle when wild ungulates were scarce. Predators shifting their activity to take advantage of secondary prey can increase predation rates on these and other alternative prey5.
American beavers (Castor candadiensis) and wolves are ideal species to understand predator habitat selection in relation to secondary prey. In most systems ungulates comprise > 70% of the wolf diet6, however beavers are often an important secondary prey in summer7. Beavers likely represent a low risk and abundant prey for wolves in summer, when ungulates are difficult to capture due to increased nutritional status. Beavers are central-place foragers and at increased risk of predation when they move on land8. For wolves, beaver availability and vulnerability likely changes throughout the year. Beavers are unavailable to wolves in winter, as they are typically under ice or within their lodges. Beavers become available to wolves in spring, when beavers leave the security of their lodges in search of forage or to disperse. Beaver vulnerability to wolf predation likely increases during fall as the frequency and duration of terrestrial foraging increases9.
Wolves appear to kill beavers by targeting them near dams, on shore, or along trails near their lodges10. Thus, we assumed that if wolves shift their habitat selection to target beavers, their selection of areas within 100 m of active beaver lodges would increase accordingly. Here, we use four years of concurrent wolf GPS data and beaver lodge monitoring to demonstrate how seasonal wolf habitat selection relates to the location of beaver lodges. We predict that wolf selection of beaver lodges will reflect the seasonal behavior of beavers and increase throughout the ice-free season.
Methods
Study area
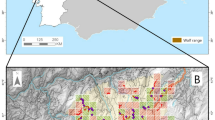
Isle Royale National Park (IRNP) is a 535-km2 archipelago in Lake Superior, Michigan, USA (Fig. 1). Over 99% of IRNP is federally designated wilderness and contains mixed boreal forests characteristic of the transition between temperate and boreal zones11. Dry mixed conifer forests of balsam fir (Abies balsamea) and white spruce (Picea glauca) dominate areas near Lake Superior and in northeastern portions of IRNP12. Upland forests are dominated by deciduous species including sugar maple (Acer saccharum) and yellow birch (Betula alleghaniensis). Mesic mixed conifer forests of white cedar (Thuja occidentalis) and black spruce (Picea mariana) are common in lowlands and swamps13. Wet meadows and marshes are common in low laying areas, near lakes, and around beaver impoundments. Other habitat types in IRNP included rocky shorelines, cliff faces, and anthropogenic structures13. Mean annual precipitation in IRNP 1991–2020 was 71 cm with 187 cm of snow; mean temperature in July was 18 C and − 13 C in January14.

Location of Isle Royale National Park within the great lakes region, USA (top panel), distribution of active beaver lodges 2018–2022 within IRNP [excluding Passage Island] (middle panel), and example of 100 m buffer around an active beaver lodge, wolf GPS locations, random points, and surrounding habitat types (bottom panel). Maps were created using Esri ArcGIS Pro, https://pro.arcgis.com/en/pro-app15 and Adobe Illustrator, https://www.adobe.com/products/illustrator16. Top panel service layer credits: Esri Canada, TomTom, Garmin, FAO, NOAA, USGS, EPA, NPS, USFWS, NRCan, Parks Canada. Middle and bottom panels: National Park Service 200017, modified by A. Sovie.
Beaver monitoring
The National Park Service conducted beaver lodge activity surveys in October 2018, 2020, 2021, and 2022. The island was divided into eight sectors and a pilot and observer flew in a float equipped fixed wing aircraft systematically across the entirety of each sector. The surveys involved flying NW-SE transects across the west end of Isle Royale, where beaver habitat is relatively rare. At the east end of Isle Royale where beaver habitat is more common, the plane flew along all drainages. The plane circled potential lodges as needed to determine whether a lodge was active18. The observer used various sign, including a food cache, freshly peeled sticks, newly cut trees or shrubs, canals, fresh mud on a lodge, or a recently repaired dam to determine if a lodge was active19,20,21. The observer located active lodges on a paper map or using a GPS. This method is highly effective at detecting active beaver lodges in IRNP with detection probability > 0.818. We excluded beaver lodges on Passage Island (where wolves do not occur) and lodges within 200 m of an active wolf den or rendezvous site. We created 100-m buffers around active beaver lodges to reflect areas where wolves hunt beavers [lodges, dams, and trails]. We considered this buffer a unique habitat type (Fig. 1). Habitat composition with the buffers included 28% dry mixed conifer forest, 20% deciduous forest, 20% wet meadows/marshes, 22% mesic mixed conifer forests, 9% water, and 2% other (Sup 1).
Wolf monitoring
Wolves were captured from four mainland source populations and translocated to IRNP September 2018–September 2019. We also captured wolves in IRNP in April 2022. Wolves were captured with foothold traps or darting and net-gunning from helicopters. Each wolf was fit with a GPS telemetry collar (Vertex Plus; VECTRONIC Aerospace, Berlin, Germany, or TGW-4470-4; Telonics, Mesa, Arizona, USA) programmed to attempt a location every 4–5 h and emit a mortality signal when stationary for 24 h. We censored the first week of locations from animals to allow them to acclimate to the radio collar. We excluded all wolf locations within 200 m of known denning sites to avoid bias from disproportionately greater use of these areas during pup rearing (NPS unpublished data). We also excluded GPS locations within Lake Superior. All wolf capture, handling, and tagging procedures were approved by the NPS Institutional Animal Care and Use Committee, supervised by NPS veterinarians, followed American Society of Mammologists guidelines22 for trapping and handling mammals, and adhered to all federal, tribal and state regulations.
Data analysis
To understand wolf spatio-temporal response to active beaver lodges, we compared use of beaver lodges to available habitat. We paired each wolf location with a random point constrained to fall within the land boundary of IRNP excluding Passage Island. We identified the habitat type of each point using the merged beaver lodge buffer and IRNP vegetation inventory map (Fig. 1)13.
We characterized wolf habitat selection using a logistic generalized linear mixed effect model (GLMM) and a logistic generalized additive mixed effect model (GAMM). GLMMs are a common class of models used to understand wildlife habit associations in diverse ecosystems23. GLMMs are extensions of commonly used linear models, are typically easier to interpret than GAMMs, and facilitate comparisons across variables. However, GLMMs assume the relationship between predictors and responses are related through a defined functional form which temporal relationships rarely follow. In contrast, GAMMs allow the response variable to vary following smoothed functions of the predictors and have been used to quantify temporal shifts in habitat use24,25,26. To leverage the advantages of both modeling approaches we first broadly characterized wolf habitat selection across seasons using a GLMM and then investigated how wolf use of selected habitats shifts at a finer temporal resolution using a GAMM.
For GLMM analysis we divided the year into four seasons (i.e. bins) based on beaver and wolf ecology. We defined spring as the average date of ice break up on interior lakes in IRNP (15 April)27 to the end of moose partition (June 14)28. We defined summer as 15 June–14 August to correspond with peak moose calf availability29. We defined fall as 15 August to the average date of complete ice cover on interior lakes in IRNP (14 November). We defined winter as the average dates of ice cover on interior lakes [14 November–14 April, 27], when beavers are primarily within their lodges or under ice and unavailable to wolves. We modeled the interaction between season and habitat use using a logit link. To account for variation between years and individual wolf behavior we used Wolf ID and year as random intercepts and Wolf ID as a random effect on habitat type slopes. We used the “brms” package30 in program R version 4.4.031 to fit the model. We used uninformative priors and ran 3 chains for 10,000 iterations with a 1000 iteration burn in. We considered the models converged if Rhat was < 1.232. We estimated the conditional log odds and a 95% credible interval (CI) by averaging 1000 posterior draws. We used the modeled log odds and 95% CI to interpret wolf habitat use, where log odds of > 0 indicate selection and < 0 avoidance. We considered estimates with 95% CIs that do not cross 0 informative.
While the buffer surrounding beaver lodges contained diverse habitats (Sup 1), lodges were disproportionally located in or adjacent to wet meadows and marshes, potentially conflating the selection of these resources. To further compare the use of these habitats with finer temporal resolution, we fit a GAMM with a logit link and fixed effect interaction between habitat type and day of year as a temporal gradient. To account for variation between years and individual wolf behavior we used Wolf ID and year as random intercepts and Wolf ID as a random effect on habitat type slopes. We used the R packaged mgcv33, to fit the model in a frequentist framework. To identify time periods with differing use we used the “plot_diff” function in the R package “itsadug”34. Plot_diff, calculates the pairwise differences between smooth terms for different levels of a factor and estimates a 95% credible interval, we considered 95% CI’s that did not contain 0 to indicate differential use of habitats.
Results
We used 834 unique active beaver lodges in our analysis, 395 [0.74 lodges/km2] in 2018, 386 [0.72 lodges/km2] in 2020, 344 [0.64 lodges/km2] in 2021, and 168 [0.31 lodges/km2] in 2022, a total decline of 57% since wolf restoration. We collected 18,932 wolf GPS locations (Table 1, Sup 1) (N2018 = 1200, N2020 = 7224, N2021 = 3690, N2021 = 6818) from 23 wolves (10 females and 13 males, Sup 1).
GLMM
The random intercept for year had a standard deviation of 0.04 (95% CI 0.0–0.18), indicating low between-year variability (Sup 3). The model included random slopes for habitat use across individual wolves. The standard deviations of the intercept was 0.13 (95% CI 0.09–0.18), and random slopes ranged from 0.25 to 0.82 (Sup 3) indicating moderate to high individual variance depending on the habitat. Wolves selected for beaver sites in summer, fall, and winter; with greatest absolute use in fall (Fig. 2, Sup 2). Wolves selected for wet meadows/marshes in all seasons, with greatest use in summer. Wolves selected for dry mixed conifer woodlands during winter and avoided them in summer. Wolves avoided deciduous forests in all seasons (Fig. 2). Wolves appeared to avoid mesic mixed conifer in winter, and selected for it in spring through fall, although only the 95% CI for summer did not overlap 0 (Fig. 2).
GAMM
Wolf selection of wet meadows and marshes increased January–July, from avoidance in January to selection in May (Fig. 3). In contrast wolves increasingly avoided beaver lodges during January–April and transitioned to selecting for lodges in June (Fig. 3). Wolves avoided beaver lodges compared to wet meadows/marshes in early spring and selected for lodges more strongly in late fall (Fig. 4).

Generalized linear mixed model mean posterior predicted log odds and 95% credible intervals of annual habitat selection by gray wolves, Isle Royale National Park, Michigan, USA, 2018–2022. Horizontal line represents log odds of 0; points above this line indicate selection while points below indicate avoidance. DMC dry mixed conifer, D deciduous, BL beaver lodges, M marsh, MMC mesic mixed conifer.

Generalized additive mixed model smoothed splines and 95% credible intervals of annual gray wolf selection of beaver lodges (orange) and wet meadows/marshes (blue) in Isle Royale National Park, Michigan, USA, 2018–2022. Horizontal line represents log odds of 0; above this line indicates selection while below indicates avoidance.

Estimated difference in the log-odds of habitat use and 95% CI between lodges and marshes/wet meadows over time, based on a logistic generalized additive model, Isle Royale National Park, Michigan, USA, 2018–2022. Horizontal line is 0, no difference, values above this line indicate higher wolf use of lodges compared to marshes.
Discussion
Predator habitat selection can have complex and cascading effects on ecosystems. Predators can reduce prey density in frequented areas through direct predation or fear-based avoidance35. Understanding how predators respond to seasonal shifts in secondary prey availability can inform our understanding of ecological systems and predation patterns. We found that wolves selected active beaver lodges in every season but spring, with greatest selection in October. Our results support our hypothesis that wolves in IRNP shift their habitat selection in response to spatial and temporal variation in prey availability and vulnerability29.
The availability and vulnerability of moose adults and calves, the primary prey of wolves, and beaver varies seasonally in IRNP and is reflected in wolf habitat selection. On IRNP adult moose vulnerability appears to peak in May and reach a nadir in September29. Moose are vulnerable to predation in early spring due to winter nutritional stress and winter tick (Dermacentor albipictus) burdens36. Vulnerable moose likely succumb to starvation or predation in spring whereas remaining moose regain fat stores after green up37,38. Similarly, moose calves are highly vulnerable to predation shortly after birth (mid-June) but can evade wolves by August. This seasonal pulse in prey vulnerability and availability is reflected in wolf habitat selection, as wolf selection of habitats associated with moose, mesic mixed conifer and wet meadows/marshes39, increased from spring to summer and declined in fall (Fig. 1). In contrast our GAMM results suggest wolves avoided beaver lodges in early spring (April–May) but selected for them during July–November, indicating a shift in foraging strategy (Fig. 2). Beavers are more vulnerable during fall through increased terrestrial foraging9. The combination of reduction in moose vulnerability and increased beaver availability from late summer to fall likely influence wolf foraging decisions.
Contrary to our expectations, our GLMM analysis indicated wolves selected for beaver lodges in winter despite our assumption that beavers are largely unavailable this time of year. However, our GAMM results clarified this finding. Using day as our temporal resolution, we found wolf selection of beaver lodges peaked in October and declined throughout winter, remaining above log odds of 0 (i.e. selection) into December. Ice cover is likely discontinuous earlier in the season and beavers may spend time above ice when air temperatures are above freezing40. During our study, maximum daytime air temperatures were regularly above freezing into December41, facilitating beaver activity past the date of ice-cover. Our GAMM results support our hypothesis that wolves target beavers as they forage in preparation for winter, then focus on other habitats when beavers spend more time in their lodges.
Our results support previous research that beavers are an important component of the non-winter wolf diet7,29. Beavers comprised a large portion of the summer wolf diet on Isle Royale before restoration7 and are an important prey item for the introduced population29. We found wolf selection of beaver lodges increased through the ice-free season, contrasting with results from an analysis of the summer diet of Isle Royale wolves conducted in 2018–2019, where the occurrence of beaver in wolf scats was high (78% of scats contained beaver hair) and consistent during May–October29. This discrepancy may be a result of methodological differences and the power to detect change between studies, or that wolf selection of beaver lodges does not necessarily result in increased predation rates.
We acknowledge several limitations of our study. Wolves may have used the areas around beaver lodges as travel corridors, water sources, or to hunt alternative prey including moose, muskrats (Ondatra zibethicus), or fish. Also, due to our 5-hour GPS relocation interval we were unable to use GPS clusters42 to determine whether a wolf was transitorily using an area near a beaver lodge or actively foraging. When investigating wolf GPS clusters in IRNP we found few beaver kills (30% of clusters, NPS unpublished data) compared to their prominence in the wolf diet (78% of scat contents29. Gable et al.,43 found that one-third of beaver kills may be missed when using a 2-hour fix rate instead of a 20-min fix rate. However, given the strength of the patterns we observed, we suggest that the ephemeral selection of areas around beaver lodges reflects wolves hunting temporally variable prey. Finally, our estimate of wolf selection for beaver lodges may be biased low, as a portion of each buffered area likely included open water that wolves are unlikely to use, while we did not restrict random points from falling into open water within the beaver lodge buffer.
Understanding how recovering predators select habitat and respond to seasonal shifts in prey availability is important for predicting and managing the ecosystem level effects of restoration. We demonstrated that wolves in IRNP shifted their habitat selection to areas near beaver lodges temporally. This behavior could have cascading effects as increased wolf presence near lodges in late summer and fall can influence beaver foraging decisions44, and subsequently forest structure. Beavers are ecosystem engineers and the impoundments they create can influence water and nutrient flows at landscape scales45,46; wolf predation can alter the duration and distribution of these impoundments9. Further, increased wolf activity in wet meadows/marshes may alter the behavior of secondary carnivores, like red foxes (Vulpes vulpes)47, reducing predation risk for smaller prey species. Similarly, increased wolf presence in areas surrounding beaver lodges may alter moose behavior in these areas44,48,49, reducing their foraging pressure on aquatic vegetation. Ecological restoration involving large carnivores could be improved by considering the temporal and spatial availability of alternative prey into project planning and implementation.
Data availability
Wildlife location data are considered sensitive by the National Park Service and are not publicly available. Non-georeferenced data is available in Dryad. Contact Mark Romanski, Isle Royale National Park Natural Resources Division Lead, with data access questions or requests.
References
Kuijper, D. P., De Kleine, C., Churski, M., Van Hooft, P. & Bubnicki, J. Jędrzejewska B. Landscape of fear in europe: wolves affect Spatial patterns of ungulate browsing in Białowieża primeval forest, Poland. Ecography 36, 1263–1275 (2013).
Kittle, A. M. et al. Landscape-level Wolf space use is correlated with prey abundance, ease of mobility, and the distribution of prey habitat. Ecosphere 8 (4) (2017).
Zabihi-Seissan, S. & Prokopenko, C. M. Vander wal, E. Wolf Spatial behavior promotes encounters and kills of abundant prey. Oecologia 200 (1), 11–22 (2022).
Valeix, M., Hemson, G., Loveridge, A. J., Mills, G. & Macdonald, D. W. Behavioural adjustments of a large carnivore to access secondary prey in a human-dominated landscape. J. Appl. Ecol. 49 (1), 73–81 (2012).
Latham, A. D. M., Latham, M. C., Knopff, K. H., Hebblewhite, M. & Boutin, S. Wolves, white-tailed deer, and beaver: implications of seasonal prey switching for woodland caribou declines. Ecography 36 (12), 1276–1290 (2013).
Mech, L. D., Boitani, L. & Wolves Behavior, Ecology, and Conservation. The University of Chicago Press, Chicago, Illinois, and London, United Kingdom, ISBN 0-226-51696-2 (2003).
Gable, T. D., Windels, S. K., Romanski, M. C. & Rosell, F. The forgotten prey of an iconic predator: a review of interactions between grey wolves Canis lupus and beavers Castor spp. Mammal Rev. 48 (2), 123–138 (2018).
McClintic, L. F., Taylor, J. D., Jones, J. C., Singleton, R. D. & Wang, G. Effects of Spatiotemporal resource heterogeneity on home range size of American beaver. J. Zool. 293 (2), 134–141 (2014).
Gable, T. D., Johnson-Bice, S. M., Homkes, A. T., Fieberg, J. & Bump, J. K. Wolves alter the trajectory of forests by shaping the central place foraging behaviour of an ecosystem engineer. Proc. Royal Soc. B, 290, (2023).
Gable, T. D., Windels, S. K., Bruggink, J. G. & Homkes, A. T. Where and how wolves (Canis lupus) kill beavers (Castor canadensis). PLoS One, 11 (12), (2016).
Sanders, S. & Grochowski, J. The forests of Isle royale national park: can we preserve this pristine wilderness in the face of climate change? Nat. Areas J. 33 (1), 66–77 (2013).
Slavick, A. D. & Janke, R. A. The vascular flora of Isle Royale National park. Mich. Botanist (USA), 26 (3), (1987).
The Nature Conservancy. USGS-NPS Vegetation Mapping Program Classification of the Vegetation of Isle Royal National Park September 9, 1999 (Midwest Regional Office. Minneapolis, 1999).
Environment Canada. Canadian climate normals 1991–2020, Thunder Bay A & Ontario Available from http://www.cmc.ec.gc.ca/climate/normals/ONTT004.HTM [accessed 13 February 2023].
Esri Inc. ArcGIS Pro version 3.3.1. Redlands, CA. (2024). https://pro.arcgis.com/en/pro-app
Adobe Inc. Adobe Illustrator. San Jose, CA. (2024). https://www.adobe.com/products/illustrator
National Park Service. Geospatial data for the vegetation mapping inventory project of isle royale national park. (2000). https://irma.nps.gov/DataStore/Reference/Profile/2233314
Romanski, M. C. Estimates of abundance and predation—the population ecology of beaver in isle royale national park. MSc thesis, Michigan Technological University, Houghton, Michigan, USA, (2010).
Hay, K. G. Beaver census methods in the Rocky mountain region. J. Wildl. Manag. 22, 395–402 (1958).
Payne, N. F. Accuracy of aerial censusing for beaver colonies in Newfoundland. J. Wildl. Manag. 45, 1014–1016 (1981).
Swenson, J. E., Knapp, S. J., Martin, P. R. & Hinz, T. C. Reliability of aerial cache surveys to monitor beaver population trends on prairie rivers in Montana. J. Wildl. Manag., 697–703, (1983).
Sikes, R. S. Guidelines of the American society of mammalogists for the use of wild mammals in research and education. J. Mammal. 97, 663–688 (2016). Animal Care and Use Committee of the American Society of Mammalogists.
Thiele, J. & Markussen, B. Potential of GLMM in modelling invasive spread. CABI Revi., 1–10, (2012).
Bollen, M., Casaer, J., Neyens, T. & Beenaerts, N. When and where? Day-night alterations in wild Boar space use captured by a generalized additive mixed model. PeerJ 12, 7390 (2024).
Polansky, L. & Robbins, M. M. Generalized additive mixed models for disentangling long-term trends, local anomalies, and seasonality in fruit tree phenology. Ecol. Evol. 3 (9), 3141–3151 (2013).
Fewster, R. M., Buckland, S. T., Siriwardena, G. M., Baillie, S. R. & Wilson, J. D. Analysis of population trends for farmland birds using generalized additive models. Ecology 81 (7), 1970–1984 (2000).
(2024). Retrieved from https://dataspace.copernicus.eu/explore-data 5 processed by Copernicus Sentinel Data & Sovie, A. March, (2024).
McGraw, A. M., Moen, R. & Schrage, M. Characteristics of post-parturition areas of moose in Northeast Minnesota. Alces 47, 113–124 (2011).
Sovie, A. R. et al. Temporal variation in translocated Isle Royale Wolf diet. Ecology Evolution, 13 (3), (2023).
Bürkner, P. C. & brms An R package for bayesian multilevel models using Stan. J. Stat. Softw. 80, 1–28 (2017).
R Core Team. R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. URL: (2024). https://www.R-project.org.
Gelman, A., Carlin, J. B., Stern, H. S. & Rubin, D. B. Bayesian Data Analysis, Third Edition (texts in Statistical science) (Chapman and Hall/CRC, 2013).
Wood, S. N. Fast stable restricted maximum likelihood and marginal likelihood Estimation of semiparametric generalized linear models. J. Royal Stat. Soc. (B). 73 (1), 3–36 (2011).
van Rij, J., Wieling, M., Baayen, R. & van Rijn, H. itsadug: Interpreting Time Series and Autocorrelated Data Using GAMMs. R package version 2.4.1., (2022).
Ripple, W. J. & Beschta, R. L. Wolves and the ecology of fear: can predation risk structure ecosystems? BioScience 54 (8), 755–766 (2004).
Lenarz, M. S., Nelson, M. E., Schrage, M. W. & Edwards, A. J. Temperature mediated moose survival in Northeastern Minnesota. J. Wildl. Manag. 73 (4), 503–510 (2009).
Tischler, K. B., Severud, W. J., Peterson, R. O. & Bump, J. K. Aquatic macrophytes are seasonally important dietary resources for moose. Diversity 11, 209 (2019).
Parker, G. R. Status Report on the Eastern Moose (Alces Alces Americana Clinton) in Mainland Nova Scotia (Environment Canada, 2003).
Allen, A. W., Jordan, P. A. & Terrell, J. W. Habitat Suitability Index Models: Moose, Lake Superior RegionVol. 82 (US Department of the Interior, Fish and Wildlife Service, Research and Development, 1987).
Lancia, R. A., Dodge, W. E. & Larson, J. S. Winter activity patterns of two radio-marked beaver colonies. J. Mammal. 63 (4), 598–606 (1982).
Environment Canada, Data, H., Bay, T. & Ontario, A. Available from https://climate.weather.gc.ca/historical_data/search_historic_data_e.html [accessed 13 February 2023].
Svoboda, N. J., Belant, J. L., Beyer, D. E., Duquette, J. F. & Martin, J. A. Identifying bobcat Lynx Rufus kill sites using a global positioning system. Wildl. Biology. 19 (1), 78–86 (2013).
Gable, T. C., Johnson-Bice, S. M., Homkes, A. T. & Bump, J. K. Differential provision roles, prey size, and prey abundance shape the dynamic feeding behavior of Gray wolves. Commun. Biology. 6, 1045 (2023).
Severud, W. J., Belant, J. L., Bruggink, J. G. & Windels, S. K. Predator cues reduce American beaver use of foraging trails. Human-Wildlife Interact. 5 (2), 296–305 (2011).
Rosell, F., Bozser, O., Collen, P. & Parker, H. Ecological impact of beavers Castor fiber and Castor canadensis and their ability to modify ecosystems. Mammal Rev., 35 (3), 248–276, (2005).
Johnston, C. A. Beaver pond effects on carbon storage in soils. Geoderma 213, 371–378 (2014).
Haswell, P. M., Jones, K. A., Kusak, J. & Hayward, M. W. Fear, foraging and olfaction: How mesopredators avoid costly interactions with apex predators. Oecologia 187, 573–583 (2018).
Pusenius, J., Kukko, T., Melin, M., Laaksonen, S. & Kojola, I. Wolf predation risk and moose movement in Eastern Finland. Wildl. Biology. 4, 1–9 (2020).
Sand, H. et al. Behavioral effects of Wolf presence on moose habitat selection: Testing the landscape of fear hypothesis in an anthropogenic landscape. Oecologia 197, 101–116 (2021).
Acknowledgements
Beaver lodge monitoring was performed under NPS contract and the NPS retains full, unrestricted use thereof. The NPS Institutional Animal Care and Use Committee approved wolf translocation and monitoring protocols in IRNP. The wolf translocation and associated monitoring required collaboration among numerous governments, agencies, institutions, and non–governmental organizations. Partners included the Grand Portage Band of Lake Superior Chippewa, Michigan DNR, Minnesota DNR, Ontario Ministry of Natural Resources and Forestry, U.S. Department of Agriculture Animal and Plant Health Inspection Service–Wildlife Services, U.S. Fish and Wildlife Service, and U.S. Geological Survey. Support from universities included Colorado State University, Michigan State University, Michigan Technological University, State University of New York College of Environmental Sciences and Forestry, Trent University, and University of Minnesota. Primary funding was provided by NPS. We thank the many volunteers and technicians that contributed to field data collection.
Author information
Authors and Affiliations
Contributions
MR and JB lead and designed wolf monitoring protocols and data collection. Beaver monitoring protocols are a long-standingprogram of the NPS AS analyzed the data and wrote the main manuscript. All authors contributed to reviewing and revising the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sovie, A.R., Romanski, M.C. & Belant, J.L. Seasonal use of American beaver lodge areas by gray wolves in Isle Royale National Park. Sci Rep 15, 31437 (2025). https://doi.org/10.1038/s41598-025-14670-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-14670-0
