
Abstract
Fibroblasts display complex functions linked to distinct gene expression profiles. These profiles influence matrix production, cell communication, and tissue development and repair. Thrombospondin-2 (TSP2), produced by fibroblasts, is a potent angiogenesis inhibitor and negatively associated with tissue repair. Single-cell RNA sequencing (scRNA-seq) on WT and TSP2 KO skin fibroblasts demonstrate distinct cell heterogeneity. Specifically, we found an enrichment of Sox10 + multipotent progenitor cells, identified as Schwann precursor cells, in TSP2 KO fibroblasts, while fibrosis-related subpopulations decreased. Immunostaining of mouse back skin wounds and cells validated the increase of this Sox10 + population. Furthermore, in silico analysis suggested enhanced PDGF-β-mediated pro-survival and inhibited BMP4-mediated differentiation signaling pathways. These molecular and functional alterations likely contribute to improved healing and increased neurogenesis in TSP2 KO wounds. Overall, our findings describe the heterogeneity of dermal fibroblasts (DFs) and identify pro-regenerative features of TSP2 KO fibroblasts.
Similar content being viewed by others


Introduction
Skin wounds in mammals heal by fibrosis, a repair mechanism that involves the rapid deposition of collagen and other extracellular matrix components that contribute to scar formation1. This fibroproliferative response, while effective in preventing infection and fluid loss, often results in scar tissue formation that differs structurally and functionally from the uninjured skin. This is due to excess deposition of collagen fibrils that leads to thickening of skin with altered mechanical properties and contributes to its inability to regenerate complex tissue structures like hair follicles and sweat glands. It is appreciated that multiple cell types, including inflammatory cells and fibroblasts, play critical roles in determining healing outcomes.
Fibroblasts are major extracellular matrix (ECM) producers and perform functions critical to wound healing2. Upon injury, activated fibroblasts migrate to the wound site and proliferate during the early phase of healing. Subsequently, they differentiate into myofibroblasts that synthesize new ECM and contract the wound, thus facilitating tissue repair. Fibroblasts also secrete various signaling molecules that recruit other cell types, such as macrophages, epithelial cells, and endothelial cells, to coordinate the healing process. Control of diverse fibroblast functions, including proliferation, differentiation, ECM production, and their communication with other cells, can influence healing outcomes.
In addition to functional diversity, fibroblasts demonstrate molecular heterogeneity across and within organs, which is also critical for tissue homeostasis2,3,4,5. For example, subsets of skin and intestine fibroblasts contribute to tissue renewal via remodeling of the ECM and serving as a stem cell niche for hair follicles or intestinal crypts, respectively6,7. These subpopulations are characterized by the expression of WNT signaling components that support epithelial and hair follicle stem cell maintenance6,7,8,9. In contrast, fibroblasts in tissues with lower cellular turnover, such as the lung and kidney, exhibit fewer stem-like markers but are nonetheless vital for repair, primarily by secreting signaling molecules2. It is appreciated that tissue-specific heterogeneity is associated with distinct functions in homeostasis and following injury.
Within tissues, fibroblast subpopulations can be categorized based on their spatial distribution. For example, cells near the upper layer of skin are defined as papillary fibroblasts (Pdgfra + CD26 + Sca-), whereas cells in the reticular dermis are called reticular fibroblasts (Pdgfra + Dlk1 + Sca-)10. Papillary and reticular fibroblasts produce distinct ECM consisting of non-fibrillary collagens and proteoglycans and well-organized fibrillary collagen bundles, respectively11,12. Due to the capability to deposit collagenous ECM, reticular fibroblasts are considered more fibrotic whereas papillary fibroblasts are more pro-regenerative13. Thus, the balance between these two populations is believed to determine the outcome of skin wound healing10. Additionally, fibroblasts with specific molecular signatures can exhibit distinct behaviors in tissue repair. For example, En1 + and Cthrc1 + fibroblasts are associated with fibrosis14,15, while Wnt1 + and Lef1 + fibroblasts are linked to reduced scarring15,16. Recent single-cell RNA sequencing (scRNA-seq) efforts integrating mouse skin and wound datasets have highlighted the spatial and temporal dynamics of fibroblasts during tissue repair, revealing that scar tissues are enriched with non-lasting, non-interconverting fibroblast populations17. Additionally, fibroblasts are also categorized via morphology at single-cell level, enabling identification of distinct senescent subtypes in aging skin18. These findings underscore the importance of elucidating fibroblast identities and behaviors to better understand the failure of regenerative healing.
Thrombospondin-2 (TSP2) is an ECM matricellular glycoprotein, whose levels of expression are negatively correlated with tissue repair outcomes. For example, mice with increased or low/null TSP2 levels display delayed or accelerated wound healing, respectively19,20,21,22. Primarily, TSP2 exerts anti-angiogenetic effects via binding to CD36 and CD47 receptors on endothelial cells23,24. Additionally, TSP2 has been implicated in ECM assembly as it was shown that TSP2 deficiency disrupts collagen fibril morphology and organization25. Moreover, TSP2 KO wounds display increased levels of soluble growth factors and enzymes, such as vascular endothelial growth factor (VEGF) and matrix metalloproteinases (MMPs)26. Together, changes in ECM assembly, angiogenesis, and growth factor availability suggest that TSP2 is a critical regulator of wound healing. Supporting this, TSP2 KO wounds exhibit accelerated epithelialization, increased angiogenesis, and reduced scar formation22,26. Wounds treated with constructs derived from TSP2 KO ECM also exhibit improved healing with enhanced cell infiltration and angiogenesis, and the emergence of new nerve bundles27,28,29,30. Collectively, these findings underscore a strong connection between TSP2 and tissue regeneration.
Differences between the healing capacities of WT and TSP2 KO skin, coupled with our understanding of fibroblasts as the major source of TSP2, prompted us to evaluate these cells in an unbiased high throughput manner. We applied scRNA-seq to dermal fibroblasts (DFs) from WT and TSP2 KO mice. In silico analysis revealed a reduction of traditional ECM-producing fibroblast populations and an increase in a Sox10⁺ population resembling Schwann cell precursors in TSP2 KO fibroblasts. Gene expression and differential pathway analyses demonstrated downregulation of BMP4 signaling and upregulation of PDGF-β signaling in TSP2 KO fibroblasts. Flow cytometry and qPCR analyses of isolated DFs confirmed the expansion of Sox10⁺ populations and increased PDGF-β expression. Given their role as nerve-associated progenitors, the increased presence of Sox10⁺ cells in TSP2 KO wounds may contribute to enhanced neurogenesis and regenerative healing. Overall, our findings reveal fibroblast heterogeneity in skin and demonstrate that TSP2 deficiency drives DFs toward a pro-regenerative state.
Method and materials
Animals
C57bl/6J and TSP2 KO mice aged 12 to 14 weeks were used. Only male mice were used for sequencing. C57bl/6J mice were purchased from Jackson Laboratory. TSP2 KO mice were generated using C57bl/6J strain as described previously25. All animal study procedures were approved by the Yale Institutional Animal Care and Use Committee (IACUC). All mice were kept in a 12-hour light/dark environment and fed ordinary feed. All authors complied with ARRIVE guidelines. All methods were performed in accordance with the relevant guidelines and regulations.
Dermal fibroblasts isolation and sequencing sample preparation
Dermal fibroblasts (DFs) were isolated from 12-week-old male WT and TSP2 KO mice dorsal skin according to the established protocol. Average body weight of mice was 25 ± 1.25 g. Briefly, mice were euthanized using CO2 inhalation at a 50% volume displacement per minute and death was confirmed by lack of reflexes, heartbeat, and respiration. Dorsal skins were shaved, excised, treated with 5% antibiotic-antimycotic (Gibco), and incubated in 25 µg/ml Trypsin (Sigma) overnight at 4 °C. After separation from the epidermis and adipose tissue, the dermis was physically disrupted and then digested using collagenase IV (Worthington Biochemical) for 3 h under 37 °C. After centrifuge, cell pellet was then resuspended in 25mM glucose Dulbecco’s Modified Eagle Medium (DMEM, Gibco) with 10% (v/v) fetal bovine serum (FBS, Peak Serum Inc) and 1% (v/v) penicillin-streptomycin (P/S, Gibco) and cultured for 3–5 days to enrich the fibroblast population.
For scRNA-seq sample preparation, after the enrichment assay, DFs were lifted using trypsin (Sigma), suspended in PBS at a concentration of 1000 cells/µl, and passed through a 100 µm cell strainer (BD Bioscience). The single-cell suspension was sent to the Yale Microarray Center, where library preparation was performed using the Chromium Next GEM single-cell 3’ v3 reagent kit (10x Genomics). The libraries were then sequenced on the Illumina NovaSeq 6000 platform with a 150 bp read length.
For bulk RNA sequencing, RNA was isolated from passage 1 DFs using the RNeasy Mini Kit (Qiagen) according to the manufacturer’s protocol. The RNA concentration was determined using a Nanodrop spectrophotometer (Thermo Scientific). Approximately 2 µg of RNA per sample was sent to the Yale Genomics Core for sequencing using the Illumina TruSeq Stranded mRNA kit (Illumina).
Single-cell RNA sequencing data analysis
Sequencing data were aligned to the mouse genome (mm10) using Cell Ranger (10x Genomics). The raw unfiltered matrix was then analyzed with the Seurat R package (version 5.0.3)31. Cells with low-quality reads (nFeatures < 300, or mitochondrial gene percentage < 0.1% or > 10%) were excluded during the QC process. A total of 18,193 and 20,134 cells remained for WT and KO samples (n = 3), respectively, and these were merged and integrated using canonical correlation analysis (CCA) with the first 40 principal components based on the top 2000 variable features. Graph-based clustering was performed using by Louvain clustering via the FindNeighbors and FindClusters function in Seurat (resolution = 0.4, dims.use = 1:40), and markers for each cluster were identified using a comprehensive Wilcoxon rank-sum test within the FindAllMarkers function (min.pct = 0.5, log fold change > 0.25). 2-dimensional embedding was performed using uniform manifold approximation and projection (UMAP) via the RunUMAP function in Seurat32.
Given the uneven distribution of cell numbers across clusters, differential gene analysis between genotypes within individual clusters was performed to minimize confounding variance. Specifically, comparisons were made between individual KO and WT samples, resulting in the generation of nine gene lists. (3WT x 3KO) A scoring value was created to reflect how many times each gene appears in the nine gene lists (1–9). P-values were integrated using Fisher’s method. Genes were ranked based on Fisher p-value, average fold change (Avg Log2FC), ratio, power (Avg Log2FC × ratio), and score (frequency across datasets). Genes with a score of 6 or higher were selected for further analysis. Downstream GO pathway and network analysis were performed using the STRING database (https://string-db.org/).
For cell-cell communication analysis, cells were processed using the NICHES package33. Autocrine signaling in endothelial, epithelial, and immune cells were excluded to focus specifically on interactions involving fibroblasts. The resulting signaling archetypes were annotated based on the sending and receiving cell types, with most interactions being related to fibroblast autocrine pathways due to the deliberate enrichment for fibroblasts in our data via selective cell culture. Cell-cell signaling edges were down-sampled to mitigate sample variation due to sample size and integrated with rpca (resolution = 0.2, dims.use = 29). Differential global cell-cell signaling between genotypes was explored using the FindMarkers function.
Violin, ridge, and feature plots of specific gene expression levels were generated using the corresponding functions in Seurat. Other visualizations, such as dot plots, histograms, and heatmaps, were generated using the ggplot2 and pheatmap packages34,35. Downstream pathway analysis, including GO pathways and GSEA, was conducted using the STRING database (https://string-db.org/) and the Web-based Gene Set Analysis Toolkit (https://www.webgestalt.org).
Flow cytometry (FACS)
A total of 1,000,000 isolated DFs were fixed using 2% PFA (J.T. Baker) for 20 min and permeabilized with 0.1% Triton-X in PBS for 15 min. Cells were treated with 1% BSA in PBS for non-specific blocking and then incubated with FACS antibodies. Untreated DFs were used as a negative control, and beads stained with corresponding antibodies were used as a positive control. All cells were passed through a 100 μm cell strainer (BD Bioscience) before being loaded onto the FACSAria II (BD Bioscience), and signals were recorded for 105 events. Laser parameters were set based on the negative and positive controls. Data analysis was then performed using FlowJo software (FlowJo, LLC).
Immunofluorescence (IF) staining
For wound tissue IF staining, 4-mm thick paraffin-embedded tissue slides from WT and TSP2 KO mouse D7 wounds were processed according to standard protocols. Primary antibody was diluted in 1% BSA in PBS and applied to the tissue area. After overnight incubation at 4 °C, slides were incubated with secondary antibody solution with DAPI for 1 h at RT and mounted using VECTASHIELD® Antifade Mounting Medium (Vector Laboratories, H-1000-10). Stained tissues were then imaged under EVOS Cell Imaging System (ThermoFisher Scientific). Images were analyzed with ImageJ (Molecular Devices).
Reverse transcription and quantitative-RT-PCR
Reverse transcription was completed with the QuantiTect Reverse Transcription Kit (Qiagen). Quantitative real-time PCR (qRT-PCR) was performed with the iTaq Universal SYBR Green One-Step Kit (Bio-Rad) on a CFX96 Touch Real-Time PCR machine (Bio-Rad). Primers were ordered from the Keck Oligonucleotide Facility (Yale University) from sequences found in PrimerBank (Harvard University) (Primer sequence: Table S1). Data were normalized to Gapdh expression.
Reagents
The following primary antibodies were used: Sox10 Antibody [Alexa Fluor 700] (1:200, Novus Biologics, NBP2-59621AF700), anti-Neurofilament H (NFH) (1:200, EMD Millipore, AB1989), anti-Vimentin (1:500, EMD Millipore, AB5733), Sox10 Rabbit pAb (1:100, ABclonal, A15100).
The following secondary antibodies were used for immunofluorescence staining or flow cytometry: anti-rabbit IgG Alexa Fluor 488 (1:1000, Invitrogen, a11008), anti-chicken IgG Alexa Fluor 488 (1:1000, Abcam, ab150169), anti-rabbit IgG Alexa Fluor 555 (1:1000, Abcam, ab150078).
Statistical analyses
Error bars represent the standard deviation (SD) unless stated otherwise. An unpaired Student’s t-test (two-tailed) was performed for comparisons between two groups. For comparisons involving three or more groups, one-way analysis of variance (ANOVA) with Tukey’s post hoc test for multiple comparisons was used. Statistical methods other than these two were specified with the results. All statistical analyses were performed using GraphPad Prism 9 or in R. Non-significant comparisons are not shown in the figures by default.
Results
Single-cell RNA transcriptomics reveals distinct mesenchymal distributions within WT and TSP2 KO skin
Isolated DFs from WT or TSP2 KO back skin (male, 12-week-old, n = 3) were subjected to single-cell RNA sequencing (scRNA-seq) (Fig. 1A). After assessing sample quality based on UMI counts, gene features, and mitochondrial percentage, we obtained 20,134 cells from KO and 18,193 cells from WT samples. Using classical markers such as Col1a1, Ptprc, Epcam, and Cdh5, we identified four major cellular lineages in the skin: mesenchymal, immune, epithelial, and endothelial cells (Fig. S1). Cell proportion analysis revealed > 95% mesenchymal abundance across samples, confirming the reliability of our mesenchymal enrichment protocol (Fig. S1).
Focusing on the mesenchymal population, we identified ten distinct fibroblast subtypes, each showing differential distribution between WT and TSP2 KO samples (Fig. 1). Based on specific marker expression, these subtypes were classified as Dcn+, Rspo3+,Dkk2+, Ebf2+, Taco1+, IFN-stimulated fibroblasts (Fb), Schwann cell precursors (ScP), vascular smooth muscle cells (VSMC), and Schwann (Sc) or fibroblast (Fb) cycling cells (Top2a+). Two-dimensional UMAP projections and proportional analyses revealed a notable enrichment of the Schwann cell precursor population in TSP2 KO samples, which is highly distinct from other mesenchymal cells and is prominently marked by Sox10, Ngfr, and Erbb3. In contrast, the largest fibroblast populations in WT skin—comprising the Dcn+, Rspo3+, and Dkk2+ subsets—were markedly reduced following TSP2 depletion.
To better understand the significance of these shifts in cell proportions, we further characterized each cluster at the molecular level. Feature plots of classical fibroblast markers (Col1a1, Pdgfra, Thy1, Fbln1, Cd34, and Loxl1) showed broad expression across the major fibroblast populations (Fig. 1E), indicating a traditionally defined fibroblast profile. Similarly, markers associated with dermal fibroblast spatial localization were dispersed across the different subpopulations (Fig. S1). Gene scoring for upper and lower dermis signatures revealed no clear spatial segregation among clusters (Fig. S1), emphasizing a molecular and functional rather than spatial distinction between different populations.
Cluster marker analysis revealed that ECM-related genes, such as Adamts2, Dkk3, Lox, and Fbn1, were enriched in the Dcn + population, suggesting a primary role in ECM production. Rspo3+ fibroblasts displayed high expression of En1, Ltbp1, Wnt5a, and Fgf10, genes associated with WNT and TGF-β signaling pathways. In contrast, Dkk2+ fibroblasts exhibited a distinct signaling profile characterized by Bmpr1b, Ptgs1, Ptgs2, and Mmp3 expression. While the major fibroblast populations shared overlapping gene expression patterns, smaller subsets exhibited more discrete profiles. For example, Taco1, a mitochondrial translational activator gene, was exclusively expressed in the Taco1+ fibroblast subset, which was nearly absent in TSP2 KO samples (Fig. S1). IFN-stimulated fibroblasts expressed a combination of interferon-responsive genes (Ifit1, Ifit3, Oasl2), suggesting a putative role in immune defense during acute infection. Ebf2, a marker for brown and beige fat precursors36, was detected in two distinct populations. However, neither population expressed additional adipocyte precursor markers such as Klf4 or Pparg (Fig. S1). One subset co-expressed smooth muscle marker (Myh11, Myom1, Cnn1), consistent with a vascular smooth muscle cell (VSMC) identity, while the other, enriched for Igfbp4 and Vit, was designated as Ebf2+ fibroblasts, potentially linked to fat metabolism.
After characterizing the traditional fibroblast populations, we next analyzed Sox10+ cells to better understand the biological significance of their expansion in TSP2 KO samples. As shown in FeaturePlot, these cells expressed multiple neural crest stem cell (NCSC) markers, including Sox2, Sox10, and Ngfr. During development and tissue maintenance, NCSCs serve as the primary source of melanocytes and can also differentiate into sensory neurons and Schwann cells that constitute the peripheral nervous system. To define the identity of the Sox10+ population, we compared their molecular profiles against known lineage markers. We found strong expression of Schwann cell lineage genes such as Erbb3, S100b, and Gap43, while melanocyte-specific markers like Mitf were absent (Figs. 1F & G). Importantly, mature Schwann cell markers, including Egr2, Pou3f1, and Mbp, were not expressed, suggesting that these cells remain in a dedifferentiated, precursor-like state. Thus, we annotated this cluster as Schwann precursors (ScP).
Together, these results reveal substantial fibroblast heterogeneity in the dermis and demonstrate a shift in cell population distributions in TSP2 KO. Specifically, populations expressing canonical skin fibroblast markers (Pdgfra, Thy1, En1, Dcn, Rspo3) were diminished in TSP2 KO skin, whereas a Schwann cell precursor-like population expressing Sox10, Ngfr, and Erbb3 was expanded.

Single cell transcriptomics reveals distinct WT and TSP2 KO DF cell population distribution. (A) Diagram of dermal fibroblast isolation and sequencing process; (B) Cell cluster UMAP plot; (C) Cell type distribution in each genotype; (D) Dotplot of selected marker genes for each cell cluster; (E) Feature plots of commonly defined fibroblast markers. (F) Feature plots and (G) Violin plots of representative markers for Schwann cell development within this population.
Three major fibroblast clusters show cluster-specific differential genes and pathways regulated by TSP2 depletion
To further investigate the impact of TSP2 deficiency beyond changes in cell distribution, we performed cluster-specific differential gene expression analyses. As expected, Thbs2 (encoding TSP2) was among the top downregulated genes across all clusters in TSP2 KO samples, confirming the reliability of the dataset. Notably, Ogn (osteoglycin), a small keratan sulfate proteoglycan, was also consistently downregulated in the major fibroblast populations (Fig. 2 and Tables S2–S6). In contrast, Tnc (Tenascin C) emerged as the most strongly upregulated gene in TSP2 KO fibroblasts (Fig. 2 and Tables S2–S6). Both Ogn and Tnc are non-structural extracellular matrix (ECM) proteins known to modulate cell behavior and matrix organization37,38 Specifically, loss of Ogn has been shown to promote smooth muscle cell proliferation and migration via the VEGF/VEGFR2 pathway 38, while elevated Tnc expression enhances fibroblast differentiation—processes critical for effective tissue repair38. Thus, the downregulation of Ogn and upregulation of Tnc in TSP2 KO fibroblasts may underlie improved cell function and ECM remodeling in the absence of TSP2.
In both the Dcn+ and Rspo3+ fibroblast populations, genes essential for cell growth and mitochondrial function were downregulated in TSP2 KO samples, including Hoxc8 and Gfer. In contrast, a cohort of genes associated with extracellular matrix remodeling, oxidative stress response, and cellular signaling, including Grem1, Sod2, Tnc, Cebpb, and Snrpf, were commonly upregulated in the TSP2 KO fraction of the Dcn+, Rspo3+, and Dkk2+ clusters (Fig. 2 and S2). Among these transcripts, Grem1 (Gremlin 1) stood out as a key regulator of cell fate and tissue remodeling. As a secreted antagonist of bone morphogenetic proteins (BMPs) and an agonist of VEGFR2 signaling, Grem1 can block BMP – induced cell differentiation and promote angiogenesis39,40. Its robust upregulation with TSP2 deficiency suggests a potential activation of anti-differentiation and pro-angiogenic pathways, potentially reshaping the fibroblast landscape in TSP2 KO skin.
Cluster-specific changes were also evident. Dcn+ fibroblasts exhibited broad dysregulation of growth factor- and ECM-related genes, including members of the insulin-like growth factor binding protein family (Igfbp2), fibroblast growth factors (Fgf10), and multiple collagen isoforms (Col18a1, Col5a3, Col4a5, Col8a1, Col11a1), along with matricellular proteins like Postn and Efemp1. Gene Ontology (GO) enrichment analysis linked these changes to processes such as morphogenesis, matrix organization, and cell adhesion.
In Rspo3+ fibroblasts, we observed downregulation of Sfrp1, a WNT signaling inhibitor, suggesting possible potentiation of intercellular WNT communication in TSP2 KO skin. Interestingly, upregulation of Trp53 (encoding p53) was also detected in this population, which activates apoptotic and cell cycle arrest pathways, possibly contributing to the observed reduced abundance of Rspo3+ cells.
Although Dkk2+ fibroblasts shared many differentially expressed genes with the Dcn+ and Rspo3+ clusters, GO pathway analysis highlighted distinct enrichment in extracellular ligand–receptor interactions, signal transduction, and BMP-related pathways, aligning with the unique molecular signature of this population, for example, the expression of Bmpr1b.
Taken together, these findings demonstrate that TSP2 deficiency reprograms fibroblast gene transcription within the skin. Our observation that multiple populations showed signs of altered BMP and WNT pathway activation suggested to us that TSP2 might play a central role in regulating signaling pathways critical for fibroblast function and skin homeostasis and repair.

Three major fibroblast clusters show cluster-specific differential genes and pathways regulated by TSP2 depletion. (A) Heatmap of highly differential genes (score ≥ 6, |power| > 1), (B) GO pathway analysis, and (C) Violin plots of highly regulated genes between WT and KO within Dcn+, Rspo3+, and Dkk2 + fibroblasts.
Sox10 + population resembles Schwann Precursors with increased Pdgfb expression in KO sample
Differential gene expression analysis of the Schwann precursor population highlighted a striking upregulation of genes associated with cell survival, angiogenesis, and tissue remodeling in the TSP2 KO group. Notably, among the top upregulated genes were Pdgfb (platelet-derived growth factor subunit B), Mcam, Moxd1, Jag1, Plp1, Cdh2, Col18a1, Sdc4, Edil3, and Anxa1 (Fig. 3 and Table S2). These factors collectively promote cell proliferation, migration, and fate stabilization through both autocrine and paracrine mechanisms41,42,43,44,45,46,47,48,49. Pdgfb emerged as a central player, known for its potent roles in promoting cell survival, proliferation, and angiogenesis. Its upregulation suggests a shift towards enhanced tissue repair and vascular remodeling programs in TSP2 KO skin. Complementary pathways were also activated: Jag1, a major Notch ligand, promotes angiogenesis and cell-cell communication47; Mcam and Cdh2 facilitate cell adhesion and motility46; Edil3 is essential for vascular development48; and Anxa1 supports VEGF-mediated endothelial migration49. Consistent with these molecular signatures, Gene Ontology (GO) enrichment analysis identified “positive regulation of endothelial cell migration,” “cell-substrate adhesion,” and “epithelial cell development” as the top biological processes affected in Schwann precursors from TSP2 KO mice.
Taken together, these data reveal that the Sox10+ population within the dermal fibroblast compartment represents a Schwann cell precursor state with a pro-survival, pro-angiogenic molecular profile. Their expansion and increased expression of regenerative pathways in the absence of TSP2 suggest that this population may contribute to a more reparative, pro-regenerative environment in TSP2 KO skin.

Sox10 + population resembles Schwann Precursors with increased Pdgfb expression in TSP2 KO group. (A) Heatmap of highly differential genes (score ≥ 6, |power| > 1, gene symbols labeled with * are associated with pro-angiogenesis), (B) Violin plots, and (C) GO pathway analysis of highly regulated genes between WT and KO Schwann Precursors.
Cell-to-cell communication analysis indicates enhanced PDGF-β and reduced BMP4 signaling in TSP2 KO cells
Having measured altered mesenchymal distributions and shifted gene expression profiles in TSP2 KO skin, we turned out attention to global cell-to-cell ligand-receptor connectivity to investigate potential systems-level mechanisms. We applied NICHES33 a computational method that samples individual pairs of cells (referred to as cell-to-cell “edges”) and quantifies/characterizes their cognate ligand-receptor profile. We queried all literature-supported ligand-receptor mechanisms present within the FANTOM5 database50 translated to yield orthologous signaling mechanisms in mouse51. Then, we applied UMAP to the ligand-receptors and signaling archetypes are annotated based on their sender and receiver identities (Fig. 4 and S3 A&B). Proportional analysis indicates that signaling involving Schwann Precursors (ScP) and Schwann Cycling cells was significantly increased in KO group compared to WT (Fig. 4 and S3C).
Differential analysis showed that TSP2 deficiency causes enrichment of PDGF-β-mediated signaling and loss of BMP4-mediated signaling (Fig. 4 and S3&4). Dotplot of BMP4- and PDGF-β-mediated circuit pairs demonstrated changes in terms of both average expression and percentage expression (Fig. S3), indicating that both mean intensity and percent of positive interactions are impacted globally. PDGF-β-mediated signaling was markedly elevated in TSP2 KO cells. Circuit activation including Pdgfb-Lrp1, Pdgfb-Pdgfrb, and Pdgfb-Itgav were upregulated in KO group, with Schwann Precursors and Schwann Cycling cells emerging as the major senders (Fig. S3). Circuit plots visually confirmed both an expanded receiving population and heightened signaling intensity, evidenced by the thickened communication edges, which represents mean connectivity of profiled intercellular edges (Fig. 4). Conversely, BMP4 signaling pathways (Bmp4-Bmpr1a/Bmpr2/Acvr2/Acvr1) were predominantly enriched in WT cells (Fig. 4C and S4A), with Dcn+ and Rspo3+ populations serving as the principal producers and receivers (Fig. 4D and S4 B&C). Loss of these interactions in KO samples likely stems from the diminished abundance of traditional fibroblast clusters, as well as increased expression of BMP4 antagonists, such as Grem1, identified in earlier differential expression analyses.
Overall, these findings indicate that TSP2 deficiency rewires intercellular mesenchymal communication networks by activating PDGF-β signaling circuits and dampening BMP4-mediated circuits, concurrent with the expansion of Schwann-like precursors and the molecular signatures previously observed. Given the well-established, fundamental roles of intercellular PDGF-β and BMP circuitry in regulating cell proliferation, migration, and differentiation within tissues41,52,53, the observed shifts help to finally explain the altered regenerative and functional potential of mesenchymal cells within TSP2 KO skin.

NICHE analysis indicates enhanced PDGF-β and reduced BMP4 intercellular signaling in TSP2 KO group. (A) The distribution of WT and KO signaling edges in a 2D UMAP plot; (B) Sending cell type and receiving cell type of each edge; (C) Volcano plot of highly regulated ligand-receptor mechanisms perturbed by KO (Supplementary file 2); (D) Circuit plot of PDGF-β and BMP4 family signaling in WT and KO cells (edge thickness is proportional to mean connectivity and frequency, respectively).
Neurogenesis is enhanced in TSP2 KO skin wounds with increased Sox10 + population and Pdgfb expression
To validate the in silico observations, we first conducted flow cytometry on isolated DFs and confirmed a significant increase in the proportion of Sox10+ fibroblasts in the KO group (Fig. 5). In parallel, qPCR analysis revealed elevated Pdgfb mRNA levels in TSP2 KO DFs, consistent with enhanced pro-survival and pro-angiogenic signaling (Fig. 5). Given that Schwann Precursors are developmental progenitors of peripheral nerve structures—including Schwann cells and sensory neurons—we reasoned that neurogenesis would be a direct and relevant functional readout of their regenerative potential. Furthermore, while previous studies have investigated angiogenesis and myofibroblast formation in TSP2 KO wounds, the impact on nerve regeneration had not yet been explored. Therefore, we focused on assessing neurogenesis as an indicator of repair.
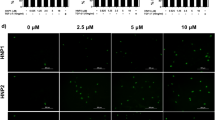
To evaluate neurogenesis, we stained day 7 (D7) back skin wounds from WT and KO mice for neurofilament heavy chain (NFH), a marker of mature nerve fibers. Quantitative analysis revealed that KO wounds exhibited a significantly greater NFH+ area relative to the total wound bed compared to WT controls (Fig. 5). Additionally, nerve bundles in KO wounds were markedly larger in diameter (Fig. 5), suggesting enhanced axonal regrowth or stabilization. Immunofluorescence staining for Sox10 further demonstrated a higher number of Sox10⁺Vimentin⁺ cells in KO wounds, present both within nerve bundles and throughout the extracellular matrix (Fig. 5 and S5), confirming the persistence and possible functional contribution of these multipotent cells during repair.
Together, these results demonstrated that TSP2 deficiency promotes an environment favorable for neurogenesis during wound healing. This highlights a previously unrecognized role for TSP2 in the control of tissue regeneration. We recently described enhanced neurogenesis in a volumetric muscle loss model in mice treated with hydrogel derived from TSP2 KO mice27. Interestingly, in a rat model of medial collateral ligament transection, addition of nerve growth factor increased nerve density and vascularity while it reduced TSP2 expression54.

TSP2 KO back skin wounds demonstrate increased neurogenesis with increased Sox10 + population and Pdgfb expression in TSP2 KO DFs. (A) Flow cytometry of Sox10 in isolated primary DFs and (B) quantification of Sox10+ (%) cells. (C) qPCR of Pdgfb in WT and TSP2 KO DFs. (D) Immunofluorescence staining of NFH and Sox10 with Vimentin in D7 WT and TSP2 KO mouse back skin wounds and quantification of (E) number of nerve bundles (NBs) in the center, (F) diameter of NBs (µm), and (G) Sox10 + area/HPF (%) across the whole wounds (dashed circles indicate nerve bundles, n = 3–4, Unpaired T-test, two-tailed, *, p < 0.05, **, p < 0.01).
Discussion
In this study, we explored the link between TSP2 and tissue regeneration by focusing on skin mesenchymal cells, a major cell type in wound repair. ScRNA-seq enabled us to investigate mesenchymal heterogeneity in great detail. We found 10 distinct mesenchymal populations. Some are clearly recognizable from the literature as the three largest fibroblast populations - Dcn+, Rspo3+, and Dkk2+, showed common fibroblast markers such as Col1a1, Pdgfa, Thy1, and Loxl1, etc55. The enrichment of extracellular matrix (ECM) genes in these clusters is consistent with the established role of fibroblasts in ECM production and dermal tissue homeostasis56. Notably, the expression of Thy1 and En1 in the major fibroblast populations supports their association with a fibrogenic lineage15,57,58,59. In contrast, other populations lacked clear in vivo precedent in the literature and were instead characterized by the expression of Taco1, Ebf2, and interferon-related genes, suggesting possible roles in mitochondrial function, metabolic regulation, and immune signaling. We also identified an interesting population expressing stem cell-associated markers including Sox10, Ngfr, and Erbb3, which may represent a Schwann precursor population with neurogenic and mesenchymal potential. Differential abundance between conditions, namely Schwann precursors being more present and fibrosis-related populations being less present in TSP2 KO, aligns with prior knowledge showing that depletion of TSP2 improves wound healing with reduced scar formation30,22.
The reduction of a major fibroblast population is accompanied by a downregulation of intercellular signaling mediated by BMP-family ligands in TSP2 KO cells. Bone morphogenetic protein 4 (BMP4), a member of the TGF-β superfamily, plays a pivotal role in cell fate determination and tissue regeneration60,61,62,63. As a key regulator of cell differentiation, BMP4 promotes the differentiation of hematopoietic, mesenchymal, and neural progenitor cells through activation of both SMAD-dependent and Ras-ERK signaling pathways61,62,64. Additionally, BMP4 has been shown to suppress stem cell signatures in the intestinal epithelium and limit the self-renewal capacity of Lgr5 + stem cells65. In human fetal lung fibroblasts, BMP4 inhibits cell proliferation and promotes myocyte differentiation60. Given its strong influence on cellular proliferation and differentiation, the downregulation of BMP4 signaling in TSP2 KO mesenchymal cells may contribute to enhanced cellular stemness.
Indeed, we observed an increased Sox10 + population in TSP2 KO skin that exhibit molecular characteristics resembling multipotent Schwann precursor cells. While in vitro experiments have demonstrated that fibroblasts can be converted into Schwann cells with specific stimulus66, the exact origin of these cells in TSP2 KO skin remains elusive. TSP2 KO ScP demonstrated elevated levels of Pdgfb, Cdh2, and Jag1 signaling, molecules which improve cell survival, enhance migration, and inhibit differentiation. Specifically, Pdgfb has been identified as a critical factor in the autocrine survival circuit of Schwann precursor cells41,42, and its enhanced expression in the TSP2 KO Sox10 + population may help to support the widely observed distribution of these cells within, specifically, TSP2 KO skin. Moreover, Schwann precursor migration depends on interactions with both cells and the ECM, as they are directed by axonal guidance. A recent study identified N-cadherin (Cdh2) as a master regulator of collective Schwann cell migration through Slit2/3-mediated contact inhibition locomotion43. In the TSP2 KO model, the increased expression of Cdh2 in ScPs, along with a more “loosened” ECM environment, likely facilitates their migration. Additionally, elevated Jag1 is associated with activation of Notch signaling which has a dual role in Schwann cell biology: while it promotes proliferation in early stages, it acts as a negative regulator of myelination during later stages of differentiation44,45. This suggests that Notch activation may contribute to the retention of a progenitor-like state in these cells. Interestingly, TSP2 has been reported to both potentiate and suppress Notch signaling through its interaction with Jag167,68, highlighting a potential regulatory mechanism that warrants further investigation. Despite the various hypotheses discussed above, the expansion of the Sox10 + population in TSP2 KO fibroblasts is discovered and validated for the first time. These cells not only exhibit molecular similarities to multipotent Schwann progenitors but are also suggested to release pro-angiogenic and regenerative ligands. This profile may enhance the regenerative potential of the fibroblast population and foster an environment conducive to tissue repair.
Although our scRNA-seq findings align with previous in vitro and in vivo observations of TSP2 knockout (KO) dermal fibroblasts, the underlying mechanistic actions require more rigorous investigation and validation. Moreover, repair is a dynamic and context-dependent process. Our current analysis is limited to unwounded skin, which constrains our ability to fully capture the roles of distinct fibroblast subpopulations during wound healing. For instance, Schwann cells have been shown to adopt a strong pro-fibrotic phenotype in keloid scar tissue compared to normal skin69,70,71,72. Thus, in a dynamic context, the distribution of cell fates and signaling activities within each subpopulation may differ significantly from what we describe here, in homeostatic conditions. To substantiate our proposed model, the spatiotemporal distribution of individual fibroblast populations throughout the wound healing process would need to be determined.
Conclusion
This study reveals that knocking out TSP2 supports pro-regenerative mesenchymal phenotypes in skin. Specifically, we observed an expansion of Sox10+ populations, which resemble Schwann precursor cells, and a reduction in fibrosis-related fibroblast populations in the TSP2 KO skin. The increased presence of Sox10⁺ cells was validated through immunostaining of both tissue and cell populations, confirming their altered distribution due to genetic knock-down. Additionally, we uncover evidence that these progenitors express pro-survival and pro-migratory molecules and may contribute to an environment that promotes tissue regeneration. We show first evidence of increased neurogenesis in TSP2 KO skin. Taken together, our findings deepen our understanding of mesenchymal cells in tissue repair, clarify the precise ways that TSP2 knock-down alters mesenchymal cell heterogeneity and intercellular communication in the skin, and illuminate new system-scale mechanisms which might help explain the well-documented enhanced tissue healing found in TSP2 KO organisms.
Data availability
All data needed to evaluate the conclusions in the paper are included in this article and its supplementary information. Datasets are deposited onto NCBI GEO (GSE283557).
References
Do, N. & Eming, S. Skin fibrosis: models and mechanisms. Curr. Res. Translational Med. 64 (4), 185–193 (2016).
Gomes, R. N., Manuel, F. & Nascimento, D. S. The bright side of fibroblasts: molecular signature and regenerative cues in major organs. NPJ Regen Med. 6 (1), 43 (2021).
Bensa, T., Tekkela, S. & Rognoni, E. Skin fibroblast functional heterogeneity in health and disease. J. Pathol. 260 (5), 609–620 (2023).
Lujano Olazaba, O., Farrow, J. & Monkkonen, T. Fibroblast heterogeneity and functions: insights from single-cell sequencing in wound healing, breast cancer, ovarian cancer and melanoma. Front. Genet. 15, 1304853 (2024).
Lynch, M. D. & Watt, F. M. Fibroblast heterogeneity: implications for human disease. J. Clin. Invest. 128 (1), 26–35 (2018).
Meran, L., Baulies, A. & Li, V. S. W. Intestinal stem cell niche: the extracellular matrix and cellular components. Stem Cells Int 2017, 7970385 (2017).
Myung, P., Andl, T. & Atit, R. The origins of skin diversity: lessons from dermal fibroblasts. Development 149(23):dev200298 (2022).
Valenta, T. et al. Wnt ligands secreted by subepithelial mesenchymal cells are essential for the survival of intestinal stem cells and gut homeostasis. Cell. Rep. 15 (5), 911–918 (2016).
Telerman, S. B. et al. Dermal Blimp1 acts downstream of epidermal TGFbeta and Wnt/beta-Catenin to regulate hair follicle formation and growth. J. Invest. Dermatol. 137 (11), 2270–2281 (2017).
Woodley, D. T. Distinct fibroblasts in the papillary and reticular dermis: implications for wound healing. Dermatol. Clin. 35 (1), 95–100 (2017).
Sorrell, J. M. & Caplan, A. I. Fibroblast heterogeneity: more than skin deep. J. Cell. Sci. 117 (Pt 5), 667–675 (2004).
Stunova, A. & Vistejnova, L. Dermal fibroblasts-A heterogeneous population with regulatory function in wound healing. Cytokine Growth Factor. Rev. 39, 137–150 (2018).
Philippeos, C. et al. Spatial and Single-Cell transcriptional profiling identifies functionally distinct human dermal fibroblast subpopulations. J. Invest. Dermatol. 138 (4), 811–825 (2018).
Tsukui, T. et al. Collagen-producing lung cell atlas identifies multiple subsets with distinct localization and relevance to fibrosis. Nat. Commun. 11 (1), 1920 (2020).
Rinkevich, Y. et al. Skin fibrosis. Identification and isolation of a dermal lineage with intrinsic fibrogenic potential. Science 348 (6232), aaa2151 (2015).
Phan, Q. M. et al. Lef1 expression in fibroblasts maintains developmental potential in adult skin to regenerate wounds. Elife 9:e60066.(2020).
Almet, A. A. et al. Integrated Single-Cell Analysis Reveals Spatially and Temporally Dynamic Heterogeneity in Fibroblast States during Wound Healing. J. Invest. Dermatol. 145(3):645-659.e25 (2025).
Kamat, P. et al. Single-cell morphology encodes functional subtypes of senescence in aging human dermal fibroblasts. Sci. Adv. 11 (17), eads1875 (2025).
Bancroft, T. et al. Up-regulation of thrombospondin-2 in Akt1-null mice contributes to compromised tissue repair due to abnormalities in fibroblast function. J. Biol. Chem. 290 (1), 409–422 (2015).
Agah, A. et al. Thrombospondin 2 levels are increased in aged mice: consequences for cutaneous wound healing and angiogenesis. Matrix Biol. 22 (7), 539–547 (2004).
Kunkemoeller, B. et al. Elevated thrombospondin 2 contributes to delayed wound healing in diabetes. Diabetes 68 (10), 2016–2023 (2019).
Kyriakides, T. R., Tam, J. W. & Bornstein, P. Accelerated wound healing in mice with a disruption of the thrombospondin 2 gene. J. Invest. Dermatology. 113 (5), 782–787 (1999).
Armstrong, L. C. et al. Thrombospondin 2 inhibits microvascular endothelial cell proliferation by a caspase-independent mechanism. Mol. Biol. Cell. 13 (6), 1893–1905 (2002).
Gao, Q. et al. Thrombospondin-1 signaling through CD47 inhibits cell cycle progression and induces senescence in endothelial cells. Cell Death Dis. 7 (9), e2368–e2368 (2016).
Kyriakides, T. R. et al. Mice that lack thrombospondin 2 display connective tissue abnormalities that are associated with disordered collagen fibrillogenesis, an increased vascular density, and a bleeding diathesis. J. Cell Biol. 140 (2), 419–430 (1998).
MacLauchlan, S. et al. Enhanced angiogenesis and reduced contraction in thrombospondin-2–null wounds is associated with increased levels of matrix metalloproteinases-2 and – 9, and soluble VEGF. J. Histochem. Cytochemistry. 57 (4), 301–313 (2009).
Chen, Z. et al. Novel muscle-derived extracellular matrix hydrogel promotes angiogenesis and neurogenesis in volumetric muscle loss. Matrix Biol. 127, 38–47 (2024).
Chen, Z. et al. Bone-derived extracellular matrix hydrogel from thrombospondin-2 knock-out mice for bone repair. Acta Biomater. 186, 85–94 (2024).
Morris, A. H. et al. Tunable hydrogels derived from genetically engineered extracellular matrix accelerate diabetic wound healing. ACS Appl. Mater. Interfaces. 10 (49), 41892–41901 (2018).
Morris, A. H. et al. Decellularized materials derived from TSP2-KO mice promote enhanced neovascularization and integration in diabetic wounds. Biomaterials 169, 61–71 (2018).
Hao, Y. et al. Dictionary learning for integrative, multimodal and scalable single-cell analysis. Nat. Biotechnol. 42 (2), 293–304 (2024).
McInnes, L., Healy, J. & Melville, J. Umap: Uniform manifold approximation and projection for dimension reduction. arXiv preprint arXiv:1802.03426, (2018).
Raredon, M. S. B. et al. Comprehensive visualization of cell–cell interactions in single-cell and Spatial transcriptomics with NICHES. Bioinformatics 39 (1), btac775 (2023).
Wickham, H., Chang, W. & Wickham, M. H. Package ‘ggplot2’. Create elegant data visualisations using the grammar of graphics. Version 2 (1), 1–189 (2016).
Kolde, R. & Kolde, M. R. Package ‘pheatmap’. R Package. 1 (7), 790 (2015).
Wang, W. et al. Ebf2 is a selective marker of brown and beige adipogenic precursor cells. Proc. Natl. Acad. Sci. 111 (40), 14466–14471 (2014).
Deckx, S., Heymans, S. & Papageorgiou, A. P. The diverse functions of osteoglycin: a deceitful dwarf, or a master regulator of disease? FASEB J. 30 (8), 2651–2661 (2016).
Katoh, D. et al. Tenascin-C induces phenotypic changes in fibroblasts to myofibroblasts with high contractility through the integrin αvβ1/transforming growth factor β/SMAD signaling axis in human breast cancer. Am. J. Pathol. 190 (10), 2123–2135 (2020).
Wang Z, Zhuang X, Chen B, Wei M. Osteoglycin knockdown promotes vascular smooth muscle cell proliferation and migration in aortic dissection via the VEGF/VEGFR2 axis. Mol Med Rep. 2021;23(1):65.
Mitola, S. et al. Gremlin is a novel agonist of the major proangiogenic receptor VEGFR2. Blood. J. Am. Soc. Hematol. 116 (18), 3677–3680 (2010).
Eccleston, P. et al. Schwann cells secrete a PDGF-like factor: evidence for an autocrine growth mechanism involving PDGF. Eur. J. Neurosci. 2 (11), 985–992 (1990).
Meier, C. et al. Developing Schwann cells acquire the ability to survive without axons by Establishing an autocrine circuit involving insulin-like growth factor, neurotrophin-3, and platelet-derived growth factor-BB. J. Neurosci. 19 (10), 3847–3859 (1999).
Hoving, J. J. et al. N-cadherin directs the collective Schwann cell migration required for nerve regeneration through Slit2/3-mediated contact Inhibition of locomotion. Elife 13, e88872 (2024).
Li, Y. et al. Notch and Schwann cell transformation. Oncogene 23 (5), 1146–1152 (2004).
Woodhoo, A. et al. Notch controls embryonic Schwann cell differentiation, postnatal myelination and adult plasticity. Nat. Neurosci. 12 (7), 839–847 (2009).
Alimperti, S. & Andreadis, S. T. CDH2 and CDH11 act as regulators of stem cell fate decisions. Stem Cell. Res. 14 (3), 270–282 (2015).
Liu, J. et al. JAG1 enhances angiogenesis in triple-negative breast cancer through promoting the secretion of exosomal lncRNA MALAT1. Genes Dis. 10 (5), 2167–2178 (2023).
Zhong, J. et al. Neovascularization of ischemic tissues by gene delivery of the extracellular matrix protein Del-1. J. Clin. Invest. 112 (1), 30–41 (2003).
Pin, A. L. et al. Annexin-1-mediated endothelial cell migration and angiogenesis are regulated by vascular endothelial growth factor (VEGF)-induced Inhibition of miR-196a expression. J. Biol. Chem. 287 (36), 30541–30551 (2012).
Ramilowski, J. A. et al. A draft network of ligand-receptor-mediated multicellular signalling in human. Nat. Commun. 6, 7866 (2015).
Raredon, M. S. B. et al. Single-cell connectomic analysis of adult mammalian lungs. Sci. Adv. 5 (12), eaaw3851 (2019).
Battegay, E. J. et al. PDGF-BB modulates endothelial proliferation and angiogenesis in vitro via PDGF beta-receptors. J. Cell Biol. 125 (4), 917–928 (1994).
Li, Z. et al. BMP4 signaling acts via dual-specificity phosphatase 9 to control ERK activity in mouse embryonic stem cells. Cell. Stem Cell. 10 (2), 171–182 (2012).
Mammoto, T. et al. Nerve growth factor improves ligament healing. J. Orthop. Res. 26 (7), 957–964 (2008).
Muhl, L. et al. Single-cell analysis uncovers fibroblast heterogeneity and criteria for fibroblast and mural cell identification and discrimination. Nat. Commun. 11 (1), 3953 (2020).
Ascensión, A. M. et al. Human dermal fibroblast subpopulations are conserved across Single-Cell RNA sequencing studies. J. Invest. Dermatol. 141 (7), 1735–1744e35 (2021).
Marangoni, R. G. et al. Thy-1 plays a pathogenic role and is a potential biomarker for skin fibrosis in scleroderma. JCI insight 7 (19) (2022).
Koumas, L. et al. Thy-1 expression in human fibroblast subsets defines myofibroblastic or lipofibroblastic phenotypes. Am. J. Pathol. 163 (4), 1291–1300 (2003).
Mascharak, S. et al. Preventing Engrailed-1 activation in fibroblasts yields wound regeneration without scarring. Science 372(6540) (2021).
Jeffery, T. K. et al. BMP4 inhibits proliferation and promotes myocyte differentiation of lung fibroblasts via Smad1 and JNK pathways. Am. J. Physiol. Lung Cell. Mol. Physiol. 288 (2), L370–L378 (2005).
Setiawan, A. M., Kamarudin, T. A., Abd, N. & Ghafar The role of BMP4 in adipose-derived stem cell differentiation: A minireview. Front. Cell. Dev. Biol. 10, 1045103 (2022).
Gustafson, B. et al. BMP4 and BMP antagonists regulate human white and beige adipogenesis. Diabetes 64 (5), 1670–1681 (2015).
Jeong, S. et al. BMP4 and perivascular cells promote hematopoietic differentiation of human pluripotent stem cells in a differentiation stage-specific manner. Exp. Mol. Med. 52 (1), 56–65 (2020).
Moon, B. S. et al. Bone morphogenetic protein 4 stimulates neuronal differentiation of neuronal stem cells through the ERK pathway. Exp. Mol. Med. 41 (2), 116–125 (2009).
Qi, Z. et al. BMP restricts stemness of intestinal Lgr5(+) stem cells by directly suppressing their signature genes. Nat. Commun. 8, 13824 (2017).
Thoma, E. C. et al. Chemical conversion of human fibroblasts into functional Schwann cells. Stem Cell. Rep. 3 (4), 539–547 (2014).
Meng, H. et al. Thrombospondin 2 potentiates notch3/jagged1 signaling. J. Biol. Chem. 284 (12), 7866–7874 (2009).
Pan, W. et al. TSP2 acts as a suppresser of cell invasion, migration and angiogenesis in Medulloblastoma by inhibiting the Notch signaling pathway. Brain Res. 1718, 223–230 (2019).
Direder, M. et al. Schwann cells contribute to keloid formation. Matrix Biol. 108, 55–76 (2022).
Direder, M. et al. The transcriptional profile of keloidal Schwann cells. Exp. Mol. Med. 54 (11), 1886–1900 (2022).
Gong, T. et al. Single-cell RNA-seq reveals the communications between extracellular matrix-related components and Schwann cells contributing to the earlobe keloid formation. Front. Med. (Lausanne). 9, 1000324 (2022).
Wei, K. et al. Schwann cells secrete IGFBP5 to facilitate the growth of keloids. Life Sci. 369, 123534 (2025).
Acknowledgements
We want to thank Yale University Keck Microarray Shared Resource, Keck DNA Sequencing Facility and Yale Harvey Cushing/John Hay Whitney Medical Library for their assistance.
Funding
This project was supported by NIH grant DK132645 and National Institute of General Medical Sciences of the National Institutes of Health under Award Number 1S10OD030363-01 A.
Author information
Authors and Affiliations
Contributions
Y.H. contributed to the design, conduction, data analysis, interpretation of all experiments. N.W. contributed to scRNA seq analysis. H.X. contributed to RNA seq data collection. D.G., H.C.H. contributed to flow cytometry. S. R. contributed to data analysis and interpretation. T.R.K. contributed to project design and data interpretation. Y.H., T.R.K., and S. R. wrote the paper and all authors provided feedback.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Huang, Y., Wang, N., Xing, H. et al. Single cell RNA seq reveals the pro-regenerative phenotype of thrombospondin-2 deficient dermal fibroblasts. Sci Rep 15, 32304 (2025). https://doi.org/10.1038/s41598-025-15839-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-15839-3
