
Abstract
Tuberculosis (TB) remains a major global health challenge. Peripheral blood immune cell composition provides valuable insights into TB progression and management. In this study, we analyzed cellular dynamics across the TB disease spectrum using 43 global transcriptomic datasets encompassing 5,902 blood samples. Distinct immune changes were identified during the early stages of TB progression. Transition from latent infection to incipient TB was associated with reduced proportions of natural killer (NK) cells. In subclinical TB, monocyte proportions increased further, accompanied by additional reductions in NK cells and B cells. These early immune shifts preceded the pronounced alterations observed in active TB, characterized by elevated monocytes and neutrophils alongside markedly decreased lymphocyte populations. During successful anti-TB treatment, immune profiles gradually normalized. Cellular dynamics were also influenced by TB burden, age, and HIV coinfection, with stronger immune responses observed in adults and in regions with lower TB burdens. Overall, this study highlights early-stage peripheral blood biomarkers, particularly NK cell changes, as potential indicators of TB progression and targets for preventive interventions.
Similar content being viewed by others

Introduction
Tuberculosis (TB), caused by Mycobacterium tuberculosis (Mtb), is one of the oldest known diseases and remains the deadliest infectious disease globally1. In 2023, approximately 10.8 million people were diagnosed with TB, leading to 1.25 million deaths worldwide1. To combat the ongoing TB epidemic and reduce mortality, the World Health Organization (WHO) set a target in 2015 to “end the TB epidemic by 2035” through proactive screening and preventive treatment for high-risk populations2. Early diagnosis and timely intervention are particularly critical for minimizing diagnostic delays and controlling the spread of TB within communities3.
In TB research, examining the cellular composition of peripheral blood can significantly support early diagnosis and intervention across multiple dimensions4. First, the immune system’s capacity to recognize and control Mtb infection is essential. Peripheral blood is an easily accessible source of immune cells, including monocytes, neutrophils, and T cells. These cells offer valuable insights into the immune responses that influence TB control and progression4. For example, CD4+ T cells, particularly those that produce interferon gamma (IFN-γ), play a key role in activating macrophages to eliminate bacteria5. However, immune response imbalances, such as excessive inflammation or immune suppression, can either improve or worsen TB outcomes6. Compared with those with active TB (ATB), people with latent TB infection (LTBI) display a distinct immune cell profile, offering insights into the immune system’s shift from containing bacteria to allowing active disease5. Second, the blood immune cell composition can indicate the TB stage and the ability of the body to fight infection. Patients with ATB often exhibit higher levels of proinflammatory cytokines and altered frequencies of monocytes and neutrophils than do those with LTBI or healthy individuals7. Additionally, blood tests revealing elevated neutrophil counts or altered monocyte‒lymphocyte ratios are commonly linked with severe or disseminated TB8. Monitoring these changes enables clinicians to predict disease outcomes and adjust treatment strategies. Third, peripheral blood biomarkers are essential for enhancing TB diagnostics and monitoring treatment progress. Cellular markers, such as specific surface protein expression levels (e.g., PD-1 on T cells or CD38 on monocytes) and cytokine production patterns, can indicate TB activity or treatment response. For example, increased CD38+ CD4+ T cell levels are associated with ATB and can help distinguish ATB from LTBI9. Thus, studying peripheral blood cellular compositions is essential for understanding immune responses in TB, assessing disease progression, and identifying diagnostic biomarkers.
The compositional changes in peripheral immune cells during TB development and treatment have been thoroughly explored via blood transcriptomes10. A pioneering study on blood transcriptomes in ATB patients reported a decrease in lymphocytes, including CD4+ T cells, CD8+ T cells, and B cells, in ATB patients within the Leicester cohort11,12. Furthermore, a reduction in polyfunctional type 17 helper T (Th17) cell-like memory T cells was observed among individuals who progressed from Mtb infection to ATB in a Peruvian TB progression cohort13. In a single-cell transcriptome analysis of peripheral blood samples from healthy controls and LTBI and ATB patients, Cai et al. reported a gradual depletion of CD3− CD7+ GZMB+ natural killer (NK) cells with TB disease progression in a Chinese cohort14. This depletion of circulating NK cells was reversed following anti-TB treatment (ATT). However, a distinct NK-like CD8+ γδ T cell subset was expanded in patients with persistent Mtb infection within the ACS cohort15. Conversely, certain myeloid cells, such as inflammatory monocytes, were found to be elevated in ATB patients11,12. For example, the number of circulating CD14+ CD16+ monocytes was elevated in ATB patients in one Chinese cohort14. In contrast, Lyu et al. reported no such changes in monocytes in another Chinese cohort16. They only reported a significant decrease in naive CD4+ T cells in ATB patients. These findings indicate substantial changes in circulating immune cells across various stages of Mtb infection, although the diversity of cellular responses across different cohorts could limit their diagnostic value for TB in practical settings. To strengthen the reliability of these cellular changes as circulating biomarkers during TB development and treatment, Roy Chowdhury et al. performed a computational deconvolution analysis of cell proportions in the peripheral blood across more than 10 clinical cohorts from three continents, including uninfected individuals (UI), LTBI, ATB, and treated TB patients, via blood transcriptomic datasets17. Their analysis revealed a consistent increase in the number of circulating NK cells in LTBI patients, a decrease in ATB, and a return to baseline following clinical recovery. Importantly, this change in circulating NK cells was further validated in an independent cohort, the South African longitudinal cohort, confirming its association with TB progression, treatment response, and the local inflammatory profile in ATB patients17. This underscores the importance and applicability of integrating heterogeneous datasets to increase the reproducibility, robustness and generalizability of the use of cellular biomarkers for TB diagnosis and treatment18,19.
While existing studies have revealed widespread changes in the circulating blood cell composition during TB development and treatment, most findings rely on a simplified binary classification of TB states—LTBI and ATB. Although this model has helped prevent millions of TB-related deaths, it has had a limited effect on reducing transmission and disease incidence, likely because millions of TB-infected individuals do not seek healthcare or receive treatment20. Recent studies have further refined the classification of asymptomatic LTBI individuals into three categories: LTBI non-progressors (LTBI NP), incipient TB patients (ITB), and subclinical TB patients (STB)21,22. Among these individuals, STB patients contribute significantly to Mtb transmission despite showing no clinical symptoms23. Therefore, a deeper understanding of stage-specific cellular compositional changes throughout the TB spectrum, particularly in the disease’s early stages, is essential for advancing knowledge of TB progression and detection22. To meet this need, we expanded our meta-analysis of blood cellular compositions in human TB by including additional public blood transcriptome datasets and categorizing TB-related samples within a framework that considers key asymptomatic stages in the TB disease spectrum20. We conducted a systematic analysis of changes in the circulating blood cell composition following Mtb infection and post-TB treatment across global TB cohorts (Fig. 1). Additionally, we examined the impact of potential confounding factors, such as local TB disease burden, age, and HIV coinfection, on these compositional changes throughout TB progression.
Materials and methods
Data collection
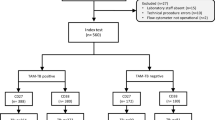
We searched for blood transcriptomic datasets related to human TB from NCBI GEO24 and EBI ArrayExpress25 in April 2024 via the terms “tuberculosis”, “transcriptome”, “Homo sapiens”, “whole blood OR peripheral blood mononuclear cells OR PBMCs” (Supplementary Figure S1). This search yielded 43 transcriptomic datasets primarily focused on diagnosing LTBI or ATB, predicting TB progression, or monitoring treatment (Supplementary Table S1; Supplementary Notes, Sect. 1). In total, the dataset comprised gene expression data from 5,902 blood samples across five continents. All raw data were downloaded and processed using a uniform preprocessing procedure to obtain protein-coding gene expression profiles (Supplementary Notes, Sect. 2). We identified that some cohorts included overlapping individuals (Supplementary Table S1). To maximize data availability and completeness, we retained these overlapping samples within their respective cohorts and analyzed each dataset as an independent cohort. These overlaps are unlikely to bias results given the size and diversity of the datasets. Leukocyte proportions were subsequently estimated from gene expression profiles using the CIBERSORTx26 cell-mixture deconvolution approach (Supplementary Notes, Sect. 3; Supplementary Figure S2).
Phenotype data processing
For decades, TB disease states have been classified using a binary model of LTBI and ATB27. While widely used, this simple model has several limitations. LTBI individuals are broadly defined as the presence of Mtb without clinical TB symptoms or infectiousness, whereas ATB is diagnosed by positive culture or molecular tests accompanied by symptoms such as fever, fatigue, appetite loss, and weight loss. To consolidate evidence on post-infection outcomes, we reviewed definitions of TB disease states from recent studies28,29,30. These criteria incorporated immunologic tests (e.g., tuberculin skin test, IFN-γ release assay), microbiologic assessments (e.g., bronchoalveolar lavage, pleural fluid, or sputum culture), radiographic findings (e.g., chest X-ray, PET/CT), and clinical symptoms. Based on these criteria, individuals with recent Mtb exposure could be classified into the following groups (Supplementary Figure S4):
-
1.
UI (Uninfected individuals): Individuals uninfected or who cleared bacteria through innate or adaptive immunity during follow-up. Samples with < 6 months of follow-up from the date of blood draw were excluded to minimize misclassification)31.
-
2.
LTBI NP (Latent TB infection, non-progressors): Infected individuals who remained TB-free during follow-up, with the same exclusion criteria as above31.
-
3.
ITB (Incipient TB): Individuals who progress to ATB during follow-up but showed no microbiological evidence, radiological abnormalities, or clinical symptoms at the time of blood draw.
-
4.
STB (Subclinical TB): Individuals who progress to ATB during follow-up, showing no clinical symptoms but presenting with immunological, microbiological, or radiological abnormalities at the time of blood draw. Cases with subsequent remission were excluded to reduce misclassification.
-
5.
ATB (Active TB): Newly diagnosed cases or cases detected through passive surveillance, defined by the presence of clinical symptoms, radiological abnormalities, and microbiological evidence.
To analyze cellular changes across TB stages, particularly the early asymptomatic stages, we redefined TB disease states in 37 cross-sectional studies based on these criteria. We also standardized the timing of blood samples for contacts who developed ATB as “time to diagnosis” across 43 datasets, enabling sequential assessment of TB progression (Supplementary Notes, Sect. 4). Healthy controls were annotated as UIs for consistency (Supplementary Table S1). Additionally, samples were grouped by TB burden, age, and HIV status to assess the influence of potential confounding factors (Supplementary Notes, Sect. 4).
Blood cellular composition comparison
To enhance robustness, we conducted multi-cohort case-control comparisons of blood cellular composition across different TB stages in both cross-sectional and longitudinal cohorts. A bivariate meta-analysis (sample size ≥ 5) with a random-effects model17 was performed using MetaIntegrator32. The resulting summary effect size (ES) indicated whether cellular proportions increased or decreased across disease stages (Supplementary Notes, Sect. 5.1). To assess the heterogeneity of ES estimates across studies, we also calculated a consistency score (C_score). To account for potential confounding factors such as country, age, and HIV coinfection, comparisons were further adjusted using linear regression (Supplementary Notes, Sect. 5.2). Group-wise analyses were then performed to evaluate compositional differences by age group, TB burden, and HIV status, with summary ES values from the meta-analysis serving as the comparison metric (Supplementary Notes, Sect. 5.3). Statistical significance was assessed using a false discovery rate (FDR) threshold of 0.1.
Results
Changes in peripheral blood cell composition in LTBI and ATB
We estimated the proportions of two myeloid cell types (monocytes and neutrophils) and four lymphocyte types (CD8+ T cells, CD4+ T cells, NK cells, and B cells) from 5,902 blood samples across all transcriptomic datasets (Supplementary Table S1; Supplementary Figure S3). Using these estimates, we first categorized TB-related samples according to the classical binary model and compared the cellular compositions of LTBI individuals and ATB patients with those of UI individuals (Fig. 1A).
In LTBI individuals, monocyte exhibited the most significant increase (ES = 0.345, FDR < 0.001), with largely consistent results (C_score = 0.429) across 14 cohorts (Fig. 2A and B; Supplementary Tables S3A, S4A). No significant changes were observed in the proportions of neutrophils, CD4+ T cells, CD8+ T cells, NK cells, or B cells (Supplementary Figure S5A; Supplementary Table S4A). In ATB patients, cellular shifts were more pronounced across 20 global cohorts (Fig. 2A and C; Supplementary Figure S5B; Supplementary Tables S3B, S4B). Monocyte (ES = 1.502, FDR < 0.001) and neutrophil proportions (ES = 0.386, FDR < 0.001) significantly increased, while CD4+ T cells (ES = -0.76, FDR < 0.001), CD8+ T cells (ES = -0.516, FDR < 0.001), NK cells (ES = -0.751, FDR < 0.001), and B cells (ES = -0.507, FDR < 0.001) significantly decreased. These compositional changes, particularly the monocyte increase, were consistent across cohorts (C_score = 0.9). Notably, these changes remained robust after adjustment for country, age, and HIV status (Supplementary Figures S6A, S6B; Supplementary Table S5A). A direct comparison of LTBI and ATB individuals across 25 cohorts revealed patterns similar to those observed between ATB and UI individuals (Fig. 2A; Supplementary Figure S5C; Supplementary Tables S3C, S4C).
Overall, these findings indicate that immune compositional changes begin in the LTBI state, characterized by monocyte expansion, are broaden to involve all major blood cell types as the disease progresses to ATB.
Changes in peripheral blood cell composition during the early stages of TB development
In the previous multi-cohort analysis, LTBI individuals were defined as those infected with Mtb but without progression to disease. Under this framework, early-stage cellular changes remain poorly understood, despite some compositional alterations observed in LTBI individuals. To address this gap, we examined cellular shifts more closely, as they may provide insights for preventive therapies and reduce Mtb transmission (Fig. 1B).
In the revised TB disease spectrum framework, asymptomatic LTBI individuals were further classified into three groups: LTBI NP, ITB, and STB. We compared blood cell proportions in these asymptomatic states with those in UI individuals across multiple global cohorts. Among LTBI NP individuals, no significant differences in myeloid or lymphocyte cell compositions were observed compared with UI individuals across six cohorts (Fig. 3A; Supplementary Figure S7A; Supplementary Tables S6A, S7A). In ITB patients, NK cells were significantly decreased in one cohort (ES = -1.249, FDR = 0.005; Fig. 3A; Supplementary Figure S7B; Supplementary Tables S6B, S7B). Among STB patients, monocyte proportions significantly increased across two cohorts (ES = 1.121, FDR < 0.013; Fig. 3A and B; Supplementary Tables S6C, S7C), whereas NK cells (ES = -0.605, FDR = 0.055) and B cells (ES = -0.839, FDR = 0.004) significantly decreased. Neutrophils tended to increase and T cells to decrease across cohorts, although these changes did not reach statistical significance (Fig. 3A and B; Supplementary Figure S7C). Importantly, these compositional shifts remained after adjusting for country, age, and HIV status, suggesting they were driven primarily by disease progression (Supplementary Figures S6C, S6D; Supplementary Table S5B).
We next examined changes between consecutive disease stages (Fig. 1B). No significant differences were found between LTBI NP and ITB, or between ITB and STB (Fig. 3A; Supplementary Figures S7D, S7E; Supplementary Tables S6D, S6E, S7D, S7E). By contrast, prominent changes were observed between STB and ATB patients across four cohorts (Fig. 3A; Supplementary Tables S6F, S7F). Among myeloid cells, monocytes showed significant increases in ATB patients (ES = 0.664, FDR = 0.027), whereas neutrophils displayed a non-significant upward trend (ES = 0.508, FDR = 0.237; Fig. 3C). Among lymphocytes, CD4+ T cells were significantly reduced in ATB (ES = -0.692, FDR = 0.001), while CD8+ T cells (ES = -0.381, FDR = 0.199), NK cells (ES = -0.415, FDR = 0.191), and B cells (ES = -0.382, FDR = 0.237) all demonstrated downward but non-significant trends (Supplementary Figure S7F).
In summary, no significant compositional changes were detected in the non-infectious stage of TB across these limited cohorts. NK cell reduction emerged early during progression to ITB, followed by increased monocytes and decreased B cells in STB. The transition from STB to ATB was marked by further monocyte expansion alongside significant CD4+ T cell depletion.
Temporal changes in peripheral blood cell composition during TB progression
Several longitudinal studies have investigated temporal changes in peripheral blood transcriptomes as TB contacts progress from TB infection to ATB28,33,34,35,36. These studies provide valuable insights into the cellular dynamics preceding clinical diagnosis, using cohorts such as Zak_SA33, Suliman_GC6-7434, Singhania_Leicester35, Tabone_Leicester28, and Roe_UK36 cohorts (Fig. 1C; Supplementary Table S8). We divided longitudinal blood samples into six time periods (Supplementary Notes, Sect. 4) preceding TB diagnosis and compared cell compositions at each interval to those of UI individuals.
Monocyte proportions began increasing as early as 360–540 days before diagnosis (ES = 0.649, FDR = 0.002) and continued to rise at 360–180 days before diagnosis (ES = 1.066, FDR = 0.016; Fig. 4A and B). Between 180 and 60 days prior to diagnosis, significant reductions in B cells were observed (ES = -0.463, FDR = 0.022; Fig. 4A and C; Supplementary Figure S8C). Interestingly, compositional changes became less pronounced as diagnosis approached, in contrast to patterns observed in cross-sectional cohorts (Figs. 4A; Supplementary Figure S8; Supplementary Tables S9). Monocytes nevertheless exhibited a consistent increase between 60 and 21 days before diagnosis (ES = 1.245, FDR < 0.001; C_score = 1; Fig. 4A; Supplementary Figure S8E). In the final 21 days before diagnosis, monocytes (ES = 1.222, FDR < 0.001), NK cells (ES = -0.978, FDR < 0.001), and B cells (ES = -0.82, FDR < 0.001; Fig. 4A and D; Supplementary Figure S8F) all underwent significant compositional changes. These results demonstrate that peripheral blood cell alterations occur in distinct temporal patterns throughout TB progression.
We further examined the compositional changes in individual TB progressors across four longitudinal cohorts: Zak_SA33, Suliman_GC6-7434, Singhania_Leicester35, and Tabone_Leicester28 cohorts (Fig. 1C). In the Zak_SA cohort, nine out of eighteen progressors exhibited the highest proportion of monocytes in the earliest time period before diagnosis (Fig. 4E). Some individuals, such as patients 06_0127 and 07_0259, showed a decrease in monocyte proportions as disease advanced. In the Suliman_GC6-74 cohort, seven of twenty progressors displayed rising monocyte proportions during follow-up (Fig. 4F). Similar but heterogeneous patterns were observed in the Singhania_Leicester (Supplementary Figure S9) and Tabone_Leicester (Supplementary Figure S10) cohorts, where only ~ 50% of progressors demonstrated monocyte increase prior to diagnosis. For NK cells, reductions were detected in eight of eighteen progressors in the Zak_SA cohort (Fig. 4E) and eight of twenty in the Suliman_GC6-74 cohort (Fig. 4F). In contrast, more than two-thirds of progressors in the Singhania_Leicester and Tabone_Leicester cohorts exhibited NK cell declines (Supplementary Figures S9, S10). Considerable heterogeneity was also evident in other blood cell types, including B cells, T cells, and neutrophils (Supplementary Figures S9-S12).
Collectively, these findings reinforce the overarching trend of monocyte expansion and concomitant NK and B cell decline during TB progression. However, substantial inter-patient and inter-cohort variability in blood cell dynamics was observed. While monocyte increases were consistent across all time periods, other blood cell populations showed fluctuating changes at different intervals. The earliest detectable alterations occurred between 360 and 60 days prior to TB diagnosis, marking a critical window in TB development.
Temporal changes in peripheral blood cell composition during ATT
In addition to pre-diagnosis changes, longitudinal cohorts have also provided insights into how peripheral blood cell compositional shifts during ATT. Using data from nine global cohorts, we tracked cellular dynamics across four intervals following treatment initiation: [7, 28], (28, 60], (60, 182], and (182, 360] days during ATT (Fig. 1D; Supplementary Table S10A). Compared with untreated ATB patients, myeloid cell populations (monocytes and neutrophils) progressively declined, while lymphocyte populations (CD4+ T cells, CD8+ T cells, NK cells, and B cells) increased (Fig. 5A; Supplementary Table S11; Supplementary Figure S13).
During the early treatment phase (7–28 days post-treatment), monocytes decreased significantly (ES = -0.702, FDR < 0.001; Fig. 5A and B), accompanied by increases in CD4+ T cells (ES = 0.32, FDR = 0.053), CD8+ T cells (ES = 0.494, FDR = 0.004), and NK cells (ES = 0.383, FDR = 0.019). Neutrophil reductions emerged between 28 and 60 days (ES = -0.538, FDR = 0.018), and B cell expansion became evident between 60 and 182 days (Fig. 5A and C). By 60–182 days and extending to 360 days post-treatment, significant shifts were observed across all major cell types, coinciding with the presumed clinical cure of ATB patients (Supplementary Figure S13).
To explore the relationship between these compositional changes and treatment outcomes, we analyzed the Thompson_SA cohort37, where patients were categorized as “cured”, “probable cured”, or “not cured” (Supplementary Table S10B). Cured patients exhibited rapid reductions in monocytes and increases in CD4+ and CD8+ T cells within the first 7 days, followed by declined neutrophils during days 14–30 days (Fig. 5D; Supplementary Table S12A). These patterns mirrored the global cohort trends. In contrast, the “probable cured” group showed more gradual changes (Supplementary Table S12B), while the “not cured” group exhibited minimal alterations – most notably, no significant changes in monocytes or B cells and only a modest NK cell increase between 120 and 180 days (Supplementary Table S12C). A similar pattern was observed in the Tabone_Leicester28 cohort (Supplementary Table S10B). Patients requiring extended ATT resembled the “probable cured” group, with significant shifts in most cell types occurring only after 60 days (Fig. 5D and E; Supplementary Table S12E). By contrast, patients on standard ATT displayed much weaker changes, limited to reduced monocytes and increased B cells after one year (Supplementary Table S12D). The weakest and most delayed responses were observed in difficult TB cases, drug-resistant TB cases, and outbreak TB strain subgroups (Fig. 5E; Supplementary Figure S14; Supplementary Tables S12F-12H). At the individual level, patient trajectories revealed notable heterogeneity in both the Thompson_SA (Supplementary Figure S15) and Tabone_Leicester (Supplementary Figure S16) cohorts. While population-level trends showed consistent reversal of TB-associated changes, patient-specific factors clearly influenced the magnitude and timing of recovery.
Overall, these findings highlight the temporal dynamics of immune recovery during ATT. The consistent pattern of declining monocytes and neutrophils alongside rising NK cells, CD4+ T cells, CD8+ T cells, and B cells reflects a reversal of TB-induced compositional shifts and is strongly associated with treatment efficacy (Fig. 5D and E). However, inter-patient variability underscores the need to account for individual and cohort-specific factors when interpreting immune responses to therapy.
Effect of confounding factors on blood cellular changes during TB development
Understanding how TB progression alters peripheral blood cell composition is crucial for identifying biomarkers and disease stages. However, these changes can be influenced by extrinsic factors. To address this, we conducted multi-cohort meta-comparisons across subgroups defined by local TB burden, age and HIV coinfection status (Fig. 1E; Supplementary Notes, Sect. 4).
First, we examined whether the local TB burden influenced the compositional changes in blood cells in different stages of TB (Supplementary Table S3). Comparisons between high- and low-burden settings revealed broadly consistent cellular patterns across disease stages, but with notable subgroup-specific differences (Fig. 6A; Supplementary Figure S17A; Supplementary Table S13). In LTBI individuals, both subgroups showed overall trends similar to the global analysis (Fig. 2A), yet neutrophils did not change significantly in either subgroup, and NK cell reductions were observed only in low-burden cohorts. Among ATB patients, cellular changes in both subgroups also mirrored global patterns (Fig. 3A). However, in asymptotic LTBI NP individuals, monocytes increased significantly only in the low-burden cohorts (Fig. 6A; Supplementary Figure S17A). More broadly, monocytes, neutrophils, NK cells, and B cells differed significantly between high- and low-burden LTBI groups; monocytes, CD4+ T cells, CD8+ T cells, and B cells differed in ATB patients; and monocytes, CD4+ T cells, NK cells, and B cells differed in LTBI NP individuals. Overall, individuals in low-burden regions exhibited more pronounced changes across stages, likely reflecting the immunological naivete of UI individuals in these settings. Second, we explored the effects of age on blood cellular changes during TB progression by grouping individuals into age categories: adolescents, adults, and seniors (Supplementary Tables S3, S6). For LTBI individuals, the compositional changes in adults (Fig. 6B) were largely consistent with those observed in the global cohort analysis (Fig. 2A; Supplementary Figure S17B; Supplementary Table S14). However, there was a significant increase in B cells exclusively in the adult subgroup. In contrast, adolescents and seniors presented less pronounced changes in blood cell composition, with no significant changes in most cell types, although adolescents presented a notable decrease in B-cell proportions at the LTBI stage (Fig. 6B; Supplementary Figure S17B). These age-related differences persisted across other disease stages, including ATB patients and individuals in asymptomatic disease states. These results suggest that adults experience more significant and consistent blood cellular changes than adolescents or seniors do. This may be explained by the generally immunocompromised status of adolescents (due to ongoing immune system development) and seniors (due to the aging immune system). Third, we also evaluated the impact of HIV coinfection. Because of limited sample sizes (Supplementary Tables S3, S6), results were largely indistinguishable from analyses of HIV-negative individuals, with overall compositional changes closely matching those in the global cohorts (Supplementary Figure S17; Supplementary Table S15).

Schematic framework of the experimental design. (A) Multi-cohort analysis of changes in circulating blood cell compositions during the latent TB infection (LTBI) and active disease (ATB) stages. (B) The latent infection stage was further subdivided into three asymptomatic stages: latent infection without disease progression (LTBI NP), incipient TB (ITB), and subclinical TB (STB). Compositional changes during the early stages of TB development were analyzed through multi-cohort comparisons. (C) Temporal changes in circulating blood cell compositions were examined in longitudinal cohorts of individuals who eventually progressed to ATB. (D) Temporal variations in blood cell proportions were analyzed in TB patients receiving anti-tuberculosis treatment (ATT). (E) The effects of factors such as TB burden, age, and HIV coinfection on cellular changes in circulating blood during TB development were also investigated.

Compositional changes in peripheral blood cells among LTBI individuals and ATB patients across global cohorts. (A) Bubble plot illustrating the compositional changes in six cell types across three comparisons: LTBI individuals versus UI patients, ATB patients versus UI patients, and ATB patients versus LTBI individuals. Red bubbles indicate an increase in the cellular composition, whereas blue bubbles indicate a decrease. The intensity of the color reflects the degree of change, with a false discovery rate (FDR) threshold of 0.1 for statistical significance. The bubble size represents the absolute magnitude of the effect size. White circles denote comparisons with no significant differences. (B) Forest plots showing the compositional changes in monocytes, neutrophils, and NK cells between LTBI individuals and UI patients. (C) Forest plots depicting the compositional changes in monocytes, neutrophils, and NK cells between ATB patients and UI patients. The GEO or ArrayExpress identifiers represent individual cohorts. Each circle indicates the standardized mean difference (effect size, ES) in cellular composition between two groups, with the circle size proportional to the cohort sample size. The horizontal strip of each circle denotes the 95% confidence interval for the ES. The diamond represents the overall difference in cellular proportions between two groups, derived by integrating ES values from all cohorts (Summary). The FDR value indicates the statistical significance of the overall cellular compositional difference across cohorts.

Compositional changes in peripheral blood cells during the early stages of TB development across global cohorts. (A) Bubble plot illustrating compositional changes in six cell types across various comparisons: LTBI NP versus UI individuals, ITB patients versus UI individuals, STB patients versus UI individuals, ITB patients versus LTBI NP individuals, STB patients versus ITB patients, and ATB patients versus STB patients. Red bubbles indicate an increase in the cellular composition, whereas blue bubbles indicate a decrease. The intensity of the color reflects the degree of change, with a false discovery rate (FDR) threshold of 0.1 for statistical significance. The bubble size represents the absolute magnitude of the effect size. White circles denote comparisons with no significant differences. (B) Forest plots showing compositional changes in monocytes, CD4+ T cells, NK cells, and B cells between STB patients and UI patients. (C) Forest plots showing compositional changes in monocytes, CD4+ T cells, NK cells, and B cells between ATB and STB patients. The GEO or ArrayExpress identifiers represent individual cohorts. Each circle indicates the standardized mean difference (effect size, ES) in cellular composition between two groups, with the circle size proportional to the cohort sample size. The horizontal strip of each circle denotes the 95% confidence interval for the ES. The diamond represents the overall difference in cellular proportions between two groups, derived by integrating ES values from all cohorts (Summary). The FDR value indicates the statistical significance of the overall cellular compositional difference across cohorts.

Temporal changes in peripheral blood cell compositions during TB progression across global cohorts. (A) Bubble plot illustrating the compositional changes in six cell types among TB contacts at six intervals relative to TB diagnosis: (-inf, -540), [-540, -360), [-360, -180), [-180, -60), [-60, -21), and [-21, 0] days. Red bubbles indicate an increase in the cellular composition, whereas blue bubbles indicate a decrease. The intensity of the color reflects the degree of change, with a false discovery rate (FDR) threshold of 0.1 for statistical significance. The bubble size represents the absolute magnitude of the effect size. White circles denote comparisons with no significant differences. Forest plots highlight specific compositional changes: (B) monocytes in TBs contact between 360 and 180 days before diagnosis; (C) B cells and NK cells in TBs contact between 180 and 60 days before diagnosis; (D) monocytes, B cells, and NK cells in TBs contact 21 days before diagnosis. The GEO or ArrayExpress identifiers represent individual cohorts. Each circle indicates the standardized mean difference (effect size, ES) in cellular composition between two groups, with the circle size proportional to the cohort sample size. The horizontal strip of each circle denotes the 95% confidence interval for the ES. The diamond represents the overall difference in cellular proportions between two groups, derived by integrating ES values from all cohorts (Summary). The FDR value indicates the statistical significance of the overall cellular compositional difference across cohorts. Temporal compositions of monocytes, NK cells, and B cells in individual ATB progressors across six intervals before diagnosis in the (E) Zak_SA and (F) Suliman_GC6-74 cohorts. The gray shaded area represents the 95% confidence interval for the mean cellular compositions in ATB progressors.

Temporal changes in peripheral blood cell compositions during ATT across global cohorts. (A) Bubble plot showing compositional changes in six cell types in ATB patients across four intervals relative to ATT: [7, 28], (28, 60], (60, 182], and (182, 360] days. Red bubbles indicate an increase in the cellular composition, whereas blue bubbles indicate a decrease. The intensity of the color reflects the degree of change, with a false discovery rate (FDR) threshold of 0.1 for statistical significance. The bubble size represents the absolute magnitude of the effect size. White circles denote comparisons with no significant differences. Forest plots illustrate specific compositional changes in monocytes, NK cells, and B cells (B) between 7 and 28 days after ATT and (C) between 60 and 182 days after ATT. The GEO or ArrayExpress identifiers represent individual cohorts. Each circle indicates the standardized mean difference (effect size, ES) in cellular composition between two groups, with the circle size proportional to the cohort sample size. The horizontal strip of each circle denotes the 95% confidence interval for the ES. The diamond represents the overall difference in cellular proportions between two groups, derived by integrating ES values from all cohorts (Summary). The FDR value indicates the statistical significance of the overall cellular compositional difference across cohorts. (D) Bubble plot showing changes in six cell types across three intervals relative to ATT: (0, 7], (14, 30], and (120, 180] days in the Thompson_SA cohort. ATB patients in this cohort were grouped by treatment outcome into cured, probable cured, and not cured subgroups. (E) Bubble plot showing changes in six cell types across seven intervals relative to ATT: (0, 7], [7, 60), (60, 120], (120, 180], (180, 240], (240, 360], and (360, inf) days in the Tabone_Leicester cohort. ATB patients in this cohort were categorized into five subgroups on the basis of clinical information related to ATT: standard ATT, extended ATT, difficult TB cases, TB drug-resistant, and outbreak TB strain subgroups. Bubble plots of the first three subgroups (standard ATT, extended ATT, and difficult TB cases) are shown. Red bubbles indicate an increase in the cellular composition, whereas blue bubbles indicate a decrease. The intensity of the color reflects the degree of change, with a FDR threshold of 0.1 for statistical significance. The bubble size represents the absolute magnitude of the effect size. White circles denote comparisons with no significant differences.

Impact of confounding factors on blood cellular changes during TB development. (A) Bar plots displaying the compositional changes in monocytes, NK cells, and B cells between TB high- and low-burden countries. # indicates a significant compositional change in multi-cohort meta-analysis with an FDR < 0.1. The error bars represent the 95% confidence intervals of the effect size (ES) values. The statistical significance of compositional differences between two subgroups was assessed via two-tailed unpaired t tests with Benjamini–Hochberg correction. The significance thresholds are denoted as follows: ns (not significant); P-value > 0.1; *: P-value < 0.1; **: P-value < 0.01; and ***: P-value < 0.001. (B) Bar plots illustrating the compositional changes in monocytes, neutrophils, and NK cells across different age groups, including adolescents, adults and seniors. The x-axis represents TB disease stages, and the y-axis denotes cohort-summary ESs.

The trajectories of peripheral blood cellular changes throughout TB development and treatment.
Together, these analyses indicate that local TB burden and age significantly influence TB-associated shifts in peripheral blood cell composition, whereas HIV coinfection could not be fully assessed due to limited data. In general, individuals from low-burden regions and adults displayed the most pronounced cellular changes, while adolescents and seniors exhibited weaker responses. These findings highlight the need to account for demographic and epidemiological context when interpreting blood-based biomarkers of TB progression.
Discussion
In this study, we performed a comprehensive analysis of cellular compositional changes in the peripheral blood of individuals with diverse TB-related clinical phenotypes. Building upon a prior multi-cohort study of immune correlates of Mtb infection17, we reanalyzed 5,902 peripheral blood samples from 43 publicly available transcriptomic datasets. A key advance of our work was the reclassification of these samples using a recently proposed framework that captures the full spectrum of TB disease38. This approach allowed us to subdivide asymptotic LTBI individuals into three biologically distinct states - LTBI NP, ITB, and STB, thus refining the characterization of early disease heterogeneity21. Our analysis also incorporated post-treatment samples, enabling us to examine immune dynamics before and after ATT. By incorporating the largest number of cohorts and samples and systematically accounting for potential confounders such as local TB burden, age, and HIV coinfection, we provide, to our knowledge, the most extensive multi-cohort investigation of cellular dynamics in TB to date.
We found that trajectories of peripheral immune cell changes emerge early, beginning during LTBI stage. The earliest signal was an increase in proportion of monocytes, followed by broader alterations as individuals progressed toward active disease (Figs. 2 and 7). The transition to ATB was marked by widespread lymphopenia and expansion of myeloid lineages, changes that were reversible following successful ATT (Figs. 5 and 7). These findings are consistent with prior reports5,8,11,13,14,15,16,17,39,40,41 and reinforce the dynamic nature of immune remodeling across the TB spectrum. Specifically, circulating monocytes, particularly CD16+ subsets, expanded with disease progression and correlated with severity42. These nonclassical monocytes play key roles in antigen presentation, T-cell stimulation, and macrophage differentiation. Neutrophils also demonstrated a dual role: protective during early infection but pathogenic during established ATB43, where elevated proportions were associated with poor clinical outcomes and delayed culture conversion44,45. Conversely, lymphocyte subsets, including IFN-γ-producing CD4+ and CD8+ T cells46, B cells47, and NK cells48, declined with advancing disease, underscoring their importance in protective immunity46,47,48. Our multi-cohort design also revealed nuanced findings not consistently reported in smaller studies. For example, while Roy Chowdhury et al. described NK cell expansion in LTBI17, we observed a trend toward decreased NK proportions in this group, though not statistically significant (Fig. 2A). This discrepancy likely reflects heterogeneity in the proportion of incipient and subclinical cases within LTBI cohorts17, as well as inter-cohort variability. Indeed, while seven of fourteen individual cohorts showed increased or near-increased NK proportions, the aggregate analysis indicated an overall decline (Fig. 2B). Such results highlight the value, and the challenge, of integrating diverse datasets, where cohort heterogeneity both enriches generalizability and complicates interpretation.
A central contribution of our work is the delineation of immune changes at asymptomatic stages of TB progression. We observed no significant compositional shifts in cell proportions in limited LTBI NP cohorts (Figs. 3 and 7). As the disease progresses to the incipient stage, a reduction in NK cells is observed, followed by a decrease in B cells during the subclinical stage. These cellular alterations are pivotal, as they mark the transition from latent infection to active disease. The earliest cellular change in TB progression is the reduction in NK cells during the ITB stage, which persists until TB diagnosis. Circulating NK cells exhibit enhanced cytotoxic activity and interact with peripheral B and T cells under inflammatory conditions to control Mtb infection17. The gradual depletion of NK cells, particularly CD3− CD7+ GZMB+ NK cells, is strongly associated with TB progression. This pattern has been observed in a Chinese cohort14 as well as in global cohorts spanning three continents17, further highlighting the significance of NK cell dynamics in TB progression. In contrast, changes in B lymphocytes occur relatively later, during the STB stage, and are marked by a significant decrease in blood B cells. This decrease has also been linked to human TB in previous studies12,35,49. The timeline of these cellular changes is consistent with the general pattern of immune responses, as adaptive immune changes often occur later than innate immune responses. Moreover, this pattern of compositional changes in circulating immune cells is recapitulated in the temporal analysis of blood cell compositions in global longitudinal cohorts (Fig. 4). The earliest compositional change in TB progression is an increase in monocytes one and a half years before diagnosis. A decrease in B cells and an increase in neutrophils subsequently emerge between six and two months before diagnosis, followed by a decline in NK cells 21 days prior to TB diagnosis. Combined with the analysis of TB progression stages over time, timepoints more than 180 days before diagnosis approximate subclinical stages at the population level, while substantial inter-individual heterogeneity remains. Taken together, our findings provide a comprehensive view of the cellular changes that occur throughout TB development, offering valuable insights into the dynamic interplay between innate and adaptive immune responses and their roles in TB progression.
Despite these consistent trends, substantial heterogeneity was evident across cohorts and individuals (Figs. 2, 3, 4 and 5; Supplementary Figures S5, S7-S13, S15, S16). This variability has also been noted in previous studies, such as differences in B-cell compositional changes across cohorts17 and varying transcriptional activities of TB signatures among individuals28. This heterogeneity may be attributed to the immune endotypes of TB patients, as TB is not a monomorphic disease50. The host response to Mtb infection involves two distinct molecular pathways and pathologies. Endotype A is characterized by increased expression of genes related to inflammation and immunity, whereas endotype B is characterized by heightened activity in metabolism and proliferation pathways. These two mechanisms may influence cellular compositional changes during TB progression differently, adding complexity to the observed heterogeneity. However, the precise correlations between these endotypes and the variability in cellular changes remain unclear and warrant further investigation to better understand the underlying mechanism driving these differences.
Cellular changes in circulating blood cells serve as valuable indicators of the disease state and patient prognosis. For example, Bogan et al. identified KLRD-1-expressing NK cells as potential biomarkers for influenza susceptibility51. In human TB, the monocyte-to-lymphocyte ratio (MLR) has been recognized as an effective biomarker for diagnosing TB and monitoring the efficacy of anti-TB therapy8. However, the most pressing challenge in TB diagnosis is the early identification of TB progressors during the asymptomatic stage52. STB patients, despite having no clinical symptoms, contribute significantly to Mtb transmission23. Differentiating ITB and STB patients from LTBI individuals and providing preventive treatment at early TB stages is the most effective strategy for TB prevention53. Unfortunately, many early-stage TB patients lack radiologic abnormalities or microbiologic evidence, complicating their identification. The blood transcriptome has been extensively investigated for TB diagnosis, leading to the discovery of several transcriptional signatures for early-stage TB detection33,34,35,36,39,54,55,56. For example, Zak et al. conducted a longitudinal study of healthy adolescents infected with Mtb in South Africa, prospectively collecting blood samples over two years and profiling their transcriptomes in this ACS cohort33. They identified a 16-gene signature capable of predicting individuals at risk of developing ATB up to 12 months preceding TB. However, a promising validation study revealed that these blood prognostic signatures meet WHO benchmark requirements only when they are applied 3–6 months before TB diagnosis31. When tested in people living with HIV prior to antiretroviral therapy, their performance did not exceed that of a simple C-reactive protein (CRP) test and fell short of WHO targets57. To improve their clinical utility, interferon-independent biomarkers need to be incorporated into these host-response signatures to increase their accuracy57. Our findings revealed a specific decrease in NK cells during the transition from LTBI NP to ITB, a change that is common across multiple global cohorts and independent of transcriptional pathway activities. This compositional change shows strong potential as a cellular biomarker to distinguish ATB progressors from non-progressors at an early stage, offering a promising tool for improving TB diagnosis and prevention. Although cell type proportions hold promise for distinguishing stages of TB progression, the development and validation of a robust and specific classifier will require additional infection-related datasets and further in-depth investigation.
This study also has several limitations. First, we restricted analyses to six major blood cell types, precluding assessment of rarer populations implicated in TB, such as the CD3− CD7+ GZMB+ NK subsets14 or NK-like CD8+ γδ T cells15. Integrating single-cell RNA-seq datasets in future work would enable higher-resolution deconvolution of blood mixtures26. Second, the number of cohorts and samples used in multicohort comparisons of certain subgroups, such as STB and ITB patients, is relatively small. This limitation arises when TB-related samples are further divided by age or HIV coinfection status, reducing the statistical power of subgroup analyses. Third, our meta-analysis was not supported by experimental validation with flow cytometry or CyTOF due to limited availability of such datasets. Nonetheless, the biological plausibility of the findings, consistency with prior studies, and observed reversibility of changes post-ATT strengthen confidence in the results. Future studies addressing these limitations could provide a more comprehensive understanding of TB progression and its associated immune dynamics.
In conclusion, our multi-cohort study maps the temporal and stage-specific evolution of immune cell composition across the TB spectrum. By integrating thousands of samples spanning diverse global populations, we identify early depletion of NK cells and later B-cell loss as hallmarks of asymptomatic progression, and we situate these events within a broader framework of immune remodeling. These findings not only deepen our understanding of TB immunopathogenesis but also highlight NK cell dynamics as a promising avenue for early biomarker development and targeted prevention strategies.
Data availability
The original data of all public datasets are freely accessible in the *GEO* or *ArrayExpress* databases, while the processed data are available in supplementary tables.
References
World Health Organization. Global Tuberculosis Report 2024. (2024).
World Health Organization. The End TB Strategy. (2015). https://www.who.int/publications/i/item/WHO-HTM-TB-2015.19
Uplekar, M. et al. WHO’s new end TB strategy. Lancet 385, 1799–1801. https://doi.org/10.1016/s0140-6736(15)60570-0 (2015).
Singhania, A., Wilkinson, R. J., Rodrigue, M., Haldar, P. & O’Garra, A. The value of transcriptomics in advancing knowledge of the immune response and diagnosis in tuberculosis. Nat. Immunol. 19, 1159–1168. https://doi.org/10.1038/s41590-018-0225-9 (2018).
Ernst, J. D. The immunological life cycle of tuberculosis. Nat. Rev. Immunol. 12, 581–591. https://doi.org/10.1038/nri3259 (2012).
Sia, J. K. & Rengarajan, J. Immunology of Mycobacterium tuberculosis infections. Microbiol. Spectr. 7. https://doi.org/10.1128/microbiolspec.GPP3-0022-2018 (2019).
Mayer-Barber, K. D. & Sher, A. Cytokine and lipid mediator networks in tuberculosis. Immunol. Rev. 264, 264–275. https://doi.org/10.1111/imr.12249 (2015).
Adane, T. et al. Accuracy of monocyte to lymphocyte ratio for tuberculosis diagnosis and its role in monitoring anti-tuberculosis treatment: systematic review and meta-analysis. Med. (Baltim). 101, e31539. https://doi.org/10.1097/MD.0000000000031539 (2022).
Kroidl, I. et al. Assessment of tuberculosis disease activity in people infected with Mycobacterium tuberculosis and living with HIV: A longitudinal cohort study. Eclinicalmedicine 49, 101470. https://doi.org/10.1016/j.eclinm.2022.101470 (2022).
Chaussabel, D., Pascual, V. & Banchereau, J. Assessing the human immune system through blood transcriptomics. BMC Biol. 8, 1–14. https://doi.org/10.1186/1741-7007-8-84 (2010).
Berry, M. P. R. et al. An interferon-inducible neutrophil-driven blood transcriptional signature in human tuberculosis. Nature 466, 973–977. https://doi.org/10.1038/nature09247 (2010).
Moreira-Teixeira, L. et al. Mouse transcriptome reveals potential signatures of protection and pathogenesis in human tuberculosis. Nat. Immunol. 21, 464–476. https://doi.org/10.1038/s41590-020-0610-z (2020).
Nathan, A. et al. Multimodally profiling memory T cells from a tuberculosis cohort identifies cell state associations with demographics, environment and disease. Nat. Immunol. 22, 781–793. https://doi.org/10.1038/s41590-021-00933-1 (2021).
Cai, Y. et al. Single-cell transcriptomics of blood reveals a natural killer cell subset depletion in tuberculosis. EBioMedicine 53, 102686. https://doi.org/10.1016/j.ebiom.2020.102686 (2020).
Roy Chowdhury, R. et al. NK-like CD8(+) gammadelta T cells are expanded in persistent Mycobacterium tuberculosis infection. Sci. Immunol. 8, eade3525. https://doi.org/10.1126/sciimmunol.ade3525 (2023).
Lyu, M. et al. Single-Cell sequencing reveals functional alterations in tuberculosis. Adv. Sci. (Weinh). 11, e2305592. https://doi.org/10.1002/advs.202305592 (2024).
Roy Chowdhury, R. et al. A multi-cohort study of the immune factors associated with M. tuberculosis infection outcomes. Nature 560, 644–648. https://doi.org/10.1038/s41586-018-0439-x (2018).
Kalesinskas, L., Gupta, S. & Khatri, P. Increasing reproducibility, robustness, and generalizability of biomarker selection from meta-analysis using bayesian methodology. PLoS Comput. Biol. 18, e1010260. https://doi.org/10.1371/journal.pcbi.1010260 (2022).
Sweeney, T. E., Haynes, W. A., Vallania, F., Ioannidis, J. P. & Khatri, P. Methods to increase reproducibility in differential gene expression via meta-analysis. Nucleic Acids Res. 45, e1. https://doi.org/10.1093/nar/gkw797 (2017).
Coussens, A. K. et al. Classification of early tuberculosis states to guide research for improved care and prevention: an international Delphi consensus exercise. Lancet Respir Med. 12, 484–498. https://doi.org/10.1016/S2213-2600(24)00028-6 (2024).
Drain, P. K. et al. Incipient and subclinical tuberculosis: a clinical review of early stages and progression of infection. Clin. Microbiol. Rev. 31, e00021–e00018. https://doi.org/10.1128/cmr.00021-18 (2018).
Stuck, L. et al. Prevalence of subclinical pulmonary tuberculosis in adults in community settings: an individual participant data meta-analysis. Lancet Infect. Dis. https://doi.org/10.1016/s1473-3099(24)00011-2 (2024).
Nguyen, H. V., Tiemersma, E., Nguyen, N. V., Nguyen, H. B. & Cobelens, F. Disease transmission by patients with subclinical tuberculosis. Clin. Infect. Dis. 76, 2000–2006. https://doi.org/10.1093/cid/ciad027 (2023).
Clough, E. et al. NCBI GEO: archive for gene expression and epigenomics data sets: 23-year update. Nucleic Acids Res. 52, D138–D144. https://doi.org/10.1093/nar/gkad965 (2024).
Sarkans, U. et al. From arrayexpress to biostudies. Nucleic Acids Res. 49, D1502–D1506. https://doi.org/10.1093/nar/gkaa1062 (2021).
Newman, A. M. et al. Determining cell type abundance and expression from bulk tissues with digital cytometry. Nat. Biotechnol. 1–10. https://doi.org/10.1038/s41587-019-0114-2 (2019).
Pai, M. et al. Tuberculosis. Nat. Rev. Dis. Primers. 2, 16076. https://doi.org/10.1038/nrdp.2016.76 (2016).
Tabone, O. et al. Blood transcriptomics reveal the evolution and resolution of the immune response in tuberculosis. J. Exp. Med. 218, e20210915. https://doi.org/10.1084/jem.20210915 (2021).
Zaidi, S. M. A. et al. Beyond latent and active tuberculosis: a scoping review of conceptual frameworks. Eclinicalmedicine 66, 102332. https://doi.org/10.1016/j.eclinm.2023.102332 (2023).
Tateosian, N. L., Morelli, M. P., Pellegrini, J. M. & Garcia, V. E. Beyond the clinic: the activation of diverse cellular and humoral factors shapes the immunological status of patients with active tuberculosis. Int. J. Mol. Sci. 24 https://doi.org/10.3390/ijms24055033 (2023).
Gupta, R. K. et al. Concise whole blood transcriptional signatures for incipient tuberculosis: a systematic review and patient-level pooled meta-analysis. Lancet Respir Med. 8, 395–406. https://doi.org/10.1016/s2213-2600(19)30282-6 (2020).
Haynes, W. A. et al. Empowering Multi-Cohort gene expression analysis to increase reproducibility. Pac. Symp. Biocomput. 22, 144–153. https://doi.org/10.1142/9789813207813_0015 (2017).
Zak, D. E. et al. A blood RNA signature for tuberculosis disease risk: a prospective cohort study. Lancet 387, 2312–2322. https://doi.org/10.1016/s0140-6736(15)01316-1 (2016).
Suliman, S. et al. Four-Gene Pan-African blood signature predicts progression to tuberculosis. Am. J. Respir Crit. Care Med. 197, 1198–1208. https://doi.org/10.1164/rccm.201711-2340oc (2018).
Singhania, A. et al. A modular transcriptional signature identifies phenotypic heterogeneity of human tuberculosis infection. Nat. Commun. 9, 2308. https://doi.org/10.1038/s41467-018-04579-w (2018).
Roe, J. et al. Blood transcriptomic stratification of short-term risk in contacts of tuberculosis. Clin. Infect. Dis. 70, ciz252. https://doi.org/10.1093/cid/ciz252 (2019).
Thompson, E. G. et al. Host blood RNA signatures predict the outcome of tuberculosis treatment. Tuberculosis 107, 48–58. https://doi.org/10.1016/j.tube.2017.08.004 (2017).
Coussens, A. K. et al. Classification of early tuberculosis states to guide research for improved care and prevention: an international Delphi consensus exercise. Lancet Respir Med. https://doi.org/10.1016/s2213-2600(24)00028-6 (2024).
Bloom, B. R. A half-century of research on tuberculosis: successes and challenges. J. Exp. Med. 220, e20230859. https://doi.org/10.1084/jem.20230859 (2023).
Kotov, D. I. et al. Early cellular mechanisms of type I interferon-driven susceptibility to tuberculosis. Cell 186, 5536–5553 e5522 (2023). https://doi.org/10.1016/j.cell.2023.11.002
Scriba, T. J. et al. Sequential inflammatory processes define human progression from M. tuberculosis infection to tuberculosis disease. PLoS Pathog. 13, e1006687. https://doi.org/10.1371/journal.ppat.1006687 (2017).
Sampath, P., Moideen, K., Ranganathan, U. D. & Bethunaickan, R. Monocyte subsets: phenotypes and function in tuberculosis infection. Front. Immunol. 9, 1726. https://doi.org/10.3389/fimmu.2018.01726 (2018).
Lowe, D. M., Redford, P. S., Wilkinson, R. J., O’Garra, A. & Martineau, A. R. Neutrophils in tuberculosis: friend or foe? Trends Immunol. 33, 14–25. https://doi.org/10.1016/j.it.2011.10.003 (2012).
Carvalho, A. C. C. et al. Pre-Treatment neutrophil count as a predictor of antituberculosis therapy outcomes: A multicenter prospective cohort study. Front. Immunol. 12, 661934. https://doi.org/10.3389/fimmu.2021.661934 (2021).
Ndlovu, L. N. et al. Increased neutrophil count and decreased neutrophil CD15 expression correlate with TB disease severity and treatment response irrespective of HIV Co-infection. Front. Immunol. 11, 1872. https://doi.org/10.3389/fimmu.2020.01872 (2020).
Musvosvi, M. et al. T-cell biomarkers for diagnosis of tuberculosis: Candidate evaluation by a simple whole blood assay for clinical translation. Eur. Respir J. 51. https://doi.org/10.1183/13993003.00153-2018 (2018).
Joosten, S. A. et al. Patients with tuberculosis have a dysfunctional circulating B-Cell Compartment, which normalizes following successful treatment. PLoS Pathog. 12, e1005687. https://doi.org/10.1371/journal.ppat.1005687 (2016).
Abebe, F. Immunological basis of early clearance of Mycobacterium tuberculosis infection: the role of natural killer cells. Clin. Exp. Immunol. 204, 32–40. https://doi.org/10.1111/cei.13565 (2021).
Krause, R. et al. B cell heterogeneity in human tuberculosis highlights compartment-specific phenotype and functional roles. Commun. Biol. 7, 584. https://doi.org/10.1038/s42003-024-06282-7 (2024).
DiNardo, A. R. et al. Gene expression signatures identify biologically and clinically distinct tuberculosis endotypes. Eur. Respir J. 60, 2102263. https://doi.org/10.1183/13993003.02263-2021 (2022).
Bongen, E., Vallania, F., Utz, P. J. & Khatri, P. KLRD1-expressing natural killer cells predict influenza susceptibility. Genome Med. 10, 45. https://doi.org/10.1186/s13073-018-0554-1 (2018).
World Health Organization. Consensus Meeting Report - Development of a Target Product Profile (TPP) and a Framework for Evaluation for a Test for Predicting Progression from Tuberculosis Infection to Active Disease. (2017). https://www.who.int/publications/i/item/WHO-HTM-TB-2017.18
Horton, K. C., Richards, A. S., Emery, J. C., Esmail, H. & Houben, R. M. G. J. Reevaluating progression and pathways following Mycobacterium tuberculosis infection within the spectrum of tuberculosis. Proc. Natl. Acad. Sci. 120, e2221186120. https://doi.org/10.1073/pnas.2221186120 (2023).
Leong, S. et al. Cross-validation of existing signatures and derivation of a novel 29-gene transcriptomic signature predictive of progression to TB in a Brazilian cohort of household contacts of pulmonary TB. Tuberculosis 120, 101898. https://doi.org/10.1016/j.tube.2020.101898 (2020).
Sivakumaran, D. et al. Host blood-based biosignatures for subclinical TB and incipient TB: A prospective study of adult TB household contacts in Southern India. Front. Immunol. 13, 1051963. https://doi.org/10.3389/fimmu.2022.1051963 (2023).
Esmail, H., Cobelens, F. & Goletti, D. Transcriptional biomarkers for predicting development of tuberculosis: progress and clinical considerations. Eur. Respir J. 55, 1901957. https://doi.org/10.1183/13993003.01957-2019 (2020).
Mann, T. et al. Blood RNA biomarkers for tuberculosis screening in people living with HIV before antiretroviral therapy initiation: a diagnostic accuracy study. Lancet Glob Heal. https://doi.org/10.1016/s2214-109x(24)00029-9 (2024).
Funding
This work was supported by the National Key R&D Program of China (Nos. 2022YFC3500200, 2022YFC3500202, 2023YFC2307300), the National Natural Science Foundation of China (Nos. 81930117, 82430122), the Jiangsu Provincial Social Development and Clinical Frontier Technology Project (BE2023790), and the Respiratory Infectious Diseases Clinical Medical Center of Suzhou (No. Szlcyxzx202108).
Author information
Authors and Affiliations
Contributions
W.G., M.W., and X.X. conceived the study. S.Z., H.Y., Q.D., K.W., Y.M., and Y.N. contributed to the data analysis. W.G., M.W., X.X., and C.C. interpreted the results, and S.Z. and W.G. wrote the manuscript. All the authors read, revised and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhu, S., Dai, Q., Wang, K. et al. Peripheral blood cellular dynamics during the progression of human tuberculosis. Sci Rep 15, 37420 (2025). https://doi.org/10.1038/s41598-025-21181-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-21181-5
