
Abstract
Mice possess hands and feet similar to humans including two phalanges for digit 1 and three phalanges in digits 2 to 5. The phalanges that make up the digits were named by their positions, however, phalanx identities between digits are unconfirmed by genetic evidence. Here we show that the proximal phalanges of digit 1 of the fore- and hindlimbs have different identities compared to the phalanges of digit 2. We found strong signals from mRNA-Seq results that unite the proximal phalanx of forelimb digit 1 with the proximal phalanx of forelimb digit 2, while the proximal phalanx of hindlimb digit 1 groups the middle phalanx of hindlimb digit 2. The same result was obtained when other clustering methods were used, indicating that our results are robust. Our results demonstrate that there are developmental differences among phalanges and that the proximal phalanx identities of digit 1 differ between the fore- and hindlimbs of the mouse.
Similar content being viewed by others

Introduction
The limbs of tetrapods have differing morphologies to satisfy their unique and specific functional requirements1,2. However, mice and humans have similar digital morphologies and the same phalangeal formula, with two phalanges in digit 1, and three phalanges in digits 2 to 5, thus mice are good models for understanding human phalanx development3. In the past few decades, the major genes, and their regulatory components, which establish the morphology and development of limbs have been identified, including how signaling explains digit identities4,5,6,7,8. However, little is known about phalanx identities.
In the mouse fore- and hindlimbs, digits 2 to 5 have similar anatomy with similar shape, size and proportion of phalanges at corresponding positions from proximal to distal ends, while digit 1 has a different anatomy with only two phalanges and different shape of proximal phalanges9. Morphology of digits reflect the differential expression of genes involved in digit formation during development, such as the expression of genes controlling cell proliferation and differentiation, and the expression of transcription factors and signaling genes that regulate gene expression10. Thus, the morphology of phalanges may also reflect the expression of genes affecting the formation of phalanges during development, and the phalanges may have different developmental programs (developmental identities). In this study, gene expression similarities are being interpreted as indicators of developmental modularity, and we defined phalanx identity according to the gene expression profile of phalanges.
Here we used mRNA sequencing (mRNA-Seq) to study phalanx identities by comparing transcriptomes generated from the phalanges of digits 1 and 2 from the mouse. We determined the developmental period for these phalanges, and then generated and analyzed transcriptome data from the relevant period to examine phalanx identities of digits 1 and 2 from the fore- and hindlimbs. mRNA-Seq technology enables an assessment of quantitative gene expression differences between samples or tissue, and thus, a homology assessment can be based on gene expression similarities and used to address questions about digit identity11. For example, the transcriptome data of digits from fore- and hindlimb has been used to determine digit identities in birds12.
Results and discussion
According to a continuous series of skeletal staining of mouse embryonic limbs from E11.25 to E19, we found that cartilage condensation of the phalanges successively occurs from proximal to distal positions at stages E12.75 - E14.75 (Fig. 1), and that all phalanges finished cartilage condensation at stages E16 - E17 (Fig. 2a). After E17, ossification centers began to appear in the phalanges (Figure S1). The skeletal staining also showed that the distal phalanges of digit 1 to 5 are similarly triangular in the fore- and hindlimbs. The middle and proximal phalanges of digit 2 to 5 are similarly isosceles trapezoidal and have similar shape, size and proportions at the corresponding positions in each stage (Fig. 2a). Although the shapes of the proximal phalanges of digit 1 in the fore- and hindlimbs are both similar to isosceles trapezoidal, the proportions of the height to the bottom base were distinct different, i.e. the proximal phalanx of digit 1 in the forelimb was much shorter and wider than that in the hindlimb (Fig. 2a). The micro-computed tomography (micro-CT) of the middle and proximal phalanges of digits 1 to 2 of an adult mouse obtained detailed and three-dimensional structures of the phalanges (Supplementary video S1-6). Consistent with the results of skeletal staining at embryonic stages, micro-CT showed the proximal phalanges of digit 1 in the forelimb was obviously different from other phalanges, because it was wider and flatter than others. However, other phalanges were similarly slender and difficult to distinguish their identities only by the morphological structure.

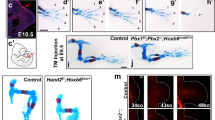
Skeletal staining from stage E12.75 to E14.75 of mouse limbs. Dorsal view of the limb is shown with the anterior up and distal to the left. (a1-e1) Skeletal staining of forelimbs. (a2-e2) Skeletal staining of hindlimbs. Red arrows indicate the position where the proximal phalanges appear, black arrows indicate the position where the middle phalanges appear, and blue arrows indicate the position where the distal phalanges appear. E, embryonic day. Bar = 1 mm.

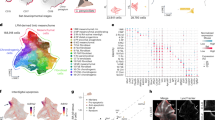
mRNA-Seq sampling and analysis of mouse phalanges. (a) Alcian-blue-stained autopods. Dorsal view of the autopod is shown with the anterior to the left and distal to the top. Red dashed lines indicate sections dissected for mRNA-Seq of the phalangeal segments. Samples for RNA-Seq were labeled E or L for stage E16 or E17, respectively, F and H for fore- and hindlimb, respectively, 1 and 2, for the distal and proximal phalanges of digit 1, respectively, and 3, 4, and 5, for the distal, middle, and proximal phalanges of digit 2, respectively. Bar = 1 mm. (b and d) Cluster analysis and heatmap of the Pearson’s correlations using all expressed genes at E16 (b) and E17 (d). To facilitate the differentiation of clusters, black indicates the distal phalanges, red indicates the proximal phalanges of forelimb digit 1 and the proximal phalanges of digit 2, and blue indicates the proximal phalanges of hindlimb digit 1 and the middle phalanges of digit 2. Red numbers at the nodes represent AU P values. (c and e) PCA plot of the 10 samples using all expressed genes at E16 (c) and E17 (e). PC scores are plotted on PC1 and PC2.
The developmental heterochrony of mouse limbs exists, i.e. mouse forelimb develops about 12 h sooner than the hindlimb before E1513,14. However, this heterochrony gradually reduced in later stages. In this study, we found that ossification centers occurred in two metacarpals (forelimb digits 3–4) and three metatarsals (hindlimb digits 2–4) at E16.75 (Figure S1). At E18.25, ossification centers appeared in the middle phalanges of forelimb digits 3–4 but not in the middle phalanx of the hindlimb. At E18.5, ossification centers took place in the proximal phalanx of the hindlimb digits 1 but not in the proximal phalanx of forelimb digits 1. Digit primordia initiated in the order of digits 4-3-2-5-1 (Figure S1). It is necessary to select developmental stages for mRNA-Seq when cartilage condensations of the two phalanges have just occurred or completed in digit 1 to reduce the effects of developmental heterochrony of mouse limbs.
Based on the above skeletal results and considerations, we performed mRNA-Seq on 20 phalanx samples from digits 1 and 2 of the fore- and hindlimbs at stages E16 and E17 (Fig. 2a) to address the question of mouse phalanx identity. A total of 27,527 and 28,766 expressed genes were detected from samples collected at E16 and E17, respectively. Hierarchical clustering, PCA analysis and K-means clustering of the mRNA-Seq data from the phalanges of digits 1 and 2 yielded two distinct clusters (Fig. 2b-e and Figure S2). The gene expression patterns from the distal phalanges of digits 1 and 2 of the fore- and hindlimbs at both developmental stages clustered together, while the patterns from the other phalanges formed a separate cluster. These results indicate that the distal phalanges of digits 1 and 2 have the same identity. Surprisingly, the gene expression patterns of the proximal phalanges of digit 1 in the fore- and hindlimbs clustered with different phalanges of digit 2 (Fig. 2b, d). At the early stage of phalangeal formation (E16), the proximal phalanx of forelimb digit 1 (EF2) clustered with both the middle and proximal phalanges of forelimb digit 2 (EF4 and EF5), with the identities of the two digit 2 phalanges not yet distinguishable. But in the hindlimb, the proximal phalanx of digit 1 (EH2) clustered with the middle phalanx of digit 2 (EH4) with a 95% approximately unbiased (AU) P value (Fig. 2b). At the later stage of phalangeal formation (E17), the proximal phalanx of forelimb digit 1 (LF2) clustered with the proximal phalanges of digit 2 of both the fore- and hindlimbs (LF5 and LH5) with a greater than 98% AU P value, while the proximal phalanx of hindlimb digit 1 (LH2) clustered with the middle phalanges of digit 2 in both the fore- and hindlimbs (LF4 and LH4) with a 99% AU P value (Fig. 2d). The AU P values were calculated via multiscale bootstrap resampling, and strongly support clustering when they are not less than 95%15. PCA results showed that the proximal phalanx of forelimb digit 1 (E/LF2) were closer to the proximal phalanx of digit 2 (E/LF5 and E/LH5) than to the middle phalanx of digit 2 (E/LF4 and E/LH4), and the proximal phalanx of hindlimb digit 1 (E/LH2) was closer to the middle phalanx of digit 2 (E/LF4 and E/LH4) than to the proximal phalanx of digit 2 (E/LF5 and E/LH5) (Fig. 2c, e). The slight segregation of the proximal phalanx in forelimb digit 1 (E/LF2) observed in PCA results may reflect its unique skeletal morphology (see above results related to Fig. 2a and Supplementary video S1-6). All these results indicate that the proximal phalanx of forelimb digit 1 has the same identity with the proximal phalanges of digit 2, and the proximal phalanx of hindlimb digit 1 has the same identity with the middle phalanges of digit 2.
To gain more insight in to the developmental differences between the proximal phalanges of forelimb digit 1 (F2) and hindlimb digit 1 (H2), we identified 755 differentially expressed genes (DEGs) between these two phalanges using standard statistical thresholds (Q-value < 0.01 and |logFC| > 1) (Figure S3 and Table S1). Among the 755 DEGs, 9 genes related to development (Dlk1, Itm2a, Rian, Angptl1, Ccn3, Ssc5d, Has2, Galnt5 and Bcat1) were selected for expression validation by Reverse-Transcription Quantitative PCR (RT-qPCR) analysis (Table S1). RT-qPCR results for these 9 genes were in accordance with the mRNA-Seq data and confirmed the accuracy of mRNA-Seq results (Fig. 3). Shh, associated with digit identity, was included in the 755 DEGs, but not members of the BMP or Smad gene families, which are major regulators of cartilage formation16,17. This may possibly be due to the mRNA-Seq samples being collected at the differentiation phase. Based on chick embryos, digit identities are likely start to form at an earlier stage prior to cartilage condensation, and interdigit signals can regulate the number of phalanges formed18,19. However, digit identities will not be fixed and irreversible until the formation of phalanges is complete5,20. Therefore, we chose E16 (cartilage condensation just occurred in the distal phalanges of hindlimb, while the condensation of other phalanges had taken shape initially) and E17 (all phalanges just finished cartilage condensation) as sampling time points. Although gene expression profiles of the two time points may reflect secondary differentiation rather than primary identity, final phalanx identities should be determined by this study.

RT-qPCR validation of the transcriptome data for nine genes at E17.
Of the 755 DEGs, 54 encode transcription factors, and 39 are associated with muscle or skeletal development or are involved in embryonic organ morphogenesis (Figure S4 and Table S1). We also identified many genes associated with neural pathways (Figure S4 and Table S1), which may be related to functions of the proximal phalanx of digit 1. These findings suggest that the gene expression differences between the proximal phalanges of digit 1 in the fore- and hindlimbs may correspond to both phalanx morphology and functional specializations. In the mouse, the forelimbs are involved in many dexterous manipulations such as manipulating small objects and eating food, which is achieved by the flexible use of the thumb, whereas the hindlimb is primarily used for support and locomotion21,22. Similar functional distinctions have been reported in humans and primates, which the dexterous thumb is associated with specific developmental programs23,24. Furthermore, the divergence of phalanx identities of mouse digit 1 corresponds to the independent evolution of digit 1 in tetrapods. The evolution of an opposable thumb is the result of important adaptations for arboreal tetrapod grasping/clutching8. In humans, the thumb shows distinctive morphological and functional divergence from other digits, which has been proposed to result from early developmental modulation and selective pressures favoring manipulation abilities24. Based on the above researches, we speculate that there are functional differences between digit 1 in mouse fore- and hindlimbs. The results show that there is little developmental difference between the distal phalanges of mouse digits 1 and 2, therefore, the functional difference between the thumb and the big toe may be influenced by the proximal phalanges of digit 1.
Although the presence of neural pathway-related DEGs points toward a functional difference in the proximal phalanges of digit 1 of the fore- and hindlimbs, it could also be the result of nerve tissue within the surrounding mesenchyme that was included in the dissection for the digit 1 proximal phalanx. Further histological studies will help confirm whether these neural pathway-related DEGs are related to the functional specialization. In this study, two sampling time points were selected. Little heterochrony was observed in the fore- and hindlimbs at E17, except for the proximal phalanges of digit 1. Although the corresponding phalanges of the fore- and hindlimbs clustered closely in Fig. 2e, it remains unclear whether the differences in the proximal phalanges of digit 1 reflect an enduring difference in segment identity or just a continued signal of heterochrony related to cartilage development and ossification timing. To determine whether there is an enduring difference in segment identity, additional sampling time points—such as during ossification, at birth, and in adulthood—are needed to investigate whether phalangeal identity remains constant. In addition, due to the irregular morphology and subjective nature of assessing the phalanges, and because we sequenced the polygonal skeletons collectively, it is impossible to determine which specific gene regulates the local morphology. In future research, single-cell sequencing that targets specific phalangeal morphologies could potentially bridge this gap by effectively linking morphological data with gene expression profiles.
In conclusion, we show that the proximal phalanx identities of digit 1 of the fore- and hindlimbs differ in the mouse, and that developmental differences exist for the proximal phalanx of digit 1 of the fore- and hindlimbs. Transcriptome similarity likely reflects morphological and functional similarity, but why and how differences in transcriptomes regulate morphology and function remain to be studied. It would also be of interest to compare human and mouse phalanges and determine if evolutionary differences exist in future experiments.
Materials and methods
Ethical use of animals
All animal experiments used in this study were approved by the Institutional Animal Care and Use Committee of Shenyang Agricultural University (Permit # 2022030706), and all procedures were conducted in accordance with the University’s Regulations for Animal Experiments.
Mouse husbandry and embryo collection
All mice (Mus musculus) used in this study were from Changsheng Biotechnology Co. LTD (Benxi City, China). ICR mice are highly fertile and the commonly used reference genome of mice is derived from C57BL/6 mice. To reduce the bias of mRNA composition caused by different breeds, better align sequencing files to the reference genome, and verify the reliability of transcriptome data better, embryos for serial skeletal staining and Reverse-Transcription Quantitative PCR (RT-qPCR) were ICR mice, and embryos for skeletal staining and mRNA-Seq at E16 and E17 stages were C57BL/6 mice. Mice were raised in a clean environment at 22 to 24℃ and 45% to 55% relative humidity, with an adequate supply of food and water and a 12 h light and 12 h dark cycle (7 pm − 7 am).
Male and female mice were placed together at 7 pm, and the precise time of mating was obtained via monitoring with a Xiaomi smart infrared camera (MJSXJ06CM, Xiaomi Corporation, Shanghai, China) in real time. Mating time was designated as day 0 of embryonic development (E0). Embryos were collected every 6 h from E8 to parturition. We excluded too large, too small and abnormal embryos, and determined the stages according to previous studies14,25,26. A total of 957 embryos was used in this study, at least 20 embryos for each development stage were used to determine morphological consistency.
Skeletal staining
For embryos at stages E11-14.75, E16 and E17, whole-mount skeletal staining was conducted according to the procedure of Nagy et al. (2009)27. Embryos were stained with Alcian blue stain (5% acetic acid, 0.03% blue in 80% ethanol) for 2 h. For embryos from stages E13 to E19, the procedure of Rigueur and Lyons (2014) was improved to perform whole-mount skeletal staining28. Here, we detail the steps that have been improved and omit the description of steps that are the same between our procedure and that of Rigueur and Lyons (2014)28. For 3.1.2 and 3.2.2 in the procedure of Rigueur and Lyons, fresh embryos were scalded in 61–65℃ water for 10–38 s, which was followed by the removal of the eyes, skin, and internal organs (except for E13-14.75). For 3.1.6 and 3.2.5 to 3.2.7 in the procedure of Rigueur and Lyons, embryos were covered with Alcian blue stain (20% acetic acid, 0.015%−0.03% blue in 80% ethanol) for approximately 2–14 h (depending upon the degree of embryonic development) and then washed 3 times with 20% acetic acid/80% ethanol for a total of 1 h. Embryos were transferred and kept in a solution of 75% ethanol in 0.05%−1% KOH for 2 h and then hydrated by successive steps of 10 min in 50% and 25% ethanol in 0.05%−1% KOH, followed by two 10 min hydration steps in 0.035%−0.5% KOH. For 3.1.7 to 3.1.8 and 3.2.8 in the procedure of Rigueur and Lyons, the embryos were then soaked in Alizarin red stain (0.035%−0.5% KOH, 0.001%−0.002% Alizarin red in water) for 2–16 h followed by soaking in 0.05%−1% KOH for ~ 24 h to clean the embryos.
Micro-computed tomography (micro-CT)
The middle and proximal phalanges of digits 1 to 2 of an adult mouse (C57BL/6, 7w, male) were imaged with a Quantum GX2 micro-CT (PerkinElmer, Waltham, Massachusetts, USA) using the following parameters: 90 kV, 80 µA, 4 min integration time, resolution 36 μm isotropic voxels.
mRNA-Seq
In this study, we determined phalanx identities by transcriptome data rather than the developmental morphology or positions of phalanges. To obtain enough RNA for transcriptome sequencing, phalanges and their surrounding mesenchyme were collected from the left and right limbs of 20 individuals from E16 and 10 individuals from E17 for each sample. Only one pooled sample was sequenced for each phalanx at each stage. Samples (E/LF1-5 and E/LH1-5) were collected as shown in Fig. 2a. These specimens were treated with 10 µl 2 M dithiothreitol (redox reagent, Sangon, Shanghai, China) and 500 µl Buffer RLT (lysis solution, Qiagen, Hilden, Germany). Total RNA was extracted using the RNeasy Mini kit (Qiagen, Hilden, Germany) and RNeasy Micro kit (Qiagen, Hilden, Germany) according to tissue amounts and treated with DNase I using RNase-Free DNase set (Qiagen, Hilden, Germany). The total RNA concentration of all samples was greater than 50 ng/µL, the total RNA quantity was greater than 0.3 µg, the OD 260/280 and OD 260/230 was greater than 1.8 and 0.5 respectively, the RNA integrity number (RIN) was between 9.6 and 10.0, and 28 S/18S was greater than 1.0. Extracted total RNA from the 20 samples was submitted to Novogene Bioinformatics Technology Co., Ltd (Beijing, China) for mRNA-Seq using the PE150 strategy on the Illumina platform. More than 20 million reads were generated per sample. Then the sequencing data were aligned to the reference genome GRCm39 using HISAT2, and transcripts were spliced with StringTie and expression was calculated29.
Clustering and PCA analysis
Normalized Trimmed Mean of M-values (TMM) were calculated from the raw data by the edgeR package in R (version 3.6.1) and were used for the following analyses30,31. Hierarchical clustering with bootstrap analyses (n = 10,000) was done using the pvclust package in R (version 3.6.1)15. Pearson correlation coefficient was calculated through the R (version 3.6.1) Lattice package, then using their Pearson correlation coefficients and TBtools-II software (version 2.042) a heatmap was generated32. The optimal number of clusters (k-value) was determined using the Elbow method, where k appears at an elbow-like turning point33. K-means clustering and two-dimension PCA analysis was performed with the OmicStudio tools at the website (http://www.omicstudio.cn). The parameter for two-dimension PCA analysis was “prcomp(data, center = FALSE, scale = TRUE)”.
Analyses of gene differential expression and gene ontology (GO)
The edgeR package was used to identify differentially expressed genes (DEGs) that clustered the different phalanges of the different limbs (proximal phalanx of forelimb digit 1 (E/LF2) with proximal phalanx of forelimb digit 2 (E/LF5) and proximal phalanx of hindlimb digit 1 (E/LH2) with middle phalanx of hindlimb digit 2 (E/LH4)31. Q-value < 0.01 and |logFC| > 1 was considered as thresholds for significant differential expression. DEGs include group A genes that were not differently expressed between proximal phalanx of forelimb digit 1 (E/LF2) and proximal phalanx of forelimb digit 2 (E/LF5), but differently between proximal phalanx of forelimb digit 1 (E/LF2) and middle phalanx of forelimb digit 2 (E/LF4) and differently between proximal phalanx of forelimb digit 2 (E/LF5) and middle phalanx of forelimb digit 2 (E/LF4). They also include group B genes that were not differently expressed between proximal phalanx of hindlimb digit 1 (E/LH2) and middle phalanx of hindlimb digit 2 (E/LH4), but differently between proximal phalanx of hindlimb digit 1 (E/LH2) and proximal phalanx of hindlimb digit 2 (E/LH5) and differently between middle phalanx of hindlimb digit 2 (E/LH4) and proximal phalanx of hindlimb digit 2 (E/LH5). We then removed the overlapping genes of groups A and B and obtained 755 DEGs for GO analysis. GO annotation for the DEGs was completed using a R (version 3.3.1) software package34. Significantly enriched GO categories were limited to corrected P < 0.05.
Reverse-transcription quantitative PCR (RT-qPCR)
From the 755 DEGs, we retained those with TMM normalized counts greater than 5 in at least two samples, and 9 genes related to development were selected from the remaining top 50 genes for RT-qPCR analysis. To obtain enough RNA for RT-qPCR, samples containing the phalanx and its surrounding mesenchyme were pooled from the left and right limbs of 20 individuals at E17. Only one pooled sample was obtained for each phalanx. Total RNA was extracted as described in the previous mRNA-Seq section. RNA was reverse transcribed into cDNA with PrimeScript FAST RT reagent Kit with gDNA Eraser (TaKaRa, Tokyo, Japan). cDNA products were then amplified using TB Green Premix (TaKaRa, Tokyo, Japan) and the following specific primers. RT-qPCR reactions were conducted in technical triplicate using a LightCycler 96 Instrument (Roche, Basel, Switzerland). mRNA expression obtained from the transcriptomic analysis (orange) was compared to the mean ± SD of gene expression levels quantified by RT-qPCR (blue) analysis (Fig. 3). Primers were synthesized by Sangon (Shanghai, China). The specificity of the amplification products was confirmed by Sanger sequencing at Sangon (Shanghai, China). Primer sequences are listed below: Dlk1, forward sequence (5’−3’): AAGAAGTCCCTTGAAACCC; reverse sequence (5’−3’): AAAGGTCGTCGTCTGAGTTC. Itm2a, forward sequence (5’−3’): TCGCCTTCAACACCCCT; reverse sequence (5’−3’): CCTGCCAAGATGAATGAGAG. Rian, forward sequence (5’−3’): CATGGCTCAGACTTCCAAT; reverse sequence (5’−3’): AGACCCAGCGAAAATTCA. Angptl1, forward sequence (5’−3’): GTCAACACCAAAGGTCAGG; reverse sequence (5’−3’): TCGTTTACAATGTTCCCGT. Ccn3, forward sequence (5’−3’): AAAGTGGACCTGTGGCTC; reverse sequence (5’−3’): TCTCACACTGGCGATTCC. Ssc5d, forward sequence (5’−3’): TCCCCCAAACCATCTCTACTCA; reverse sequence (5’−3’): CGTGTTGGAAGCAAACTGGTATT. Has2, forward sequence (5’−3’): AAGAAGTCCCTTGAAACCC; reverse sequence (5’−3’): AAAGGTCGTCGTCTGAGTTC. Galnt5, forward sequence (5’−3’): AAGTTCGGATTCTTCGCC; reverse sequence (5’−3’): CACATCACCTGTTGCGTTCT. Bcat1, forward sequence (5’−3’): CTCCCTCACCCTGTACCA; reverse sequence (5’−3’): CAGTAGCCCTTCCAGCAC. Reference gene ACTB, forward sequence (5’−3’): TGAGAGGGAAATCGTGCG; reverse sequence (5’−3’): CAAGAAGGAAGGCTGGAAAA.
Data availability
Datasets generated during the current study are available in the GEO database at https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE264526.
References
Kavanagh, K. D. et al. Developmental Bias in the Evolution of Phalanges. Proc. Natl. Acad. Sci. 110, 18190–18195, (2013). https://doi.org/10.1073/pnas.1315213110
Young, J. J. & Tabin, C. J. Saunders’s framework for Understanding limb development as a platform for investigating limb evolution. Dev. Biol. 429, 401–408. https://doi.org/10.1016/j.ydbio.2016.11.005 (2017).
DeLaurier, A. et al. The mouse limb anatomy atlas: an interactive 3D tool for studying embryonic limb patterning. BMC Dev. Biol. 8, 83. https://doi.org/10.1186/1471-213X-8-83 (2008).
Hiscock, T. W., Tschopp, P. & Tabin, C. J. On the formation of digits and joints during limb development. Dev. Cell. 41, 459–465. https://doi.org/10.1016/j.devcel.2017.04.021 (2017).
Huang, B. & Mackem, S. Rethinking positional information and digit identity: the role of late interdigit signaling. Dev. Dyn. 251, 1414–1422. https://doi.org/10.1002/dvdy.440 (2022).
McQueen, C. & Towers, M. Establishing the pattern of the vertebrate limb. Development 147 https://doi.org/10.1242/dev.177956 (2020).
Petit, F., Sears, K. E. & Ahituv, N. Limb development: A paradigm of gene regulation. Nat. Rev. Genet. 18, 245–258. https://doi.org/10.1038/nrg.2016.167 (2017).
Zhu, J., Patel, R., Trofka, A., Harfe, B. D. & Mackem, S. Sonic Hedgehog is not a limb morphogen but acts as a trigger to specify all digits in mice. Dev. Cell. 57, 2048–2062e4. https://doi.org/10.1016/j.devcel.2022.07.016 (2022).
Delgado, I., Torres, M. & Gradients Waves and Timers, an overview of limb patterning models. Semin Cell. Dev. Biol. 49, 109–115. https://doi.org/10.1016/j.semcdb.2015.12.016 (2016).
Stewart, T. A. et al. Evidence against tetrapod-wide digit identities and for a limited frame shift in bird wings. Nat. Commun. 10, 3244. https://doi.org/10.1038/s41467-019-11215-8 (2019).
Tschopp, P. & Tabin, C. J. Deep homology in the age of Next-Generation sequencing. Philos. Trans. R Soc. B Biol. Sci. 372, 20150475. https://doi.org/10.1098/rstb.2015.0475 (2017).
Wang, Z., Young, R. L., Xue, H. & Wagner, G. P. Transcriptomic analysis of avian digits reveals conserved and derived digit identities in birds. Nature 477, 583–586. https://doi.org/10.1038/nature10391 (2011).
Duboc, V. & Logan, M. P. O. Regulation of limb bud initiation and limb-type morphology. Dev. Dyn. 240, 1017–1027. https://doi.org/10.1002/dvdy.22582 (2011).
Wanek, N., Muneoka, K., Holler-dinsmore, G., Burton, R. & Bryant, S. V. A staging system for mouse limb development. J. Exp. Zool. 249, 41–49. https://doi.org/10.1002/jez.1402490109 (1989).
Suzuki, R., Shimodaira, H. & Pvclust An R package for assessing the uncertainty in hierarchical clustering. Bioinformatics 22, 1540–1542. https://doi.org/10.1093/bioinformatics/btl117 (2006).
Lopez-Rios, J. The many lives of SHH in limb development and evolution. Semin Cell. Dev. Biol. 49, 116–124. https://doi.org/10.1016/j.semcdb.2015.12.018 (2016).
Zhou, Z. et al. Neogenin regulation of BMP-Induced canonical Smad signaling and endochondral bone formation. Dev. Cell. 19, 90–102. https://doi.org/10.1016/j.devcel.2010.06.016 (2010).
Suzuki, T., Hasso, S. M. & Fallon, J. F. Unique SMAD1/5/8 Activity at the Phalanx-Forming Region Determines Digit Identity. Proc. Natl. Acad. Sci. 105, 4185–4190, (2008). https://doi.org/10.1073/pnas.0707899105
Dahn, R. D. & Fallon, J. F. Interdigital regulation of digit identity and homeotic transformation by modulated BMP signaling. Science 289, 438–441. https://doi.org/10.1126/science.289.5478.438 (2000).
Zhu, J. et al. Uncoupling Sonic Hedgehog control of pattern and expansion of the developing limb bud. Dev. Cell. 14, 624–632. https://doi.org/10.1016/j.devcel.2008.01.008 (2008).
Barrett, J. M., Raineri Tapies, M. G. & Shepherd, G. M. G. Manual dexterity of mice during Food-Handling involves the thumb and a set of fast basic movements. PLoS One. 15, e0226774. https://doi.org/10.1371/journal.pone.0226774 (2020).
Schnerwitzki, D., Englert, C. & Schmidt, M. Adapting the pantograph limb: differential robustness of Fore- and hindlimb kinematics against genetically induced perturbation in the neural control networks and its evolutionary implications. Zoology 157, 126076. https://doi.org/10.1016/j.zool.2023.126076 (2023).
Marzke, M. W. Precision grips, hand morphology, and tools. Am. J. Phys. Anthropol. 102, 91–110, (1997). https://doi.org/10.1002/(SICI)1096-8644(199701)102:1<91::AID-AJPA8>3.0.CO;2-G.
Rolian, C., Lieberman, D. E. & Hallgrímsson, B. The Coevolution of human hands and feet. Evolution 64, 1558–1568. https://doi.org/10.1111/j.1558-5646.2010.00944.x (2010).
Theiler, K. The House Mouse: Atlas of Embryonic Development 2nd edn pp. 53–131 (Springer-Verlag, 1989).
Musy, M. et al. A quantitative method for staging mouse embryos based on limb morphometry. Development 145 https://doi.org/10.1242/dev.154856 (2018).
Nagy, A., Gertsenstein, M., Vintersten, K. & Behringer, R. Alcian blue staining of the mouse fetal cartilaginous skeleton: Fig. 1. Cold Spring Harb Protoc. 2009 (pdb.prot5169). https://doi.org/10.1101/pdb.prot5169 (2009).
Rigueur, D. & Lyons, K. M. Whole-Mount skeletal staining. Methods Mol. Biol. 1130, 113–121. https://doi.org/10.1007/978-1-62703-989-5_9 (2014).
Pertea, M., Kim, D., Pertea, G. M., Leek, J. T. & Salzberg, S. L. Transcript-level expression analysis of RNA-seq experiments with HISAT, stringtie and ballgown. Nat. Protoc. 11, 1650–1667. https://doi.org/10.1038/nprot.2016.095 (2016).
Robinson, M. D. & Oshlack, A. A. Scaling normalization method for differential expression analysis of RNA-Seq data. Genome Biol. 11, R25. https://doi.org/10.1186/gb-2010-11-3-r25 (2010).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. EdgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. https://doi.org/10.1093/bioinformatics/btp616 (2010).
Chen, C. et al. TBtools-II: A one for all, all for one bioinformatics platform for biological Big-Data mining. Mol. Plant. 16, 1733–1742. https://doi.org/10.1016/j.molp.2023.09.010 (2023).
Edmonds, R. D. et al. Transcriptomic response of murine liver to severe injury and hemorrhagic shock: A Dual-Platform microarray analysis. Physiol. Genomics. 43, 1170–1183. https://doi.org/10.1152/physiolgenomics.00020.2011 (2011).
Young, M. D., Wakefield, M. J., Smyth, G. K. & Oshlack, A. Gene ontology analysis for RNA-Seq: accounting for selection bias. Genome Biol. 11, R14. https://doi.org/10.1186/gb-2010-11-2-r14 (2010).
Acknowledgements
This research was funded by a grant from the Department of Science & Technology of Liaoning Province of China (no. 2024-MS-099).
Author information
Authors and Affiliations
Contributions
Conceptualization, S.Z. and Z.W.; methodology, Z.W.; validation, W.D., D.G., J.C. and J.Y.; formal analysis, W.D. and S.B.; investigation, W.D., D.G. and J.C.; resources, S.Z. and Z.W.; writing—original draft preparation, W.D.; writing—review and editing, Z.W. and D.I.; visualization, W.D. and Z.J.; supervision, Z.W.; project administration, Z.W.; funding acquisition, Z.W. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All animal experiments were conducted in accordance with the ARRIVE Guidelines and the Institutional Animal Care and Use Committee of Shenyang Agricultural University following approval by the Animal Ethics Committee (Permit # 2022030706).
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Dang, W., Gao, D., Jia, Z. et al. Proximal phalanges of digit 1 have different identities in the forelimbs and hindlimbs of the mouse. Sci Rep 15, 37417 (2025). https://doi.org/10.1038/s41598-025-21196-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-21196-y
