
Abstract
Rice (Oryza sativa L.), a staple food for over half of the global population, is highly sensitive to low temperatures, particularly during early growth stages, resulting in significant yield losses. Addressing cold stress during the early stages of rice cultivation by screening rice genotypes can ensure optimal crop establishment and subsequent growth. An experiment was carried out by selecting ten rice genotypes for evaluation for cold tolerance through laboratory-based germination assays followed by subsequent screening with respect to biometric and biochemical parameters. Meher exhibited the highest germination rate (95%) under cold stress conditions, while Jajati and Ranidhana showed minimal variation in germination (2–7%). PERCOL (Percentage of seeds with coleoptile superior to 5 mm) and REDCOL (Percentage of reduction in coleoptile length) values remained consistent for Jajati and Pradeep. Cold stress led to notable reductions in plant height (9.2–31.1%) and the number of productive tillers per plant (36.8–37.1%). Samanta and Birupa recorded the highest number of productive tillers under control and cold stress conditions, respectively. Panicle exsertion was marginally impacted in Ranidhana (2.9%) and substantial decline recorded in Meher (19.2%), highlighting genotype-specific resilience. Spikelet fertility and 100-seed weight declined by 5.2–7.2% and 15–18%, respectively, in Samanta, and Jajati. Seed yield per plant was highest in Birupa (12.25 g/plant) under cold stress conditions. GGE biplot analysis indicated Kharavela and Birupa as high yielders under control and cold stress, respectively. Highest proline (4.6 mg/g) and carbohydrate content (18.08%) were noted in Samanta. Ranidhana and Samanta resulted in stable yield across environments indicating broader adaptability.
Similar content being viewed by others

Introduction
Rice is a crucial staple food crop ranking third worldwide and serving energy source for half of the global population1. This essential grain not only provides a significant source of calories but also plays a vital role in the economies of many countries. Beyond its nutritional value, its cultivation and trade are deeply intertwined with cultural practices and traditions, making it central to food security and livelihoods for millions. Unlike wheat and barley, rice plants, especially the Indica subspecies, are more vulnerable to cold stress as well as fluctuations in temperature regime, severely impacting rice productivity2. Cold stress, mainly in the cold regions of tropical and subtropical areas, hinders germination, delays seedling growth and inhibits flowering, including pollen viability, thus posing a major threat to the sustainability of rice production3,4. Prevalence of suboptimal temperature at reproductive stages of rice also has a negative bearing on the grain yield5. Low-temperature stress has been emerged as a substantial challenge for rice cultivation in almost all major countries contributing significantly to the global rice production, including Korea and Japan6. The global impact of cold stress on rice is highlighted by the decrease in rice production in areas of high-latitude and high-altitude regions of tropical countries like the Philippines and Thailand, along with instances of severe yield reduction in Northern Japan, Australia, Brazil and Argentina7. Yield reduction as a result of cold stress to the tune of 38.6%8, 0.65 million tonnes in Japan9, 3–5 million tonnes in China and 3.9 tonnes per hectare in Korea10 has been reported from 2007 to 2015. Although about 87% of global rice production is met by the eleven Asian countries significantly contributing to global food security11, low-temperature stress impacts rice production in many Asian countries12,13. Low-temperature stress is responsible for more than 50% yield loss excluding diseases and pest attacks14. The dry season rice of India, boro rice faces cold stress during seedling stage across 4 million hectares of rice areas15 including 60% of the area of North-eastern and western hill states of Himalayas16.
Cold stress impairs rice growth and development by poor seedling development, yield potential, and affecting overall crop performance2,3. However, the effect of cold stress on morphological and physiological traits of rice varies is stage dependent with distinct response observed during vegetative and reproductive stages. Additionally, low temperature (< 20 °C) delays rice germination and seedling establishment17, reduces seedling vigour18, and reduces the number of tillers19 by enhancing the mortality rate19,20,21,22. Cold stress adversely affects sink formation during the reproductive stage22,23 manifested by reduction in flowering and delayed panicle initiation24. Further cold stress is responsible for panicle sterility and poor seed setting, subsequently affecting grain quality and yield19,25.
About 60% of the north-eastern and western Himalayan hilly regions, is subjected to cold stress as a result of the cold irrigation water from melted snow combined with low ambient air temperature exacerbates cold stress in sensitive indica species26. Boro rice, normally cultivated from November to May in Eastern India and adjoining areas, is subjected to low temperature at both the seedling and reproductive stages. Selecting cold stress-tolerant genotypes for optimising yield in early sown indica genotypes is crucial27. A number of studies have already been conducted to screen genotypes for cold tolerance. However, characterization of genotypes with an optimum yield and nutritional richness suited to for resilient rice production while conserving the genetic diversity has not been explored. With changing climate scenarios, the response of genotypes is variable, thus making it necessary to identify location-specific resilient genotypes for better adaptability under fluctuating temperature regimes as well as climatic conditions. The characterization of the existing genotypes which are tolerant to diseases and insects and with enriched nutritional profile will further contribute to food security aligning with SDG 2 of UN Sustainable Development Goals (SDG). Based on this fact, our research focused on the initial screening of ten genotypes with uniform maturity durations for cold tolerance based on different morphological characters at the seedling and reproductive stages.
Materials and methods
The experiment was conducted at the Dr. G. V. Chalam Seed Testing Research Laboratory, AICRP on Seed Technology Research, NSP (Crops). In this study, ten medium duration rice genotypes namely, Bhanja, Bhuban, Birupa, Jajati, Kharavela, Meher, Pradeep, Pratap, Ranidhana and Samanta were collected from the EB-II section of the Central Research Farm, OUAT Bhubaneswar.
Laboratory conditions
In laboratory, 100 seeds of each genotype were placed in each petridish (90 mm × 15 mm) on 9 cm wet filter paper and each genotype was replicated three times in Completely Randomized Design (CRD). Before placing the seeds for germination, all the seeds were washed in 70% aqueous ethanol for 30 s followed by immersion in 5% aqueous sodium hypochlorite for 20 min, and then rinsed six times with distilled water. Seeds were then placed on Petri dishes containing two layers of germination paper, wet with distilled water and 1 ml of Benomyl solution 2.5 ppm to avoid contamination. The petri dishes were kept in a growth chamber at a light/dark cycle of 14/10 h with a light intensity of 200 μ mole photons/m2/s. The temperature of the growth chamber was set at 27 ℃ for control condition and 15 °C for cold treatment28. After 4 days of treatment, the chamber temperature was adjusted to 27 ℃, all petri dishes were kept uncovered for 10 days without interruption maintaining standard conditions of moisture and light. Here we had used a germinator (INDOSAW, single chamber type) with program for 24 h. Healthy seedlings were collected and characterized for seedling parameters for evaluation of cold tolerance of rice genotypes.
-
1. Germination percentage (%): The number of germinated seeds was counted and the percentage of germination was calculated using the following formula:
$${\text{Germination percentage}}\left( {\text{\% }} \right){\text{ = Number of normal seedlings/Total no. of seeds planted}} \times {100}{\text{.}}$$ -
2. Percentage of seeds with coleoptile superior to 5 mm (PERCOL): It was obtained by considering all the germinated seeds after 14 days of the cold treatment and by counting the percentage of coleoptile with length superior to 5 mm29.
$${\text{PERCOL = }}\left( {\text{number of seeds with coleoptile > 5 mm}} \right) \times {100/20}$$ -
3. Percentage of reduction in coleoptile length (REDCOL): It was obtained through comparison of average coleoptile length 14 days after germination at 15 °C (cold treatment) and at 27 °C (control), and calculating the percentage of reduction in coleoptile length by germination under cold temperature29
$${\text{REDCOL = }}\left[ {\left( {{\text{col}}{\text{. length under cold temperature }} \times {100}} \right){\text{/ col}}{\text{. length under control}}} \right]{\text{{-}100}}$$
where coleoptile length is the average of the 20 seeds evaluated per replication per genotype.
Pot culture
Well germinated seeds of both cold treated seeds and control (normal seed) of each genotype (5 numbers each) were transplanted in the pot (30 cm × 25 cm). Pots were filled with sandy loam soil and grown outdoors to simulate field conditions during Rabi, 2022–23. Plants were watered two times a day. Before sowing, GROMOR fertilizer was applied at 25 g/pot. Urea was top dressed with 9 g/pot at mid tillering and panicle initiation stage to each pot along with other standard cultivation practices. A Completely Randomized Design was used with each of the genotypes replicated thrice under control and cold stress, and observations for various growth and yield attributes were recorded for each genotype from each replication.
The injury scoring was determined in the rice seedlings considering seedling colour, spikelet fertility from tillering to maturity. The score uses a scale of 1 (highly cold tolerant) to 9 (highly cold sensitive) in accordance with the standard evaluation system for rice that was developed by the International Rice Research Institute14. After 14 days of cold treatment, under stress condition, this experiment is performed by scoring seedling, 1 = dark green seedling, 2–3 = light green seedling, 5 = yellow seedling, 7 = brown seedling, 9 = dead seedlings30.
Biochemical parameters
Various biochemical parameters were analysed by collecting fresh leaf samples from the seedlings at 30 DAS (Days After Sowing), 60 DAS and 90 DAS The proline content, total soluble protein, carbohydrate and Malondialdehyde (MDA) content were determined31,32,33,34.
Proline content
Fresh leaf samples of 200 mg from seedlings were homogenized in 3% (w/v) aqueous sulfosalicylic acid (5 ml) followed by filtration through Whatman No. 2 paper. The filtrate was then centrifuged at 4000 rpm for 15 min and 5 ml each of glacial acetic acid and acid ninhydrin were added to 5 ml of supernatant. Then it was subjected to hot water bath (100 °C) for 1 h. After heating, tubes were cooled in an ice bath, followed by the addition of 5 ml toluene. The mixture was shaken for 20–30 s, and the upper toluene layer was collected and brought to room temperature. The intensity of the resulting red color was measured at 520 nm using a UV–Vis spectrophotometer. Proline concentration was determined using a standard curve and expressed as mg/g fresh weight31.
Total soluble protein content
Fresh leaf of 1 g was macerated in a 10 ml solution of 10% TCA followed by centrifuging at 5000 rpm for 10 min32. The supernatant was discarded and the residue was mixed thoroughly with 10 ml of 1 N NaOH using a glass rod. After centrifuging again at 10,000 rpm for 10 min at 40 °C, 0.2 ml of the sample extract was pipetted into a test tube followed by dilution to 2 ml with water. In each test tube, including the blank, 10 ml of reagent C was added. After 10 min, 1 ml of reagent D was mixed followed by incubation at room temperature in the dark for 30 min. The appearance of a blue colour indicated the reaction completion which was measured at 660 nm. A standard protein curve was prepared and from this curve, the sample protein content was calculated as mg/g FW of the sample.
Carbohydrate content
Carbohydrate content of leaf samples was estimated by Anthrone method using 0.1 g leaf sample33 by Eq. (1).
Malondialdehyde (MDA) content
Lipid peroxidation indicated by malondialdehyde content of leaves was estimated34 by sampling 0.2 g of sample homogenized in 3 ml of 50 mM phosphate buffer at pH 7.0. The resulting homogenate underwent centrifugation at 15,000 g for 15 minutes. Then, 2.0 ml of 0.5% thiobarbituric acid (TBA) in 20% trichloroacetic acid (TCA) was added to 2 ml of supernatant. The mixture was then subjected to hot water bath for 30 minutes followed by ice bath. After a further centrifugation at 10,000 g for 10 minutes, the absorbance of the supernatant was measured at 532 nm and 600 nm using a spectrophotometer and MDA content was calculated by the extinction coefficient, which is equal to 155 mM −1cm−1 using the standard Eq. (2).
Data and statistical analysis
Data were analyzed by Analysis of Variance using R studio and means comparison by Tukey’s test (p = 0.05). Relationships among the measured characteristics were established through Pearson’s coefficient of correlation. These statistical methods provided insights into variable relationships and enabled thorough examination of the data patterns and associations. The GGE (Genotype + Genotype-by-Environment Interaction) Biplot analysis was carried out to identify best genotypes with respect to germination percentage and yield. The first Principal Components (PC1) scores of genotypes and environments against their respective scores for second Principal Components (PC2) were generated by subjecting the singular-value decomposition of environment-centered35.
Result
Effect on seedling growth
As a result of cold treatment, the growth parameters were decreased in all genotypes (Fig. 1) compared to control with respect to germination, PERCOL and REDCOL. Moreover, significant variation was observed among genotypes under cold treatment (Table 1) showing significant interactions of genotypes with varied temperature (Table 2).

Response of different genotypes under control and cold stress conditions.
Germination percentage
The germination percentage (Table 2) revealed significant variation with genotypes Meher (95%) and Ranidhana (86%) demonstrating highest germination percentage under control and cold stress conditions, respectively. Jajati recorded significantly lower germination percentage (75%) under control whereas, lowest germination percentage (58%) was observed in Bhanja under cold stress among all the genotypes. Minimal differences were recorded in germination percentages (2–7%) for genotypes Jajati, Ranidhana, Samanta, Birupa and Bhuban suggesting their relatively stable germination percentage under varied temperature conditions.
PERCOL
The PERCOL value serves as an important criteria for differentiating various genotypes for their cold tolerance. The results of our experiment (Table 2) revealed that Pratap was least affected by cold temperature with 10.75% of seeds with coleoptile length greater than 5 mm (PERCOL) followed by Ranidhana (9.86%). The lowest PERCOL value was recorded in Bhanja (5.46%) which remained statistically identical to Birupa (5.94%) and Pradeep (6.12%).
The boxplot (Fig. 2A) illustrated that Pratap was least affected with highest median PERCOL value (~ 10%) followed by Ranidhana. Additionally, Bhanja recorded the lowest median value of PERCOL with Kharavela and Meher performed moderately in terms of PERCOL. Further, the figure depicted highest spread in Pratap indicating variability whereas, Ranidhana demonstrated consistently higher value. Pradeep and Jajati had lower median value (6–7%) with no variability, suggesting higher consistency.

PERCOL and REDCOL of genotypes under cold stress conditions.
REDCOL
Alike, significant variation in percentage of seed with reduction in coleoptile length greater than 5 mm (Table 2) among genotypes was observed with higher percentage of reduction in coleoptile length in genotypes such as Ranidhana (4.75%). This was closely followed by Pratap (4.68%) and lowest value was recorded in Samanta (3.06%). Ranidhana, Kharavela and Pratap resulted in statistically similar reduction in coleoptile length under cold stress.
The boxplot analysis of genotypes for REDCOL (Fig. 2B) highlighted significant variation among genotypes with highest median value being observed for Ranidhana. Meher showed moderate median value for REDCOL (~ 4%) with significant variability. Samanta had the lowest median value with negligible variability. Jajati and Pradeep exhibited moderate value of REDCOL along with lower variability indicated by narrower spread.
Effect of cold stress on the morphological characters of rice genotypes at the vegetative and reproductive stage
Effect on growth attributes
Various growth attributes such as plant height, no. of tillers per plant were significantly influenced by cold stress and control conditions (Fig. 3). The plant height ranged from 90.50 to 110 cm under control condition with genotype Samanta exhibiting the highest plant height (110 cm). Jajati reached up to 84.03 cm height followed by Samanta with a plant height of 83.94 cm under cold stress condition. The plant height was reduced by 9.1–31.1% under cold stress compared to control with the highest reduction of plant height in Kharavela (31.1%) and lowest in Pradeep (9.1%). The plant height was reduced by 9.4% in Jajati, whereas Ranidhana and Samanta resulted in 10.1% and 23.6% reduction in plant height, respectively.

Effect of varied temperature treatment on growth attributes of rice in pot culture.
The number of tillers per plant varied significantly among the genotypes, with Kharavela demonstrating highest tillers per plant (15.5) under control conditions whereas, Samanta was superior in terms of tillers per plant (8.25) under cold stress. The number of productive tillers per plant exhibited significant reduction to the extent of 36.8–37.1% relative to control. Samanta recorded highest number of productive tillers per plant under control conditions, whereas, Birupa was superior under cold stress.
Effect on yield attributes and yield
The results of our experiment (Fig. 4) showed that Bhanja was least affected by cold stress with respect to days to 50% flowering requiring 85 days. In contrast, in Ranidhana, time taken to reach 50% flowering was highest with a value of 106 days. Similarly, under control conditions, Jajati exhibited longest duration to reach 50% flowering (117 days) among genotypes, whereas, remained at par with Ranidhana under cold stress. Panicle length was longest in Ranidhana under control conditions and in Pradeep under cold stress conditions. Jajati recorded the maximum reduction in panicle length (- 51.5%) under cold stress. Panicle exsertion was recorded to be highest for Ranidhana followed by Jajati and Pradeep whereas, Meher had the lowest value. Further, panicle exsertion was reduced by 2.9% and 19.2% for Ranidhana and Meher, respectively, implying less significant effect of cold stress on Ranidhana.

Effect of cold stress on yield attributes and yield of rice in pot culture.
Spikelet fertility was highest in Samanta and Birupa under control and cold stress conditions, respectively. The spikelet fertility was least affected by cold stress in Birupa and Kharavela. However, there was significant reduction in spikelet fertility to the tune of 5.2–7.2% in other genotypes such as Ranidhana, Samanta and Jajati. The 100 seed weight ranged from 1.76 to 2.55 g under control conditions and resulted in a decline of 15–18% under cold stress. Comparison of seed yield per plant revealed, Jajati outperformed under control conditions, whereas, Birupa recorded the highest yield under cold stress conditions remaining at par with Jajati.
Correlation coefficient between the seedling and reproductive characters
The association between laboratory and field parameters under cold temperature aids in selection for cold tolerance genotypes. Hence, correlation analysis was carried out among seed, seedling and yield parameters and the results were presented in Fig. 5.

Correlation analysis of seedling and yield parameter among genotypes [V1: Germination (%), V2: PERCOL (%),V3: REDCOL (%), V4: Days to 50% flowering ,V5: Days to complete flowering,V6:Panicle exsertion%,V7: Plant height (cm),V8:No. of tiller/plant,V9: No. of Productive tillers/plant, V10 :Panicle length (cm), V11: Spikelet fertility (%),V12: 100 seed weight (g), V13: yield per plant (g). The size and color intensity of each circle indicate the strength of the Pearson correlation coefficient between variable pairs. Blue circles represent positive correlations, whereas red circles denote negative correlations. Darker and larger circles correspond to stronger correlations, while lighter and smaller circles indicate weaker associations.]
The germination percentage recorded significant positive correlation with number of productive tillers per plant (r = 0.674*) at 5% level of significance. Whereas, a non- significant positive correlation was observed with PERCOL (r = 0.400), REDCOL (r = 0.342), days to 50% flowering (r = 0.409) and number of tillers per plant (r = 0.314), spikelet fertility (r = 0.197) and yield per plant (r = 0.606). Days to complete flowering, panicle exsertion, plant height, panicle length were negatively correlated with germination percentage. PERCOL demonstrated non-significant correlation with REDCOL (r = 0.089) at 5% level of significance. However, relatively higher positive correlation of PERCOL was observed with days to 50% flowering, days to complete flowering, panicle exsertion, plant height, number of tillers per plant and number of productive tillers per plant, spikelet fertility, 100 seed weight and yield per plant whereas, negative correlation with other parameters. Contrary to this, REDCOL showed negative correlation with days to complete flowering, plant height, number of tillers per plant, panicle length and spikelet fertility. Days to 50% flowering was negatively correlated with panicle length (r = − 0.654), whereas remained non-significant with remaining traits. Panicle exsertion showed strong significant correlation with spikelet fertility (r = 0787**) at 1% level of significance but non-significant correlation with others. Number of productive tillers per plant exhibited higher significant positive correlation with spikelet fertility (r = 0.745*) and yield per plant (r = 0.645*) at 5% level of significance. Panicle length and 100 seed weight showed non-significant correlation, whereas, spikelet fertility demonstrated significant correlation with yield per plant (r = 0.653*) at 5% level of significance.
GGE analysis for germination percentage
The GGE (Genotype + Genotype-by-Environment Interaction) Biplot analysis was carried out to distinguish the best genotypes across various environments. The which-won-where plot (Fig. 6A) of germination percentage capturing 68.21% and 31.79% variation by the principal components 1 and 2 (PC1 and PC2), respectively. The rays divided the polygon into 4 sectors with 2 mega environments. The genotype Meher and Ranidhana performed best in control and cold stress conditions respectively. Jajati and Bhanja performed well in different sectors indicating distinct environment and there was significant interaction of environments on genotypes. The genotypes Samanta, Kharavela situated near origin indicating relatively stable performance across environments.

GGE biplot analysis for germination percentage of genotypes under varied conditions.
The Mean vs. stability analysis (Fig. 6B) characterized the genotypes based on performance and consistency across locations. The figure showed that the genotypes Meher, Pratap and Ranidhana had higher germination percentage over mean performance, whereas, the genotypes Bhanja and Jajati performed poorly in terms of mean germination percentage. The genotypes Ranidhana and Birupa had higher germination percentage under cold stress conditions with higher variability whereas, Kharavela being centrally located exhibited stable germination percentage.
GGE analysis for yield
The GGE analysis was carried out to identify best genotypes in terms of yield. The principal components 1 (PC1) and principal components 2 (PC2) captured 73.14% and 26.86% of the variations respectively (Fig. 7).

GGE biplot analysis for yield of genotypes under varied conditions.
The Which-won-where plot (Fig. 7A) indicated that the rays divided the polygon into 6 sectors with two mega environments. The genotypes Kharavela and Birupa performed best under control and cold stress conditions, respectively. The genotypes Jajati, Bhanja and Meher performed better in separate environment indicating distinct environment requirement. The genotypes Ranidhana and Samanta being situated near the origin indicated stable performance in terms of yield.
Figure 7B showed the Mean vs stability analysis of genotypes and helps to distinguish genotypes on the basis of mean performance and stability across environments. Pratap, Meher and Kharavela demonstrated higher mean performance, whereas, genotypes Bhanja and Jajati yielded lower than average. Ranidhana and Samanta were comparatively stable performer in terms of yield. Birupa and Pradeep showed better adaptability under cold stress conditions whereas, Kharavela showed better adaptability under control conditions.
Ranking of genotypes (Fig. 7C) showed that Birupa, Samanta and Pradeep were closer to concentric circle indicating ideal genotypes. Bhuban, Ranidhana and Bhanja were moderate performer with respect to yield under cold stress and can be ranked as Bhuban > Ranidhana > Bhanja.
The Fig. 7D characterized the genotypes based on their discriminativeness and representativeness and highlighted that, Jajati and Meher were high yielder across environments. Bhanja, Ranidhana, and Kharavela being far from AEC (Average Environment Coordination) axis and higher discriminativeness indicating suitability in a specific environment. Samanta and Bhuban were clustered near centre indicating average performance across environments.
Classification of rice genotypes based on injury scoring at the seedling, spikelet fertility and tillering to maturity stage
Rice seedlings displayed diverse response to cold stress at 15 °C temperature. Consequently, genotypes were categorized into tolerant, moderately tolerant and susceptible based on injury scores, reflecting visual seedling appearance under cold stress conditions. This classification helps in identifying genotypes with better performance under cold stress (Table 3). Among the genotypes, Jajati was identified as highly tolerant (Injury score 0–1), two of the genotypes viz. Ranidhana and Samanta were considered as tolerant (Injury score 2–3). Moreover, Birupa genotype was found to be moderately tolerant (Injury score 4–5), Bhuban demonstrated moderate sensitivity (Injury score 6–7), whereas, Pratap, Meher, Kharavela, Bhanja and Pradeep were highly sensitive to cold stress (Injury score 8–9).
Biochemical parameters
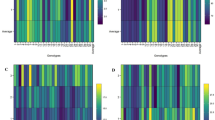
Significant differences among genotypes for accumulation of various biochemical parameters were observed (Fig. 8) on a temporal scale. The proline content was relatively higher at 30 DAS followed by gradual decline up to 90 DAS. Pratap exhibited 55% higher proline accumulation (12.88 mg/g) compared to Samanta at 30 DAS, whereas, Jajati and Samanta noted highest accumulation of proline at 60 (6.06 mg/g) and 90 DAS (4.96 mg/g), respectively. On contrary, Bhuban recorded lowest proline content at both 60 and 90 DAS with a reduction of 41.9% and 48.3% relative to Jajati and Samanta, respectively. With respect to protein content, genotypes such as Samanta (0.86 mg/g), Bhuban (2.43 mg/g) and Kharavela (2.58 mg/g) recorded highest protein content at 30, 60 and 90 DAS. Samanta closely followed Kharavela with a slight reduction of only 4.2% in protein content. Birupa resulted in 50.2% reduction in protein content (1.23 mg/g) at 90 DAS compared to Samanta. The carbohydrate content was highest in Bhuban (4.68%), Jajati (8.35%) and Samanta (18.08%) at 30, 60 and 90 DAS, respectively. Jajati and Ranidhana closely followed Samanta with a slight decline to the extent of 5.8 and 8.6%, respectively relative to Samanta. The analysis of Malondialdehyde (MDA) content highlighted highest content in Ranidhana (3.76 µmole/g) followed by Jajati (3.22 µmole/g) at 30 DAS. Birupa recorded highest value at 60 DAS which was 16.8% and 21.1% higher compared to Ranidhana and Samanta, respectively. At 90 DAS, the MDA content was highest in Pradeep (3.36 µmole/g) and lowest in Kharavela. Ranidhana and Samanta showed a reduction in MDA content by 11.6% and 32.7%, respectively.

Response of various biochemical parameters among genotypes under under cold stress condition on a temporal scale.
Discussion
Seed and seedling parameter
Germination percentage is one of the important parameter for screening cold tolerance of genotypes as cold stress reduced germination percentage due to greater sensitivity of rice low temperature21. In our study, Ranidhana was least affected by cold temperature with highest germination percentage underscoring its relative tolerance at early stage. Minimal differences in germination percentage among genotypes Jajati, Ranidhana, Samanta, Birupa and Bhuban under control and cold conditions signifies their relatively stable germination under low temperature and cold stress moderately impacted theses genotypes at early stage. The inhibition of germination during the early growth stage and subsequent seedling injury under cold stress was reported by several workers2,36. The reduced germination under cold stress might be attributed to reduced metabolic activity leading to inactivation of enzymes which plays a key role in germination2,28 . Cold stress tolerant genotypes exhibited higher germination percentage which might be owing to better membrane stability and osmolyte accumulation facilitating water uptake for germination. Genotypes exhibiting higher germination percentage showed better adaptability under cold stress15.
The PERCOL value which recorded seeds with coleoptile length superior to 5 mm identifies resilience of genotypes to growth conditions and it serves as a crucial parameter for differentiating cold sensitive and tolerant genotypes37. Ranidhana was least affected by cold temperature indicated by highest PERCOL value. Ranidhana genotype with higher germination rate under cold stress along with minimal difference of germination rate between control and cold stress and with highest PERCOL value suggests tolerance to cold temperature at germination and seedling growth. REDCOL, which measures reduction in coleoptile length37, is also considered to be one of the important parameter for evaluating cold tolerance during germination in rice29,38. The coleoptile length was reduced in genotypes in response to cold stress as a result of effect of cold temperature on cellular elongation and division in addition to reduced metabolic activity29,36. Increased metabolic activity with less reduction in coleoptile length in genotypes can serve as an indicator for initial screening against cold tolerance. In our study, Samanta exhibited lowest REDCOL value with relatively higher germination percentage indicating relative tolerance to cold and resilience at seedling stage.
Growth attributes
The perusal of data also revealed significant reduction in plant height, number of tillers per plant in cold stress compared to control. The reduction in the number of leaves and tillers per plant in response to low temperature is an important criteria in understanding the plant responses to cold stress3 as productive tillers per plant contributes to the final yield of the plant under stress condition corroborated by significant correlation observed in our study (Fig. 5). The detrimental effect of cold stress on rice tillers characterized by decline in their numbers under cold stress36. This might be due to reduction in seedling stem elongation under cold stress leading to decreased plant height7. Samanta with higher plant height and productive tillers per plant indicated stable performance under cold stress condition at tillering stage. Birupa with highest productive tillers per plant under cold stress suggested better adaptability under cold stress at maximum vegetative growth stage.
Yield attributes and yield
Likewise significant reduction in various yield attributes and yield of rice genotypes were observed under cold stress conditions. The days to 50% flowering was delayed under cold stress with Bhanja requiring lesser days to reach 50% flowering. This might be attributed to disruption in essential physiological processes thereby hampering flowering and delaying maturity, growth and development under cold stress condition39. Genotypes with delayed 50% flowering might experience late-season maturity or yield reduction as a result of extension of the duration of the crop thereby indicating sensitivity to cold stress. Reduction in panicle length and panicle exsertion percentage were observed under cold stress from our study. Similar findings of reduction of panicle length and reduced panicle exsertion were reported under cold stress40. Complete panicle exsertion under cold stress can lead to higher spikelet fertility as well as more number of filled grains per panicle playing a crucial role in characterizing cold stress tolerance41 and vital in determining yield as demonstrated by the stronger positive correlation (Fig. 5). Spikelet sterility is crucial for evaluating cold tolerance in rice during booting and flowering because of stronger association of spikelet fertility with grain yield (Fig. 5). Highest spikelet fertility in Birupa and lowest in Meher indicates relatively tolerance and sensitivity to cold stress. Cold tolerance during reproductive stages is crucial for the survival of pollen, seed formation and optimal grain filling, ensuring maximum yield39. Yield per plant was observed to be highest in Birupa indicating better adaptability under cold stress and lowest in Bhanja. The reduction in yield in susceptible genotypes might be pertaining to reduction in percentage of fertile grains under cold stress conditions9,41. Further GGE analysis for yield revealed that Kharavela was the highest yielder under control whereas Birupa outperformed under cold stress conditions. Meher and Jajati were high yielders under control. Samanta and Ranidhana were the most stable performer in terms of yield across environment.
Biochemical parameters
The results of our study demonstrated proline content was gradually declined from from vegetative to tillering stage. Proline plays a vital role in defence mechanism against abiotic stress42 by eliminating excess H+ ions and maintaining optimal cytosolic pH during oxidative respiration. Proline accumulation under low temperatures enhanced adaptive response to cold stress43 as a result of stabilization of polyribosome and increasing water binding capacity while acting as reservoir of carbon and nitrogen13. Relatively higher proline accumulation in Samanta at 60 and 90 DAS indicated the adaptability of the genotype under cold stress with minimal oxidative stress as significant correlation had been reported between cold tolerance and accumulation of proline44. Similarly, highest proline content in Jajati at 60 DAS indicating significant osmotic adjustment in cold stress thereby making it tolerant at later growth stages, whereas, Bhuvan was susceptible to cold stress induced damage due to less accumulation of proline. The increase in protein content might be as a result of upgradation of various cold regulated proteins which serves as an indicator of cold stress tolerance45. Highest protein content in Kharavela, Samanta, and Bhuban might have been attributed to upregulation of cold stress related proteins critical for survival for cellular functioning under cold stress, highlighting their relative tolerance to cold stress. The carbohydrate content is crucial for synthesis of soluble sugars such as sucrose, hexose, raffinose, glucose, fructose and trehalose. These are by-product of degradation of starch and sucrose induced as a result of cold stress induced expression of enzymes thereby helping in osmotic adjustment while protecting cell membrane46. These sugars serve as compatible solutes as well as osmoprotectants against freezing damage serving as an important indicator of cold stress47. Carbohydrate content was more pronounced in Bhuban indicating significant osmotic adjustment while contributing to reserve of energy under cold stress thereby making it tolerant at earlier stages, whereas Ranidhana and Samanta were tolerant to cold stress at later stages. MDA is a by-product of lipid peroxidation caused by oxidative stress and used to identify stress index47. Exposure to cold stress at different stages increased MDA content in genotypes indicating the accumulation of reactive oxygen species48. Though, Ranidhana recorded highest MDA value at 30 DAS, there was reduction in MDA content at other stages which might be as a result of increased antioxidant defense system in response to initial exposure to cold stress. Samanta was moderately impacted by cold stress indicated by stable MDA content signifying wider adaptability on exposure to cold stress at different stages. Highest MDA content of Pradeep may be attributed to cold stress induced oxidative stress thereby damaging cellular membrane indicating relatively lower tolerance to cold stress. Ranidhana and Samanta were tolerant to cold stress with higher proline and carbohydrate content along with stable MDA value, whereas, Jajati was relatively stable indicated by moderate proline content. Genotypes such as Kharavela and Birupa showed moderate tolerance to cold stress. Genotypes Meher, Bhanja, Bhuba, Pradeep and Pratap were less resilient biochemically to cold stress indicating relative sensitivity.
The evaluation of ten rice genotypes for different parameters at different growth stages under contrasting conditions demonstrated marked variation in germination, seedling growth and yield parameters among genotypes signifying the differential response. Genotypes such as Ranidhana, Samanta, and Birupa exhibited relatively stable performance at different growth stages under cold stress, indicating inherent physiological and biochemical resilience to cold stress. However, the present study was carried out under pot culture conditions, which, although useful for initial screening of genotypes based on morpho-physiological parameters, imposed certain limitations. The nutrient and water acquisition can be limited as a result of restricted rooting volume. Further, absence of field level microclimatic conditions and inter-plant competition can alter tillering behaviour. Though, the relative ranking of genotypes can provide significant insights about response of different genotypes to cold stress, the exact magnitude of yield and stress can be further validated through multi-location and field-based studies for adaptability of identified genotypes under contrasting environments.
Conclusion
The temperature plays a crucial role in regulating the growth behaviour and grain yield of rice with low temperature adversely affects by interfering with plant metabolism and influencing various physiological and biochemical parameters. The response of different genotypes differ by varied temperature conditions. The growth and yield attributing characters reduced as a result of cold stress during germination. Initial screening of genotypes for cold stress at early stages of growth can optimize the crop production under cold stress conditions. Various parameters including minimal difference in reduction in coleoptile length as well as germination percentage facilitated screening at germination and seedling stage. Selecting genotypes with higher productive tillers per plant, higher spikelet fertility and 100 seed weight can optimize yield under field condition. In our study, Ranidhana and Samanta were most adaptable under contrasting conditions at different growth stages. Birupa and Jajati had specific environment requirement and can result optimum yield under cold stress and control, respectively indicating specific adaptability. Although initial screening on the basis of seedling parameters and germination percentages demonstrated significant reduction on exposure to cold stress, subsequent growth and yield attributes in the field were determined by the interaction of the genotype with prevailing climatic factors. Genotypes with broader adaptability can show resilience under cold stress with minimal impact on yield. This underscores the selection of genotypes based on initial screening followed by further genomic and molecular studies can optimize the yield under cold stress condition thereby contributing to food security. Further study can be undertaken under multiple locations experiencing diverse temperature regimes for getting deeper insights about response of genotypes to cold stress. Further, identification of QTL’s for traits associated with cold tolerance at different stages along with marker assisted selection in genotypes with broader genetic base and nutritional value in various regions can be undertaken for hastening up breeding programme while conserving the germplasm.
Data availability
All data generated or analysed will be made available on request.
References
Sen, S., Chakraborty, R. & Kalita, P. Rice—Not just a staple food: A comprehensive review on its phytochemicals and therapeutic potential. Trends Food Sci. Technol. 97, 265–285. https://doi.org/10.1016/j.tifs.2020.01.022 (2020).
Lone, J. A. et al. Cold tolerance at germination and seedling stages of rice: Methods of evaluation and characterization of thirty rice genotypes under stress conditions. Int. J. Curr. Microbiol. Appl. Sci. 7, 1103–1109. https://doi.org/10.20546/ijcmas.2018.701.134 (2018).
Ye, C. et al. Cold tolerance in rice varieties at different growth stages. Crop Pasture Sci. 60, 328–338. https://doi.org/10.1071/CP08352 (2009).
Dong, J. et al. Physiological and genome-wide gene expression analyses of cold-induced leaf rolling at the seedling stage in rice (Oryza sativa L.). Crop J. 7, 431–443. https://doi.org/10.1016/j.cj.2019.03.002 (2019).
Ndour, D. et al. Agro-morphological evaluation of rice (Oryza sativa L.) for seasonal adaptation in the Sahelian environment. Agronomy 6, 8. https://doi.org/10.3390/agronomy6010008 (2016).
Singh, B. K. et al. Cold stress in rice at early growth stage: An overview. Int. J. Pure Appl. Biosci. 5, 407–419. https://doi.org/10.18782/2320-7051.2750 (2017).
Cruz, R. P. D. et al. Avoiding damage and achieving cold tolerance in rice plants. Food Energy Secur. 2, 96–119. https://doi.org/10.1002/fes3.29 (2013).
Unan, R. et al. Cold stress reduces rice grain yield in temperate conditions. Revista Brasileira de Engenharia Agrícola e Ambiental. 26(12), 947–952. https://doi.org/10.1590/1807-1929/agriambi.v26n12p947-952 (2022).
Shimono, H. et al. Low temperature-induced sterility in rice: Evidence for the effects of temperature before panicle initiation. Field Crops Res. 101, 221–231. https://doi.org/10.1016/j.fcr.2006.11.008 (2007).
Gautam, A. G. & Suresh, J. QTL based screening of rice cultivars from Andhra Pradesh showing cold resistance at germination stage. Environ. Ecol. 35, 278–284 (2017).
Bandumula, N. Rice production in Asia: Key to global food security. Proc. Nat. Acad. Sci. India Sect. B Biolog. Sci. 88, 1323–1328 (2018).
Zhang, H. et al. Major QTLs reduce the deleterious effects of high temperature on rice amylose content by increasing splicing efficiency of Wx pre-mRNA. Theor. Appl. Genet. 127, 273–282. https://doi.org/10.1007/s00122-013-2216-4 (2014).
Zhang, Q. et al. Rice and cold stress: Methods for its evaluation and summary of cold tolerance-related quantitative trait loci. Rice 7, 24. https://doi.org/10.1186/s12284-014-0024-3 (2014).
IRRI. Standard Evaluation System for Rice International Rice Research Institute, Manila, (2002).
Pradhan, S. K. et al. Population structure, genetic diversity and molecular marker-trait association analysis for high temperature stress tolerance in rice. PLoS ONE 11(8), e0160027. https://doi.org/10.1371/journal.pone.0160027 (2016).
Najeeb, S. et al. Genetics and breeding of low-temperature stress tolerance in rice. In: Rice Improvement: Physiological, Molecular Breeding and Genetic Perspectives. pp. 221–280 https://doi.org/10.1007/978-3-030-66530-2_8 (2021).
Hussain, S. et al. Temperature extremes: Impact on rice: Growth and development. In: Plant Abiotic Stress Tolerance https://doi.org/10.1007/978-3-030-06118-0_6 (2019).
Ali, M. G., Naylor, R. E. L. & Matthews, S. Distinguishing the effects of genotype and seed physiological age on low temperature tolerance of rice (Oryza sativa L.). Exp. Agric. 42, 337–349. https://doi.org/10.1017/S0014479706003619 (2006).
Fujino, K. et al. Mapping of quantitative trait loci controlling low-temperature germinability in rice (Oryza sativa L.). Theor. Appl. Genet. 108, 794–799. https://doi.org/10.1007/s00122-003-1509-4 (2004).
Farrell, T. C. et al. Genotypic variation for cold tolerance during reproductive development in rice: Screening with cold air and cold water. Field Crops Res. 98, 178–194. https://doi.org/10.1016/j.fcr.2006.01.003 (2006).
Baruah, A. R. et al. Cold tolerance at the early growth stage in wild and cultivated rice. Euphytica 165, 459–470. https://doi.org/10.1007/s10681-008-9763-4 (2009).
Oliver, S. N., Dennis, E. S. & Dolferus, R. ABA regulates apoplastic sugar transport and is a potential signal for cold-induced pollen sterility in rice. Plant Cell Physiol. 48, 1319–1330. https://doi.org/10.1093/pcp/pcm100 (2007).
Kuroki, M. et al. A quantitative trait locus for cold tolerance at the booting stage on rice chromosome 8. Theor. Appl. Genet. 115(5), 593–600. https://doi.org/10.1007/s00122-007-0589-y (2007).
Shrestha, S. et al. Phenological responses of upland rice grown along an altitudinal gradient. Environ. Exp. Bot. 89, 1–10. https://doi.org/10.1016/j.envexpbot.2012.12.007 (2013).
Jena, K. K. et al. Identification of cold-tolerant breeding lines by quantitative trait loci associated with cold tolerance in rice. Crop Sci. 51, 517–523. https://doi.org/10.2135/cropsci2010.12.0733 (2012).
Akter, N. et al. Phenotypic and molecular characterization of rice genotypes’ tolerance to coldstress at the seedling stage. Sustainability. 14, 4871. https://doi.org/10.3390/su14094871 (2022).
Suh, J. P. et al. SSR analysis of genetic diversity and cold tolerance in temperate rice germplasm. Plant Breed. Biotechnol. 1, 103–110. https://doi.org/10.9787/PBB.2013.1.2.103 (2013).
Hsu, C. H. & Hsu, Y. T. Biochemical responses of rice roots to cold stress. Bot. Stud. 60, 14. https://doi.org/10.1186/s40529-019-0262-1 (2019).
Cruz, R. P. D. & Milach, S. C. K. Cold tolerance at the germination stage of rice: Methods of evaluation and characterization of genotypes. Sci. Agric. 61, 1–8. https://doi.org/10.1590/S0103-90162004000100001 (2004).
Kim, S. I. & Tai, T. H. Evaluation of seedling cold tolerance in rice cultivars: a comparison of visual ratings and quantitative indicators of physiological changes. Euphytica 178(3), 437–447. https://doi.org/10.1007/s10681-010-0343-4 (2011).
Bates, L. S. et al. Rapid determination of free proline for water-stress studies. Plant Soil. 39, 205–207. https://doi.org/10.1007/BF00018060 (1973).
Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. https://doi.org/10.1016/0003-2697(76)90527-3 (1976).
Yoshida, S. et al. Laboratory Manual for Physiological Studies of Rice (International Rice Research Institute, 1976).
Heath, R. L. & Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 125, 189–198. https://doi.org/10.1016/0003-9861(68)90654-1 (1968).
Yan, W. & Tinker, N. A. An integrated biplot analysis system for displaying,interpreting, and explori ng genotype X environment interaction. Crop Sci. 45, 1004–1016 (2005).
Sharma, N., Reinke, R. & Sacks, E. J. Comparison of methods to evaluate rice (Oryza sativa) germplasm for tolerance to low temperature at the seedling stage. Agronomy 11, 385. https://doi.org/10.3390/agronomy11020385 (2021).
Sharifi, P. Evaluation on sixty-eight rice germplasms in cold tolerance at germination stage. Rice Sci. 17, 77–81. https://doi.org/10.1016/S1672-6308(08)60103-0 (2010).
Priyanka, K. et al. Response of rice seedlings to cold tolerance under boro conditions. SABRAO J. Breed. Genet. 47, 185–190 (2015).
Suh, J. P. et al. Identification and analysis of QTLs controlling cold tolerance at the reproductive stage and validation of effective QTLs in cold-tolerant genotypes of rice (Oryza sativa L.). Theor. Appl. Genet. 120, 985–995. https://doi.org/10.1007/s00122-009-1226-4 (2010).
Andaya, V. C. & Mackill, D. J. Mapping of QTLs associated with cold tolerance during the vegetative stage in rice. J. Exp. Bot. 54, 2579–2585. https://doi.org/10.1093/jxb/erg243 (2003).
Cruz, R. P. D., Milach, S. C. K. & Federizzi, L. C. Rice cold tolerance at the reproductive stage in a controlled environment. Sci. Agric. 63, 255–261. https://doi.org/10.1590/S0103-90162010000600008 (2006).
Perveen, S. et al. Low temperature stress induced changes in biochemical parameters, protein banding pattern and expression of Zat12 and Myb genes in rice seedling. J. Stress Physiol. Biochem. 9, 193–206 (2013).
Zhang, Y. et al. Chitooligosaccharide plays essential roles in regulating proline metabolism and cold stress tolerance in rice seedlings. Acta Physiol. Plant. 41, 1–11. https://doi.org/10.1007/s11738-019-2864-3 (2019).
Liu, Z. et al. Arbuscular mycorrhizal fungi enhanced rice proline metabolism under low temperature with nitric oxide involvement. Front. Plant Sci. 13, 962460. https://doi.org/10.3389/fpls.2022.962460 (2022).
Pan, X. et al. Transcriptional and physiological data revealed cold tolerance in a photo-thermo sensitive genic male sterile line Yu17S. BMC Plant Biol. 22, 44. https://doi.org/10.1186/s12870-022-03437-8 (2022).
Seydel, C. et al. Temperature-induced dynamics of plant carbohydrate metabolism. Physiol. Plant. 174, e13602. https://doi.org/10.1111/ppl.13602 (2022).
Wang, S. Q. et al. Application of brassinolide alleviates cold stress at the booting stage of rice. J. Integr. Agric. 19, 975–987. https://doi.org/10.1016/S2095-3119(19)62639-0 (2020).
Zhao, Y. et al. Effect of low temperature on chlorophyll biosynthesis and chloroplast biogenesis of rice seedlings during greening. Int. J. Mol. Sci. 21(4), 1390. https://doi.org/10.3390/ijms21041390 (2020).
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
T.B. performed Conceptualization, Methodology, Writing—Original draft preparation; S.P performed Coceptualization, Data curation, Investigation, Supervision; T.J. performed Formal analysis, Software, Visualization; S.P. performed Supervision, Validation; S.M. performed Validation, Writing—Reviewing and Editing; K.K. performed Visualization, Formal analysis; M.K. performed Formal analysis; S.KM performed data curation; M.K.R. supervised and performed Validation.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Consent for publication
All the authors have provided informed consent for publication of the manuscript.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Bala, T., Pradhan, S., Jena, T. et al. Cold stress resilience in rice: genotypic variation, yield traits, and GGE biplot insights. Sci Rep 15, 37674 (2025). https://doi.org/10.1038/s41598-025-21562-w
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-21562-w
