
Abstract
The boosted Sterile Insect Technique (SIT) by releasing sterilized males coated with the larvicide pyriproxyfen has been suggested for the control of Aedes container-breeding mosquitoes. In 2023, a boosted-SIT field trial was applied in Greece against Aedes albopictus by weekly releases of pyriproxyfen-coated sterile males at the beginning (12 May–16 June) and peak (1 September–6 October) of mosquito activity, including two Mark-Release-Recapture sessions in July. No overall effect on egg densities was noted in the treated site, while a significant decrease compared to a control site was recorded in specific locations. The egg hatching rate in the treated site (56–57%) was lower than the control site (84–85%) during the release cycles and one month following the second release cycle. During the second release cycle and the following month, an overall suppression of the adult population was achieved (85–93% reduction compared to the control site), and high boosted sterile-to-wild male ratio (15:1) and competitiveness index of boosted-SIT males (0.46) were recorded, respectively. The suppression of both adult and egg populations was more pronounced a month after the conclusion of releases, indicating a residual activity of boosted-SIT. Our findings demonstrate the potential of boosted-SIT against Ae. albopictus particularly in suppressing adult population.
Similar content being viewed by others

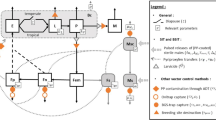

Introduction
Aedes albopictus (Skuse 1894), also known as the Asian tiger mosquito, is a container breeding mosquito that poses a considerable nuisance to humans due to its aggressive daytime biting behavior and a serious public health risk due to its ability to transmit harmful diseases such as dengue, chikungunya and Zika viruses1. It is considered the most invasive mosquito in the globe, currently occurring in 126 countries world-wide2. In Europe, since its first detection in Albania in 19793, Ae. albopictus has been established in 14 countries including Croatia, Italy, Germany, Greece, Spain, France, Portugal, and Switzerland4. Aedes albopictus was first reported in northwestern Greece in 2003–2004, and since then it has been widely dispersed across the country and the Attica Region including the Municipality of Athens5,6. In recent years, no local transmission of Aedes-borne diseases has been reported in Greece7. However, the widespread distribution and high population densities of the vector Ae. albopictus across the country poses a significant risk of autochthonous disease transmission, like the sporadic cases of locally transmitted dengue virus reported between 2010 and 2024 in Croatia, France, Spain, and Italy, as well as chikungunya virus outbreaks reported in Italy and France between 2007 and 20248.
It is expected that, Ae. albopictus will continue to expand its geographical distribution beyond its current range9, highlighting that one of the main reasons for the success of its invasion is the lack of efficient control methods10. The primary obstacle to effectively controlling Aedes container-breeding mosquitoes is the difficulty in finding and accessing the plethora of breeding sites that are often cryptic in domestic environments11,12. In addition, the use of synthetic insecticides, primarily pyrethroids and organophosphates in space spraying applications that target Ae. albopictus adults resting outdoors face several constraints related to high operational cost, low efficacy, development of mosquito resistance, and environmental and human safety issues13,14,15. Because currently the control of Ae. albopictus relies mainly on conventional larvicidal and adulticidal treatments, sometime coupled with source removal campaigns, it is necessary to identify new alternative integrated management strategies combining chemical and non-chemical control methods13,16.
The Sterile Insect Technique (SIT) has been used as an eco-friendly species-specific alternative control approach against mosquitoes and other insects. The standard SIT is based on the systematic mass releases of males sterilized by means of irradiation to mate with wild females and reduce their reproductive success leading eventually to local population decline17,18. SIT was initially tested against several mosquito species in the 1960, ’70, and ‘8019 and successively abandoned for approximately twenty years. The interest on SIT against mosquito vectors resurged since the 200020. In 2024, 17 on-going standard SIT projects were censed on a world-wide scale against Ae. albopictus, with varying results of induced sterility and population suppression. Only two of them, namely in Italy and Spain, have been performed under the pre-operational phase-III, and none under the final large-scale operational phase-IV, according to phase conditional approach for testing SIT against Aedes mosquitoes established by the WHO Vector Control Advisory group21,22.
In 2018, a phase-II small scale field trial of standard SIT was performed against Ae. albopictus for the first time in Greece in a 5-ha area (Vravrona, Attica Region) during the peak mosquito activity (September–November), resulting in significant reduction of egg hatch rates in comparison to untreated control sites23. The year after, from June to September 2019, the trial was repeated at a wider site (10 ha) in the same area (Vravrona, Attica Region) as SIT phase-II small scale field trial again for the suppression of Ae. albopictus, reaching a maximum of 78% induced sterility and egg reduction rates24. For the successful implementation, the SIT needs to be applied on wide area due to immigration or introduction of already mated females in the SIT area from adjacent areas25,26. SIT has been successfully implemented in large scale applications in the past resulting in eradication of tsetse flies, screwworms and Mediterranean fruit flies18. However, the implementation of the SIT against mosquitoes in large-scale trials is challenging because mosquitoes develop large populations that require the release of large number of sterile males to achieve population suppression, which implies high operational cost13,17. Hence, the SIT should not be considered as a standalone solution but as a part of an integrated vector management program combined with other control methods18,22.
In 2014, Bouyer & Lefrançois27 proposed for the first time the boosted-SIT concept for the control of Aedes mosquitoes, which relies on the release of sterilized males coated with pyriproxyfen (PPF), a larvicide (juvenile hormone analog) that does not affect adult survival. The wild females are contaminated with the larvicide during mating or mating attempts with the PPF-coated sterile males. Contaminated females transfer PPF to the oviposition sites resulting to the cessation of offspring production in case of successful mating with sterile males and to adult emergence inhibition of larvae produced from other females mated with wild-fertile males. Following the promising results of auto-dissemination stations of PPF for Ae. albopictus and Ae. aegypti management11,28,29 and the auto-dissemination of PPF by releasing PPF-treated males of Ae. albopictus30, the boosted-SIT is based on the same PPF auto-dissemination principle to a wider scale area though wild females mated with PPF coated sterile males27. By boosting SIT with PPF, it is estimated that the number of released sterile males needed to eliminate Ae. albopictus and Ae. aegypti population can be reduced by over 95%31. The boosted-SIT that was implemented in 2021 in phase-II pilot-scale field trials in La-Reunion against Ae. aegypti and in Spain against Ae. albopictus efficiently suppressed populations of both Aedes species in the treated sites32. Furthermore, the boosted-SIT against Aedes vectors has been proposed as a new strategy to prevent the establishment of other invasive mosquito species sharing the same breeding sites33.
In a plethora of published papers, SIT and IIT (Incompatible Insect Technique) have demonstrated good suppression capacity of mosquito populations when targeting either Ae. albopictus or Ae. aegypti24,26,34,35,36,37,38,39. These trials tracked SIT/IIT efficacy using a range of measures, such as adult female abundance, egg abundance, and egg hatching rate. Additional metrics have been considered to assess the quality of released sterile males such as the sterile-to-wild male ratio and sterile male mating competitiveness. Herein, we report the results from a phase-II pilot scale boosted-SIT field trial that was conducted for the first time in Greece from May to October 2023 aiming to suppress Ae. albopictus population. We assessed the performance of boosted-SIT in a treated site compared to a control site in terms of adult and egg population reduction as well as reduction of egg hatching rate, estimating also boosted sterile-to-wild male ratio and boosted SIT male mating competitiveness.
Materials and methods
Study area
The boosted-SIT field trial was implemented at a 15-ha site (treated site) in Vravrona area, located in the municipality of Markopoulo, east of Athens, in Attica Region, Greece. The treated site was a partially isolated semi-urban area surrounded by the sea in the north and high wooded hills in the south, west and east (Fig. 1). The nearest urban area was approximately 1.5 km far from the treated site. To assess the efficacy of the boosted-SIT, an untreated control site (Artemida), sized 5-ha, was established in a semi-urban area located 4.5 km north of the treated site in the municipality of Artemida-Spata, Attica Region (Fig. 1). During the field trial, no chemical treatments were performed, neither in public areas against mosquitoes nor in the treated and control sites.

Study sites selected for the field experiment of boosted-SIT to suppress wild populations of Aedes albopictus. Trials implemented from May to November 2023 in the outskirts of Athens (Greece) in Vravrona (treated site) and Artemida (control site). Mosquito populations were monitored with sampling stations (ovitraps + HLC) located at the black points drawn on the map. Maps on the left were created with ArcGIS Pro (3.0.0), available from Esri (Marathon Data, GR)]. The background map in the main plot on the right was retrieved from the Google Maps Platform https://mapsplatform.google.com/, using functions available in the ggmap package for R https://github.com/features/packages version 4.0.0, together with a private API key.
Production and transportation of sterile males
The production of sterile Ae. albopictus males was performed at the mass rearing facilities of Centro Agricoltura Ambiente “G. Nicoli” (CAA), Italy, following the methodology outlined by Bellini et al.25 and Malfacini et al.40,41. The mosquito colony used in the current field trial was established in CAA facilities from Ae. albopictus eggs collected from the Vravrona area following the IAEA mosquito mass rearing protocols42,43. At the pupal stage, males were exposed to a dose of 40 Gy irradiation using an X-ray irradiator (Radgil2, Gilardoni, Italy). This irradiation level achieves the best possible compromise between sterility and the performance of sterile males. It is noted that the quality of the resultant adults and the level of sterility attained are significantly impacted by the age of the male mosquito pupa at the time of irradiation (measured from pupation to the irradiation moment)44. Prior to packaging, adult male mosquitoes were kept in a large cooling cabinet at 8 ± 1 °C, 85 ± 5% relative humidity for about 15 min. After that, batches of approximately 1000 cold-shock anesthetized sterile males were transferred into small plastic cups of 350 ml volume and 10 cm diameter (5 cm height) that were stacked vertically in cylindrical cardboard containers, and placed inside a polystyrene container with enough phase-changing materials (PCM) to keep the temperature at approximately 12 °C, and then transported by express courier service from the production facility (CAA) to Athens (Mastronikolos et al.)45. As soon as they arrived, sterile males were transferred to the release site.
Coating of sterile males with PPF
In the laboratory, a powder mixture was prepared containing 40% PPF (purchased by Sigma Aldrich, > 98% purity) and 60% fluorescent dye powder (fluorescent powder mix, DayGlo, Cleveland, USA). Plastic cups of 100-mL were coated with 10 mg of this formulation for each 1000 adult males, following the PPF dosage applied to Ae. albopictus and Ae. aegypti in boosted SIT trials in Spain and La Reunion, respectively32. To ensure that the dust mixture adheres to the walls of the dusting container, the inside surfaces were rubbed with sandpaper to create a rough surface. The container was then shaken vigorously to coat the inner surfaces evenly. In the release site, batches of 1000 cold-shock anesthetized sterile males were immediately introduced into the plastic containers, which were smoothly rotated for 30 s (ca. 25 rotations) to coat the sterile males uniformly.
Release of PPF-coated sterile males
The releases of PPF-coated sterile males were made on a roughly regular grid of 25 permanent release stations covering the treated site in Vravona area (Fig. 2), with a release density ranging from 2333 to 4679 PPF-coated sterile males/ha/week (Table 1). PPF-coated sterile males were released on a weekly basis at the beginning (six releases in the period 12 May to 16 June 2023) and at the peak (six releases in the period 1 September to 6 October 2023) of Ae. albopictus activity as previously determined for the study area in Vravrona by Stefopoulou et al.7. Batches of 1000 PPF-coated sterile males were released to cup-shaped paper containers (diameter of upper base 12 cm and lower base 11 cm, 8 cm height) that were hung by rope on fences and tree branches. At each release date the paper cups were evenly distributed among release stations to achieve uniform distribution of PPF-coated sterile males in the treated site. The inner surface of each container was previously scratched to provide the sterile male mosquitoes with a place to rest. The hanging rope was coated with vaseline (Vaseline Original Pure Petroleum Jelly, Unilever, London, UK) to prevent any access to predatory insects (e.g., ants). All releases were conducted approximately 1.5 h after receipt of shipment from Italy (usually 5–7 p.m.). In the release site, soon after coating the sterile males with the powder mixture, batches of 1000 anesthetized PPF-coated sterile males were transferred from the plastic PPF-coating containers to the release paper container cups and left there for 2 h to regain their fly activity. PPF-coated sterile males that died during the release process or were unable to fly from the paper containers were removed and counted.

Spatial coverage of release stations of boosted sterile males during the field trial implemented in Vravrona (Greece) in 2023. Basemap–Satellite image: Esri World Imagery (ArcGIS Pro 3.0.0), available from Esri (Marathon Data, GR).
Between the two release cycles (May–June) and (September–October), a Mark-Release-Recapture (MRR) trial was implemented in the treated site including two mass releases of PPF-coated sterile males to assess the flight distance capacity and longevity of boosted sterile males, on 14 and 21 July (results not presented herein). In the MRR releases, 10,000 boosted-sterile males were released in the center and the rest were evenly distributed among the 25 releasing stations of the treated area. Table 1 summarizes the weekly releases, including the quantity of boosted-sterile males delivered per week, the associated mortality rate, the contamination by females, and the estimated density of released boosted-sterile males per hectare.
Evaluation of the efficacy of boosted SIT application
The efficacy of boosted-SIT against Ae. albopictus was evaluated by a network of permanent sampling stations consisting of ovitraps and Human Landing Catches (HLC). The spatial coverage of sampling stations for each sampling method on the treated site is presented in Fig. 3. A network of 20 ovitraps in the treated site and 7 ovitraps in the control site was set up on 5 May 2023. Each ovitrap consisted of a 1.5 L black plastic container filled with 1.0 L water with a wooden strip (tongue depressor) measuring 150 × 18 × 1.6 mm as oviposition substrate. The oviposition substrates were collected weekly, until 11 November 2023, and transferred to the laboratory for egg counting and egg hatching rate following the step wise procedure as outlined by Balatsos et al.24, by adopting established protocols for egg transportation16 and egg hatching25. In the laboratory, the oviposition substrates were kept at 25 ± 1 °C, 80% RH, 14:10 L:D for one day and then placed in a container with a saturated solution of potassium sulphate (K2SO4) for a minimum of six days to complete embryonation. Then, the eggs were submerged for 20–24 h in 700 mL of deionised water containing 2 mL of hatching solution [12.5 g broth (Nutrient Broth OXOID) + 2.5 g yeast powder/100 mL of deionised water] and egg hatching rate was determined by counting the hatched and unhatched eggs under stereomicroscope.

Spatial coverage of entomological collections implemented in 2023, in the outskirts of Athens (Greece) to assess the efficacy of a boosted SIT to suppress wild Aedes albopictus populations monitored with oviposition traps (eggs), or Human Landing Catches (adults). The background map was retrieved from the Google Maps Platform https://mapsplatform.google.com/, using functions available in the ggmap package for R https://github.com/features/packages version 4.0.0, together with a private API key.
The collection of Ae. albopictus adults with HLC-technique was performed 1 and 6 days after each release date, from 5:00 p.m. to 7:00 p.m., at 10 permanent sampling stations in the treated site and 5 permanent sampling stations in the control site. At each sampling station, adult males and females approaching an operator with exposed limbs were collected for 5 min using a manual battery-operated aspirator. A total of 10 operators, rotated at each sampling date, were employed to prevent possible bias related to individual attractiveness on mosquito catches. The mosquitoes were transferred separately from each sampling station to plastic containers for further identification and counting in the laboratory. All Ae. albopictus specimens collected with the HLC-technique were meticulously examined under stereoscope with UV-light source to detect the fluorescent dust marking, and sorted as colored PPF-coated sterile males, non-colored wild males and non-colored wild females.
Data analysis
Relative mosquito population density
The observed mosquito density—i.e., trap-level mosquito count \({y}_{i}\), was standardized into relative density (noted \(\Theta\)) to discard the effect of population size, and to remove any time trend in relative density, other than the exposure to boosted-SIT, for example the influence of meteorological variables such as rainfall and day/night temperature on Ae. albopictus population dynamics. The expected density \({E}_{i}\) for exposure period \(i\), was the average of trap-level observed densities in the control site during the same period. The relative density \({\Theta }_{i}\) for exposure period \(i\) was the trap-level observed density, divided by the expected density: \({\Theta }_{i}={y}_{i}/{E}_{i}\). Data were grouped by time periods (approx. 1 month each) to avoid unstable estimates for the relative density.
Separate analyses were conducted for eggs and adults. To compare the efficacy of boosted-SIT on eggs and adults, the analysis was restricted to the common treatment exposure periods from May to November. Figure 4 shows the temporal coverage divided in 5 periods of entomological surveys collecting eggs using ovitraps and adults using HLC, and releases of PPF-coated sterile males in Vravrona area, Attica Region, in 2023, to assess the efficacy of boosted-SIT in suppressing Ae. albopictus populations. Once disseminated in the breeding sites, pyriproxyfen can have a long-lasting effect as an inhibitor of larval growth46,47. The persistent effect of boosted-SIT was evaluated with data collected during periods 2, 3, and 5.

Time coverage of entomological collections and releases of sterile males, implemented in the outskirts of Athens (Greece) in 2023 to assess the efficacy of a boosted-SIT to suppress Aedes albopictus populations. Mosquito population was monitored with ovitraps (eggs), and Human Landing Catches (adults).
The five periods were defined as follows:
-
1.
12/05–17/06/2023: population suppression during the 1st release cycle at the beginning of mosquito activity
-
2.
18/06–14/07/2023: persistent population suppression following the 1st release cycle
-
3.
30/07–01/09/2023: population suppression of a mark-release-recapture experiment implemented in mid-July
-
4.
02/09–07/10/2023: population suppression during the 2nd release cycle at the peak of mosquito activity
-
5.
08/10–10/11/2023: persistent population suppression following the 2nd release cycle
In addition, the expected density Ei (with i being the index for the period) was itself standardized to force the relative density \({\Theta }_{i}\) to be 1 for the first period. To this end, each Ei was multiplied by a factor \(k = \overline{y}_{1} /E_{1}\), with \(\overline{y}_{1}\) the average observed density in the treated site during period 1.
Egg hatching rate and competitiveness of sterile males
The egg hatching rate and its 95% credible interval was estimated with a Bayesian mixed-effect binomial logistic regression of the observed ratio of the number of hatching eggs to the total number of eggs corrected with the number of damaged eggs. This ratio was further used to estimate the competitiveness of sterile males, that is, their capacity to mate with fertile females in the presence of wild males48,49.
with \(R\) the estimated ratio of sterile to wild males. Bouyer & Vreysen49 showed that when this ratio is greater than four, the estimator of competitiveness provides strongly biased estimates. When this situation was met, \({C}_{i}\) was set to NA, as well as its credible limits.
Statistical model
The same statistical model was fitted for eggs and adults. The modeling approach was taken from Moraga50. A mixed-effect Poisson model of mosquito density was used to assess the efficacy of boosted SIT on Ae. albopictus density, and its spatial variations. For the observed density \(y_{i} \left( {i = 1, \ldots ,I:{\text{exposure period}}} \right)\), the spatial Poisson model was:
with \(E_{i}\), and \(\Theta_{i}\) the expected and relative density. For the trap \(j = 1, \ldots ,J\), the model was:
-
\(\eta_{i,j}\) was the linear predictor for trap \(j\) at exposure \(i\),
-
\(x_{k} \cdot (k = 2, \ldots ,I)\) was an indicator variable taking the value 1 for exposure \(k\), and 0 elsewhere,
-
\(b_{1}\) was the intercept i.e., the fixed-effect coefficient corresponding to the reference exposure \(x_{1}\), and \(b_{k}\) was the fixed-effect coefficient associated with \(x_{k}\),
-
\(u_{j}\) was the trap-level realization of the spatial random effect \(u\), with \(\mathop \sum \limits_{j = 1}^{J} u_{j} = 0\), and variance \(\sigma_{u}^{2}\).
With this parameterization, associated with the standardization of the expected density explained in the former section, the fitted relative density for exposure period \(i\) was an estimate of the suppression of the population of Ae. albopictus. It was the average of the exponentiated linear predictors for the \(J\) traps in the treated site:
A modified version of the Besag-York-Mollié model51,52 was used to estimate the variance of \(u\), \(\sigma_{u}^{2}\). To this end, \(u\) was split into (i) a spatially-structured component with a conditional auto-regressive (CAR) distribution, and (ii) a purely-random, unstructured, white noise component. Goodness-of-fit for these models is shown on SI Fig. S3 for eggs, and Fig. S4 for adults. Observed and fitted means were quite close. Plots of Pearson’s residuals did not reveal alarming trends. Model coefficients were estimated in a Bayesian framework with integrated nested Laplace approximation (INLA): the Bayesian process was approximated by analytical solutions, allowing a quick convergence53. The spatial Poisson model was fitted with the function ‘inla’ in the R package INLA53, with these settings:
-
Input data were not aggregated, i.e., one line in the dataset corresponded to a single trapping session.
-
The expected density was defined by the argument ‘E’ in the call to ‘ìnla’.
-
Priors for the precision of model hyper-parameters (i.e., variance of the components of \(u\)), were provided by the BYM2 version of the spatial variance estimation procedure52.
The success probability of boosted SIT was used to identify traps associated with low mosquito density, with respect to their expected density \(E\). We compared the fitted relative density \(\hat{\Theta }\) to a pre-defined threshold \(\Theta^{ - } = 0.20\). To estimate this indicator, 10,000 samples were drawn in the posterior marginals of the fitted relative density, and the proportion of simulated \(\hat{\Theta }_{sim} \le \Theta^{ - }\) was computed: boosted SIT was successful if \(P\left( {\hat{\Theta }_{sim} \le \Theta^{ - } } \right) \ge 0.95\).
Persistence of the effect of boosted SIT
The assessment of persistence of boosted-SIT was based on the difference in relative density between periods 1 and 2, as well as between periods 4 and 5. Under the assumption of no persistence, we expected that the later relative density was not lower than the earlier one, that is we expected to find a non-positive difference in relative density (null hypothesis \({\mathcal{H}}_{0}\)). To test this assumption, 1000 sets of simulated relative densities were drawn in the posterior distribution of the fitted values. For each simulation, the differences in fitted relative density was computed between periods 1 and 2 (\({\delta }_{1-2}\)), as well as between periods 4 and 5 (\({\delta }_{4-5}\)). For each set of these 1000 simulated differences \({\delta }_{x,y}\), we computed its mean and 95% credible interval (defined by quantiles 2.5% and 97.5%), and \(p=P\left(\delta x,y<0\right)\). If \(p\ge 0.95\), \({\mathcal{H}}_{0}\) was accepted.
Results
At the 15-ha treated site in Vravrona area, Attica Region, a total of 727,914 PPF-coated sterile males of Ae. albopictus were released throughout the trial. The mean density of releases was 3466 ± 193 (S.E.M.) PPF-coated sterile males/ha per week (Table 1).
The egg hatching rates in the treated and control sites (groups), as well as mating competitiveness of boosted males, are shown in Table 2 and Fig. 5. The egg hatching rate in the treated site (56–57%) was lower than the control site (84–85%) during the release cycles and one month following the second release cycle. The boosted sterile-to-wild male ratios during the first (1st period) and second (4th period) release cycles were 2.76 and 15.16, respectively. The mating competitiveness of boosted-SIT males was 0.18 and 0.13 during the 1st and 2nd period and then increased to 0.46 at the 5th period, i.e., the month after the second release cycle.

(a) Egg hatching rates in the treated and control site and their 95% lower and upper credible limits, (b) boosted sterile male competitiveness index and its 95% lower and upper credible limits, and (c) boosted sterile-to-wild males ratio, recorded during a boosted SIT trial in Vravrona, Attica Region, Greece, from May to November 2023. Note: The estimated value of competitiveness index during the fourth period of exposure was associated with a high ratio between sterile and wild males, therefore this value is omitted.
The mean relative density of collected eggs was > 0.92 throughout the 5 designated exposure periods, i.e., < 8% egg population reduction in the treated site compared to the control site (Figs. 6, 7). A great spatial variability in relative density and probability of success in the reduction of both egg and adult population densities was noticed among the specific locations of the treated site (Figs. 7, 8 and SI Figs S1, S2). Notably, a spotted successful reduction of egg population in terms of relative densities (i.e., > 80% egg population reduction compared to control site) and/or probability of success (> 95% probability to reach > 80% population reduction compared to control) was observed particularly in southeastern locations of treated site during the first release cycle (1st period) and the subsequent month (2nd period) as well as during the second release cycle (4th period) and the subsequent month (5th period). The boosted-SIT had no effect (Θ = 1.00; P = 0) on adult population densities during the first release cycle (1st period) (Fig. 8 and SI Fig S2). A meaningful reduction of adult population in terms of relative densities (Θ = 0.07–0.15, i.e., a 85–93% reduction) and probability of success (P = 0.9) was observed in the treated site during the second release cycle (4th period) and one month following the cease of releases (5th period). The MRR releases did not affect the egg population densities but exerted a 40% overall reduction of adult population in the treated site the following exposure month (August).

Observed (a), and relative (b) density of Aedes albopictus eggs and adults sampled during a boosted SIT trial in Vravrona (n = 20 ovitraps/collection day) with Artemida (n = 10 ovitraps/collection day) as the control site, from May to December 2023. The relative density was the observed density in traps from the treated site, divided by the expected density, i.e., the standardized and average density from traps in the control site. The violin plot shows the distribution of density.

Efficacy of boosted SIT in suppressing the Aedes albopictus population of eggs in Vravrona (Attica, Greece) during a field trial from May to November 2023. Relative density (θ) of eggs, i.e., density of eggs in traps located in the treated site divided by the average density of eggs in the control site during the same period. The white-border polygons were defined by a Voronoi tessellation of trap locations. The background maps were retrieved from the Google Maps Platform https://mapsplatform.google.com/, using functions available in the ggmap package for R https://github.com/features/packages version 4.0.0, together with a private API key.

Efficacy of boosted SIT in suppressing the Aedes albopictus population of adults in Vravrona (Attica, Greece) during a field trial from May to November 2023. Relative density (θ) of adults, i.e., density of adults collected with HLC in the treated site divided by the average density of adults in the control site during the same period. The white-border polygons were defined by a Voronoi tessellation of trap locations. The background maps were retrieved from the Google Maps Platform https://mapsplatform.google.com/, using functions available in the ggmap package for R https://github.com/features/packages version 4.0.0, together with a private API key.
Table 3 summarizes the key-findings in the treated site of the current study regarding suppression of egg and adult population densities, egg hatching rates, boosted-SIT male competitiveness and boosted sterile-to-wild male ratio.
Regarding the assessment of persistence of the effect of boosted SIT on relative density of eggs:
-
between periods 1 and 2: \({\delta }_{1-2}=0.117\), 95% credible interval: [− 0.171; − 0.058], \(P\left({\delta }_{1-2}<0\right)>0.999\), hence we accepted the null assumption of no persistence;
-
between periods 4 and 5: \({\delta }_{4-5}=0.584\), 95% CI: [0.512; 0.648], \(P\left({\delta }_{4-5}<0\right)<0.001\), hence we rejected the null assumption of no persistence.
Regarding the assessment of persistence of the effect of boosted SIT on relative density of adults:
-
between periods 1 and 2: \({\delta }_{1-2}=0.404\), 95% credible interval: [0.276; 0525], \(P\left({\delta }_{1-2}<0\right)<0.001\), hence we rejected the null assumption of no persistence;
-
between periods 4 and 5: \({\delta }_{4-5}\)= 0.087, 95% CI: [0.071; 0.1015], \(P\left({\delta }_{4-5}<0\right)<0.001\), hence we rejected the null assumption of no persistence.
Discussion
The results presented herein demonstrate for the first time the feasibility of boosted-SIT for the suppression of Ae. albopictus in Greece by releasing sterile males coated with PPF in a phase-II small scale field trial. The boosted-SIT application resulted in significant reduction of either egg or adult densities compared to control in several distinct locations of the treated site. The great spatial variability in the reduction of egg and adult population densities may be attributed to inhomogeneity in microclimatic environments such as vegetation and the availability of breeding sites. No remarkable effect on egg density was noted in the treated site overall throughout the trial, while a significant decrease was limited to specific locations at the southeastern part of the treated site during the first release cycle from May until mid-June and the periods following the completion of release cycles. The windy weather that was often noticed at the northern part of the treated site may have drifted released boosted SIT-males to southern locations resulting to increased sterilization of wild females and dissemination of PPF into breeding sites. The southeastern locations were less vegetated with fewer potential natural breeding sites, hence likely infested with lower mosquito populations, and this may have accounted for the stronger suppression in egg densities observed there.
The effect of boosted-SIT on mosquito adults showed a different temporal and spatial pattern. During the first cycle of releases at the beginning of mosquito activity from May to mid-June, no effect was observed against adult population, while after stopping releases a significant reduction (40%) of adult population was recorded. Interestingly, a satisfactory success in suppressing the adult population in the whole treated site was noted during the second release cycle from 1 September until 6 October reaching 85% population reduction compared to control and this effect was significantly increased (93%) next month after the cease of releases until 11 November. The time delay for effective adult population reduction may be justified by the fact that the boosted-SIT does not act directly to wild adult population but actually to the offspring through sterilization of wild females by sterile males and inhibition of the growth of larvae and pupae to adult stage into the breeding sites due to the PPF action27. After the conclusion of releases on 6 October, the suppression of both adult and egg population continued and was even more intense the next month, indicating the residual activity of boosted-SIT on Ae. albopictus population. This persistent effect may be the combination of induced sterility in wild females as the standard SIT effect which is anticipated the following mosquito generation, along with the inhibition of mosquito adult emergence from the breeding sites due to PPF insecticidal long-lasting effect on immature stages22,27,46,47.
The overall efficacy of the boosted-SIT was more pronounced against adults than eggs of Ae. albopictus considering that the standard SIT as part of the boosted-SIT concept does not necessarily affect the egg density despite the induced sterility caused to wild females. In a standard SIT trial performed during the peak of mosquito activity (September–October) in the same area of smaller scale (5-ha) in 2018 an induced egg sterility of 40–84% did not result to a significant reduction of egg density23, a phenomenon that was also observed in Italy where based on several field pilot trials it was estimated that induced egg sterility below 50% may not lead to any reduction of Ae. albopictus’ egg population densities25. The pilot SIT trials in Italy were performed with continuous releases at the rate of 896–1590 SIT males/ha/week, while in the current trial the releases of boosted-SIT males (3466/ha/week) were interrupted during mosquito season. The standard SIT trial in 2018 and the current boosted-SIT trial in the same area were not performed throughout the mosquito season, and this may have contributed to their ineffectiveness to mosquito egg population. However, the phase-II standard SIT trial that was performed at 10-ha of the same study area of Vravrona in 2019 by weekly releases of sterile Ae. albopictus males from May to September, gave > 50% reduction of egg density in September, reaching 78%, while the reduction of egg density continued within a range of 70–78% for up to 4 weeks after the completion of the sterile male releases24. It is noted that the standard SIT trial in 2019 was performed with continuous weekly releases throughout the season of mosquito activity from May to September, whereas in our study the releases were interrupted during the summer. Although the three SIT field trials in 2018, 2019, and 2023 in Greece were conducted in the same study area of Vravrona, there were differences in the test designs e.g., the use or not of PPF, the number and season of releases, and the size of treated site, that may have contributed to differences in the efficacy against Ae. albopictus populations. The limited success of boosted-SIT against egg densities in our study may also be attributed to immigration of adult females of Ae. albopictus in the treated site from adjacent areas as noted in the boosted-SIT trial in Spain by Bouyer et al.32. Nevertheless, the effect of PPF on the survival of immature stages and adult emergence from breeding sites27 may have resulted to the significant reduction of Ae. albopictus adults compared to limited decline in egg densities. This protective action against immigration of viable eggs is a major benefit of the boosted-SIT.
In the current study, the hatching rate of eggs in the treated site was lower than in the control site throughout the trial, reaching 56% during the first and second release cycle versus 84% in the control site, and 57% one month after the second release cycle versus 85% in the control site. Similarly, Balatsos et al.24, reported a significant decrease in the egg hatching rate in the standard SIT plot compared to the control plot, with an average rate of induced sterility of 49%. The reduction of egg hatching rate compared to control in the current boosted-SIT trial is close to the 50% reduction observed during other field SIT trials23,24,38,54,55,56, but it seems to be lower than the egg hatching reduction level of 70–80% commonly associated with successful population suppression SIT trials24,25,38. Nevertheless, despite the considerable reduction in egg hatching rates, no remarkable overall suppression of egg population was observed in the treated site of the current study. This finding aligns with observations from standard SIT pilot field studies in Italy25 where egg densities were not clearly affected when induced egg sterility was below 50%, and a positive correlation between egg densities and egg hatching rate was observed when induced egg sterility was 70–80%. The performance of boosted-SIT with the release of PPF-coated Ae. albopictus males in our study in Greece resembles to the boosted-SIT applied in a 35-ha area in La Vilavella, Valencia, Spain in 202132. Following the same methodology for data analysis, the mean success rate in suppression of egg population of Ae. albopictus (i.e., probability to achieve > 80% reduction compared to control) at the treated site in Spain, was considerably low (P = 0.11–0.29) over the exposure periods like in our field trial in Greece (P = 0.05–0.20 for exposure periods 1,2,4 & 5). However, in both countries the boosted SIT resulted in successful egg population reduction (P > 0.95) in specific locations of the treated sites. Although the effect of boosted-SIT on Ae. albopictus adult population recorded differently in Spain than in Greece, i.e., emerged adults from ovitraps in Spain Vs adults collected with HLC in Greece, in the trials of both countries the method was more efficient in suppressing adult than egg population of Ae. albopictus. The effect of boosted-SIT against adult population in Spain was more intense at the beginning of releases and mosquito activity in June reaching 98% population suppression while in Greece significant suppression of adult population was observed in September–October. Differences in the performance of the same method between the two countries may be attributed to differences in the quantity (2712 and 3466 boosted-SIT males/ha/week in Spain and Greece, respectively) and frequency of releases (3 releases per week continuously in Spain Vs weekly release interrupted during summer in Greece) and in boosted sterile-to-wild male ratio during the releases (2.76–15.16 in Greece Vs 6.0–62.2 in Spain).
The efficiency of boosted-SIT in the current trial in Greece reaching 85–93% suppression of adult population is good compared to standard SIT phase-II trials against Ae. albopictus worldwide like in China where 40% reduction of female population was achieved in a 1.2-ha area by releasing 166,666 sterile males/week/ha57, in Croatia where 45% adult reduction was achieved in a 30-ha area by releasing 2000 sterile males/week/ha22, in Mauritius where 63.6% female reduction was achieved by releasing 20,000 sterile males/week/ha58, in Sri Lanka where 95.5% female reduction was achieved in a 30-ha area by releasing 3300 sterile males/week/ha22, and in Switzerland where 66.7% reduction of female population was achieved in a 45-ha area by releasing 3000 sterile males/week/ha22. The high suppression of adult population that was observed during the second release cycle and one month later complies with the considerably high boosted sterile-to-wild male ratio (15.16) and mating competitiveness index of boosted-SIT males (0.46) that were recorded for those two exposure periods, respectively. For standard SIT programs against Aedes spp. a minimum sterile-to-wild male ratio of 10:1 is recommended for efficient reduction of mosquito populations59. Moreover, a minimum value of 0.2 for sterile male competitiveness index (i.e., 5 times more likely for a female to be mated with a wild male than a boosted-SIT male), has been proposed by Bouyer and Vreysen49 to conduct cost-efficient pilot phase-II SIT field trials. SIT or SIT/IIT field projects with successful suppression of Ae. albopictus population in China39, Italy25, Mauritius58 and Albania60 reported a sterile male competitiveness value above the threshold of 0.2. In addition, the two MRR releases in mid-June causing an overall 40% reduction in adult population in August, may have also contributed to the reduction of population densities in September–October.
For the current study some limitations related primarily to test design are acknowledged. As a result of limited resources, the current study design did not include a control standard SIT site to compare the standard-SIT with boosted-SIT and assess the contribution of PPF. Hence, our study aimed to assess the potential of the boosted-SIT overall, i.e., SIT plus PPF dissemination, against Ae. albopictus by comparison with an untreated control site. Further studies are deemed necessary to determine the dissemination of PPF to breeding sites and its contribution to the standard SIT on the suppression of mosquito population densities. Due to limited resources, we followed an interrupted release schedule of boosted-SIT males targeting the beginning (May–June) and the peak (September–October) of Ae. albopictus activity. Nevertheless, continuous releases throughout the mosquito season may have resulted in more intense, sustained, and even suppression of Ae. albopictus densities in the study area. Another limitation is the difference in the size of the treated (15 ha) and control (5 ha) sites, which may have affected the comparability of mosquito populations. This potential study bias is, however, mitigated by the fact that the treated and control sites are similar semi-urban, coastal, green areas. In addition, a considerable mortality was recorded for SIT males (23% on average) due to shipment, which is slightly higher than the previously reported23,24,45 mortality of SIT males (approx. 16%) transported from mosquito rearing facilities of CAA in Italy to Greece. Mortality of SIT-males due to shipment may have affected their quality, particularly in terms of longevity, as previously reported by Balatsos et al.61. Reduced lifespan of transported sterile males may trigger increasing the numbers of released boosted-SIT males to achieve an effective ratio of boosted sterile-to-wild male ratio and adequately competitive boosted-SIT males in the field.
In conclusion, the current phase-II small scale field trial conducted for the first time in Greece indicated the potential of boosted-SIT against Ae. albopictus, particularly in suppressing mosquito adult population. A significant persistent effect after the end of releases against both adult and egg population densities was also observed, underscoring the residual effect of the method by sterilizing wild females via standard SIT and affecting the survival of immature stages in breeding sites by the PPF larvicidal action. Further laboratory, semi-field and wider scale field studies are deemed necessary to substantiate the added value of PPF when combined with the standard SIT to support the application of boosted-SIT as an alternative integrated mosquito management method against Ae. albopictus. For upscaling the boosted-SIT to pre-operational phase III in Greece, the most critical step forward, which is currently explored, is a large investment for mass rearing and mass sterilization facilities, including quality control procedures, and subsequently the deployment of the intervention to wider geographical scale with continuous season-long releases, and the performance of cost-effectiveness studies. Then, the most important activity towards operational phase IV is the establishment of a sustainable management plan, preferably autonomous, for implementing the intervention taking decisions based on the continuous collection of entomological and epidemiological data from the target area62. Within the framework of the IAEA TC Project RER/5/022, Greece, Albania, and Croatia have also received basic equipment for mass production to support the generation of sterile males for research purposes and small-scale field trials. Thus, potential regulatory and public acceptance issues with releasing pesticide-coated insects should be also explored when planning to move towards operational phase. With careful consideration of all available mosquito-control methods, including source reduction and community engagement, boosted-SIT should be viewed as a part of an area-wide integrated vector management strategy that minimizes risks to the environment and human health.
Data availability
The datasets used for the efficacy assessment are provided within the supplementary information files.
References
Becker, N. et al. Mosquitoes: Identification Ecology and Control (Springer International Publishing, Cham, 2020).
Wilkerson, R. C., Linton, Y.-M. & Strickman, D. Mosquitoes of the World (Johns Hopkins University Press, Baltimore, 2021).
Adhami, J. & Reiter, P. Introduction and establishment of Aedes (Stegomyia) albopictus skuse (Diptera: Culicidae) in Albania. J. Am. Mosq. Control Assoc. 14, 340–343 (1998).
ECDC. Aedes albopictus—Current known distribution: July 2024. https://www.ecdc.europa.eu/en/publications-data/aedes-albopictus-current-known-distribution-july-2024 (2024).
Giatropoulos, A. et al. Records of Aedes (Stegomyia) cretinus in Greece before and after the invasion of Aedes (Stegomyia) albopictus (Diptera: Culicidae). Eur. J. Entomol. 121, 199–205. https://doi.org/10.14411/eje.2024.022 (2024).
Badieritakis, Ε et al. Aedes albopictus (Skuse, 1895) (Diptera: Culicidae) in Greece: 13 years of living with the Asian tiger mosquito. Parasitol. Res. 117, 453–460. https://doi.org/10.1007/s00436-017-5721-6 (2018).
Stefopoulou, A. et al. Knowledge, attitude, and practices survey in Greece before the implementation of sterile insect technique against Aedes albopictus. Insects 12, 212 (2021).
ECDC. Dengue—Annual Epidemiological Report for 2022. https://www.ecdc.europa.eu/en/publications-data/dengue-annual-epidemiological-report-2022 (2024).
Kraemer, M. U. G. et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 4, 854–863. https://doi.org/10.1038/s41564-019-0376-y (2019).
Medlock, J. M. et al. An entomological review of invasive mosquitoes in Europe. Bull. Entomol. Res. 105, 637–663. https://doi.org/10.1017/S0007485315000103 (2015).
Caputo, B. et al. The “auto-dissemination” approach: Approach: A novel concept to fight Aedes albopictus in urban areas. PLoS. Negl. Trop. Dis. 6, e1793 (2012).
Medlock, J. M. et al. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 12, 435–447. https://doi.org/10.1089/vbz.2011.0814 (2012).
Achee, N. L. et al. Alternative strategies for mosquito-borne arbovirus control. PLoS Negl. Trop. Dis. 13, e0006822. https://doi.org/10.1371/journal.pntd.0006822 (2019).
Corbel, V. et al. Tracking insecticide resistance in mosquito vectors of arboviruses: The worldwide insecticide resistance network (WIN). PLoS Negl. Trop. Dis. 10, e0005054. https://doi.org/10.1371/journal.pntd.0005054 (2016).
Qi, Z. et al. Effects of prenatal exposure to pyrethroid pesticides on neurodevelopment of 1-year- old children: A birth cohort study in China. Ecotoxicol. Environ. Saf. 234, 113384. https://doi.org/10.1016/j.ecoenv.2022.113384 (2022).
Bellini, R. et al. Practical management plan for invasive mosquito species in Europe: I. Asian tiger mosquito (Aedes albopictus). Travel Med. Infect. Dis. 35, 101691. https://doi.org/10.1016/j.tmaid.2020.101691 (2020).
Haramboure, M. et al. Modelling the control of Aedes albopictus mosquitoes based on sterile males release techniques in a tropical environment. Ecol. Model. 424, 109002. https://doi.org/10.1016/j.ecolmodel.2020.109002 (2020).
Lees, R. S., Carvalho, D. O. & Bouyer, J. Potential Impact of Integrating the Sterile Insect Technique into the Fight Against Disease-Transmitting Mosquitoes. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management (eds Dyck Victor, A. et al.) 1081–1118 (CRC Press, Boca Raton, 2021).
Benedict, M. Q. Sterile insect technique: Lessons from the past. J. Med. Entomol. 58, 1974–1979. https://doi.org/10.1093/jme/tjab024 (2021).
Bellini, R. et al. Use of the Sterile Insect Technique Against Aedes albopictus in Italy: First Results of a Pilot Trial. In Area-Wide Control of Insect (eds Pests Vreysen, M. J. B. et al.) 505–515 (Springer, Netherlands, Dordrecht, 2007). https://doi.org/10.1007/978-1-4020-6059-5_47.
WHO & IAEA. Guidance framework for testing the sterile insect technique as a vector control tool against aedes-borne diseases. https://www.who.int/publications/i/item/9789240002371.
Bouyer, J. Current status of the sterile insect technique for the suppression of mosquito populations on a global scale. Infect. Dis. Poverty 13, 68. https://doi.org/10.1186/s40249-024-01242-z (2024).
Balatsos, G. et al. Reduction in egg fertility of Aedes albopictus mosquitoes in Greece following releases of imported sterile males. Insects 12, 110. https://doi.org/10.3390/insects12020110 (2021).
Balatsos, G. et al. Sterile insect technique (SIT) field trial targeting the suppression of Aedes albopictus in Greece. Parasite 31, 17. https://doi.org/10.1051/parasite/2024020 (2024).
Bellini, R., Medici, A., Puggioli, A., Balestrino, F. & Carrieri, M. Pilot field trials with Aedes albopictus irradiated sterile males in Italian urban areas. J. Med. Entomol. 50, 317–325. https://doi.org/10.1603/me12048 (2013).
Tur, C. et al. Suppression trial through an integrated vector management of Aedes albopictus (Skuse) based on the sterile insect technique in a non-isolated area in Spain. Insects 14, 688. https://doi.org/10.3390/insects14080688 (2023).
Bouyer, J. & Lefrançois, T. Boosting the sterile insect technique to control mosquitoes. Trends Parasitol. 30, 271–273. https://doi.org/10.1016/j.pt.2014.04.002 (2014).
Chandel, K. et al. Targeting a hidden enemy: pyriproxyfen autodissemination strategy for the control of the container mosquito Aedes albopictus in Cryptic habitats. PLoS Negl. Trop. Dis. 10, e0005235 (2016).
Devine, G. J. et al. Using adult mosquitoes to transfer insecticides to Aedes aegypti larval habitats. Proc. Natl. Acad. Sci. USA. 106, 11530–11534. https://doi.org/10.1073/pnas.0901369106 (2009).
Mains, J. W., Brelsfoard, C. L. & Dobson, S. L. Male mosquitoes as vehicles for insecticide. PLoS Negl. Trop. Dis. 9, e0003406. https://doi.org/10.1371/journal.pntd.0003406 (2015).
Pleydell, D. R. J. & Bouyer, J. Biopesticides improve efficiency of the sterile insect technique for controlling mosquito-driven dengue epidemics. Commun. Biol. 2, 201; https://www.nature.com/articles/s42003-019-0451-1 (2019).
Bouyer, J. et al. Suppression of Aedes mosquito populations with the boosted sterile insect technique in tropical and mediterranean urban areas. Sci. Rep. 15, 17648. https://doi.org/10.1038/s41598-025-02795-1 (2025).
Bouyer, J., Iyaloo, D. & Baldet, T. Preventing the establishment of invasive exotic mosquitoes. Trends Parasitol. https://doi.org/10.1016/j.pt.2025.06.001 (2025).
Martín-Park, A. et al. Pilot trial using mass field-releases of sterile males produced with the incompatible and sterile insect techniques as part of integrated Aedes aegypti control in Mexico. PLoS Negl. Trop. Dis. 16, e0010324. https://doi.org/10.1371/journal.pntd.0010324 (2022).
Kittayapong, P. et al. Combined sterile insect technique and incompatible insect technique: The first proof-of-concept to suppress Aedes aegypti vector populations in semi-rural settings in Thailand. PLoS Negl. Trop. Dis. 13, e0007771. https://doi.org/10.1371/journal.pntd.0007771 (2019).
Mains, J. W., Brelsfoard, C. L., Rose, R. I. & Dobson, S. L. Female adult Aedes albopictus suppression by Wolbachia-infected male mosquitoes. Sci. Rep. 6, 33846 (2016).
Mains, J. W., Kelly, P. H., Dobson, K. L., Petrie, W. D. & Dobson, S. L. Localized control of Aedes aegypti (Diptera: Culicidae) in Miami, FL, via inundative releases of Wolbachia-infected male mosquitoes. J. Med. Entomol. 56, 1296–1303. https://doi.org/10.1093/jme/tjz051 (2019).
Gato, R. et al. Sterile insect technique: Successful suppression of an Aedes aegypti field population in Cuba. Insects 12, 469. https://doi.org/10.1093/jme/tjz051 (2021).
Zheng, X. et al. Incompatible and sterile insect techniques combined eliminate mosquitoes. Nature 572, 56–61. https://doi.org/10.1038/s41586-019-1407-9 (2019).
Malfacini, M. et al. Aedes albopictus sterile male production: Influence of strains, larval diet and mechanical sexing tools. Insects 13, 899. https://doi.org/10.3390/insects13100899 (2022).
Malfacini, M. et al. Effect of 2 sex-sorting time schedules on SIT facility management. J. Insect Sci. 23, 9. https://doi.org/10.1093/jisesa/iead060 (2023).
Balestrino, F., Puggioli, A., Gilles, J. R. L. & Bellini, R. Validation of a new larval rearing unit for Aedes albopictus (Diptera: Culicidae) mass rearing. PLoS ONE 9, e91914. https://doi.org/10.1371/journal.pone.0091914 (2014).
Maiga, H., Mamai, W., Yamada, H., Argilés Herrero, R. & Joint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture, V. (Austria). Guidelines for Mass-Rearing of Aedes Mosquitoes. Version 1. https://inis.iaea.org/records/ssx7t-tar88 (2020).
Balestrino, F. et al. γ ray dosimetry and mating capacity studies in the laboratory on Aedes albopictus males. J. Med. Entomol. 47, 581–591. https://doi.org/10.1603/me09272 (2010).
Mastronikolos, G. D. et al. Quality control methods for Aedes albopictus sterile male transportation. Insects 13, 179. https://doi.org/10.3390/insects13020179 (2022).
Ohba, S. et al. The effect of pyriproxyfen as a “population growth regulator” against Aedes albopictus under semi-field conditions. PLoS ONE 8, e67045. https://doi.org/10.1371/journal.pone.0067045 (2013).
Hustedt, J. C., Boyce, R., Bradley, J., Hii, J. & Alexander, N. Use of pyriproxyfen in control of Aedes mosquitoes: A systematic review. PLoS Negl. Trop. Dis. 14, e0008205. https://doi.org/10.1371/journal.pntd.0008205 (2020).
Fried, M. Determination of sterile-insect competitiveness. J. Econ. Entomol. 64, 869–872. https://doi.org/10.1093/jee/64.4.869 (1971).
Bouyer, J. & Vreysen, M. J. B. Yes, irradiated sterile male mosquitoes can be sexually competitive!. Trends Parasitol. 36, 877–880. https://doi.org/10.1016/j.pt.2020.09.005 (2020).
Moraga, P. Geospatial Health Data: Modeling and Visualization with R-INLA and Shiny (Chapman and Hall/CRC, New York, 2019).
Besag, J., York, J. & Mollié, A. Bayesian image restoration, with two applications in spatial statistics. Ann. Inst. Stat. Math. 43, 1–20. https://doi.org/10.1007/BF00116466 (1991).
Riebler, A., Sørbye, S. H., Simpson, D. & Rue, H. An intuitive Bayesian spatial model for disease mapping that accounts for scaling. Stat. Methods Med. Res. 25, 1145–1165. https://doi.org/10.1177/0962280216660421 (2016).
Rue, H. et al. Bayesian Computing with INLA: A Review. Annu. Rev. Stat. Appl. 4, 395–421. https://doi.org/10.1146/annurev-statistics-060116-054045 (2017).
Gouagna, L. C. et al. Strategic approach, advances, and challenges in the development and application of the SIT for area-wide control of Aedes albopictus mosquitoes in Reunion Island. Insects 11, 770. https://doi.org/10.3390/insects12030191 (2020).
Becker, N. et al. Integrated control of Aedes albopictus in southwest Germany supported by the sterile insect technique. Parasit. Vectors 15, 9. https://doi.org/10.1186/s13071-021-05112-7 (2022).
Bouyer, J. When less is more: Accounting for overcompensation in mosquito SIT projects. Trends Parasitol. 39, 235–237. https://doi.org/10.1016/j.pt.2023.02.001 (2023).
Zhang, D. et al. Mating harassment may boost the effectiveness of the sterile insect technique for Aedes mosquitoes. Nat. Commun. 15, 1980. https://doi.org/10.1038/s41467-024-46268-x (2024).
Iyaloo, D. P., Bouyer, J., Facknath, S. & Bheecarry, A. Pilot suppression trial of Aedes albopictus mosquitoes through an Integrated Vector Management strategy including the Sterile Insect Technique in Mauritius. Preprint at https://doi.org/10.1101/2020.09.06.284968 (2020).
Oliva, C. F. et al. Sterile Insect Technique (SIT) against Aedes species mosquitoes: A roadmap and good practice framework for designing, implementing and evaluating pilot field trials. Insects 12, 191. https://doi.org/10.3390/insects12030191 (2021).
Velo, E. et al. A mark-release-recapture study to estimate field performance of imported radio-sterilized male Aedes albopictus in Albania. Front. Bioeng. Biotechnol. 10, 833698. https://doi.org/10.3389/fbioe.2022.833698 (2022).
Balatsos, G. et al. Residual longevity of recaptured sterile mosquitoes as a tool to understand field performance and reveal quality. Insects 15, 826. https://doi.org/10.3390/insects15110826 (2024).
Bouyer, J., Yamada, H., Pereira, R., Bourtzis, K. & Vreysen, M. J. B. Phased conditional approach for mosquito management using sterile insect technique. Trends Parasitol. 36, 325–336. https://doi.org/10.1016/j.pt.2020.01.004 (2020).
Acknowledgements
We thank the City Council of the Municipality of Markopoulo Mesogaias (Greece) for granting permission to conduct our field experiment in the Vravrona area. We are also grateful to Evangelia Zavitsanou for her valuable support in maps’ creation and we sincerely thank the reviewers for their constructive comments, which helped to improve the quality and clarity of this work.
Biocide risk assessment
All procedures employed in the current study complied with international guidelines and with current national legislation. The laboratories involved in this project are regularly inspected by local officials to ensure that they comply with all relevant national and international laws, conventions, guidelines and codes of conduct and safety. As part of the implementation of Article 56 of EU Regulation 528/2012, we submitted a detailed risk assessment report on the potential effects on human and animal health, as well as environmental impact, from the field trial involving the release of biocide-treated sterile males and the current trial was approved by the Hellenic Ministry of Rural Development and Food (Authorization No. ΑΔΑ: ΨΖΗΗ4653ΠΓ-Γ1Ξ).
Institutional review board statement
The strain of Aedes albopictus used in this study was established using mosquito eggs collected from ovitraps in the Vravrona area (Markopoulo Mesogaias, Greece). The collection areas were public and not privately owned or protected. Mosquito egg collections from the field did not involve endangered or protected animal species. Consequently, the establishment of the laboratory mosquito strain did not require a specific permit. The Ethics Committee of Benaki Phytopathological Institute concluded that the current study was conducted in accordance with the Ethics Code for Research; therefore, no special permit was required.
Funding
This study was supported by the projects: A) the project “moSquITo: Innovative approaches for monitoring and management of the Asian tiger mosquito with emphasis on the Sterile Insect Technique (ΤΑΕΔΚ06173)”, financed by the National Recovery and Resilience Plan, “Greece 2.0” and EU Funding—Next Generation EU; B) the project entitled “A systematic surveillance of vector mosquitoes for the control of mosquito-borne diseases in the Region of Attica”, financed by the Region of Attica; C) the project “E4Warning: Eco-Epidemiological Intelligence for early Warning and response to mosquito-borne disease risk in Endemic and Emergence setting” (https://www.e4warning.eu) financed by Horizon Europe grant no. 01086640; D) the TC Project RER/5/022, “Establishing Genetic Control Programmes for Aedes Invasive Mosquitoes”, and the programme of Coordinated Research Activities, “Quality control bioassays for irradiated Aedes albopictus males” (CRP Contract No: 23915), financed by the IAEA.
Author information
Authors and Affiliations
Contributions
A.M., J.B., D.P.P and N.T.P. concepted research; A.G., G.B., J.B., N. T. P., D.P.P. and A.M. designed research; A.G., G.B., V.K., J.B., N. T. P., D.P.P., I.M. and A.M. performed research; J.B. and R.L. analyzed data; A.G., A.M., D.P.P., N.T.P., R.B., and A.P interpreted data; A.G., J.B., and R.L. wrote the first version of the paper; All authors corrected the final version of the paper.
Corresponding author
Ethics declarations
Competing interests
Jérémy Bouyer is co-founder and CEO of MoSITouch, a company offering vector control services, particularly the boosted sterile insect technique.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.




Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Giatropoulos, A., Balatsos, G., Karras, V. et al. Suppression of the Asian tiger mosquito (Aedes albopictus) populations using the boosted sterile insect technique in Greece. Sci Rep 15, 38738 (2025). https://doi.org/10.1038/s41598-025-22454-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-22454-9

{kind=link}
{kind=link}
{kind=link}
{kind=link}