
Abstract
Potassium (K) is essential for the productivity of tropical grasses, but its optimal supply remains unclear. This study evaluated the effects of K on gas exchange, photosynthetic rate (A), forage production (FP), and root development (RP) in Panicum maximum cultivars. The experiment was conducted using a randomized block design in a 6 × 4 factorial scheme (six cultivars: Tanzânia, Quênia, Mombaça, Zuri, Massai and Tamani; and four K rates: 0, 205, 410 and 820 mg dm−3). There was interaction between cultivars and K rates for A and stomatal conductance (gs), with linear increases in A for all cultivars. The gs response in the Zuri cultivar was quadratic, with a maximum of 5.32 mmol m−2 s−1 at the dose of 410 mg dm−3 K, and linear for the other cultivars. The CO2 concentration (Ci) and leaf temperature (Tleaf) were not influenced by the K dose or by the cultivars, maintaining an average of 129.28 ppm and 29.32 °C, respectively. Transpiration (E) increased by 0.018 mmol m−2 s−1 with increasing K doses. The chlorophyll content fitted a quadratic model, with a maximum of 35 SPAD at the dose of 530 mg dm−3. The FP increased linearly for all cultivars, with the highest FP in the cultivars Quênia, Mombaça and Zuri. Increasing K doses improved A and gs, reflecting in higher FP. The cultivars Quênia and Mombaça showed the greatest increases in FP up to 820 mg dm−3 K, while the maximum efficiency for RP occurred between 205 and 410 mg dm−3, suggesting that excessive K supply may not proportionally increase root development.
Similar content being viewed by others

Introduction
Modern agriculture prioritizes forage species that respond efficiently to soil fertility management1. However, the productive capacity of forage plants is directly influenced by the availability of growth factors such as nutrients and water1. These factors are also among the most influential in determining plant responses to climate change and their ability to cope with environmental stress2.
In this context, cultivars main of Panicum maximum—Tanzânia, Mombaça, Massai, Quênia, Zuri, and Tamani—meet the market demand for more productive plants with better nutritional quality. The Mombaça grass stands out for its high vegetative vigor and productivity3, while Tanzânia presents a canopy architecture more favorable to grazing, with a higher leaf-to-stem ratio and easier management4. The Massai cultivar, of short stature, has a high tiller density, strong resistance to pasture spittlebugs (Mahanarva spp.), good drought tolerance, and adaptation to soils of lower fertility, being especially recommended for small ruminants5,6. The Quênia cultivar, with intermediate height, combines tender stems and soft leaves with high productive potential, enhancing animal intake and pasture structure maintenance7. The Zuri cultivar is tall, with thick stems and broad leaves, in addition to resistance to leaf spot caused by Bipolaris maydis, making it suitable for intensive systems8. Finally, the Tamani cultivar, also of short stature, is notable for its high tiller density, good nutritional value, and flexible management, being appropriate for rotational grazing and crop-livestock integration systems7.
Among the essential nutrients for plants, potassium (K) plays a crucial role in physiological processes such as photosynthesis and stomatal regulation, which directly impact the growth and productivity of forage plants9, especially when grown in acidic and low-fertility soils, such as those of the Brazilian Cerrado10. This situation is especially relevant in sandy soils, such as Quartzarenic Neosols, which have low water and nutrient retention capacity, favoring leaching and hindering the availability of K for plants.
K deficiency can lead to significant economic losses, including reduced forage production caused by potential changes in photosynthesis and stomatal function, as well as visual symptoms such as chlorosis and leaf necrosis11,12. These changes result in reduced efficiency of CO2 absorption and tissue hydration, impairing plant metabolism13. Although the role of pot K in forage production is well recognized, knowledge about its physiological effects—such as on gas exchange and its relationship with forage and root production—remains limited for Panicum maximum grown in low-fertility soils, where potassium is deficient and other nutrient limitations may also be present. It is important to consider that the availability of one nutrient can influence the uptake and efficiency of others.
Thus, understanding the physiological changes promoted by K in forage plants becomes essential for the development of more efficient and sustainable management strategies, aiming to maximize productivity and ensure the sustainability of production systems in soils with macro and micronutrient deficiencies. Therefore, the hypothesis tested was that adequate K supply improves gas exchange and increases forage and root production in Panicum maximum cultivars. The objective of this study was to evaluate the effects of different doses of K on physiological and productive parameters in six cultivars of this species.
Results
Physiological variables
There was an interaction between cultivar and K dose for leaf photosynthesis rate (A) (P = 0.0001) and stomatal conductance (gs) (P = 0.0011) (Table 1). Increasing K doses resulted in a linear increase in A for all cultivars, with estimated increments of 0.287; 0.246; 0.207; 0.269; 0.238 µmol/m2s−1 in the cultivars Tanzânia, Quênia, Mombaça, Massai, and Tamani, respectively, for each mg dm−3 K added to the soil. The A of the Zuri cultivar was adjusted to a quadratic regression model, with a maximum point estimated at a dose of 205 mg dm-3.
Without K fertilization, there was no difference between cultivars for A. At the dose of 205 mg dm−3 K, the highest A was observed in the cultivar Zuri, the lowest in Mombaça and intermediate levels in the other cultivars. At the doses of 410 and 820 mg dm−3 K, the highest A was recorded in the cultivar Tanzânia. The lowest A at the dose of 410 mg dm−3 K were in the cultivars Mombaça and Zuri, and at the dose of 820 mg dm−3 K in the cultivar Zuri, with intermediate values in the other cultivars.
The gs fitted a linear increasing regression model in the cultivars Tanzânia, Quênia, Mombaça, Massai, and Tamani, with values of 0.025; 0.024; 0.023; 0.032; 0.024 mmol m−2 s−1, respectively. In the Zuri cultivar, gs followed a quadratic regression model, with the maximum point estimated at the dose of 446.5 mg dm−3 K, reaching 0.010 mmol m−2 s−1. There was a difference between cultivars only at the 820 mg dm−3 K dose, with the lowest gs observed in the Zuri cultivar, and no difference for the other cultivars. The Ci (internal CO2 concentration) did not fit regression equations, and no differences were observed between cultivars, with an average of 128.80 mmol m−2 s−1.
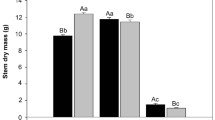
There was an effect of K doses on leaf transpiration (E) (P = 0.0001), and chlorophyll concentration (P = 0.0001) (Fig. 1), as well as an effect of cultivar on E (P = 0.0005) and chlorophyll concentration (P = 0.0019). The effect of K doses on E followed a linear regression model, with an estimated increase of 0.0018 mmol m−2 g−1 in E for each mg dm−3 K added to the soil. The chlorophyll concentration followed a quadratic regression model, with the maximum point estimated at a dose of 574.2 mg dm−3 K, with a concentration of 29.84 SPAD units per evaluated leaf. There was no effect of K doses (P = 0.1155) and cultivars (P = 0.2686) on leaf temperature, with an average of 29.32 °C.

Leaf transpiration (mmol m−2 g−1) (A); and SPAD chlorophyll index (B), in Panicum maximum cultivars in response to different potassium doses.
The highest E was observed in the Quênia and Mombaça cultivars, the lowest in the Massai and Tamani cultivars, and intermediate in the others. Higher chlorophyll concentrations (P = 0.0001) were estimated in the Massai cultivar, with lower concentrations in the others (Table 2).
Forage and root production
There was an interaction between cultivar and K dose for forage production (g/pot) (P = 0.0001) (Table 3). Increasing linear responses were observed in the cultivars Tanzânia, Quênia, Mombaça, Zuri, Massai, and Tamani, with estimated increases of 0.16; 0.20; 0.16; 0.14; 0.10; and 0.12 g of dry matter (g/pot), respectively, for each mg dm⁻3 K added to the soil. In the absence of K fertilization, the highest forage production was observed in the Mombaça cultivar, with lower productions in the Tânzania, Massai and Tamani cultivars. At doses of 205 and 820 mg dm⁻3 K, the highest forage productions were observed in the Quênia, Mombaça, and Zuri cultivars, while at the 410 mg dm⁻3 K dose, the highest forage productions were observed in the Quênia and Zuri cultivars, with the lowest in the Massai cultivar.
There was an interaction between cultivar and K dose for root production (P = 0.0001; Table 4), with estimated linear increments of 0.080; 0.156; 0.147; and 0.098 g per pot of roots for each mg dm−3 of K in the cultivars Quênia, Zuri, Massai, and Tamani, respectively. The root production of the Tanzânia and Mombaça cultivars followed a quadratic regression model, with the estimated maximum points at doses of 505 mg K dm−3 and 547 mg K dm−3, respectively.
In the absence of K fertilization, the highest root production was recorded in the cultivars Quênia, Mombaça, and Tamani, and the lowest in the others. At the 205 mg K dm−3 dose, the highest root production was in the cultivars Zuri and Tamani, and the lowest in Tanzânia. At the 410 mg K dm−3 dose, the highest production was in Mombaça and the lowest in the others, while at the 820 mg K dm−3 dose, the highest root productions were in the cultivars Mombaça, Zuri, Massai, and Tamani.
Discussion
The linear increase in A (Table 1) is explained by the high demand for K required by forage plants to activate enzymes involved in ATP production. The greatest stimulation of ATP production sites is maintained by K ions14. In the absence of fertilization, the similar behavior observed among cultivars regarding A reflects a common physiological response to K deficiency, which reduces photosynthetic activity9. At other K doses, the higher A values found in the Tanzânia cultivar may be attributed to the increased energy demand for shoot development in comparison to the use of organic reserves15. The association of higher A with lower root production in this cultivar supports this interpretation.
The linear behavior of gs in response to increasing K levels is related to the elevated photosynthetic rates (Table 1), since higher photosynthetic activity requires more K to regulate stomatal dynamics16. In contrast, the Zuri cultivar exhibited a peak gs value at the estimated K dose of 446.5 mg dm⁻3, indicating an optimal point of stomatal efficiency, followed by partial closure at higher doses—likely a mechanism to avoid excessive water loss. This may represent an adaptive strategy under high K availability, balancing CO2 assimilation with transpiration.
Despite the varying K levels, internal CO2 concentration (Ci) remained relatively stable, which can be explained by the relationship between gs and A17. A cultivar with high CO2 uptake (high gs) and high A may maintain Ci levels similar to a cultivar with lower values of both variables due to a physiological balance in gas exchange18.
Transpiration rate (E) also responded linearly to soil-applied K (Fig. 1), in line with the increase in gs. According to Pompeu et al. [18], this enhances water use efficiency, favoring K absorption and leading to improved forage production in the aerial part, as supported by the results of this study. Additionally, the absence of significant changes in leaf temperature (Tleaf) suggests that the cooling effect of transpiration was sufficient to offset the increased energy absorbed by the leaves. Under these conditions, the plant maintains a balanced leaf temperature13.
K also functions as an osmotic regulator, facilitating guard cell expansion and stomatal opening19. Higher K doses enhance stomatal conductance and, consequently, gas exchange and transpiration, supporting efficient regulation of Tleaf20. As for chlorophyll concentration, a quadratic response to K levels was observed (Fig. 1), with a peak of 29.84 SPAD units at a dose of 574.2 mg dm−3. This initial increase is due to K’s role in optimizing photosynthetic and physiological processes. However, at higher doses, nutrient imbalances may occur—particularly reduced uptake of magnesium, iron, and zinc, essential cofactors for chlorophyll synthesis19. This suggests that beyond ionic competition, K accumulation may disrupt metabolism and chloroplast function, resulting in decreased chlorophyll levels1.
Leaf transpiration rates were higher in the Quênia and Mombaça cultivars, likely due to their greater leaf length and width, while lower rates were recorded in Massai and Tamani (Table 2). This aligns with findings that larger leaf areas increase water demand through enhanced canopy photosynthesis21. In parallel, higher SPAD values in the Massai cultivar compared to others (Table 2) may be linked to a greater number of new leaves, driven by its higher tillering capacity22. These new leaves possess elevated photosynthetic potential and contribute to the more intense green coloration.
A significant increase of approximately 50% in both forage (Table 3) and root production (Table 4) was observed with the application of the minimum K dose (205 mg dm−3), underscoring the macronutrient’s critical role in plant physiology and growth. This initial rise likely results from improved photosynthetic efficiency and translocation of photoassimilates, fulfilling basic nutritional demands for biomass accumulation23. However, with increasing K doses, gains in forage production became less pronounced (Table 3), suggesting potential limitations due to insufficient availability of other macronutrients, particularly nitrogen (N). Ashworth et al.24 emphasize the need for balanced K and N application to maximize yield. This reinforces the idea that an adequate nutrient balance, not merely K supply, is essential for sustaining high productivity, rendering excess K ineffective in the absence of sufficient N.
Overall, both in the absence of fertilization and across the evaluated K doses, the Massai and Tamani cultivars consistently exhibited lower forage yields than Tanzânia, Quênia, Mombaça, and Zuri. This reflects intrinsic genetic differences, with Massai and Tamani presenting smaller plant sizes and a lower stem fraction in total forage mass25.
Conclusion
The physiological responses, forage, and root production of Panicum cultivars varied significantly according to the interaction between K doses and cultivar. Although increasing K doses generally enhanced photosynthesis, transpiration, and stomatal conductance, the magnitude and pattern of these responses differed among cultivars. For instance, while cultivars such as Tanzânia and Quênia showed linear increases across all variables, others like Zuri exhibited quadratic responses with optimal performance at intermediate K levels. Similarly, forage and root production did not increase uniformly among cultivars, with Quênia and Zuri standing out at intermediate doses and Mombaça at the highest dose. These results underscore the importance of considering cultivar-specific responses when determining optimal K fertilization strategies. A balanced nutrient supply remains essential, but K recommendations must be tailored to each cultivar’s physiological and productive behavior under varying soil K levels.
Materials and methods
Location, period, and experimental design
The experiment was conducted in a greenhouse at Embrapa Beef Cattle, Campo Grande–Mato Grosso of Sul (0°26′47,2" S; 54°43′23,6" O), between July 2019 and January 2020 (168 days).
The greenhouse without automated control of relative air humidity. Temperature regulation was achieved through an evaporative cooling system consisting of exhaust fans and a wet wall (PAD) located at one end of the structure. When the internal temperature reached 32 °C, sensors activated the system, causing water to flow through the PAD and the exhaust fans to operate, drawing in cooler, more humid air. This process was automatically halted when the temperature returned to 30 °C. Although the system is designed to maintain a temperature of approximately 25 °C, the average temperature during the 168-day experimental period was around 30 °C, except during specific periods when gas exchange measurements were performed. It is important to note that the system only controls high temperatures, with no mechanism for heating the environment.
The experimental design was a randomized block design in a 6 × 4 factorial scheme, with six cultivars (Panicum maximum cvs. Tanzânia, Quênia, Mombaça, Zuri, Massai, and Tamani) and four K doses: 0, 205, 410, and 820 mg dm−3, with three replications. The K doses were initially defined with the aim of evaluating K deficiency symptoms, based on the soil analysis and the nutrient requirements of the forage grasses. It is important to note that, in pot experiments, nutrient doses are typically higher than those applied under field conditions due to restricted root development and limited soil exploration. Under these confined conditions, a greater nutrient supply is required to adequately meet plant demands.
Although the experiment was conducted in pots inside a greenhouse, where environmental conditions are theoretically controlled, the presence of exhaust fans and an evaporative cooling system at one end of the structure could influence the performance of treatments located closer to these systems. Therefore, a randomized block design was adopted to minimize potential environmental variation within the greenhouse.
Preparation and conduct of the experiment
The sowing was performed in plastic pots with a volume of 2.55 dm−3, previously perforated, filled with a layer of gravel (2 cm) and soil classified as Typical Quartzarenic Neosol—Arenosols26, collected from the 0–20 cm depth. The soil presented the following chemical composition: pH (CaCl2) of 5.79; calcium (Ca2⁺) concentration of 1.70 cmolc/dm3; magnesium (Mg2⁺) of 1.07 cmolc/dm3; potassium (K+) of 0.15 cmolc/dm3; and no detectable aluminum (Al3+), with a value of 0.00 cmolc/dm3. The potential acidity (H+ + Al3⁺) was 1.15 cmolc/dm3. The sum of bases reached 2.92 cmolc/dm3. The potential cation exchange capacity (CEC) was 4.07 cmolc/dm3, while the effective CEC matched the sum of bases at 2.92 cmolc/dm3. Base saturation was 71.7%, and aluminum saturation was 0.00%. The soil also had an organic matter content of 1.49%, and available phosphorus (P) was measured at 44.6 mg/dm3.
Before sowing, a pre-planting fertilization was done with 54.68 mg dm−3 P; 1378 mg dm−3 of dolomitic limestone, aimed at increasing calcium and magnesium levels in the soil and correcting its acidity; 67.63 mg dm−3 S; 11.02 mg dm−3 Zn; 11.02 mg dm−3 Cu; 2.76 mg dm−3 B; and 1.37 mg dm−3 Mo. Nitrogen fertilization was applied 30 days after germination, with a dose of 100 mg dm−3 of nitrogen, totaling 255 mg per pot, using urea as the nitrogen source. Additionally, after each grass harvest, the same dose of 100 mg dm−3 of nitrogen was reapplied, in the form of urea and potassium chloride (KCl), according to the proposed treatments.
Soil moisture was maintained at approximately 80% of the pot capacity, a practical equivalent to field capacity for disturbed soils in pot experiments. Pot capacity was determined by the difference between the weight of the dry soil (sieved through a 2 mm mesh) and the weight of the saturated soil, obtained after slowly adding water until dripping began at the bottom of the container. Once dripping stopped, the final weighing allowed estimation of the amount of water retained, representing the maximum water-holding capacity of the system. Although frequent weighing was not performed throughout the 168-day experiment, irrigation was routinely adjusted to keep soil moisture close to this reference value.
Sowing was carried out on July 30, 2019, with 50 seeds per pot. Thinning began on the 15th day after sowing and was performed weekly until six plants remained per pot, which occurred around day 35. Fertilization started at this point, with K solution applied using a graduated pipette. The use of a nutrient solution promotes a more uniform distribution of nutrients within the pot, increases their availability, and reduces losses. The first application was performed 35 days after planting, and subsequent applications were carried out after each evaluation cut, at 28-day intervals.
The Panicum maximum cultivars (Tanzania, Kenya, Mombasa, Zuri, Massai, and Tamani) were selected and released by the breeding program of the Brazilian Agricultural Research Corporation (Embrapa Gado de Corte)27. Accordingly, Embrapa itself provided the genetic material (seeds) used to conduct this study.
Soil moisture was maintained at approximately 80% of the pot capacity, a practical equivalent to field capacity for disturbed soils in pot experiments. Pot capacity was determined by the difference between the weight of the dry soil (sieved through a 2 mm mesh) and the weight of the saturated soil, obtained after slowly adding water until dripping began at the bottom of the container. Once dripping stopped, the final weighing allowed estimation of the amount of water retained, representing the maximum water-holding capacity of the system. Although frequent weighing was not performed throughout the 168-day experiment, irrigation was routinely adjusted to keep soil moisture close to this reference value.
Physiological measurements
To evaluate the gas exchange in the cultivars, an infrared gas analyzer LI-6400 (Li-cor, Lincoln NE, USA) was used, with photosynthetically active radiation of 1000 micromoles (µmol m⁻2 s⁻1). The evaluations were performed weekly after each cut. Two representative tillers were selected for each cultivar, and the second youngest fully expanded leaf (with visible ligule) was identified. The measurement chamber was positioned at the central part of the leaf, always between 9:00 AM and 2:00 PM (Brasília time), as this period is considered to have the highest photosynthetic activity. The variables analyzed were leaf photosynthesis rate (A, µmol CO2 m⁻2 s⁻1), stomatal conductance (gs, mmol CO2 m⁻2 s⁻1), internal CO2 concentration (Cᵢ, ppm), leaf transpiration (E, µmol H2O m⁻2 s⁻1), and leaf temperature (Tleaf, in °C).
The chlorophyll concentration, expressed in SPAD units, was estimated using a SPAD-502 Chlorophyll Meter (Soil and Plant Analysis Development®). After each cut, a leaf from a tiller of each experimental unit was selected and marked with a permanent marker for weekly monitoring at the middle third of the leaf.
Forage biomass yield
The established cutting height for the taller plants (Tanzânia, Quênia, Mombaça, and Zuri) was 20 cm from the soil level, and for the shorter plants (Massai and Tamani), it was 15 cm. After the last sampling, a sixth cut was made close to the soil, and this material was dried in a forced-air oven, weighed, and the forage mass was summed to account for the forage production.
For the forage mass evaluation, the forage from each pot was cut at the pre-established residue height. All samples were placed in a labeled paper bag. The samples were then dried in a forced-air oven at 55 °C until a constant weight was obtained, after which they were weighed on an analytical balance (0.01 g) to determine the dry matter. Forage production was obtained per pot by summing the forage mass from all cuts.
Root biomass yield
After the final cut, irrigation was stopped to halt respiratory and enzymatic processes. The entire contents of the pots (roots and soil) were transferred to a wooden tray, and the roots were separated from the soil using running water with the aid of a set of aluminum sieves with 2.00 mm and 1.00 mm mesh sizes. After this procedure, the material was placed in paper bags, labeled, and taken to a forced-air oven at 55 °C for 72 h. After drying, the material was removed from the bags and weighed on an analytical balance (0.01 g) to estimate root production in grams (g).
Statistical analysis
Data were tested for homogeneity of variances with the Bartlett’s test, and normality of data-distribution (PROC UNIVARIATE), with the Shapiro–Wilk’s test, which are necessary prerequisites for analysis of variance. The data were subjected to analysis of variance using the SAS GLM procedure (v. 9.4, SAS Inc., Cary, NC, USA).
The statistical model adopted considered the fixed effects of cultivars, K doses, and their interaction. Cultivars, treated as a qualitative variable, were evaluated using the F-test, and when significant (P < 0.05), means were compared using Tukey’s test at a 5% significance level. K doses, treated as a quantitative variable, were analyzed using polynomial regression, testing both linear and quadratic models. The selected model was the one with significant regression coefficients (P < 0.05) and the highest coefficient of determination (R2).
Data availability
The data that support this study cannot be publicly shared due to ethical or privacy reasons of the Brazilian Agricultural Research Corporation (Embrapa) and may be shared upon reasonable request to the corresponding author if appropriate.
References
Lopes, M. N. et al. Gas exchange in Massai grass fertilized with nitrogen and grazed by sheep. Biosci. J. 36, 152–160 (2020).
Terrer, C. et al. Nitrogen and phosphorus constrain the CO2 fertilization of global plant biomass. Nat. Clim. Chang. 9, 684–689 (2019).
Euclides, V. P. B. et al. Maintaining post-grazing sward height of Panicum maximum (cv. Mombaça) at 50 cm led to higher animal performance compared with post-grazing height of 30 cm. Grass Forage Sci. 73, 174–182 (2018).
Difante, G. S. et al. Ingestive behaviour, herbage intake and grazing efficiency of beef cattle steers on Tanzania guineagrass subjected to rotational stocking managements. Rev. Bras. Zootec. 38, 1001–1008 (2009).
Gurgel, A. L. C. et al. Estrutura do pasto e desempenho de ovinos em capim-massai na época seca em resposta ao manejo do período das águas. Bol. Ind. Anim. 74, 86–95 (2017).
Santana, L. S. et al. Pasture structure and sheep performance supplemented on different tropical grasses in the dry season. Rev. Mex. Cienc. Pecuar. 11, 89–101 (2020).
Tesk, C. R. et al. Herbage responses of Tamani and Quênia guinea grasses to grazing intensity. Agron. J. 112, 2081–2091 (2020).
Mauri, A. C. et al. Biomass production and nutrient accumulation in some important tropical forage grasses for use in integrated crop-livestock systems. Aust. J. Crop Sci. 18, 894–899 (2024).
Ahmad, I. & Maathuis, F. J. M. Cellular and tissue distribution of potassium: physiological relevance, mechanisms and regulation. J. Plant Physiol. 12, 1–7 (2014).
Silveira, M. L. et al. Potassium and phosphorus fertilization impacts on bermudagrass and limpograss herbage accumulation, nutritive value, and persistence. Crop Sci. 57, 2881–2890 (2017).
Guo, J. et al. Growth, photosynthesis, and nutrient uptake in wheat are affected by differences in nitrogen levels and forms and potassium supply. Sci. Rep. 9, 1–12 (2018).
Nieves-Cordones, M., Alemán, F., Martínez, V. & Rubio, F. K⁺ uptake in plant roots: The systems involved, their regulation and parallels in other organisms. J. Plant Physiol. 171, 688–695 (2014).
Taiz, L., Zeiger, E., Moller, I. M. & Murphy, A. Fisiologia e desenvolvimento vegetal. (Artmed, Porto Alegre, 2017).
Prajapati, K. & Modi, H. A. The importance of potassium in plant growth: A review. Indian J. Plant Sci. 1, 177–186 (2012).
Chen, S., Wang, D., Lin, J. & Xu, G. Z. The comprehensive regulation of light intensity and photoperiod on growth and yield of virus-free potato under the same daily light integral. Agronomy 15(4), 898 (2025).
Tränkner, M., Tavakol, E. & Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 163, 414–431 (2018).
Sousa, R. O. et al. Photosynthetic efficiency and water status as determinants for the performance of semiarid-adapted cotton cultivars under drought in greenhouse. Agronomy 15(2), 500 (2025).
Pompeu, R. C. F. F. et al. Características morfofisiológicas do capim-Aruana sob diferentes doses de nitrogênio. Rev. Bras. Saúde Prod. Anim. 11, 1187–1210 (2010).
Marschner, P. Marschner’s mineral nutrition of higher plants 3rd edn. (Academic Press, 2012).
Xie, K. et al. Leaf photosynthesis is mediated by the coordination of nitrogen and potassium: the importance of anatomical-determined mesophyll conductance to CO2 and carboxylation capacity. Plant Sci. 290, 110–126 (2020).
Silva, P. S. S., Leite, R. D. C., Carneiro, J. S. S., Freitas, G. A. & Silva, R. R. Mombaça grass development with partial replacement of potassium fertilizer by sodium chloride and the effects of adding calcium. Trop. Grassl. Forrajes Trop. 8, 195–202 (2020).
Veras, E. L. L. et al. Tillering and structural characteristics of potassium cultivars in the Brazilian semiarid region. Sustainability 12, 1–11 (2020).
Hafsi, C., Debez, A. & Abdelly, C. Potassium deficiency in plants: Effects and signaling cascades. Acta Physiol. Plant. 22, 1–16 (2014).
Ashworth, A. J. et al. Switchgrass forage yield and compositional response to phosphorus and potassium. Agrosyst. Geosci. Environ. 2, 1–8 (2019).
Martuscello, J. A. et al. Produção e morfogênese de capim BRS Tamani sob diferentes doses de nitrogênio e intensidades de desfolhação. Bol. Ind. Anim. 76, 1–10 (2019).
Soil Survey Staff. Soil taxonomy: a basic system of soil classification for making and interpreting soil surveys. 2nd edn, (USDA-Natural Resources Conservation Service, Washington, DC, 1999). (USDA Agric. Handb. 436).
Fonseca, D. M. & Masrtuscello, J. A. Plantas Forrageiras, 2ª edição (ed. Fonseca, D. M., Masrtuscello, J. A.) 122–163 (editora UFV, 2022).
Acknowledgements
The authors thank the Embrapa Beef Cattle, Federal University of Mato Grosso of Sul Foundation, through the Postgraduate Program in Animal Science, the National Council for Sci-entific and Technological Development (CNPq), the Higher Education Personnel Improvement Coordination (CAPES, Finance Code 001) and the Foundation for the Support of the Development of Education, Science and Technology of the State of Mato Grosso of Sul (FUNDECT).
Author information
Authors and Affiliations
Contributions
E.L.LV., G.S.D., A.R.A., M.C.M.M., D.B.M., V.P.B.E., P.E.T. and A.L.C.G designed the study. E.L.L.V., J.G.R., J.C.S.S., G.O.A.M. and M.G.P performed the experiment and collected data. E.L.L.V., G.S.D., A.L.C.G., J.C.S.S. and P.E.T analyzed the data. A.L.C.G conducted statistical analysis. E.L.L.V. and J.G.R., wrote the manuscript. All authors read and critically revised drafts for intellectual content and provided approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
de Lima Véras, E.L., dos Santos Difante, G., de Araújo, A.R. et al. Gas exchange and productivity responses of Panicum maximum cultivars to increasing soil potassium levels in a greenhouse study. Sci Rep 15, 40704 (2025). https://doi.org/10.1038/s41598-025-24330-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-24330-y
