
Abstract
Tillage after the main crop is harvested may impact the growth of roots of the successive stubble cover crops (CCs). A three-year field experiment was conducted at the warm and dry Ruzyně and cold and wet Lukavec sites from 2021 to 2023. The root length density (RLD) to a depth of 60 cm was determined for seven variants of CCs, namely, two single species and five mixtures of two species: white mustard (Sinapis alba L.), phacelia (Phacelia tanacetifolia, Benth.), clover (Trifolium alexandrinum L.) and camelina (Camelina sativa (L.) Crantz), cannabis (Cannabis sativa L.) and setaria (Setaria italica (L.) Beauv.), buckwheat (Fagopyrum esculentum Moench) and phacelia, buckwheat and mustard, and oats (Avena sativa L.) and vetch (Vicia sativa L.). Two types of tillage, namely, mouldboard ploughing (PLO) and reduced shallow tillage (RET), were examined. Tillage did not significantly influence the RLD of the surface or subsoil layers or the average RLD of the 0–60 cm layer at either site, but the impact of tillage on the RLD interacted with CC variant. Reduced tillage decreased the RLD of the 0–60 cm layer by 0.25 cm cm-3 on average in mustard, phacelia, and the mixture of buckwheat and phacelia and increased it by 0.24 cm cm-3 in the other variants at Ruzyně. At Lukavec, RET decreased the RLD by 0.23 cm cm-3 in phacelia and mixtures of cannabis and setaria, buckwheat and phacelia, and mustard and oats, whereas the increase in the other variants was negligible (0.06 cm cm-3). The CC variants significantly affected the RLD of the 0–60 cm and 0–10 cm layers at both sites and in both tillage treatments, whereas this effect was not confirmed in the subsoil layer. The average RLD of 0–60 cm zone of the mixture of oats and vetch was greatest in both tillage treatments and at both sites (1.55–2.09 cm cm-3), followed by that of mustard (1.20–1.68 cm cm-3) and the mixture of buckwheat and mustard (1.05–1.51 cm cm-3). Reduced tillage increased the proportion of the root length in the top 30 cm layer by 3.81% and 3.58% on average at Ruzyně and Lukavec, respectively, and slightly reduced it in the subsoil layer at both sites. At both sites, the aboveground dry mass of CCs corresponded significantly with the RLD of the 0–60 cm layer and the subsoil layer (40–50 and 50–60 cm) under PLO. The observed differences in the root density and distribution of CCs, as well as their responses to tillage, could be important for the uptake of limited resources, water, and nutrients and should be considered when designing CC mixtures.
Similar content being viewed by others

Introduction
Cover crops (CCs) provide several benefits for soil quality and the environment, particularly by protecting the soil from erosion and by the depletion of residual (nitrate) nitrogen and its accumulation in biomass, which results in a decreased risk of N leaching during the nonvegetation period1. The input of organic matter, its contribution to soil carbon sequestration or disease suppression due to the fungicidal, nematocidal, and bactericidal activities of root exudates and decayed biomass and improvements in soil quality have been reported for various site conditions and cropping systems2,3,4,5,6. The introduction of CCs into cropping systems is part of the concept of climate-smart agriculture7,8.
The expected benefits depend on sufficient CC growth, which is often limited by unfavourable conditions for stubble CCs after the main crop is harvested. The establishment of stubble CCs is typically impaired by a shortage of soil water and available nutrients and by residues of the main crops, mainly straw from cereals, oilseed rape, and maize, in temperate cropping systems5,7,9.
Tillage systems with varying intensities and residue incorporation rates are employed after main crop harvesting and may impact CC root growth. Stubble CCs are mostly sown after reduced tillage, shallow chisel ploughing or harrowing, and, less often, nontilled soil in the Czech Republic (CR). Emergence is usually worse in reduced tillage (RET) than after ploughing10,11. Shallow tillage is often preferred by farmers because it uses less energy than ploughing and is expected to conserve water in the soil. The impacts of tillage after harvest on soil properties have often been studied, but the results depend on numerous conditions, and their relationship to the root growth of CCs is unclear12. Root growth under reduced tillage or no-till was reduced compared with that under a mouldboard-ploughed system, at least in some layers, which was attributed to increased penetration resistance13. Good emergence and root growth are essential prerequisites for available soil resource utilization during relatively short periods of late summer and autumn before the onset of winter14. The root traits of CC species and cultivars have been studied with respect to the depletion of residual nitrate from the top and subsoil layers15,16 and other ecosystem services17 or in relation to carbon input/deposition and rhizosphere microcosms6. An improvement in soil conditions, especially the amelioration effect on compacted soils, has been reported18,19. In this context, the morphology of the CC root system is important; for example, Hudek et al. reported that compared with thicker taproots, CCs with a fine root system are more beneficial for creating pore space20, and Blanco-Canqui concluded that mixing fibrous-rooted CCs with tap-rooted CCs may provide complementary benefits to reduce the risk of compaction21. Different root growth dynamics, profile distributions, and penetration depths of various species in CC mixtures could facilitate the more efficient use of the often-limited water and nutrient resources after the main crop is harvested22,23. In this study, we compared the growth of the most widely used CCs in the Czech Republic, mustard and phacelia, as single species and in a mixture with buckwheat, a fast-growing species. The effects of tillage on the shoot growth of CCs have been reported previously19; however, data on the effects of tillage on the roots of CCs are scarce24. Owing to methodological difficulties and laboriousness, even fewer data are available on CC roots in mixtures of two or more species under field conditions6.
A better understanding of the response of root growth of CC species to tillage would contribute to the formulation of selection guidelines for different soil and climatic conditions. A dense and deep root system enables plants to utilize water and nutrients from the soil effectively16,20,25.
This study aimed to determine the effects of conventional and reduced tillage practices after the main cereal crop is harvested on the root length distribution in topsoil and subsoil in CCs grown alone and as a mixture of species. We hypothesized that compared with reduced tillage, ploughing would significantly improve root growth, i.e., greater root length. However, the effects of tillage might differ among the studied species and mixtures.
Materials and methods
Experimental sites
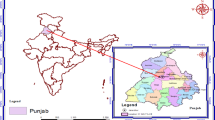
A three-year field experiment was conducted from 2021 to 2023 at the Czech Agrifood Research Center in Prague, Ruzyně (RUZ, https://www.carc.cz), and the Lukavec experimental station (LUK, https://www.spslukavec.cz) (Fig. 1). The map of the orography of the Czech Republic was prepared in ARCGis 10.6.1 software environment (ArcGIS Enterprise, Redlands, CA, USA) using ZABAGED® file datasets, which are provided as open data, free of charge under the Creative Commons CC BY 4.0 license (https://geoportal.cuzk.cz/(S(xha41rssilscp4polc0u1pxz))/Default.aspx?lng=EN&mode=TextMeta&text=dSady_zabaged&side=zabaged&menu=24). The experiment at RUZ was performed in the same field (50°5′5.053" N, 14°18′5.536" E). At LUK, the experiment was conducted in two adjacent fields from 2021 to 2022 (49°33′22.85" N 14°58′43.671" E) and in a nearby field in 2023 (49°34′0.309" N 14°59′0.637" E). The experimental fields were conventionally farmed with regular autumn ploughing to a depth of 18–22 cm before the start of this experiment. Cereals or oilseed rape were grown in previous years. The experimental sites have different soil and climate conditions; Ruzyně is 340 m a.s.l. in a warm region, with deep clay-loam soil developed on loess (degraded chernozem). The average (1991–2020) temperatures and total precipitation at this site are 9.6 °C and 497.5 mm, respectively; daily meteorological data at the site are available (www.carc.cz/meteostanice). Lukavec is located in a mildly warm region with Cambisol (brown soil), loamy-sandy soil, with an average (1991–2020) temperature and total precipitation of 7.5 °C and 694 mm, respectively. The basic site and soil characteristics are shown in Table 1, and the temperature and precipitation during the experimental years are shown in Fig. 2.

Position of the field experiments at Ruzyně and Lukavec.

Precipitation and temperature (at 2 m) from 2021–2023 and the average values for 1991–2020 at Ruzyně and Lukavec. Precipitation is the amount accumulated from July 1 to March 31.
Variants of soil tillage
CCs were sown after the combined harvest of winter wheat (2021) and spring barley (2022, 2023) at RUZ and after the harvest of winter barley (2021, 2023) and winter wheat (2022) at LUK. The height of the stubble ranged from 6–8 cm, and the straw was removed. The main cereal crops were harvested in July, followed by tillage within 14 days and sowing of CCs within several days (LUK) or another 10–14 days (RUZ). The experiment involved two variants of tillage after the main crop was harvested: conventional mouldboard ploughing to a depth of 20–24 cm and smoothing the soil surface with harrows (PLO) and reduced tillage with chisel ploughing to a depth of 8–10 cm (RET). The CCs were seeded using a PaL Ripper HD 302 seed drill (P & L, spol. s r. o., Czechia, pal@pal.cz) at RUZ and a Pöttinger Vitasem 302-disc seed drill (PÖTTINGER Landtechnik GmbH, Austria, info@poettinger.at) at LUK. The sowing rates are shown in Table 2.
Variants of cover crops
Two single crops (mustard and phacelia) and five two-species mixtures were studied (Table 2). The seven CC variants were white mustard (Mus), phacelia (Pha), clover and camelina (CloCam), cannabis and setaria (CanSet), buckwheat and phacelia (BucPha), buckwheat and mustard (BucMus), and oats and common vetch (OatVet). A split-plot experimental design was applied. The tillage system consisted of seven CC plots each replicated three times. The plot sizes were 36 and 60 m2 at the RUZ and LUK sites, respectively. No fertilizers or herbicides were applied after the main crops were harvested or during CC growth. The CCs were sown on August 19, 2021, August 11, 2022, and August 24, 2023, at RUZ, and on August 9, 2021, July 28, 2022, and August 2, 2023, at LUK. CC seeds were purchased from specialized companies (SEED SERVICE, s.r.o. Vysoké Mýto, CZ, https://seedservice.cz/osiva, and PRO SEEDS s.r.o., Havlíčkův Brod, CZ, https://www.proseeds.cz/osiva/). The seeds of all the CCs and cultivars are commercially produced, can be purchased and are commonly used by farmers in the Czech Republic.
Root study
The roots of the CC species cannot be easily distinguished from those of volunteer cereal plants after destructive soil sampling with a soil auger. Therefore, the following approach was adopted as a compromise. Ten to fourteen days before soil sampling, the shoots of volunteer plants and sporadic weed individuals (if any) in the vicinity (less than 10 cm) of the selected CC individuals were removed. At the time of sampling, the roots of dead plants were damaged and could be distinguished by their brown colour; thus, they were removed during the cleaning of the root samples.
Three replicates of the soil with roots were sampled at the time of aboveground biomass harvest before winter. Each replicate was formed by mixing two soil samples taken by a soil root corer 8 cm in diameter (Eijkelkamp, NL). The sampler was situated with a plant in the centre (a single species) or with the centre between two plants of each species adjacent to each other (less than two centimetres) to model their interactions. An effort was made to select plants representing the plant variability in the plots. Roots were collected away from areas with many volunteer cereal plants or sporadic perennial weeds. The soil samples were taken in 10 cm segments to 60 cm depth and soaked in 5% Na2CO3·10H2O to improve disaggregation, and the roots were subsequently washed on a set of sieves (2- and 0.5 mm mesh sizes). The soil and organic particles were removed by sedimentation and decantation. Finally, the remaining, primarily organic, residues were manually cleaned from the roots using tweezers. The length of the roots was measured manually26. The dry root weight was determined at 65 °C to a constant weight. The average root length density, RLD (cm cm−3 soil), root dry mass density, RMD (mg cm−3 soil), and specific root length, SRL (cm mg-1 of dry root), in a sample in the 10 cm soil segments were calculated. The effects of experimental factors on the RLD values in the 0–10 cm topsoil, 40–50 cm, and 50–60 cm subsoil layers indicated the differences in root distribution and depth important for the depletion of nitrogen and water, and the average RLD in the 0–60 cm layer was used to analyse the effects of year, tillage, and CC variant. The relative distribution of root length in a sample was calculated as the proportion of the RL in a given layer to the total root length in the sample at 0–60 cm.
Aboveground biomass
The plants (including cereal volunteer plants) from 0.50 m2, formed from two 0.25 m2 samples from two places, on every plot, were cut at the soil surface, and the dry weights (at 65 °C) of the CCs and cereal volunteers were determined. Here, only the aboveground dry mass (ADM) of CC plants is presented.
Statistical analysis
The effects of year, tillage, and CC variant on root length density were analysed separately for the two sites using categorical ANOVA. Tillage was used as the main plot, and CC variant was used as the split-plot treatment for data analysis. A fixed-effects model was used. Because some RLD data were not normally distributed according to the Shapiro‒Wilk test (p < 0.05), all the data were log-transformed for analysis. Differences between means were evaluated by Tukey’s HSD or Dunnett’s test if Levene’s test at p < 0.05 did not confirm homogeneity of variance. In cases where log-transformed data did not meet the requirement of normality, the nonparametric Kruskal‒Wallis test was employed (p < 0.05). The ADM data from Ruzyně did not have a normal distribution and were transformed using the cube root, followed by ANOVA. At Lukavec, the nonparametric Kruskal‒Wallis test was used (p < 0.05), as the transformed data were not normally distributed. Pearson’s correlation analysis was performed to analyse the relationships between the ADM and the RLD or RMD. The STATISTICA 14 program (StatSoft, Inc., Tulsa, OK, USA) was used.
Results
Effects of tillage on the root length density in soil
Tillage did not significantly (p < 0.05) influence the RLD at either site or at any of the analysed depths (Table 3). The average RLD at 0–60 cm was almost the same under PLO and RET, with values of 1.17 and 1.20 cm cm-3, respectively, at RUZ (Table 4). At LUK, the values were similar, namely, 1.20 and 1.10 cm cm-3 under PLO and RET, respectively.
The analysis revealed a significant, mostly weak interaction (p > 0.05) between tillage and the CC variant for the average RLD at 0–60 cm at both sites and for 0–10 cm at RUZ, whereas no interaction was detected between tillage and year (Table 3). A high RLD at 0–60 cm was observed in Mus, Pha, and BucPha, with values 0.25 cm cm-3 higher on average (120–136% of RET) under PLO, whereas the RLD values under RET of CloCam, CanSet, BucMus, and OatVic were 0.24 cm cm-3 greater (105–155% of PLO) at RUZ. At LUK, the interaction was relatively weak; a 0.23 cm cm-3 higher RLD on average was observed in Pha, CanSet, BucMus, and OatVic (106–135% of RET) under PLO, and a 0.06 cm cm-3 lower RLD was detected in Mus, CloCam, and BucPha (102–110% of PLO). However, most (16 out of 21 pairs) of the differences between PLO and RET for the same CC variant were not significant at p < 0.05 (Table 3).
Root length density of cover crop variants
The ANOVA results revealed a highly significant (p < 0.001) effect of the CC variants and year on the RLD at 0–60 cm and the top 0–10 cm at RUZ and LUK, whereas the effect was not significant at subsoil depths of 40–50 and 50–60 cm. Compared with the other variants, OatVet (1.79 cm cm-3), Mus (1.46 cm cm-3), and BucMus (1.39 cm cm-3) had significantly higher RLD (averaged over tillage variants and years) at 0–60 cm. At LUK, OatVet (1.82 cm cm-3) had the highest RLD, followed by Mus (1.14 cm cm-3) and BucMus (1.17 cm cm-3). The lowest average RLD at 0–60 cm was found in CanSet, Pha, and BucPha (RUZ) or CloCam (LUK) (0.74–1.06 cm cm-3) (Table 4).
The average RLD was significantly lower in 2021 (1.06 cm cm-3) than in 2022 (1.22 cm cm-3) and 2023 (1.27 cm cm-3) at RUZ. The order was similar at LUK, with the RLD significantly higher in 2023 (1.31 cm cm-3) than that in 2021 (1.03 cm cm-3) and 2022 (1.11 cm cm-3).
Effects of tillage on the root length distribution in the soil and specific root length
The roots of all the CCs were concentrated in the topsoil (Fig. 3). On average, 46.7% (PLO) and 49.3% (RET) of the RLs in a sample were found in the 0–10 cm zone at RUZ. At LUK, the decrease in roots with depth was less steep, and the proportions of RL in the topsoil were low: 38.4% (PLO) and 42.5% (RET) (Fig. 4). Reduced tillage increased the proportion of root length in the top 30 cm by 3.81% and 3.58% on average at Ruzyně and Lukavec, respectively, and slightly reduced it in the subsoil at both sites. The average proportions of RL in the deepest monitored zone, at 50–60 cm depth, reached 2.83% (PLO) and 2.22% (RET) at RUZ and 1.20% (PLO) and 1.10% (RET) at LUK. Despite the small proportion, the RLD in the subsoil layers (40–50 cm and 50–60 cm) was significantly and positively correlated with the average RLD throughout the inspected soil profile, with r values ranging from 0.90 to 0.95 at RUZ. At LUK, the correlation was weaker, with r values ranging from 0.56 to 0.87.

Distribution of root length density in the soil profile at Ruzyně and Lukavec. PLO and RET denote the ploughing and reduced tillage treatments, respectively. Variants: mustard (Mus), phacelia (Pha), clover and camelina (CloCam), cannabis and setaria (CanSet), buckwheat and phacelia (BucPha), buckwheat and mustard (BucMus), and oat and vetch (OatVet).

Relative distributions of root length (RL) in the soil profile (in % of total RL at 0–60 cm in a sample) at Ruzyně and Lukavec. PLO and RET denote the ploughing and reduced tillage treatments, respectively. Variants: mustard (Mus), phacelia (Pha), clover and camelina (CloCam), cannabis and setaria (CanSet), buckwheat and phacelia (BucPha), buckwheat and mustard (BucMus), and oat and vetch (OatVet).
The SRL was greater at RUZ than at LUK, and the value decreased approximately linearly with depth. The values averaged over the variants increased from 0.54 cm mg-1 at 0–10 cm to 7.28 cm mg-1 at 50–60 cm under PLO and from 0.55 to 13.02 cm mg-1 under RET at RUZ. The corresponding values at LUK increased from 0.52 cm mg-1 at 0–10 cm to 4.22 and 3.28 cm mg-1 at 40–50 cm and at 50–60 cm, respectively (PLO), and from 0.49 to 4.17 (40–50 cm) and 3.72 cm mg-1 (50–60 cm) under RET (Fig. 5).

Distribution of specific root lengths of cover crops in the soil profile. PLO and RET denote the ploughing and reduced tillage treatments, respectively. Variants: mustard (Mus), phacelia (Pha), clover and camelina (CloCam), cannabis and setaria (CanSet), buckwheat and phacelia (BucPha), buckwheat and mustard (BucMus), and oat and vetch (OatVet). The average values of the SRL in the layers are shown (Average).
Aboveground dry matter yield
The effects of tillage and CC variants on ADM yield were similar at both sites. The average ADM was significantly greater under PLO (170.0 and 81.8 g m2) than under RET (84.4 and 52.4 g m2) at RUZ and LUK, respectively. Mus and OatVic produced the highest ADM at both sites, followed by Pha. The lowest ADM was observed for CanSet at both sites (Table 5). The correlation coefficients between the average ADM of the CCs at LUK and RUZ were 0.82 (PLO) and 0.84 (RET) (n = 7, p < 0.05).
The relationships between ADM (averaged over time) and RLD at 0–60 cm were significant at RUZ and LUK for all combinations except for RUZ/RET, whereas the relationships between ADM and RMD were not significant (r from 0.20 to 0.60) (Fig. 6). At both sites, the ADM significantly corresponded to the RLD in the top 0–10 cm and subsoil layers (40–50 and 50–60 cm, respectively) under PLO (r > 0.76; n = 7; p < 0.05) and nonsignificantly under RET (r from 0.51 to 0.73).

The correlations between average aboveground dry mass yield and average root length or root dry mass at 0–60 cm at Ruzyně (Ruz) and Lukavec (Luk). Tillage treatments included ploughing (PLO) and reduced shallow tillage (RET). * denotes significance at p < 0.05 (n = 7).
Discussion
The aim of this study was to verify the hypothesis of improved root growth in selected CCs under conventional tillage compared with reduced tillage. The experimental data did not confirm the hypothesis that tillage (PLO and RET) imposes a systematic, significant effect on the average root length density in the 60 cm zone of the examined soil or in the individual top and subsoil soil layers. A possible reason for this finding could be a significant interaction between tillage and the CC variants. Furthermore, the distribution of roots showed increased growth in the topsoil under RET and slightly less growth in the subsoil at both sites. As documented in numerous studies, reduced tillage influences soil physical conditions and may affect crop shoot and root growth. However, data on stubble CCs are sporadic16. Research has often focused on using CCs to improve the soil conditions of subsequent crops19,24,27,28. In our experiment, conventional tillage and deep ploughing were regularly used in cereals preceding the stubble CCs.
Unlike tillage, the CC variants significantly affected the average RLD of the 0–60 cm and 0–10 cm layers but not of the subsoil layers at both sites. Similar results for the average and topsoil analyses are logical because 0–10 cm represented 38.4–49.3% of the total length of the examined zone. Substantial differences in the shoot growth of CCs have often been documented12,24,29. Comparisons of the RLD of CCs with those in other studies are not easy because of differences in soil and weather conditions, precrop and tillage, sowing and harvest terms, or length of growth. The highest RLD values in this experiment were observed in OatVic, Mus, and BucMus, whereas relatively low values were observed in CanSet, Pha and CloCam. The similar order of variants in different soil and climatic conditions confirms the importance of the characteristics of the given species. The high root density of mustard is consistent with the vigorous shoot and root growth of Brassicaceae species (rape, kale, oilseed radish), which corresponds to a high demand for and depletion of nitrogen29. Relatively cold weather in autumn is suitable for cereals used as CCs (mostly rye), which form dense root systems in the topsoil, as reflected by the distribution of roots in the soil. The reduced shoot and root growth of cannabis and setaria was probably caused by their sensitivity to low autumn temperatures at the soil surface30. Despite the different conditions, the root length data are comparable to those published; for example, Hudek et al. reported 90-day container experiment values of 2.06, 4.9, 4.45, and 4.09 cm cm-3 for phacelia, mustard, oats, and buckwheat, respectively, at 0–15 cm and 0.12, 0.39, 0.26, and 0.29 cm cm-3 at 45–60 cm20.
Although different methods for collecting roots and aboveground parts were applied, a positive relationship was found between the average RLD values at 0–60 cm and the ADM. Furthermore, the CCs with high aboveground mass presented high root density in the subsoil layers (40–50 and 50–60 cm), which may be important for the depletion of leached nitrate. The average RLD at 0–60 cm and in the topsoil at Ruzyně also showed a significant interaction between the CC variant and the experimental year, which complicates the generalization of the results14. On the other hand, the effects of interactions between tillage and the experimental year on the RLD were not significant. A comparison of precipitation and temperature at Ruzyně does not explain the low RLD in 2021 and the high RLD in 2022 and 2023. Additionally, the RLD did not respond to high precipitation in 2022.
The significant interaction between the tillage treatment and CC variant suggests that the changes in soil conditions affected CC species differently. These differences may be related to the different root morphologies of CC species. For example, white mustard (Sinapis alba L.), like many other Brassicaceae species, has a strong taproot, the roots of phacelia are concentrated in the upper soil31, and hairy vetch has a primary root that branches into several lateral roots of similar diameter32. PLO increased the RLD in Mus, Pha, and BucPha at RUZ. These results do not reveal a straightforward relationship between species root traits and different types of tillage. Oats and rye, which are often used as CCs, have fibrous root systems that may explain why the root density in the subsoil was lower in compacted soil than in loose soil. This effect was significantly less pronounced in rapeseed and forage radish18. Our data did not confirm this assumption, and the observations do not seem generally valid. For example, Wong et al. reported that compared with rapeseed and kale, winter triticale formed coarser roots and exhibited deeper rooting17. In the lighter soil at LUK, the impact of tillage on the RLD was relatively low.
The effect of tillage on SRL was not observed at LUK, probably because of the different soil textures; clay loam at Ruzyně generally provides less suitable conditions for the penetration of thick roots. Notably, low root density, i.e., small root samples, in the subsoil may increase the degree of sampling error because soil particles may adhere to the root tissues33. A decrease in SRL with depth in CCs and large differences between CC species, as observed in our experiment, were reported by Hudek et al.20. Additionally, similar to the authors, a high SRL was observed in mustard and oats.
The depth of CC roots is important for depleting residual nitrate and for the uptake of water15,16. Overwintering CCs reach the 60–90 cm layer, and approximately 90% of the root mass and length are concentrated in the 0–15 cm topsoil zone17. In our study, the CC plants reached depths of more than 50 cm in a relatively short time before the onset of winter at both sites. The data on the RLD in the deepest monitored soil layer may indicate the potential for N depletion from the subsoil, which is especially important under the soil-climate conditions at Lukavec, a relatively wet site with light soil and high contents of coarse sand and stones in the subsoil, making it prone to nitrate leaching34. Even a low root density enables plants to take up mobile nitrate35,36. The limit of 1.0 or 0.5 cm cm-3 is sometimes given as a sufficient root density for nitrogen nitrate uptake. In our experiment, the average RLD decreased below 0.5 cm cm-3 between depths of 15 and 35 cm under RET and between 25 and 45 cm under PLO at RUZ, suggesting differences in depletion potential. High root density at 50–60 cm was observed in OatVic, BucMus, and Mus at both sites, but the differences were not significant.
This methodical approach did not enable us to distinguish the roots of individual species in mixtures. CCs are now grown almost exclusively as mixtures of two or more species; therefore, their different properties can be advantageous. Compared with single-species stands, growing mixtures of CCs provides several benefits, for example, increased rhizodeposition and improved nitrate depletion because of differences in root growth rates, extent, and depth21,22,23. Data from the literature concerning CC mixtures have not confirmed greater root length in CC mixtures than in monocultures37,38. However, in container experiments, increased root lengths, densities, and areas were observed in mixtures, for example, in mixtures of vetch and rye compared with rye alone39,40. In our experiment, compared with the mixture including phacelia, the mixture of buckwheat with mustard had a greater RLD, which agrees with the lower RLD in single phacelia than in single mustard. There was some tendency towards a higher RLD for the mixtures than for the single species under RET.
The sampling of roots poses a challenge, as the roots of crop and volunteer or weed species can be distinguished after destructive soil sampling with a soil auger only via sophisticated, advanced methods41. Simplified approaches are often used; for example, Wong et al. advise “[taking] care to avoid dense patches of weed within plots”17, and Amsili and Kaye justified nonrandom biomass sample locations (avoiding nontarget species) by focusing measurements on the variation caused by the species of interest rather than other types of field-scale variations, such as weeds or areas with uneven stands6. Bodner et al. placed an auger with the plant at the center42. Container experiments partially avoid these difficulties20,39,40. Our approach enabled us to preserve the competition effect of neighbouring plants for most of the growth period. Increased root growth could not be excluded, but CC plants occupied the topsoil after two months. We used the root length near the plants to compare the CC variants. The roots can expand to a great distance from the plant6, but according to visual inspection of the excavated plants, the roots were concentrated near the base of the plants. It could be assumed that plants with more extensive root systems also have greater lateral extents and vice versa. As a result, the observed differences between CC species would be even more distinct.
Notably, our data cannot be simply extrapolated to the entire area of a plot because of the sampling approach. In addition, reduced tillage significantly increased the spatial variability in CCs and cereal volunteer plants because of the uneven accumulation of postharvest residues of previous cereal crops and the different drying conditions of the loosened soil surface. The primary objective of our study was to compare the effects of different tillage systems on the roots of CCs, and this objective has been achieved, even if the data are interpreted as indicators of root size. The exact determination of root size would require sampling at different distances from a single plant while considering the different numbers and uneven distances of other individuals or employing the monolith method. To obtain more precise data on the influence of various factors on root growth and distribution in CC species mixtures, new methods and approaches are necessary41.
In conclusion, the expected negative impact of reduced tillage on root length and weight was not confirmed. The results revealed large differences between CC species or their mixtures and, at the same time, indicated different responses of CC species to tillage. These differences could be important for the uptake of limited resources, water and nutrients, and taking these differences into account is recommended when designing CC mixtures.
Data availability
The datasets used and analysed during the current study will be made available by the authors upon reasonable request to the corresponding author (jan.haberle@carc.cz).
Abbreviations
- RLD:
-
Root length density (cm cm-3 of soil)
- RMD:
-
Root dry mass density (mg cm-3 of soil)
- SRL:
-
Specific root length (cm mg-1 of dry root)
- ADM:
-
Aboveground dry mass yield (kg m-2)
- PLO:
-
Conventional mouldboard ploughing tillage
- RET:
-
Reduced tillage with chisel ploughing
- RUZ, and LUK:
-
Ruzyně and Lukavec experimental sites, respectively
- Mus:
-
White mustard
- Pha:
-
Phacelia
- CloCam:
-
Clover and camelina
- CanSet:
-
Cannabis and setaria
- BucPha:
-
Buckwheat and phacelia
- BucMus:
-
Buckwheat and mustard
- OatVet:
-
Oats and common vetch
References
Thapa, R., Mirsky, S. B. & Tully, K. L. Cover crops reduce nitrate leaching in agroecosystems: A global meta-analysis. J. Environ. Qual. 47, 1400–1411 (2018).
Ghidoli, M. et al. Camelina sativa (L.) crantz as a promising cover crop species with allelopathic potential. Agronomy 13, 2187 (2023).
Engedal, T. et al. Plant organ rather than cover crop species determines residue incorporation into SOC pools. Soil Biol. Biochem. 200, 109616 (2025).
Adetunji, A. T., Ncube, B., Mulidzi, R. & Lewu, F. B. Management impact and benefit of cover crops on soil quality: A review. Soil Tillage Res. 204, 104717 (2020).
Koudahe, K., Allen, S. C. & Djaman, K. Critical review of the impact of cover crops on soil properties. Int. Soil Water Conserv. Res. 10, 343–354 (2022).
Amsili, J. P. & Kaye, J. P. Root traits of cover crops and carbon inputs in an organic grain rotation. Renew. Agric. Food Syst. 36, 1–10 (2020).
Quintarelli, V. et al. Cover crops for sustainable cropping systems: A review. Agriculture 12, 2076 (2022).
Erekalo, K. T. et al. Review on the contribution of farming practices and technologies towards climate-smart agricultural outcomes in a European context. Smart Agric. Technol. 7, 100413 (2024).
Scavo, A. et al. The role of cover crops in improving soil fertility and plant nutritional status in temperate climates. A review. Agron. Sustain. Dev. 42, 93 (2022).
Waring, E. R., Pederson, C., Lagzdins, A., Clifford, C. & Helmers, M. J. Water and soil quality respond to no-tillage and cover crops differently through 10 years of implementation. Agric. Ecosyst. Environ. 360, 108791 (2024).
Haramoto, E. R. Species, seeding rate, and planting method influence cover crop services prior to soybean. Agron. J. 111, 1–11 (2019).
Harasim, E., Antonkiewicz, J. & Kwiatkowski, C. A. The effects of catch crops and tillage systems on selected physical properties and enzymatic activity of loess soil in a spring wheat monoculture. Agronomy 10, 1–12 (2020).
Kadžienž, G., Munkholm, L. J. & Mutegi, J. K. Root growth conditions in the topsoil as affected by tillage intensity. Geoderma 166, 66–73 (2011).
Herrera, J. M., Feil, B., Stamp, P. & Liedgens, M. Root growth and nitrate-nitrogen leaching of catch crops following spring wheat. J. Environ. Qual. 39, 845–854 (2010).
Kristensen, H. L. & Thorup-Kristensen, K. Root growth and nitrate uptake of three different catch crops in deep soil layers. Soil Sci. Soc. Am. J. 68, 529–537 (2004).
Thorup-Kristensen, K. Are differences in root growth of nitrogen catch crops important for their ability to reduce soil nitrate-N content, and how can this be measured?. Plant Soil 230, 185–195 (2001).
Wong, K. M. et al. Cover crop cultivars and species differ in root traits potentially impacting their selection for ecosystem services. Plant Soil 500, 279–296 (2024).
Chen, G. & Weil, R. R. Penetration of cover crop roots through compacted soils. Plant Soil 331, 31–43 (2010).
Gajewski, P., Majchrzak, L., Bocianowski, J. & Kaczmarek, Z. Effects of cover crops and tillage methods on selected physical and water retention properties of Luvisol. Int. Agrophys. 39, 73–86 (2025).
Hudek, C., Putinica, C., Otten, W. & De Baets, S. Functional root trait-based classification of cover crops to improve soil physical properties. Eur. J. Soil Sci. 73, e13147 (2022).
Blanco-Canqui, H. Do cover crop mixtures improve soil physical health more than monocultures?. Plant Soil 495, 99–112 (2024).
Engedal, T. et al. Legume-based cover crop mixtures can overcome trade-offs between C inputs, soil mineral N depletion and residual yield effects. Agric. Ecosyst. Environ. 349, 108408 (2023).
Restovich, S. B., Andriulo, A. E. & Portela, S. I. Cover crop mixtures increase ecosystem multifunctionality in summer crop rotations with low N fertilization. Agron. Sustain. Dev. 42, 19 (2022).
Büchi, L., Wendling, M., Amossé, C., Necpalova, M. & Charles, R. Importance of cover crops in alleviating negative effects of reduced soil tillage and promoting soil fertility in a winter wheat cropping system. Agric. Ecosyst. Environ. 256, 92–104 (2018).
Luo, Z. et al. Soil-smart cropping for climate-smart production. Geoderma 451, 117061 (2024).
Tennant, D. A test of a modified line intersect method of estimating root length. J. Ecol. 63, 995–1001 (1975).
Williams, S. M. & Weil, R. R. Crop cover root channels may alleviate soil compaction effects on soybean crop. Soil Sci. Soc. Am. 68, 1403–1409 (2004).
Grunwald, D. & Koch, H. J. Cover crop effects on fibrous roots and growth of subsequent sugar beet. Plant Soil 495, 487–498 (2024).
Cottney, P., Black, L., White, E. & Williams, P. N. Organic manures integrated with cover crops affect both cover crop and commercial crop performance including soil health: Effect on cover crop growth, nitrogen uptake, and soil mineral N. Agriculture 15, 1–18 (2025).
Haberle, J., Chuchma, F., Raimanova, I. & Wollnerova, J. Agroclimatic zoning of temperature limitations for growth of stubble cover crops. Climate 13, 1–16 (2025).
Hampl, U. Gründüngung (Leopold Stocker Verlag, 1996).
Kutschera, L., Lichtenegger, E. & Sobotik, M. Wurzelatlas Der Kulturpflanzen Gemäßigter Gebiete: Mit Arten Des Feldgemüsebaues (DLG-Verlag GmbH, 2009).
Janzen, H. H., Entz, T. & Ellert, B. H. Correcting mathematically for soil adhering to root samples. Soil Biol. Biochem. 34, 1965–1968 (2002).
Kautz, T. et al. Nutrient acquisition from arable subsoils in temperate climates: A review. Soil Biol. Biochem. 57, 1003–1022 (2013).
Kuhlmann, H., Barraclough, P. B. & Weir, A. H. Utilization of mineral nitrogen in the subsoil by winter wheat. Z. Pflanzenernähr. Bodenkd. 152, 291–295 (1989).
Haberle, J. & Káš, M. Simulation of nitrogen leaching and nitrate concentration in a long-term field experiment. J. Cent. Eur. Agric. 13, 416–425 (2012).
Bukovsky-Reyes, S., Isaac, M. E. & Blesh, J. Effects of intercropping and soil properties on root functional traits of cover crops. Agric. Ecosyst. Environ. 285, 106614 (2019).
Yu, Y. et al. Estimation of runoff mitigation by morphologically different cover crop root systems. J. Hydrol. 538, 667–676 (2016).
Saleem, M. et al. Cover crop diversity improves multiple soil properties via altering root architectural traits. Rhizosphere 16, 100248 (2020).
Guardia, G. et al. Balancing legume-cereal proportions in cover crop mixtures to minimize N2O emissions. Geoderma 454, 117195 (2025).
Rewald, B., Meinen, C., Trockenbrodt, M., Ephrath, J. E. & Rachmilevitch, S. Root taxa identification in plant mixtures—current techniques and future challenges. Plant Soil 359, 165–182 (2012).
Bodner, G., Himmelbauer, M., Loiskandl, W. & Kaul, H. P. Improved evaluation of cover crop species by growth and root factors. Agron. Sustain. Dev. 30, 455–464 (2010).
Funding
This research was funded by the MINISTRY of AGRICULTURE of the CZECH REPUBLIC, grant numbers QK21010308 and MZe-RO0425.
Author information
Authors and Affiliations
Contributions
Conceptualization, J.H., M.K. and P.S.; field experiments M.K., methodology of root study, P.S. and J.H.; formal analysis, I.R., M.M. and M.L.; sampling and root investigation, P.S. and M.M.; resources, M.K.; writing—original draft preparation, J.H.; writing—review and editing, J.H., M.L., and I.R.; visualization, M.L.; project administration and funding, I.R., M.K. All authors have read, reviewed and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
The experiment complied with relevant institutional, national, and international guidelines and legislation.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Svoboda, P., Raimanova, I., Moulik, M. et al. Effects of tillage on the root length and depth distribution of stubble cover crops. Sci Rep 15, 40788 (2025). https://doi.org/10.1038/s41598-025-24494-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-24494-7
