
Abstract
Exogenous hormones are crucial for regulating reproductive output at various developmental stages in plants, including the medicinal species Pseudostellaria heterophylla, which has a chasmogamous (CH)–cleistogamous (CL) mixed mating system. This study examined the influence of phytohormones on reproductive output at different growth stages and aimed to identify strategies for optimizing genetic breeding. Phytohormones such as gibberellic acid, abscisic acid, indole-3-acetic acid, and zeatin were applied at concentrations of 0, 10, 50, 100, and 200 mg/L during the sprouting and pre-flowering stages of CH and CL flowers. Results indicated that 10 mg/L ABA during the pre-flowering stage increased CH flower numbers by about 30%, while 200 mg/L ABA reduced CH flower production by over 40%. Spraying 200 mg/L GA3 at the sprouting stage resulted in a 150% increase in CL flowers. Other hormones had negligible effects on CL flowers, with only a 9.4% reduction when 100 mg/L ZT was applied. Additionally, 200 mg/L GA3 during the pre-flowering stage significantly increased root weight by 55.6%. In conclusion, applying exogenous hormones at specific stages can effectively regulate flower numbers and root weight, aiding genetic breeding efforts.
Similar content being viewed by others


Introduction
Plants rely on the regulation of specific hormone levels to initiate each developmental stage, as these hormones play a crucial role in triggering specific gene expression patterns that govern various physiological processes1,2,3,4,5,6. Plant hormones regulate flowering by modulating epigenetic inheritance, acting as signals coordinating flowering in higher plants2,7. Exogenous hormones, in addition to endogenous hormones, play a significant role in plant tissue and organ differentiation, as well as in processes such as flowering, fruiting, maturation, senescence, dormancy, and germination2,8. The influence of exogenous hormones on plant development is well documented8,9. However, research on the effect of exogenous hormones on flowering and fruiting has been carried out on hermaphroditic or dioecious plants, while research focusing on the dimorphic mixed-mating species remains limited10.
The dimorphic mixed-mating plant, also known as the chasmogamous (CH)- cleistogamous (CL) system, refers to plants that can produce both CH and CL flowers sequentially on the same plant11,12. These two types of flowers serve distinct functions with a mixed mating system to achieve sexual reproduction. CH flowers attract pollinators with their bright corollas and nectar, facilitating cross-pollination and the production of outcrossed seeds. However, they can also produce self-pollinated offspring through self-pollination or geitonogamy13,14. In contrast, CL flowers lack corollas and nectar glands, have fewer stamens than CH flowers, and contain many ovules. This leads to automatic self-pollination and seed production14. Consequently, these plants produce seeds with two distinct genetic traits.
In natural environments, the relative proportions of CH and CL flowers, as well as the balance between self-pollinated and cross-pollinated seeds, are influenced by a variety of environmental and biological factors11,15,16. For example, when CH flowers cannot pollinate or produce fruits due to unfavorable conditions or insufficient pollinators, plants may redirect resources towards producing more CL flowers. This shift increases the production of self-pollinated seeds. Consequently, the reproductive strategy of plants in the CH-CL system is highly adaptable, allowing them to regulate reproductive output based on environmental conditions12.
P. heterophylla(Miq.) Pax ex Pax et Hoffm. (Caryophyllaceae) is a dwarf (10–20 cm tall) perennial herb with a dimorphic mixed-mating system native to eastern China17. In the study area, the flowering period of CH flowers ranges from mid-April to late May at the top of the plant. CH flowers are large, insect-pollinated, with five petals and sepals each, white petals, ten stamens, and a three-lobed stigma. CL flowers appear after the CH flowers and bloom until September. They are small, closed, lack petals, have four sepals (pale purple) and two stamens and occur in the lower part of the plant. The underground fleshy roots (Radix) overwinter and propagate vegetatively, with one root typically forming one plant. The underground fleshy roots of the species are widely used in traditional medicine and serve as the primary raw material for medicinal and health products. However, continuous cultivation has led to the degeneration of variety, with diseases becoming increasingly severe, resulting in reduced yields18. Consequently, there is a pressing demand for breeding disease-resistant, high-yielding new varieties to improve fleshy root yield18.
Our study investigates the role of exogenous hormones (gibberellic acid (GA3), abscisic acid (ABA), indole-3-acetic acid (IAA), and zeatin (ZT)) in regulating reproductive output at various developmental stages in P. heterophylla, which exhibits a CH-CL mixed mating system, although, the previous studies have shown that the number and weight of P. heterophylla roots were significantly increased after IAA treatment19, however, there are still deficiencies in research in related fields. The aim is to identify optimal hormone concentrations that promote CH or CL flower production and enhance root weight. The findings will contribute to breeding strategies focused on improving yield and disease resistance in this species.
Materials and methods
Plant materials
The P. heterophylla used in this study is a common plant in Liaoning Province, China, often growing on sunny hillsides. Before the experiment, wild P. heterophylla were transplanted into the homogenous garden experimental field. The wild habitat and the cultivated garden were several kilometers apart, and the environmental conditions were the same. (P. heterophylla can be harvested from the wild, and there are no ethical or legal issues.). Specific permission was not required for the study at our site. Endangered or protected species do not grow in the study site.
Following the CH flowering period, adventitious roots form in the transition zone of the rootstock, gradually swelling and maturing by the end of the CL flowering period in autumn. These roots develop into 3–6 unevenly sized fleshy structures capable of becoming independent individuals in the following year. This species’ reproductive strategy involves outcrossed seeds from CH flowers, self-pollinated seeds from CL flowers, and vegetative propagation through fleshy roots, contributing to its complex fine-scale spatial genetic structure (Fig. 1).

Dimorphic flowers and fleshy roots of Pseudostellaria heterophylla. (A) chasmogamous flower, (B) cleistogamous flower, and (C) fleshy roots.
Experimental block design
The experiment was conducted in a homogeneous garden on the Liaodong University campus in Dandong City, Liaoning Province, Northeast China (latitude 40°08, longitude 123°03′), covering approximately 400 m2. In early November 2022, fleshy roots were planted in nutrient bowls with a diameter and a height of 10 cm, each holding 500 g of native soil. To ensure uniformity, excessively large or small fleshy roots were removed. One plant was planted per nutrient bowl. The plants were divided into groups containing 30 plants, resulting in 49 groups and 1470 nutrient bowls planted. The samples were managed under normal overwintering conditions (In winter, fleshy roots can overwinter naturally.) (Fig. 2).
The experiment used a randomized block design, divided into three groups based on treatment times: A, B, and C. Group A represented the sprouting stage P. heterophylla shoots before flower buds appeared (March 25), with hormone spraying once daily for three consecutive days. Group B included plants in the pre-flowering stage (April 17) with most buds visible, receiving hormone spraying for three consecutive days. Group C involved continuous hormone spraying during both two stages.

Experimental block design in the homogeneous garden.
Exogenous hormone treatments
Four hormones, GA3, ABA, IAA, and ZT, were tested in the experiment at five concentration levels (0, 10, 50, 100, and 200 mg/L), with 0 mg/L serving as the control. Whole-plant spraying was performed between 9:00 AM and 11:00 AM for each experimental group, continuing until stems and leaves were saturated, the amount of each spray is 10 ml. Each concentration level was applied to 30 plants as replicates.
Data collection and analysis
Observation began three days after the final hormone spraying, with weekly recordings of the number of CH and CL flowers throughout the growth period. The number of new flowers was cumulatively added to the original record. Harvesting of the fleshy roots began in late September after the aboveground parts started to wither. Root weight and total weight were measured. Reproductive parameters were compared under different treatments using the MANOVA test with a significance set at 0.05. All statistical analyses were done using SPSS (22.0) software.
Results
The effect of exogenous hormones on the flowering frequency of CH plants
Effect of GA3 on CH flowers
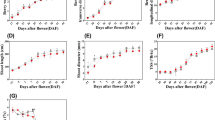
In the sprouting stage (Group A), the application of various concentrations of GA3 significantly (F = 77.89, P < 0.001) promoted CH flower numbers, with the high concentrations (200 mg/L) resulting in 21.6% per plant compared to the control group (Fig. 3). During the pre-flowering stage (Group B), a low concentration of 10 mg/L GA3 is more effective, leading to significant (F = 109.19, P < 0.001) 25.3% increase in CH flower numbers. In comparison, high concentration (200 mg/L) of GA3 suppresses CH flowering (Fig. 3). Continuous application of GA3 across both stages (Group C) does not show significant effects, with high concentrations exhibiting inhibitory effects (Fig. 3).
Effect of ABA on CH flowers
The application of ABA at a low concentration (10 mg/L) promotes CH flowering when applied during the sprouting or pre-flowering stages (Fig. 3). Particularly, during the pre-flowering stage, 10 mg/L ABA resulted in a significant (F = 97.32, P < 0.001) 31.7% increase in CH flower numbers compared to the control group (Fig. 3). However, the high-concentration of ABA (200 mg/L) inhibits CH flowering at all stage, with a remarkable (F = 136.77, P < 0.001) 41.9% reduction in CH flower numbers than the control group during the pre-flowering stage (Fig. 3).
Effect of IAA on CH flowers
The effects of IAA supplementation on CH flowers are complex and do not exhibit a consistent pattern. Application of 50 mg/L IAA during the sprouting stage increases CH flower numbers by 14.7% though the effect is not statistically significant (F = 11.35, P = 0.057; Fig. 3). However, a high concentration of 200 mg/L IAA applied during the pre-flowering stage results in a significant (F = 22.46, P < 0.05) increase of 19.1% in CH flower numbers. Continuous application of IAA across both stages either suppresses CH flowering or has no significant effect (Fig. 3).
Effect of ZT on CH flowers
Supplementation with ZT consistently impacts CH flowers, with no significant effect at low (10 mg/L) to medium (100 mg/L) concentrations. However, at high concentrations (200 mg/L), it exhibits an inhibitory effect during both the sprouting and pre-flowering stages (Fig. 3). Continuous application of 200 mg/L ZT across both stages significantly (F = 127.54, P < 0.001) reduces CH flower numbers by 38.7% compared to the control group (Fig. 3).
Effect of exogenous hormones on the number of CL flowers.

Effects of gibberellic acid (GA3), abscisic acid (ABA), indole-3-acetic acid (IAA), and zeatin (ZT) on CH flower numbers at different growth stages in Pseudostellaria heterophylla.(A) sprouting stage; (B) pre-flowering stage; (C) both stages combined. hormones concentrations unit is mg/L.
Effect of GA3 on CL flowers
Supplementation with high concentration GA3 (200 mg/L) is most effective during the sprouting stages, significantly (F = 197.26, P < 0.001) increasing CL flower numbers by 150.94% from an average of 5.3 ± 1.6 flowers per plant in the control group to 13.3 ± 3.4 flowers (Fig. 4). This effect is significantly stronger than observed during the pre-flowering and continuous application across both stages (Fig. 4).
Effect of ABA on CL flowers
Low-concentration ABA (10 mg/L) during the sprouting stage significantly (F = 168.34, P < 0.001) increases CL flower numbers by 141.51% from an average of 5.3 ± 1.6 flowers per plant to 12.8 ± 2.7 flowers (Fig. 4). While there is slight promotion of CL flowers during the pre-flowering stage or with continuous application, the effect is not as significant as during the sprouting stage. A high concentration of ABA (200 mg/L) shows no significant impact and even exhibits some inhibitory effect (Fig. 4).
Effect of IAA on CL flowers
Supplementation of low-concentration 10 mg/L IAA during the sprouting stage increases CL flower numbers by approximately 79.25%. However, the effect gradually diminishes with increasing concentration (Fig. 4). In the pre-flowering and continuous application stages, only the 100 mg/L IAA treatment promotes CL flower numbers, though the effect is less pronounced than the low-concentration treatment during the sprouting stage (Fig. 4).
Effect of ZT on CL flowers
During the pre-flowering stage, low-concentration ZT promotes CL flowers, while high-concentration ZT treatment reduces CL flowers, though not significantly (F = 15.42, P > 0.05) from 5.3 ± 1.6 in the control group to 4.8 ± 0.9, a 9.4% reduction (Fig. 4). Continuous treatment at both the pre-flowering and two stages periods shows a moderate promotion effect on CL flowers, with increasing concentration enhancing the promotion effect. However, it is less effective than the low-concentration treatment during sprouting (Fig. 4).

Effects of gibberellic acid (GA3), abscisic acid (ABA), indole-3-acetic acid (IAA), and zeatin (ZT) on CL flower numbers at different growth stages in P. heterophylla. (A) Sprouting stage; (B) pre-flowering stage; (C) both stages combined.
Effect of exogenous hormones on root weight P. heterophylla
GA3 significantly increases the root weight of P. heterophylla, particularly at a high concentration (200 mg/L) during the pre-flowering stage, where the fleshy root weight increases from 0.9 ± 0.3 g (control) to 1.4 ± 0.4 g, a 55.6% increase (F = 297.26, P < 0.001; Fig. 5). Continuous application from the sprouting stage to pre-flowering stages results in reduces in total root weight, indicating inhibitory effects (Fig. 5). Other hormones show minimal or no significant effect on root weight. For example, 100 mg/L IAA during the sprouting stage increases root weight by only 5.5% with no significant difference (F = 7.446, P > 0.05; Fig. 5). Certain concentrations, such as 50 mg/L ZT during the sprouting stage, inhibit root weight, reducing it to 0.4 ± 0.2 g (F = 99.13, P < 0.001; Fig. 5).

Effects of gibberellic acid (GA3), abscisic acid (ABA), indole-3-acetic acid (IAA), and zeatin (ZT) on fleshy root weight at different growth stages in Pseudostellaria heterophylla. (A) Sprouting stage; (B) pre-flowering stage; (C) both stages combined.
Discussion
Plant flowering is largely regulated by the levels of endogenous hormones, which are influenced by environmental factors. In addition, the application of exogenous hormones can also exert significant effects on flowering patterns providing a foundation for utilizing these exogenous hormones to alter the reproductive strategies of CH-CL plants18,19,20,21.
Role of exogenous hormones in regulating CH flower production
Flower bud formation is regulated by a complex interplay of external environmental factors and endogenous plant hormone signals, with GA3 playing a central role in this process8. GA3 signaling modulated the expression of genes responsible for flower formation in leaves, with flower initiation signals transmitted from leaves to apical meristem, activating the ‘flowering gene’9. In our study, both GA3 and ABA effectively promote the number of CH flowers, although the time of application and concentration had varying effects. Our findings are consistent with previous research reports showing that GA3 and ABA can increase the number of flowers in Rhododendron and Vanilla22,23. The application of GA3 on young leaves has been shown to advance flowering and increase flower numbers in strawberries24.
However, continuous application of hormones from the sprouting stage to the pre-flowering stage yielded only modest effects on flower promotion, suggesting both time and concentration of exogenous hormone application are crucial for optimal results. We speculate that the flowering time and sequence of mixed mating plants, such as P. heterophylla, are influenced by environmental factors, such as light duration and temperature11. Moreover, the number of flowers is intricately related to the level and ratio of endogenous hormones25. Continuous application of exogenous hormones may lead to the accumulation of excessive hormone levels or alter endogenous hormone ratios, thereby reducing their effectiveness in promoting flower formation.
Influence of exogenous hormones on CL flower development
Early application of GA3 and ABA effectively promoted the formation of CL flowers in P. heterophylla. However, treatment with ZT at a high concentration during the pre-flowering stage resulted in a relative reduction in the number of CL flowers, though this difference was not statistically significant (P > 0.05). CL flowers in plants with a CH-CL mating system, such as P. heterophylla, adapt a feature that enables self-pollination, particularly in heterogeneous environments. The reproductive strategy is characterized by reducing floral structures such as petals, stamens, and pollen, allowing plants to allocate more resources to seed production. Research indicated that approximately 90% of P. heterophylla seeds originate from CL flowers12,17.
The exogenous application of plant hormones has been shown to promote the formation of CH flowers21, and as demonstrated in our study, these hormones can also enhance CL flower production. However, ZT at high concentrations showed a contrasting effect, exhibiting some inhibitory impact on CL flower formation. This observation diverges from the general inhibitory effects of high concentrations of the four hormones on CH flower production. It is hypothesized that this phenomenon may be due to the role of ZT in promoting more branching and leaf growth at high concentrations. This increased vegetative growth could lead to a redistribution of resources within the plant, affecting the energy allocation towards reproductive structures, including CL flowers.
Role of exogenous hormones in root growth variation
Fleshy roots developed from adventitious roots, as the reproductive output form of P. heterophylla, influence the genetic structure of its population12. Only high concentration (200 mg/L) GA3 treatment during the pre-flowering stage significantly promoted root weight among the four exogenous hormones used. The effects of the other hormones were not significant, including 100 mg/L IAA treatment, which slightly increased root weight (Fig. 4). ABA or ZT treatments did not promote root weight or even had inhibitory effects at some concentrations. This contrasts with previous studies, showing that IAA is key for root growth during the nutritional growth period (CL flowering period) of P. heterophylla and that ZT can also promote root weight26. This discrepancy is likely due to the difference in hormone treatment timing. In the above studies, treatment was applied during the CL flowering period, whereas our study focused on the sprouting or the pre-flowering period of CH, leading to different results.
GA3 regulates plant growth and development in the middle stage of flower development, while IAA and cytokinin (CK) regulate stigma and stamens growth and development during the middle and late stages, and ABA participates in flower bud differentiation and stigma and stamens growth and development in the late stage27. Therefore, different treatment timings likely explain the contrasting results.
The current study results showed all four exogenous hormones promoted CL flower formation; they had no significant effect on root weight and, in some cases, inhibitory effects. Correlation analysis between CL flower number and root weight revealed a significant negative correlation (y=-0.017x + 3.096, r2 = 0.377, p < 0.05), indicating a trade-off relationship between resources allocated to CL flowers and root weight during the entire growth and development period of P. heterophylla. Resources are initially allocated to CL flowers and seeds but depleted later in adventitious root expansion, which reduces root weight. Resource allocation in reproducing plants is shaped by physiological trade-offs, where increased investment in one activity must be offset by reduced investment in others28. Plants with plastic reproductive strategies display such trade-offs29,30. However, no trade-off relationship was found between the CH flowers, CL flowers and root weight (p > 0.05) because the number of CH flowers is few, and their resources used for flowering and seeds are mostly from mother roots, which have a low proportion. Even if exogenous hormones promote the CH flower numbers and seed sets, their influence on root weight remains insignificant (author, unpublic data).
Conclusion
In conclusion, exogenous hormones, particularly GA3 and ABA, significantly influence flowering and root growth in P. heterophylla. GA3 and ABA promoted CH and CL flower numbers, with the most significant effects observed at specific concentrations during the sprouting and pre-flowering stages. However, continuous hormone application across both stages resulted in modest effects, highlighting the importance of optimal timing and concentration for achieving desired outcomes. ZT, especially at high concentrations, exhibited inhibitory effects on CH flowers and root weight, suggesting a trade-off in resource allocation between vegetative and reproductive growth. The negative correlation between CL flower numbers and root weight further supports this hypothesis of resource allocation trade-offs. Overall, the results emphasize the complex role of exogenous hormones in regulating plant growth and reproductive strategies, providing insights into optimizing flowering and root development in P. heterophylla.
Data availability
Data is provided within the manuscript or supplementary information files.
References
Davis, S. J. Integrating hormones into the floral-transition pathway of Arabidopsis Thaliana. Plant. Cell. Environ. 32(9), 1201–1210 (2009).
Campos-Rivero, G. et al. Plant hormone signaling in flowering: an epigenetic point of view. J. Plant. Physiol. 214, 16–27 (2017).
Marhava, P. et al. A molecular rheostat adjusts auxin flux to promote root protophloem differentiation. Nature 558(7709), 297–300 (2018).
Nguyen, T. N., Tuan, P. A., Mukherjee, S., Son, S. & Ayele, B. T. Hormonal regulation in adventitious roots and during their emergence under waterlogged conditions in wheat. J. Exp. Bot. 69(16), 4065–4082 (2018).
Chen, K. et al. Abscisic acid dynamics, signaling, and functions in plants. J. Integ Plant. Biol. 62(1), 25–54 (2020).
Sakuraba, Y. et al. Multilayered regulation of membrane-bound ONAC054 is essential for abscisic acid-induced leaf senescence in rice. Plant. Cell. 32(3), 630–649 (2020).
Waadt, R. et al. J. I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell. Bio. 23(10), 680–694 (2022).
Conti, L. Hormonal control of the floral transition: can one catch them all? Dev. Biol. 430(2), 288–301 (2017).
Yang, Z. Q. et al. Changes of endogenous hormone level during flower development in on-season and off-season Longan. J. Anhui Agric. Sci. 47, 47–74 (2019).
Minter, T. C. & Lord, E. M. Effects of water stress, abscisic acid, and gibberellic acid on flower production and differentiation in the cleistogamous species Collomia grandiflora Dougl. Ex Lindl. (Polemoniaceae). Am. J. Bot. 70(4), 618–624 (1983).
Culley, T. M. & Klooster, M. R. The cleistogamous breeding system: a review of its frequency, evolution, and ecology in angiosperms. Bot. Rev. 73(1), 1–30 (2007).
Zhang, L. H., Sun, Q., Zhao, J. M. & Zhang, Y. W. Plasticity in the reproductive strategy of a clonal cleistogamous species, Pseudostellaria heterophylla. Plant. Ecol. 219, 1493–1502 (2018).
Le Corff, J. Effects of light and nutrient availability on chasmogamy and cleistogamy in an understory tropical herb, Calathea micans (Marantaceae). Ame J. Bot. 80(12), 1392–1399 (1993).
Koul, M. & Sharma, N. Rates and pattern of ovule abortion vis-à-vis in situ pollen germination in some populations of Trifolium fragiferum L. J. Bio. 37, 1067–1077 (2012).
Li, Q. et al. Expression of B-class MADS-box genes in response to variations in photoperiod is associated with chasmogamous and cleistogamous flower development in Viola Philippica. BMC Plant. Biol. 16, 1–14 (2016).
Barnett, L. L., Troth, A. & Willis, J. H. Plastic breeding system response to day length in the California wildflower Mimulus Douglasii. Am. J. Bot. 105(4), 779–787 (2018).
Hu, W. Z., Zhao, J. M. & Zhang, Y. W. Fitness advantage and maintenance mechanisms of dimorphic mixed-mating plants. Biodivers. Sci. 27(04), 468–474 (2019).
Hu, D. J., Shakerian, F., Zhao, J. & Li, S. P. Chemistry, Pharmacology and analysis of Pseudostellaria heterophylla: a mini-review. Chin. Med. 14(1), 21 (2019).
Zheng, W. et al. Effects of exogenous Paclobutrazol and GA3 treatment on the accumulation of endogenous IAA and transcriptional level of related genes in the development of tuberous toot of Pseudostellaria heterophylla. Bio Bull. 35(2), 46–52 (2019).
20 Mesejo, C. et al. Gibberellin reactivates and maintains ovary-wall cell division causing fruit set in parthenocarpic Citrus species. Plant. Sci. 247, 13–24 (2016).
Chang, M. Z. & Huang, C. H. Effects of GA3 on promotion of flowering in Kalanchoe spp. Sci. Hortic. 238, 7–13 (2018).
Zhang, X. M., Yi, Y., Yang, H., Gao, W. & Wang, M. Y. Effect of external hormone on the florescence regulation of Rhododendron irroratum in Bai-li Du-juan tourist attractions. North. Hortic. 12, 170–173 (2009).
Wang, H. et al. Effects of plant external hormones on flower bud differentiation of vanilla. Chin. J. Trop. Agric. 36, 72–76 (2016).
Paroussi, G., Voyiatzis, D. G., Paroussis, E. & Drogoudi, P. D. Growth, flowering and yield responses to GA3 of strawberry grown under different environmental conditions. Sci. Hortic. 96(1–4), 103–113 (2002).
Kolachevskaya, O. O. et al. Auxin synthesis gene tms1 driven by tuber-specific promoter alters hormonal status of Transgenic potato plants and their responses to exogenous phytohormones. Plant. Cell. Rep. 36, 419–435 (2017).
Li, J. et al. Regulation of GA3 on the development of tuberous root and gibberellins biosynthesis of Pseudostellaria heterophylla. Mol. Plant. Breed. 16(20), 6867–6874 (2018).
Wen, J. Y. et al. Development process of pistils and stamens and effect of endogenous hormone content on style growth in Plumbago auriculata Lam. With distyly. Plant. Sci. J. 4(02), 263–270 (2016).
Barrett, S. C. Influences of clonality on plant sexual reproduction. P Natl. Sci. 112(29), 8859–8866 (2015).
Zhang, D. et al. GA4 and IAA were involved in the morphogenesis and development of flowers in Agapanthus praecox ssp. Orientalis. J. Plant. Physiol. 171(11), 966–976 (2014).
Fazlioglu, F. & Bonser, S. P. Phenotypic plasticity and specialization in clonal versus non-clonal plants: A data synthesis. Acta Oecol. 77, 193–200 (2016).
Acknowledgements
The authors thank Chunfeng Yang for his valuable suggestions on an earlier version of this manuscript. We also thank Shengjun Huang and Lihui Zhang for their help in the field.
Funding
This research was supported by grants from the National Natural Science Foundation of China (32272757), Liaoning province science and technology plan-combined plan (2024JH2/102600132), Changchun Normal University (CSJJ2023008ZK).
Author information
Authors and Affiliations
Contributions
Yanwen Zhang and Jimin Zhao designed the experiments. Qi Sun and Lei Wu collected data. Qi Sun wrote the original manuscript. Yanwen Zhang and Lei Wu revised the manuscript and figures. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
All plant materials were used in accordance with national and international standards and local laws and regulations. The use of all plant materials does not pose any risk to other species in nature. No specific permission is required for the collection of all samples described in this study. The specimens of the research species were kept in the herbarium of Liaodong University, No. CP2002-3, and the appraiser by Professor Zhang Yanwen.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sun, Q., Wu, L., Zhao, JM. et al. Exogenous hormones regulate the sequentially reproductive output in Pseudostellaria heterophylla with a chasmogamous–cleistogamous mixed mating system. Sci Rep 15, 41580 (2025). https://doi.org/10.1038/s41598-025-25479-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-25479-2
