
Abstract
This study investigates the role of water level fluctuations (WLF) in shaping phytoplankton community structure and gross primary productivity (GPP) in Hongfeng Reservoir, a karst reservoir in Southwest China, through long-term monitoring (2016–2023) across seven sampling sites. Results revealed substantial seasonal and interannual variations in water levels (maximum fluctuation range: 9.78 m) and nutrient concentrations, with total nitrogen (TN) and phosphorus (TP) peaking during wet seasons. Chlorophyta dominated species diversity (42–48%), while Cyanophyta exhibited the highest relative abundance. GPP fluctuated markedly between the dry (586.09 mg C/m²/d) and wet seasons (3018.66 mg C/m²/d), driven by phytoplankton abundance and community stability. The Bray-Curtis (BC) dissimilarity index declined over time, showing a negative correlation with WLF (r = -0.03) and a significant negative impact on GPP. WLF indirectly enhanced GPP by altering community dynamics. Findings highlight WLF as a critical regulator of phytoplankton structure and productivity, emphasizing the importance of hydrological management in mitigating algal blooms and balancing ecological stability. This study provides actionable insights for optimizing water level regulation to sustain reservoir ecosystem health in karst regions.
Similar content being viewed by others
Introduction
In recent years, global climate change and frequent extreme weather have severely threatened aquatic ecosystems1,2. By the end of this century, human-induced climate change may exacerbate warmth and aridity in many arid and semi-arid regions, and drought-induced water level decline will exacerbate eutrophication in water bodies, leading to increased cyanobacterial biomass and dominance in tropical aquatic systems3. Global climate change has also resulted in significant alterations in temperature and water level fluctuations (WLF) in reservoirs worldwide. Moreover, water temperature directly influences phytoplankton in reservoirs through physiological and individual developmental processes (e.g., cell growth and division) while WLF primarily affects it indirectly by altering key environmental parameters such as water flow velocity, transparency, and nutrient availability4,5.Reservoirs provide a range of services to humans and are a key component of global geographical, ecological, and hydrological processes6. The vulnerability of reservoirs to climate change may result in substantial alterations in water levels, storage capacity, and inundation areas. Moreover, WLF can even affect the slope stability in reservoirs7. Reservoirs in karst areas are crucial for groundwater development and water supply due to their complex hydrogeological structures and effective storage and runoff capacities8. In addition, karst reservoirs are characterized by pronounced fluctuations in water volume, elevated concentrations of calcium and magnesium ions, substantial water level variability, and strong connectivity with groundwater systems9.
Water level fluctuations (WLF) influence phytoplankton community dynamics, biomass, and species composition by altering mixing processes, biogeochemical cycles, oxygen dynamics, and photosynthetic activity10,11,12. For example, WLF can reduce cyanobacterial biomass by changing hydrodynamic conditions13, while scientific water level management maintains healthy aquatic plant beds to improve water quality14. Amplitude, frequency, and duration of WLF are considered the most relevant factors affecting the functioning of aquatic ecosystems15,16,17. Intense WLF can impact water quality, potentially triggering algal blooms by altering phytoplankton community structure18.The water level of a reservoir is often regarded as a crucial indicator of the water balance within the reservoir system, determined by the distribution of hydrological processes in the catchment area, such as precipitation, runoff generation, and evapotranspiration19. WLF have significant effects on the functional and structural attributes of aquatic biological communities20. The interannual and intra-annual fluctuations in water level are considered to be key drivers that control phytoplankton productivity and species diversity21. Intense WLF may even alter environmental parameters such as nutrient concentrations, light availability, and temperature, thereby influencing phytoplankton primary productivity and community composition22.
Previous studies on water level fluctuations (WLF) in reservoirs focused on the relationships between WLF and water quality, water trophic state, phytoplankton, and littoral plants. However, as far as we know, research on how WLF impact phytoplankton community structure and gross primary productivity (GPP) in karst reservoirs remains relatively limited. Hongfeng Reservoir is a major drinking water source for Guiyang, providing daily water supply for the city’s residents. The stability and safety of its aquatic ecosystem directly impact regional public health security and social sustainable development. This unique functional role renders the reservoir significantly important for research. Thus, the objectives of this study are: (1) to examine the trends in WLF and physicochemical parameters of Hongfeng Reservoir; (2) to analyze the variations in phytoplankton community structure and stability; and (3) to elucidate the spatiotemporal patterns of primary productivity in Hongfeng Reservoir, thereby revealing the mechanisms by which WLF influence primary productivity.
Materials and methods
Research area
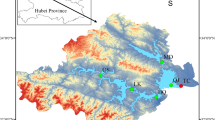
The Hongfeng Reservoir is located in Guiyang City, Guizhou Province, southwestern China (20° 26ʹ–26°35ʹ N, 106° 19ʹ–106° 28ʹ E). The reservoir was constructed by damming the Maotiao river in 1960. The reservoir is primarily divided into two zones, the northern and southern reservoir areas (Fig. 1). The Hongfeng Reservoir is located upstream of the Maotiao River, a major tributary of the Wujiang River in central Guizhou Province, with a basin area of 1,596 km2. The reservoir can store 6.01 × 108 m3 of water, and the mean water depth is 11 m. The Hongfeng Reservoir basin is in a subtropical monsoon humid climate zone with an average annual temperature of 14.4 °C and an average annual precipitation of 1,174.7 ~ 1,386.1 mm. The reservoir supplies 70% of Guiyang’s urban water consumption and has played a vital role in Guizhou’s socioeconomic development.

Sampling Area (In a and b, color gradients represent elevation, where red indicates high altitude and blue indicates low altitude.)
Sample collection and analysis
A total of 7 sampling points were selected from upstream to downstream along the reservoir, namely Pianshanzhai (PSZ), Yaodong (YD), Dam (DB), Huayudong (HYD), Xijiaoshuichang(XJSC), Houwu (HW) and Sanchahe (SCH) (Fig. 1). This study divided each year into four distinct hydrological periods for analysis and collected eight years of sample data spanning 2016 to 2023. Surface water samples, were collected using a water sampler, and properly stored in 1.5 L containers for subsequent analysis later in the laboratory. A portable multi-parameter water quality analyzer (model HI 98194; Harderwood, Italy) was used to measure water temperature (WT, °C), dissolved oxygen (DO, mg/L), and pH. Transparency (SD) was measured in situ with a Secchi disk (model XH-SSP; Xinhu Company, Shanghai, China), and photosynthetically active radiation (PAR) intensity was measured with a split-type spectral radiometer (model AZ8583) at Hongfeng Reservoir.
Laboratory analysis
This study used standard methods issued by SEPB (2002) to analyze water quality, determining the following indicators: total nitrogen (TN, mg/L), ammonia nitrogen (NH₃-N, mg/L), total phosphorus (TP, mg/L), permanganate index (CODₘₙ, mg/L). Furthermore, this study used the spectrophotometric method specified in the standard method of the National Water Conservancy Industry (SL 88-2012) to determine chlorophyll-a (Chl-a, mg/m³). For phytoplankton, 15 mL of Lugo reagent (1:100) was added to 1.5 L samples, which were settled for 24–48 h and concentrated to 80 mL. Phytoplankton were identified and counted in a 0.1 mL chamber using an optical microscope (OLYMPUS CX43) at 400× magnification.
Data analysis
Four different hydrological periods were used in this study: the dry season (DS, from December to March), the rising period (RP, from April to May), the wet season (WS, from June to July), and the falling period (FP, from August to November). The classification of these four hydrological periods was mainly based on water level variation.
Calculation of WLF
Water level data was obtained from the Hongfeng Reservoir Authority in Guizhou Province. Daily water level data from Hongfeng Reservoir were used to calculate monthly mean water levels through mathematical operations. The daily dynamic data were also analyzed to determine annual fluctuation amplitudes and extreme values. Finally, both daily and monthly water level dynamics were correlated with monthly phytoplankton sampling data to explore potential relationships.
Calculation of euphotic depth
Zeu: euphotic depth. Kd (PAR): diffuse attenuation coefficient of photosynthetically active radiation. Because water transparency also indicates the degree of light attenuation in water and is a conventional and easily obtainable parameter, the diffuse attenuation coefficient of photosynthetically active radiation was calculated using water transparency23.
The value of f in this study was 1.4. This study calculated the diffuse attenuation coefficient of photosynthetically active radiation using the transparency of the Hongfeng Reservoir water body and then calculated the euphotic depth (Zeu) of the water body. The euphotic depth is defined as the depth of the water column where photosynthetically active radiation (PAR) is ≥ 1% of the surface irradiance.
Calculation of GPP
Gross primary productivity (GPP) was estimated based on the vertically generalized production model (VGPM) following Behrenfeld and Falkowski24.
PPeu: primary productivity from surface to true light layer (mg C/m2/d). \(\:{P}_{opt}^{B}\): maximum carbon fixation rate of the water column. Cz: Chlorophyll-a concentration at depth z. dz: thickness of water layer. Ez: PAR intensity at depth z. Eopt: PAR intensity at the depth of\(\:\:{P}_{opt}^{B}\). βd: the initial slope of the P-I curve (the correspondence between photosynthesis rate and light intensity). Dirr: photoperiod. Zeu: euphotic depth. Further simplifying the model yields the core formula for calculating GPP.
Copt: surface chlorophyll-a (Chl.a) concentration. E0: PAR intensity on the lake surface. Zeu: true light layer depth (When the water depth is less than the true light layer depth, use water depth instead). Dirr: photoperiod. The photosynthesis of chlorophyll-a is mainly controlled by enzymes, and enzyme activity is largely controlled by temperature. Therefore, it is generally believed that \(\:{P}_{opt}^{B}\) is a function of surface temperature and has multiple conversion relationships.
Statistics
The sampling area map were created using ArcMap (Version 10.8, URL: https://www.arcgis.com/). The Bray-Curtis dissimilarity index (BC; where a larger BC value indicates greater community instability, and a smaller value indicates greater stability) was calculated using the “vegan” package in R (version 4.0.0). Phytoplankton dominance was calculated using the “readxl” and “openxlsx” packages. Daily and monthly dynamics of water levels, violin plots of physicochemical indicators, pie charts of phytoplankton structural composition, abundance proportion charts, trends of the Bray-Curtis dissimilarity index over seasons, relationships between the average Bray-Curtis index and water level changes during different hydrological periods, and fitting analyses of the Bray-Curtis index and GPP were constructed using Origin 2024b. IBM SPSS Statistics 26 was used for conducting univariate significance analysis of various physicochemical indicators. In R (version 4.0.0), the “FactoMineR” and “factoextra” packages were employed to perform principal component analysis (PCA) on phytoplankton abundance data across different hydrological periods. Principal component analysis (PCA) was performed on the phytoplankton abundance matrix to reveal primary variation in community structure across periods. Using the vegan package in R, envfit() was then applied to fit environmental variables (e.g., DO, pH, TN, TP, WT, SD) to the PCA ordination, with arrow direction indicating association trends and length reflecting correlation strength, identifying drivers of phytoplankton dynamics. This study utilized R packages (version 4.0.0) including devtools, linket, readxl, tidyverse, openxlsx, plspm, dplyr, ggplot2, and ggpubr to perform Spearman correlation analysis and partial least squares modeling (PLS). This analysis aims to investigate the effects of water level fluctuations on phytoplankton community stability and gross primary productivity (GPP).
Results
Changes in water level fluctuations
The water level of Hongfeng Reservoir exhibited significant variations across different years and hydrological periods. Over the eight-year study period, the interannual water level ranged from 1230.03 m to 1239.81 m. The maximum inter-annual fluctuation amplitude over the 8-year period was 9.78 m, while the average annual fluctuation amplitude was 5.49 m. Whereas the minimum and maximum intra-annual fluctuation amplitudes were 2.76 m (in 2019) and 6.11 m (in 2020), respectively (Fig. S1a, Fig. S1b). Notably, water levels in 2023 were consistently among the lowest observed throughout the entire eight-year period. Across different hydrological periods, the mean water level fluctuation amplitudes were 5.41 m (dry season); 4.55 m (rising period); 5.99 m (wet season); and 6.03 m (falling period). The fluctuation amplitudes were smallest during the falling period (6.27 m) and largest during the dry season (8.98 m), with the wet season showing the second-largest amplitude (8.84 m) (Fig. S1c). In the monthly fluctuation range of water levels, the largest fluctuation occurred in June 2017 with an amplitude of 4.15 m, while the smallest fluctuation was observed in January 2019, where the water level fluctuated by only 0.03 m (Fig. S1d).
Changes in physical and chemical factors
Interannually, with TN and TP exhibiting significant responses to interannual water level variations (Fig. S2). The results of one-way ANOVA indicated that the physicochemical characteristics of Hongfeng Reservoir varied significantly among different hydrological periods (n = 672, P < 0.01). Among them, WT (n = 672, p < 0.001) (Fig. 2a), TN (n = 672, p < 0.001), CODMn (n = 672, p = 0.003), and NH3-N (n = 672, p < 0.001) varied significantly, with ranges of WT (6.7–29.2 ℃), TN (0.56–3.70 mg/L), CODMn (1.40-4.00 mg/L), and NH3-N (0.004–0.612 mg/L). Additionally, TN, TP, CODMn, and NH3-N all reached their maximum values during the wet season, with concentrations of 3.70 mg/L, 0.08 mg/L, 4.00 mg/L, and 0.612 mg/L, respectively (Fig. 2b and c). From the dry season (DS) to the falling period (FP), WT, TN, TP, CODMn, NH3-N and pH generally exhibited an increasing trend, while SD and DO showed a decreasing trend (Fig. 2d). SD and DO reached their minimum values of 0.5 m and 4 mg/L during the wet season (WS) and falling period (FP), respectively (Fig. 2).

Environmental changes during the hydrological periods. Violin plots showing the changes during the hydrological periods i.e. dry season (DS), rising period (RP), wet season (WS) and falling period (FP) of (a) water temperature (WT), transparency (SD), and dissolved oxygen (DO), (b) total nitrogen (TN) and permanganate index (CODMn), (c) total phosphorus (TP) and Ammonium nitrogen (NH3-N) and (d) pH. The data include the entire eight years dataset. The symbol “*” represents significant differences (p < 0.05).
Changes in phytoplankton community structure and stability in reservoir
During this study, a total of 119 species from 7 phyla of phytoplankton were identified, including 55 Chlorophyta species, 30 Bacillariophyta species, 22 Cyanophyta species, 4 Dinophyta species, 4 Euglenophyta species, 3 Chrysophyta species and 1 Cryptophyta species. Comparative analysis of phytoplankton community composition across different hydrological periods revealed the identification of 111, 114, 113, and 94 species during the dry season, rising period, wet season, and falling period, respectively. Among them, the proportion of Chlorophyta was the largest, reaching 42%, 43%, 47%, and 48%, respectively (Fig. 3a). Across different hydrological periods, the phytoplankton community in Hongfeng Reservoir was predominantly composed of Chlorophyta, Bacillariophyta, and Cyanophyta, with Chlorophyta representing the highest species richness, followed by Bacillariophyta and Cyanophyta; phytoplankton species diversity was the highest during the rising period and lowest during the falling period, while the wet season exhibited greater species richness than the dry season (Fig. 3a). The dominant species (with a dominance value > 0.02) were identified, and the top five species ranked by dominance in different hydrological periods were dry season (Pseudanabaena limnetica, Cyclotella meneghiniana, Syneddra acusva, Aulacoseira granulata, Cryptomonas sp.), rising period (Pseudanabaena limnetica, Cyclotella meneghiniana, Aulacoseira granulata, Syneddra acusva, Asterionella sp.), wet season (Pseudanabaena imnetica, Cyclotella meneghiniana, Syneddra acusva, Limnothrix sp., Achnanthes sp.), falling period (Pseudanabaena limnetica, Cyclotella sp., Syneddra acusva, Merismopedia punciata, Achnanthes sp.) (Table S1). The dominant algal species in Hongfeng Reservoir were primarily Pseudanabaena limnetica, Cyclotella sp., and Syneddra acusva. Across different hydrological periods, Cyanophyta consistently exhibited the highest relative abundance in Hongfeng Reservoir, with the following order: wet season > falling period > rising period > dry season. Phytoplankton abundance gradually increased from the dry season to the wet season but slightly declined during the falling period (Fig. S3). Four distinct clusters were identified in the PCA analysis related to hydrological periods (DS, FP, RP, WS). Specifically, the dry season (DS, blue dots) lies in Dim1 negative, Dim2 positive (variance contributions: 32.8%, 24.6%), associated with SD and CODMn. The flood (FP, yellow triangles) clusters in both Dim1 and Dim2 negative ranges, linked to NH3-N. The recession period (RP, gray squares) locates in Dim1 positive, Dim2 central, correlating with DO, pH, and TP. The wet season (WS, red crosses) occupies both Dim1 and Dim2 positive ranges, associated with precipitation, GPP, and WT. Dim1 and Dim2 capture 32.8% and 24.6% of total variance respectively, illustrating key gradients across these periods in the ordination space (Fig. 3b).
Analysis of Bray-Curtis dissimilarity for phytoplankton in Hongfeng Reservoir revealed a general declining trend in Bray-Curtis values from 2016 to 2023. Between 2016 and 2021, Bray-Curtis values exhibited significant fluctuations, while fluctuations were smaller from 2022 to 2023. The lowest Bray-Curtis value (0.09) was recorded in 2019, and the highest value (0.91) occurred in 2021 (Fig. S4a). Analysis of Bray-Curtis dissimilarity values across hydrological periods revealed a general decline in Bray-Curtis values from the dry season to the falling period, indicating a gradual increase in community stability as water levels rose from low to high. Furthermore, during the high-water period in the wet season, Bray-Curtis values exhibited a negative correlation with WLF (correlation coefficient = -0.03) within a specific water level range (1231.64–1238.64 m) (Fig. S4b, Fig. S5).

Structural composition of phytoplankton and principal component analysis with other physicochemical indices. (a) Structural composition of phytoplankton in different hydrological periods, (b) Principal Component Analysis (PCA) biplot illustrating the relationships between environmental variables and sampling periods across different seasons. The first two principal components (Dim1 and Dim2) explain 32.8% and 24.6% of the total variance, respectively. Seasons/Periods abbreviations: Dry Season (DS), RP (Rainy Period), Wet Season (WS), falling period (FP). Environmental variables: Total Nitrogen (TN), Total Phosphorus (TP), Ammonia Nitrogen (NH₃-N), pH, Secchi Depth (SD), gross primary productivity (GPP), Water Temperature (WT).
Changes in growth primary productivity
The GPP of Hongfeng Reservoir varied significantly across different hydrological periods, showing increases primarily during the rising period, wet season, and falling period, while decreasing during the dry season (Fig. 4). The mean GPP during the wet season (3018.66 mg C/m²/d) was significantly higher than that during the dry season (586.09 mg C/m²/d), rising period (2030.75 mg C/m²/d), and falling period (1448.80 mg C/m²/d). The wet season exhibited the largest GPP variation range (285.35–8809.87 mg C/m²/d), including the maximum GPP value (8809.87 mg C/m²/d), while the dry season showed the smallest GPP variation range (73.37–5284.35 mg C/m²/d), including the minimum GPP value (73.37 mg C/m²/d). Except for the SCH sampling site, GPP at most sampling sites was higher during the wet season and falling period compared to the dry season and rising period. Additionally, results from the PLS model indicated that WLF exerted an overall indirect positive effect on GPP, with this effect being particularly evident during the wet season (Figs. 4 and 5). Specifically, during different hydrological periods, water level fluctuations (WLF) exert indirect positive effects on gross primary productivity (GPP) in the wet season and dry season (Fig. S6).
Chl-a concentrations at most sampling sites were highest during the wet season and lowest during the dry season. The mean Chl-a concentration during the wet season (15.70 mg/m³) was significantly higher than that during the dry season (4.12 mg/m³), rising period (8.95 mg/m³), and falling period (8.91 mg/m³). The dry season exhibited the largest variation range in Chl-a (0.35–49.6 mg/m³), while the rising period showed the smallest range (1.28–15.1 mg/m³) (Fig. 4).

Changes in gross primary productivity (GPP) and chlorophyll-a (Chl.a) during different hydrological periods.

Relationships among water level fluctuation (WLF), Bray-Curtis values (BC), and gross primary productivity (GPP). The partial least squares (PLS) model plots show (a) the direct and indirect impact relationships among WLF, physico-chemical indices, and GPP, and (b) the correlation between water level fluctuation, physico-chemical factors, and Bray-Curtis Values.

Impact analysis of Bray-Curtis values (BC) on gross primary productivity (GPP). (a) Fitting analysis between community Bray-Curtis values and gross primary productivity, (b) Trends in productivity and Bray-Curtis values at different stages.
Discussion
The effects of regulated WLF on nutrient dynamics
Over long-term WLF, nutrient concentrations in reservoirs are influenced by water levels, subsequently affecting water quality. During periods of intense WLF, these fluctuations can become the dominant factor. Currently, the world is facing significant challenges from global warming, which promotes the proliferation of many algal species in warm and nutrient-rich environments25. With the increasing frequency of extreme climate events, significant changes in water levels can impact the physical, chemical, and biological processes of reservoir ecosystems, compromising their health and integrity26. For instance, WLF can indirectly influence phytoplankton communities by altering physicochemical conditions27. This study primarily focuses on comparing differences between seasons (i.e., periods with distinct water levels). This is because analyses conducted by season exhibit more pronounced differences, and furthermore, subdividing the analysis into individual seasons yields greater reference value for the present study. The results of this study indicate that water levels in Hongfeng Reservoir exhibit significant variability over long-term scales, with a maximum fluctuation amplitude of 9.78 m over an eight-year period. Among different interannual fluctuations, the smallest fluctuation (2.76 m) occurred in 2019, while the largest (6.11 m) was observed in 2020 (Fig. S1). TN and TP showed monthly fluctuations, with TN exhibiting greater variability and increasing with rising water levels, while TP changes were relatively minor (Fig. S1). Water level fluctuation may change the interaction between water and sediment, affecting N and P availability28. Overall, nutrient concentrations in Hongfeng Reservoir showed an increasing trend from the low-water dry season to the high-water wet season, likely due to increased rainfall during the wet season enhancing surface runoff and introducing substantial external nutrients, thereby elevating nutrient concentrations. Numerous studies have investigated the effects of long-term WLF. For example, lowering water levels can increase nutrient concentrations and enhance cyanobacterial proliferation and dominance29. The long-term WLF have led to a general increasing trend in total phosphorus concentration in surface sediments; moreover, the alternating processes of sediment deposition and hydraulic erosion under prolonged water level variations significantly enhance the downstream migration of phosphorus30. Furthermore, prolonged water-level fluctuations have led to a significant reduction in riparian plant species composition within canyon geomorphic areas31.
The effects of WLF regulation on the structure and stability of phytoplankton communities in reservoir
Complex aquatic interactions exist between reservoir water level, hydrogeochemical cycling, and community distribution32,33. The communities of large plants, phytoplankton, microorganisms, and benthic invertebrates had been shown to vary with changes in water levels in reservoirs34. Consequently, the composition and stability of the phytoplankton community vary across different hydrological periods. The fluctuation of the water level in Hongfeng Reservoir was negatively correlated with the Bray Curtis (BC) index of the phytoplankton community (Fig. S4). In highly dynamic reservoir systems, WLF significantly influence physicochemical conditions and phytoplankton community dynamics35. Across the dry season, rising period, wet season, and falling period, the phytoplankton community was predominantly composed of Chlorophyta, with Cyanophyta exhibiting the highest relative abundance in all periods. From the dry season to wet season, both phytoplankton species richness and abundance showed a gradual increasing trend, while from the wet season to falling period, they declined. These findings suggested that WLF are a key factor driving differences in phytoplankton community structure across hydrological periods. Moreover, from the dry season to the wet season, both the number of phytoplankton species and phytoplankton abundance showed a gradual increasing trend. From the wet season to the falling period, both the number of species and phytoplankton abundance decrease. This indicated that one important factor causing the differences in phytoplankton communities during different hydrological periods is WLF. This finding aligns with previous research conducted in Zhoucun Reservoir, which revealed that rainfall-induced nitrogen variability constitutes a primary determinant of phytoplankton dynamics across contrasting hydrological years36. In contrast, other relevant studies have collectively demonstrated that phytoplankton community dynamics are regulated by complex interactions among multiple environmental factors, including water temperature, thermal stratification, light availability, nutrient accessibility, and WLF37. A decline in water levels may increase sediment resuspension and turbidity, thereby influencing phytoplankton biomass and community structure38. Across interannual variations and hydrological periods, the Bray-Curtis (BC) dissimilarity values exhibited an overall declining trend, with the lowest mean BC value occurring in 2019 and the highest in 2021. Additionally, BC values during the wet season were generally lower than those during the dry season, indicating higher phytoplankton community stability in the wet season compared to the dry season. Species diversity can increase or decrease community stability39. Community stability largely depends on its ability to resist external disturbances40. This study demonstrates that the stability of the phytoplankton community in Hongfeng Reservoir is significantly influenced by WLF. This result is consistent with previous research indicating that seasonal WLF in Poyang Lake play a crucial role in phytoplankton succession41. Furthermore, studies had shown that water levels at 14–15 m play a decisive role in shaping phytoplankton community patterns and succession through the combined effects of scouring, dilution, and biological competition42,43.
The effects of phytoplankton community stability on GPP under water level fluctuation regulation
Based on the overall results of the 8-year study, hydrodynamic changes are key drivers of the productivity and structure of freshwater ecosystems, and they indirectly influence reservoir primary productivity by regulating phytoplankton community composition and nutrient loading (Fig. 6a and b). Analysis stratified by hydrological periods reveals that water level fluctuations (WLF) exert indirect positive effects on the wet season and dry season (Fig. S6). Other studies have also highlighted that hydrological systems are critical drivers of freshwater ecosystem productivity and structure, and they are increasingly affected by human activities44. This study demonstrates that the Bray-Curtis index of phytoplankton community in Hongfeng Reservoir exhibited a negative correlation with primary productivity from 2016 to 2023 (Fig. 6). Specifically, the gross primary productivity (GPP) decreased with increasing Bray-Curtis index, indicating that the primary productivity of phytoplankton declined as the community stability decreased (Fig. 6a). These findings highlight the critical role of phytoplankton in primary productivity, nutrient cycling, and energy transfer within aquatic ecosystems45. During the eight-year monitoring period in Hongfeng Reservoir, significant changes occurred in GPP and community BC values across the four different hydrological periods. From the dry season to the wet season, GPP gradually increased. WLF strongly affected the GPP of phytoplankton46, and increases in interannual WLF lead to increases in plant productivity47. The GPP of phytoplankton in Hongfeng Reservoir varied across different hydrological periods, and the overall BC value of the phytoplankton community showed a decreasing trend. Compared to the dry season and rising period, GPP was higher during the wet season, reaching its maximum value. From the dry season to the wet season, phytoplankton GPP gradually increased, indicating that WLF positively influenced primary productivity by regulating phytoplankton community composition and stability as water levels transitioned from low (dry season) to high (wet season) (Fig. 6b). Specifically, GPP increased with rising water levels. Previous studies had demonstrated a coupling relationship between reservoir WLF and the content and types of organic matter in the water48. For example, water level decline has been identified as the primary driver of spatiotemporal variations in annual and seasonal primary productivity of vegetation in lake-floodplain ecosystems49. WLF significantly influenced nutrient dynamics and biological succession patterns, with high-amplitude groundwater level variations enhancing productivity in downstream areas44,50. Notably, phytoplankton primary productivity has surpassed submerged macrophyte productivity since 2000, becoming the dominant energy flow in the food web39.
Conclusions
In Hongfeng Reservoir, both interannual variations and hydrological periods considerably influenced water levels and physicochemical factors. Nutrient concentrations, phytoplankton species richness, and primary productivity increased from the dry season to the wet season as water levels rose, followed by a gradual decline during the falling period. Thus, regulating water levels in Hongfeng Reservoir represents a simple and effective approach to controlling nutrient concentrations and phytoplankton growth. Furthermore, during the high-water period in the wet season, phytoplankton community stability improved with increasing water levels within a specific range (1231.64–1238.64 m). As a critical environmental driver, WLF significantly regulate the phytoplankton community structure and its stability, providing a theoretical basis for controlling algal blooms through water level regulation. Therefore, WLF are a critical driver of the phytoplankton community structure and primary productivity in Hongfeng Reservoir. Moreover, the high water levels during the wet season indirectly enhance primary productivity by influencing community structure and stability. These findings provide a feasible solution for managing phytoplankton growth and community stability in reservoirs. This study will contribute to a deeper understanding of the stability and sustainability of aquatic ecosystems and provide a scientific basis for further research in related fields.
Data availability
All data generated or analysed during this study are included in this published article [and its supplementary information files].
References
Priya, A. Impact of climate change and anthropogenic activities on aquatic Ecosystem – a review. Environ. Res. 238, 117233 (2023).
Tilman, D. & Extinction Climate change and the ecology Ofhomo sapiens. J. Ecol. 110, 744–750 (2022).
Brasil, J., Attayde, J. L., Vasconcelos, F. R., Dantas, D. D. F. & Huszar, V. L. M. Drought-Induced Water-Level reduction favors cyanobacteria blooms in tropical shallow lakes. Hydrobiologia 770, 145–164 (2016).
Selmeczy, G. B. et al. Old sins have long shadows: climate change weakens efficiency of trophic coupling of Phyto- and zooplankton in a deep Oligo-Mesotrophic lowland lake (Stechlin, Germany)—a causality analysis. Hydrobiologia 831, 101–117 (2019).
Wu, Z. et al. Do the key factors determining phytoplankton growth change with water level in china’s largest freshwater lake? Ecol. Indic. 107, 105675 (2019).
Li, X., Ye, X., Yuan, C. & Xu, C. Can water release from local reservoirs Cope with the droughts of downstream lake in a large River-Lake system? J. Hydrol. 625, 130172 (2023).
Xiong, X. et al. Unsaturated slope stability around the three Gorges reservoir under various combinations of rainfall and water level fluctuation. Eng. Geol. 261, 105231 (2019).
Zhang, W., Pan, X., Liang, J., Zeng, J. & Song, C. Study on the hydrogeological structure of a karst subterranean river and seepage of a karst reservoir: A case study of the Yibasan reservoir in Yunnan Province, China. Water 16, 92 (2024).
Bao, Q. et al. Response of OC, TN, and TP deposition mediated by aquatic photosynthetic community structures in shallow karst surface waters under different land uses. Environ Res. 223, 115488 (2023).
Fadel, A., Atoui, A., Lemaire, B. J., Vinçon-Leite, B. & Slim, K. Environmental factors associated with phytoplankton succession in a mediterranean reservoir with a highly fluctuating water level. Environ. Monit. Assess. 187, 633 (2015).
Kann, J. & Walker, J. D. Detecting the effect of water level fluctuations on water quality impacting endangered fish in a Shallow, hypereutrophic lake using Long-Term monitoring data. Hydrobiologia 847, 1851–1872 (2020).
Su, H., Wang, R. & Feng, Y. Anaysis of water temperature flucyuations in lake Jamno. J. Ecol. 109, 3182–3194 (2021).
Li, Y. et al. Reservoir Regulation-Induced variations in water level impacts cyanobacterial bloom by the changing physiochemical conditions. Water Res. 259, 121836 (2024).
Lu, J., Bunn, S. E. & Burford, M. A. Effects of water level fluctuations on nitrogen dynamics in Littoral macrophytes. Limnol. Oceanogr. 63, 833–845 (2018).
Mäemets, H., Laugaste, R., Palmik, K. & Haldna, M. Response of primary producers to water level fluctuations of lake peipsi. P Est. Acad. Sci. 67, 231 (2018).
A, R. et al. Human-Induced water loss potentially threatens sustainable development of sandy regions in China. Commun. Earth Environ. 6, 68 (2025).
Liu, Q., Gan, L., Wu, H., Liang, L. & Yan, D. Water level fluctuations control wetland hydrological connectivity indriving the integrity of wetlands. J Hydrol. 657, 133095 (2025).
Liao, N., Zhang, L., Chen, M., Li, J. & Wang, H. The influence mechanism of water level operation on algal blooms in Canyon reservoirs and bloom prevention. Sci. Total Environ. 912, 169377 (2024).
Han, Q. et al. Assessing alterations of water level due to environmental water allocation at multiple Temporal scales and its impact on water quality in Baiyangdian Lake, China. Environ. Res. 212, 113366 (2022).
Bondar-Kunze, E., Tritthart, M. & Hein, T. The influence of short term water level fluctuations and desiccation stress on periphyton development at a riparian zone of a large regulated river. Fundam Appl. Limnol. 186, 283–296 (2015).
Wang, K. et al. Altered energy mobilization within the Littoral food web in new habitat created by Climate-Induced changes in lake water level. Front Ecol. Evol. 10, 886327 (2022).
Jia, J. et al. Phytoplankton community Composition, carbon Sequestration, and associated regulatory mechanisms in a floodplain lake system. Environ. Pollut. 306, 119411 (2022).
Kratzer, S., Hakansson, B. & Sahlin, C. Assessing Secchi and photic zone depth in the Baltic sea from satellite data. Ambio 32, 577–585 (2003).
Behrenfeld, M. J. & Falkowski, P. G. A consumer’s guide to phytoplankton primary productivity models. Limnol. Oceanogr. 42, 1479–1491 (1997).
Dai, Y. et al. Coastal phytoplankton blooms expand and intensify in the 21St century. Nature 615, 280–284 (2023).
Liu, Q. et al. Relationship between dissolved organic matter and phytoplankton community dynamics in a Human-Impacted subtropical river. J. Clean. Prod. 289, 125144 (2021).
Ye, L., Tan, L., Wu, X., Cai, Q. & Li, B. L. Nonlinear causal analysis reveals an effective water level regulation approach for phytoplankton blooms controlling in reservoirs. Sci. Total Environ. 806, 150948 (2022).
Hu, J. et al. Variation in resource allocation strategies and environmental driving factors for different Life-Forms of aquatic plants in cold temperate zones. J. Ecol. 109, 3046–3059 (2021).
Liu, J. et al. Water-Level fluctuations are key for phytoplankton taxonomic communities and functional groups in Poyang lake. Ecol. Indic. 104, 470–478 (2019).
Yu, H., Xu, S., Tian, W., Zhu, L. & Sun, Y. Impact of Long-Term water level fluctuation on the Distribution, Transport, and fate of phosphorus in reservoir sediment. Environ. Sci. Pollut Res. 26, 33146–33156 (2019).
Jian, Z. et al. Long-Term responses of riparian plants’ composition to water level fluctuation in china’s three Gorges reservoir. PLoS One. 13, e0207689 (2018).
Cruz-Ramírez, A. K., Salcedo, M. Á., Sánchez, A. J., Barba Macías, E. & Mendoza Palacios, J. D. Relationship among physicochemical Conditions, Chlorophyll-a Concentration, and water level in a tropical River–Floodplain system. Int. J. Environ. Sci. Technol. 16, 3869–3876 (2019).
Schrofner Brunner, B. et al. Macroalgae maintain growth outside their observed distributions: implications for Biodiversity-Ecosystem functioning at landscape scales. J. Ecol. 111, 1362–1373 (2023).
Wu, Q., Wang, F., Chen, Y., Zou, W. & Zhu, Z. Diazotrophic community in the sediments of Poyang lake in response to water level fluctuations. Front. Microbiol. 15, 1324313 (2024).
Wang, L., Tan, L. & Cai, Q. Distinct differences of vertical phytoplankton community structure in mainstream and a tributary Bay of the three Gorges Reservoir, China. Front Plant. Sci. 15, 1381798 (2024).
Qiu, X., Huang, T. & Zeng, M. Differences in phytoplankton dynamics and community structure between a wet year and dry year in the Zhoucun reservoir. J. Freshw. Ecol. 31, 377–391 (2016).
Lewis, A. S. L. et al. Reservoir drawdown highlights the emergent effects of water level change on reservoir Physics, Chemistry, and biology. J Geophys. Res. -Biogeosci. 129, e2023JG007780 (2024).
Porcel, S. et al. Structure of plankton and waterbird communities under water level fluctuations: two case studies in shallow lakes of the Patagonian steppe. Aquat. Sci. 86, 69 (2024).
Zhang, X., Tan, L., Cai, Q. & Ye, L. Environmental factors indirectly reduce phytoplankton community stability via functional diversity. Front Ecol. Evol. 10, 990835 (2022).
Allen, S. et al. Interannual stability of phytoplankton community composition in the North-East Atlantic. Mar. Ecol. Prog Ser. 655, 43–57 (2020).
Qian, K., Liu, X. & Chen, Y. Effects of water level fluctuation on phytoplankton succession in Poyang Lake, China – a five year study. Ecohydrol Hydrobiol. 16, 175–184 (2016).
Bonato Asato, A. E. et al. Seasonal shifts in plant diversity effects on Above-Ground–Below‐Ground phenological synchrony. J. Ecol. 108, 1649–1663 (2020).
Liu, X., Qian, K. & Chen, Y. Effects of water level fluctuations on phytoplankton in a Changjiang river floodplain lake (Poyang lake): implications for dam operations. J. Gt Lakes Res. 41, 770–779 (2015).
Onandia, G. et al. Biogeochemistry of natural ponds in agricultural landscape: lessons learned from modeling a kettle hole in Northeast Germany. Sci. Total Environ. 634, 1615–1630 (2018).
Toporowska, M., Rechulicz, J., Adamczuk, M. & Niedźwiecki, M. The role of abiotic and biotic environmental factors in shaping epiphyton on common Reed in Shallow, hydrologically Transformed, temperate lakes. Knowl Manag Aquat. Ecosyst. 419, 18 (2018).
Guimarais-Bermejo, M. O., Merino-Ibarra, M., Valdespino-Castillo, P. M., Castillo-Sandoval, F. S. & Ramírez-Zierold, J. A. Metabolism in a deep hypertrophic aquatic ecosystem with high Water-Level fluctuations: A decade of records confirms sustained net heterotrophy. PeerJ 6, e5205 (2018).
Li, X., Cui, B., Yang, Q. & Lan, Y. Impacts of water level fluctuations on detritus accumulation in lake Baiyangdian, China. Ecohydrology 9, 52–67 (2016).
Peng, J., Hu, Z., Feng, D. & Wang, Q. Variations of organic matter content and type within the sequence stratigraphic framework of the lacustrine Deep-Water Dongyuemiao Formation, Sichuan Basin, Western China. Mar. Pet. Geol. 149, 106104 (2023).
Ye, X., Meng, Y., Xu, L. & Xu, C. Net primary productivity dynamics and associated hydrological driving factors in the floodplain wetland of china’s largest freshwater lake. Sci. Total Environ. 659, 302–313 (2019).
Collins, S. V. et al. Regional response of the coastal aquifer to hurricane Ingrid and sedimentation flux in the Yax Chen cave system (Ox Bel Ha) Yucatan, Mexico. Palaeogeogr Palaeoclimatol Palaeoecol. 438, 226–238 (2015).
Funding
This work was supported by the National Natural Science Foundation of China (No. 42463007), the Guizhou Provincial Science and Technology Program (RC [2020] 6009-2, YWZ [2023]006, FQ [2023]010).
Author information
Authors and Affiliations
Contributions
Yugui Peng: Conceptualization, Methodology, Investigation, Writing-Original Draft. Jing Xiao: Investigation, Formal analysis, Data Curation. Qian Chen:Data Curation. Jiangzhi Luo: Data Curation. Taibo Gu: Investigation, Data Curation.Xiaochuan Song: Investigation, Data Curation. Bo Yang: Investigation, Data Curation. Mengshu Han: Data Curation. Si Zhou: Data Curation. Yulin Li: Data Curation. Qiuhua Li: Software, Investigation, Data Curation, Writing-Original Draf Supervision. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Peng, Y., Li, Q., Xiao, J. et al. Hydrology drives phytoplankton succession and productivity coupling in plateau hydropower reservoirs with implications for ecosystem health management. Sci Rep 15, 42085 (2025). https://doi.org/10.1038/s41598-025-25966-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-25966-6
