
Abstract
This study aims to explore the relationship between Helicobacter pylori (H. pylori) infection, particularly CagA-positive strains, and Clonorchiasis. A total of 309 civil servants who underwent health check-ups and 73 hospitalized patients with Clonorchiasis from May 2019 to April 2023 were included. A control group of 100 healthy individuals matched by age, gender, and residence was included. H. pylori and Clonorchiasis antibodies in civil servants were detected using ELISA. Western blotting was used to identify H. pylori strains in hospitalized patients. Among civil servants, H. pylori and Clonorchiasis antibody positivity rates were 43.7% and 23.9%, respectively. H. pylori antibody positivity was comparable between those with (45.9%) and without Clonorchiasis (43.0%) infection. H. pylori infection among Clonorchiasis patients (61.6%) was slightly higher than in healthy controls (44.0%). Multifactorial analysis identified consuming raw fish (61.145; 22.263–167.93; 0.000) and CagA + H. pylori infection (3.7; 1.233–11.103; 0.020) as independent risk factors for Clonorchiasis. The rate of CagA + H. pylori infection is significantly higher in patients with Clonorchiasis than in healthy individuals. CagA + H. pylori infection and a history of consuming raw fish are independent risk factors for current Clonorchiasis.
Similar content being viewed by others
Introduction
Clonorchiasis is a zoonotic disease characterized primarily by damage to the hepatobiliary system, caused by adult Clonorchis sinensis (C. sinensis) parasitizing the bile ducts. Taking C. sinensis as an example, this strain has been endemic in China for over 2000 years, with more than 15 million people in countries such as China, Japan, North Korea, and Vietnam currently at risk of infection1,2. The most severe complication of Clonorchiasis infection is cholangiocarcinoma, which the International Agency for Research on Cancer (IARC) of the World Health Organization (WHO) has classified as a Group I carcinogen for cholangiocarcinoma. Due to its complex life cycle and wide range of definitive hosts, it poses a significant public health burden3. Differences in dietary habits among humans determine the heterogeneity of Clonorchiasis infection within populations, while climatic conditions, sanitation, economic status, and educational level are closely related to the prevalence and reinfection rates. Moreover, patients with high infection intensity before treatment are particularly prone to reinfection4,5,6,7.
Different species of Clonorchiasis do not overlap in geographic distribution and are located in different climatic zones, but their life cycles and pathogenicity are similar8. After Clonorchiasis infects the human body, it induces a complex gene-protein-metabolism regulatory network in the host’s liver, causing a series of immune pathological damages to the biliary tract through mechanical injury and the direct or indirect effects of excretory-secretory products (ESPs)9. ESPs mediate intercellular communication between biliary epithelial cells and hepatic stellate cells, inducing epithelial-mesenchymal transition (EMT) and biliary fibrosis by affecting cytokine secretion and protein expression levels, which promotes malignant transformation of cells in the biliary microenvironment and late-stage hepatobiliary abnormalities10. The role of host immune cells varies during different stages of infection, with CD4 + T cell subsets playing a dominant role11. Liver damage characterized by collagen deposition and inflammatory cell infiltration can occur within 24 h after Clonorchiasis infection12, with early-stage responses primarily driven by macrophage-induced innate immunity and Th1-mediated cellular immunity. As the duration of infection increases, the levels of inflammatory cytokines (especially IL-6 and IL-1β) decrease, while anti-inflammatory cytokines, particularly IL-4, increase, indicating that a Th2-type immune response is triggered during the adult stage of Clonorchiasis to control excessive inflammation and promote tissue repair13. Additionally, the imbalance between regulatory T cells (Treg) and Th17 cells can also facilitate the fibrotic process14. IL-10 plays a crucial role throughout the infection process by inhibiting Th1 cell secretion of IFN-γ and IL-2, inducing Th2 cells to secrete IL-4 and IL-5, and protecting the host liver from damage by suppressing Th17 cell responses15. Therefore, the immune pathological damage of Clonorchiasis is the result of the combined actions of multiple cytokines.
Helicobacter pylori (H. pylori), classified as a Group I carcinogen, is a Gram-negative spiral bacterium first discovered and cultured in gastric biopsy samples by Robin Warren and Barry Marshall16. It utilizes adhesins (such as blood group antigen-binding adhesin, BabA; sialic acid-binding adhesin, SabA), enzymes (such as urease, Ure), toxins (such as vacuolating cytotoxin gene, VacA), and effector proteins (such as cytotoxin-associated gene A, CagA) to participate in colonization, immune evasion, and disease induction17. After entering the stomach, H. pylori releases virulence factors CagA and VacA, interacting with the host’s gastric microenvironment and altering host cell signaling pathways, which induce, control, and regulate inflammatory responses in the gastric mucosa, maintaining chronic inflammation via epigenetic pathways in gastric epithelial cells18. These virulence factors primarily promote disease development by inducing changes in host inflammatory cytokines such as interleukin (ILs), tumor necrosis factor-alpha (TNF-α), and gamma interferon (IFN-γ)19. Research has confirmed a synergistic pathogenic effect between H. pylori and pathogens such as EBV, HBV, HCV, Brucella, and Orientia tsutsugamushi, indicating that co-infection may influence disease progression20,21,22,23,24. Additionally, H. pylori plays a protective role in asthma and inflammatory bowel disease (IBD) by balancing the homeostasis of pro-inflammatory and anti-inflammatory microbes, with this immunomodulatory effect being more pronounced in CagA-positive H. pylori strains25,26.
The relationship between parasitic infections and H. pylori has been a longstanding topic of discussion. Some studies have found that parasitic infections can improve the clinical outcomes of H. pylori-related diseases. It is reported that the potential impact of co-infection with H. pylori and helminths on the immune response to Mycobacterium tuberculosis, inducing a Th2-type immune response to H. pylori27. In sub-Saharan Africa, a “dichotomy” is observed, with a 2/3 H. pylori infection rate and an extremely low incidence of gastric adenocarcinoma (2–3%). Researchers agree that this may be related to the high burden of soil-transmitted helminth (STH) infections in the area, where gastrointestinal helminths reduce the risk of H. pylori-induced gastric adenocarcinoma by inducing immune tolerance28. However, other studies also found inconsistent results, suggesting that co-infection of intestinal parasites and H. pylori may not always be protective. In certain contexts, H. pylori infection has been proven to relate to the increased risk of hepatobiliary abnormalities and cholangiocarcinoma, and co-infection with helminths may exacerbate tissue injury through chronic inflammation 29,30. These inconsistent findings underscore the complex mechanisms underlying the impact of H. pylori co-infection on clinical outcomes. Both Clonorchiasis and H. pylori mediate immune responses through the balance of CD4 + T cell subsets, leading us to hypothesize that there may be some cross-influences between these biological behaviors. Therefore, we designed this cross-sectional study to explore the synergistic effects of H. pylori infection on current Clonorchiasis infection.
Methods
Study design and participants
We conducted a cross-sectional study ranging from May 2019 to April 2023 in two hospitals, including the Health Examination Center of Affiliated Hospital of Youjiang Medical University for Nationalities and the Infectious Disease Department of Beihai People’s Hospital. A total of 482 participants were enrolled in this study, including (1) 309 civil servants undergoing routine health check-ups, (2) 73 hospitalized patients with current C. sinensis infection, and this group was further divided into H. pylori-positive (n = 45) and H. pylori-negative (n = 28), and (3) 100 healthy individuals.
The inclusion criteria were as follows: (1) individuals aged 18–65 years; (2) confirmed H. pylori or C. sinensis infection status by serological or parasitological methods; (3) written informed consent provided. The exclusion criteria were as follows: (1) history of liver or biliary tract diseases unrelated to C. sinensis; (2) immunodeficiency or use of immunosuppressive drugs; (3) incomplete clinical or laboratory data; (4) Patients who cannot cooperate.
General information
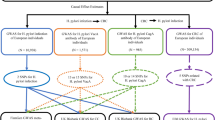
A total of 309 healthy individuals were recruited from the Health Examination Center of Affiliated Hospital of Youjiang Medical University for Nationalities, including 275 males and 34 females, with an average age of 44.34 ± 8.01 years. Seventy-three hospitalized patients with current Clonorchiasis were recruited from the Infectious Disease Department of Beihai People’s Hospital, including 57 males and 16 females, with an average age of 42.15 ± 9.44 years. The control group comprised 100 healthy individuals matched for gender and age from Beihai People’s Hospital during the same period, including 73 males and 27 females, with an average age of 43.29 ± 8.97 years. Detailed grouping of the study population and the technical approach is shown in Fig. 1. No significant difference was found in gender (P = 0.978) and age (P = 0.553).

Detailed grouping of the study population and the technical approach.
Diagnosis of Clonorchiasis was based on the standards of the former Ministry of Health of the People’s Republic of China WS309—2009, using a modified Kato thick-smear technique to detect C. sinensis eggs in the feces and duodenal fluid of the subjects as the basis for diagnosis. Enzyme-linked immunosorbent assay (ELISA) was used to detect C. sinensis antibodies in the patients’ blood, supplementing the pathogenic diagnosis31, along with epidemiological history and clinical manifestations for confirmation.
Based on the presence of clinical symptoms, the collected cases of Clonorchiasis infection were divided into two categories: (1) current infection, defined by the presence of C. sinensis eggs in feces and/or typical clinical symptoms, with or without positive C. sinensis antibodies; (2) past infection, defined by positive C. sinensis antibodies alone, with no egg detection in feces and no clinical manifestations. one being current infection, characterized by positive C. sinensis antibodies, detection of eggs in feces, and/or characteristic clinical symptoms; the other being past infection, indicated solely by positive C. sinensis antibodies, negative fecal egg detection, and/or absence of clinical manifestations. Exclusion criteria included individuals with other liver and biliary diseases, immunodeficiency, or those undergoing related treatments. All case data collection was approved by the ethics review committees of the two hospitals. All patients were informed of the risks and benefits of participating in the study before it began, and they voluntarily participated in the research and signed informed consent forms. Based on H. pylori detection results, the 73 patients with current Clonorchiasis infection were divided into H. pylori positive (45 cases) and negative (28 cases), with a comparison of general information between the two groups detailed in Table 1.
Methods
Peripheral blood samples were collected from 309 healthy individuals, and ELISA was used to simultaneously detect overall infection rates of H. pylori antibodies and C. sinensis antibodies in the general population. Based on the positivity of C. sinensis antibodies, the H. pylori infection status of the subjects was analyzed. In addition, the 100 healthy individuals recruited from Beihai People’s Hospital were matched controls used specifically for comparative analysis with 73 hospitalized clonorchiasis patients. Peripheral blood samples were collected from 73 hospitalized patients with current C. sinensis infection and 100 healthy individuals, and H. pylori antibodies were detected using the Western blot method (products from Shenzhen Blot Biological Company). The reagents were purchased from Shenzhen Blot Biological Company (batch number: 161216), and the experimental procedures were strictly followed according to the instructions. The Western blot method can detect antibodies against H. pylori cytotoxin CagA (bands at 128 kD and 116 kD), vacuolating cytotoxin-associated protein VacA (bands at 95 kD and 91 kD), and urease ureB (66 kD) and urease ureA (30 kD). Diagnosis criteria: (1) A positive result for either urease A or urease B, or both, indicates H. pylori strain infection; (2) If condition (1) is met, and CagA and/or VacA are positive, it indicates infection with CagA + strains. Declaration: All experimental methods in this study were strictly carried out in accordance with relevant guidelines and regulations, to ensure the scientific nature and ethical compliance of the research.
Statistical methods
A database was established using SPSS 27.0 statistical software. Continuous variables were expressed as mean ± standard deviation (x ± s), and the categorical variables were presented as frequency and percentage (n, %). The comparison of H. pylori infection status between the two groups was conducted using the chi-square test. Univariate analysis of risk factors for current C. sinensis infection was performed using the chi-square test, and variables with statistically significant factors were included in the logistic regression model for multivariate analysis, with a significance level of α = 0.05, using two-sided tests.
Results
Survey of H. pylori and C. sinensis infection status among civil servants
A total of 482 participants were included in this study, the baseline characteristics of C. sinensis infection patients stratified by H. pylori infection are listed in Table 1. By the ELISA method, antibodies against H. pylori and C. sinensis were tested in 309 civil servants. The results showed that the H. pylori antibody positivity rate was 43.7% (135/309) (95% CI: 38.13–49.25%), and the C. sinensis antibody positivity rate was 23.9% (74/309) (95% CI: 19.16–28.73%). Further analysis of H. pylori infection status between C. sinensis-infected and non-infected individuals revealed that the H. pylori antibody positivity rate among C. sinensis-infected individuals was 45.9% (34/74), which was similar to the 43.0% (101/235) in non-infected individuals, with no statistical difference (P = 0.654). A further analysis of H. pylori infection status in individuals with current infection, past infection, the results showed that the H. pylori antibody positivity rate was 57.1% in the current C. sinensis infection group, and 43.0% in the healthy control group. Although a slightly higher positive percentage of current infection than the healthy control group, the difference was not statistically significant (P = 0.299). In contrast, patients with past C. sinensis infection showed no significant increase in H. pylori antibody positivity compared with non-infected individuals. See Fig. 2 for details.

Survey of H. pylori and C. sinensis infection status among civil servants using ELISA.
Infection status of different H. pylori strains in patients with current c.sinensis infection
The Western blot method was used to detect H. pylori antibodies in 73 patients with current C. sinensis infection and 100 healthy controls, followed by strain identification. The results showed that the H. pylori infection rate in patients with current C. sinensis infection was 61.6%, slightly higher than the rate of 44.0% in healthy individuals, but this difference was not statistically significant (P = 0.249). However, the infection rate of CagA + strains in patients with current C. sinensis infection was 56.2%, significantly higher than the 28.0% in healthy individuals (χ2 = 13.960, P < 0.001, indicating a significant difference. The infection rate of CagA strain in healthy infected individuals was 16.0%, significantly higher than that in current C. sinensis infected individuals (P = 0.033). See Fig. 3 for details.

Infection status of different H. pylori strains in patients with current C.sinensis infection using the Western blot method.
Multivariate regression analysis of the promoting effect of CagA + H. pylori infection on current C. sinensis infection
Factors such as patient gender, residence, history of consuming raw fish, and H. pylori CagA status (CagA⁺/CagA⁻) were incorporated into the multivariate regression model, and the results indicated that a history of consuming raw fish (61.145; 22.263–167.93; 0.000) and CagA + H. pylori infection (3.7; 1.233–11.103; 0.020) were independent risk factors for current C. sinensis infection. See Fig. 4.

Multivariate regression analysis of the relationship between CagA + H. pylori infection and current C. sinensis infection.
Discussion
Currently, over 35 million people globally are affected by C. sinensis infection, but only about 1.5–2 million exhibit symptoms or complications. Among individuals with current infection, some patients present acute symptoms such as fever, jaundice, and liver function abnormalities, while others show more subtle symptoms, only testing positive for eggs in fecal examinations32. The clinical phenotypic differences of C. sinensis disease are the result of the combined effects of the host’s immune status, genetic background, characteristics of the infecting strain, infection load, and the host’s age, gender, nutritional status, and environmental factors. H. pylori infection primarily occurs in childhood, with its prevalence increasing with age, while Clonorchiasis predominantly affects adults. Poor dietary habits and unfavorable socioeconomic conditions are common risk factors for both H. pylori and Clonorchiasis infection. Additionally, both Clonorchiasis and H. pylori mediate immune responses through the balance of CD4 + T cell subsets, leading us to hypothesize potential cross-interactions between these two microorganisms. The classification of infections, particularly past infections, involves a combination of approaches. For instance, serological testing is commonly used, where IgM antibodies indicate recent infection and IgG antibodies reflect past exposure due to their prolonged presence. Moreover, advances in molecular diagnostics, including metagenomic sequencing, facilitate direct detection of pathogen nucleic acids, enhancing the identification of current or prior infections, especially in immunocompromised individuals. The integration of immunological evidence, clinical coding, symptom timing, and molecular methods thus provides a comprehensive framework for accurately categorizing infections in both clinical and research settings.
The clinical phenotypic differences of C. sinensis disease are the result of the combined effects of the host’s immune status, genetic background, characteristics of the infecting strain, infection load, and the host’s age, gender, nutritional status, and environmental factors. H. pylori infection primarily occurs in childhood, with its prevalence increasing with age, while Clonorchiasis predominantly affects adults. Poor dietary habits and unfavorable socioeconomic conditions are common risk factors for both H. pylori and Clonorchiasis infection. Additionally, both Clonorchiasis and H. pylori mediate immune responses through the balance of CD4 + T cell subsets, leading us to hypothesize potential cross-interactions between these two microorganisms.
In this study, we first employed the ELISA method to detect both H. pylori antibodies and C. sinensis antibodies in 309 civil servants undergoing health check-ups. The results indicated that the infection rate of C. sinensis among healthy individuals in this region was as high as 23.9%, likely related to local dietary habits that favor raw freshwater fish consumption. Subsequent multivariable regression analysis further confirmed that a history of consuming raw fish is one of the independent risk factors for C. sinensis infection. Our study also found that the H. pylori infection rates among individuals with and without C. sinensis infection was 45.9% and 43.0%, respectively, with no statistically significant difference. However, the H. pylori infection rate among individuals with current C. sinensis infection reached 61.5%. Although this did not show statistical significance, it suggests that H. pylori may facilitate the invasion and pathogenicity of C. sinensis in the human body through certain mechanisms, such as raw fish consumption, poor hygiene. The antibody results might reflect prior exposure or involve serological cross-reactivity rather than active synergy. Additionally, selecting hospitalized patients may introduce bias toward more severe cases. Finally, the cross-sectional design limits causal inference. Future longitudinal and mechanistic studies are needed to clarify these relationships.
H. pylori infection often occurs in childhood and, if not eradicated, can persist for a lifetime. We can’t help but wonder whether the colonization of H. pylori during childhood prepares the ground for the invasion of Clonorchiasis. H. pylori downregulates inflammation and controls the host’s immune response through various virulence factors33. Its induced chronic active inflammation reduces the gastrointestinal mucosal barrier function, creating a suitable environment for the colonization and development of Clonorchiasis. Theoretically, if an individual has a prior H. pylori infection, especially a CagA-positive infection, this may lead to a certain degree of immunosuppression or immune tolerance. When such an individual subsequently gets infected with Clonorchiasis, the immune microenvironment changes caused by the previous H. pylori infection may facilitate the persistence of Clonorchiasis and disease progression. Furthermore, H. pylori infection may also indirectly affect the host’s susceptibility and response to Clonorchiasis by influencing the composition of the gastrointestinal microbiota. The gut microbiota is closely related to the development and function of the host immune system34, and the dysbiosis caused by H. pylori infection may further impact the host’s defense against other pathogens. Therefore, we speculate that H. pylori infection could be a factor influencing the clinical heterogeneity of symptoms after Clonorchiasis infection.
To clarify the interaction between the two, we selected 73 hospitalized patients with current C. sinensis infection from another hospital and used the Western blot method to detect H. pylori antibodies. Strain identification was performed based on the presence of CagA and/or VacA regions. The results showed that the H. pylori infection rate among patients with current C. sinensis infection was 61.6%, slightly higher than the rate of 44.0% in healthy individuals, although this difference was not statistically significant. However, the infection rate of CagA + strains in patients with current C. sinensis infection was 56.2%, significantly higher than the 28.0% in healthy individuals, indicating a significant association between CagA + H. pylori strains and current C. sinensis infection.
In the hamster model established by Dangtakot et al., the colonization rate of H. pylori in the hepatobiliary system in the co-infection group with H. pylori and C. sinensis was higher than that in the single infection group, and it increased the severity of hepatobiliary abnormalities, manifested as more severe periductal fibrosis, cholangitis, and biliary hyperplasia35. This may result from one or more of the following reasons: 1. The local inflammation and tissue damage in the hepatobiliary system caused by Clonorchiasis provide a more favorable environment for H. pylori colonization36; 2. The urease produced by H. pylori neutralizes gastric acid, creating a more suitable weakly acidic environment for its survival37. After Clonorchiasis infection, physiological changes such as bile stasis or alterations in bile duct pH facilitate the survival and proliferation of H. pylori. Additionally, bile acids can enhance the translocation of H. pylori’s CagA protein into host cells, amplifying the effects of CagA to promote inflammation and damage38; 3. Co-infection by both pathogens interferes with the host’s immune response39, weakening the ability to clear H. pylori; 4. Researchers speculate that H. pylori may exhibit a “ride-along” phenomenon in the lifecycle of Clonorchiasis, where L-fucose mediates H. pylori colonization in the Clonorchis intestine40, followed by the transmission of H. pylori carried by Clonorchiasis larvae in the hepatobiliary system41,42; 5. H. pylori may first cause damage in the stomach and then infect the biliary system via ascending duodenal infection or through the portal venous circulation 43; 6. Some virulence factors of H. pylori strains (CagA and VacA) may enhance their colonization ability in the hepatobiliary system44. Of course, the interactions between the two pathogens are not unidirectional. The inflammatory response induced by H. pylori infection may lead to changes in the biliary and intestinal mucosa, providing a suitable environment for the colonization and development of Clonorchis, while the Treg cells induced by the infection suppress the inflammatory response45, thereby reducing the host’s defense against Clonorchis infection.
We conducted a multifactorial regression analysis on the factors that may influence current C. sinensis infection. The results suggest that CagA-positive H. pylori infection is one of the independent risk factors for current C. sinensis disease. Virulence factors like CagA enable H. pylori to activate the production of inflammatory cytokines and chemokines, leading to leukocyte recruitment. Suyapoh. W et al. observed that in patients infected with both CagA-positive H. pylori and C. sinensis, there was a significant increase in leukocyte infiltration and lymphocyte aggregation, indicating a synergistic effect of these two carcinogens in inducing severe inflammatory responses, goblet cell metaplasia, and abnormal proliferation39. Notably, the polymorphism of the EPIYA sequence in the CagA protein is closely related to the severity of disease in patients with Clonorchiasis infection, with the EPIYA-AB’C type (containing the variant EPIYT B) leading to more severe periductal fibrosis, while CagA types containing multimerization (CagA multimerization, CM) sequences were significantly associated with the degree of fibrosis42. Other studies have shown that CagA also induces persistent advanced periductal fibrosis (APF) after praziquantel treatment, even leading to recurrence, affecting patient prognosis46. These findings emphasize the importance of monitoring the CagA protein in the management and treatment of biliary diseases in cases of co-infection with Clonorchiasis and H. pylori.
This study has some limitations. As a cross-sectional design, it cannot establish causality or the sequence of infections. The lack of mechanistic data, such as cytokine profiles and functional assays, limits our understanding of the underlying biological interactions. In addition, potential confounders such as socioeconomic status, comorbidities, and hygiene practices were not fully accounted for and may have influenced the results. Antibody-based testing may also reflect prior rather than active infection. These factors should be addressed in future longitudinal and mechanistic studies.
In summary, the infection rate of CagA-positive H. pylori in patients with current C. sinensis infection is significantly higher than that in healthy individuals, and CagA-positive H. pylori strains are independent risk factors for current C. sinensis infection, suggesting that H. pylori carrying the CagA gene may have a synergistic effect on the occurrence and development of Clonorchiasis.
Most current studies primarily focus on the relationship between H. pylori and Opisthorchis viverrini co-infections, likely due to the distribution characteristics of Clonorchiasis in their respective locations. Our study provides supplementary insights into the synergistic pathogenic effects of C. sinensis and H. pylori. Although we have observed a potential association between CagA-positive H. pylori and current C. sinensis infection, possibly promoting more severe pathological damage and even the development of cholangiocarcinoma, there are still some research limitations. There is a lack of in-depth understanding of the molecular mechanisms by which CagA-positive H. pylori and C. sinensis co-infections synergistically affect host cells. More research is needed to clarify how these pathogens jointly influence signaling pathways and gene expression in biliary epithelial cells. The host immune response to H. pylori and C. sinensis infections is very complex and exhibits individual variability. The diversity of the host immune background may not have been adequately considered in studies, potentially affecting a comprehensive understanding of pathogen virulence. Furthermore, long-term follow-up studies on co-infected patients are crucial for understanding their relationship with the progression of C. sinensis and assessing the long-term effects of potential therapeutic interventions.
Data availability
Some of the data in this study are cited from publicly available literature, and the specific sources have been indicated in the text. Other datasets generated and analyzed during the research process are not publicly available due to containing patient privacy information. They can be obtained from the corresponding author upon reasonable request.
References
McManus, D. P. Recent progress in the development of liver fluke and blood fluke vaccines. Vaccines (Basel) 8(3), 553 (2020).
Zhu, X. P. & Su, C. Human parasitology (People’s Medical Publishing House, 2018).
Qian, M. B., Keiser, J., Utzinger, J. & Zhou, X. N. Clonorchiasis and opisthorchiasis: Epidemiology, transmission, clinical features, morbidity, diagnosis, treatment, and control. Clin. Microbiol. Rev. 37(1), e0000923 (2024).
Li, Z. et al. Clonorchis sinensis reinfection rate and reinfection determinants: A prospective cohort study in Hengxian County, Guangxi, China. J. Infect. Dis. 225(3), 481–491 (2022).
Qian, M. B. & Zhou, X. N. Children education for sustainable control of liver fluke infections. Infect. Dis. Poverty 11(1), 119 (2022).
Tidman, R., Kanankege, K. S. T., Bangert, M. & Abela-Ridder, B. Global prevalence of 4 neglected foodborne trematodes targeted for control by WHO: A scoping review to highlight the gaps. PLoS Negl. Trop. Dis. 17(3), e0011073 (2023).
Zheng, J. X. et al. Natural variables separate the endemic areas of Clonorchis sinensis and Opisthorchis viverrini along a continuous, straight zone in Southeast Asia. Infect. Dis. Poverty 13(1), 24 (2024).
Pakharukova, M. Y., Zaparina, O., Hong, S.-J., Sripa, B. & Mordvinov, V. A. A comparative study of Helicobacter pylori infection in hamsters experimentally infected with liver flukes Opisthorchis felineus, Opisthorchis viverrini, or Clonorchis sinensis. Sci. Rep. 11(1), 7789 (2021).
Zhao, L. & Peng, X. H. Advances in the immunologic mechanism of hepatobiliary fibrosis induced by Clonorchis sinensis. J. Parasit. Biol. 18(05), 609–613 (2023).
Yi, J., Jeong, J. H., Won, J., Chung, S. & Pak, J. H. The crosstalk between cholangiocytes and hepatic stellate cells promotes the progression of epithelial-mesenchymal transition and periductal fibrosis during Clonorchis sinensis infection. Parasit. Vectors 17(1), 151 (2024).
Zhang, B. B. et al. Increased hepatic Th2 and Treg subsets are associated with biliary fibrosis in different strains of mice caused by Clonorchis sinensis. PLoS ONE 12(2), e0171005 (2017).
Wu, Y. et al. Multilayer omics reveals the molecular mechanism of early infection of Clonorchis sinensis juvenile. Parasit. Vectors 16(1), 285 (2023).
Zhan, T. et al. Multi-omics approaches reveal the molecular mechanisms underlying the interaction between Clonorchis sinensis and mouse liver. Front. Cell Infect. Microbiol. 13, 1286977 (2023).
Yan, C. et al. The dynamics of Treg/Th17 and the imbalance of Treg/Th17 in Clonorchis sinensis-infected mice. PLoS ONE 10(11), e0143217 (2015).
Zhang, B. et al. IL-10 regulates Th17 response to inhibit hepatobiliary injury caused by Clonorchis sinensis infection in C57BL/6J mice. Front. Cell Infect. Microbiol. 12, 994838 (2022).
Marshall, B. J. & Warren, J. R. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 1(8390), 1311–1315 (1984).
Mohammadzadeh, R., Menbari, S., Pishdadian, A. & Farsiani, H. Helicobacter pylori virulence factors: Subversion of host immune system and development of various clinical outcomes. Expert Rev. Mol. Med. 25, e23 (2023).
Baj, J. et al. Helicobacter pylori virulence factors-mechanisms of bacterial pathogenicity in the gastric microenvironment. Cells 10(1), 27 (2020).
Luo, C. T., Sun, L. M. & Cen, Z. Research status of the relationship between different virulent Helicobacter Pylori infection and inflammatory factors. Med. Innov. China 21(08), 165–169 (2024).
Giammanco, A. et al. Helicobacter pylori and epstein-barr virus co-infection in gastric disease: What is the correlation with p53 mutation, genes methylation and microsatellite instability in a cohort of sicilian population?. Int. J. Mol. Sci. 24(9), 8104 (2023).
Wang, J. et al. Helicobacter pylori infection may increase the risk of progression of chronic hepatitis B disease among the Chinese population: A meta-analysis. Int. J. Infect. Dis. 50, 30–37 (2016).
Attallah, A. M. et al. Prevalence of Helicobacter pylori infection in patients with chronic hepatitis C. J. Genet. Eng. Biotechnol. 20(1), 13 (2022).
Afrasiabian, S., Mohsenpour, B., Ghaderi, E., Nadri, S. & Hajibagheri, K. Does Helicobacter pylori infection play a role in susceptibility to brucellosis?. Jpn. J. Infect. Dis. 70(6), 672–674 (2017).
Du, Q. et al. CagA-positive Helicobacter pylori may promote and aggravate scrub typhus. Front. Microbiol. 15, 1351784 (2024).
Durazzo, M., Adriani, A., Fagoonee, S., Saracco, G. M. & Pellicano, R. Helicobacter pylori and respiratory diseases: 2021 update. Microorganisms 9(10), 2033 (2021).
Bretto, E. et al. Helicobacter pylori in inflammatory bowel diseases: Active protagonist or innocent bystander?. Antibiotics (Basel) 13(3), 267 (2024).
Bustamante-Rengifo, J. A., Astudillo-Hernández, M. & Del Pilar, C.-O. Effect of Helicobacter pylori and helminth coinfection on the immune response to Mycobacterium tuberculosis. Curr. Microbiol. 78(9), 3351–3371 (2021).
Setshedi, M. & Watermeyer, G. The impact of Helicobacter pylori and intestinal helminth infections on gastric adenocarcinoma and inflammatory bowel disease in Sub-Saharan Africa. Front. Med. (Lausanne) 9, 1013779 (2022).
Rao, B. et al. Dysbiosis in the human microbiome of cholangiocarcinoma. Front. Physiol. 12, 715536 (2021).
Dangtakot, R. et al. CagA(+) Helicobacter pylori infection and N-nitrosodimethylamine administration induce cholangiocarcinoma development in hamsters. Helicobacter 26(4), e12817 (2021).
Sun, X. Z. et al. Overview of diagnostic methods for Clonorchis sinensis. Jilin Anim. Husb. Vet. Med. 43(09), 117–118 (2022).
Liu, K. et al. Spatio-temporal disparities of Clonorchis sinensis infection in animal hosts in China: A systematic review and meta-analysis. Infect. Dis. Poverty 12(1), 97 (2023).
Kusters, J. G., van Vliet, A. H. & Kuipers, E. J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 19(3), 449–490 (2006).
Zheng, D., Liwinski, T. & Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 30(6), 492–506 (2020).
Dangtakot, R. et al. Coinfection with Helicobacter pylori and Opisthorchis viverrini Enhances the Severity of Hepatobiliary Abnormalities in Hamsters. Infect. Immun. 85 (4), e00009-17. https://doi.org/10.1128/iai.00009-17 (2017).
Avilés-Jiménez, F. et al. Microbiota studies in the bile duct strongly suggest a role for Helicobacter pylori in extrahepatic cholangiocarcinoma. Clin. Microbiol. Infect. 22(2), 178-e11 (2016).
Mu, T. et al. Helicobacter pylori intragastric colonization and migration: Endoscopic manifestations and potential mechanisms. World J. Gastroenterol. 29(30), 4616–4627 (2023).
Noto, J. M. et al. Iron deficiency linked to altered bile acid metabolism promotes Helicobacter pylori-induced inflammation-driven gastric carcinogenesis. J. Clin. Invest. 132(10), e147822 (2022).
Suyapoh, W. et al. Synergistic effects of cagA+ Helicobacter pylori co-infected with Opisthorchis viverrini on hepatobiliary pathology in hamsters. Acta Trop. 213, 105740 (2021).
Thanaphongdecha, P. et al. Adherence of Helicobacter pylori to Opisthorchis viverrini gut epithelium and the tegument mediated via L-fucose binding adhesin. Parasitology 149(10), 1374–1379 (2022).
Itthitaetrakool, U. et al. Chronic Opisthorchis viverrini infection changes the liver microbiome and promotes Helicobacter growth. PLoS ONE 11(11), e0165798 (2016).
Deenonpoe, R. et al. Elevated prevalence of Helicobacter species and virulence factors in opisthorchiasis and associated hepatobiliary disease. Sci. Rep. 7, 42744 (2017).
Cen, L. et al. Helicobacter pylori infection of the gallbladder and the risk of chronic cholecystitis and cholelithiasis: A systematic review and meta-analysis. Helicobacter 23(1), e12457 (2018).
Hatta, M. N. A., Mohamad Hanif, E. A., Chin, S. F. & Neoh, H. M. Pathogens and carcinogenesis: A review. Biology (Basel) 10(6), 533 (2021).
Schroeter, C. B. et al. Crosstalk of microorganisms and immune responses in autoimmune neuroinflammation: A focus on regulatory T cells. Front. Immunol. 12, 747143 (2021).
Phung, H. T. T. et al. Persistent advanced periductal fibrosis is associated with cagA-positive Helicobacter pylori infection in post-praziquantel treatment of opisthorchiasis. Helicobacter 27(4), e12897 (2022).
Acknowledgements
Not applicable.
Funding
This work was supported by National Key Research and Development Program of China (No.2023YFC2605400) and Key R&D Program Project in Guangxi (No.2023AB04032).
Author information
Authors and Affiliations
Contributions
C.L., Z.L. and J.C. jointly conceived the research plan. F.D. collected the basic data from the clinical frontline. Y.L. assisted in integrating resources and participated in data organization. Z.L. provided historical medical record data. Q.D. carried out preliminary data analysis. J.P. assisted in interpreting the results. Y.N. provided reference opinions based on clinical experience. F.H. wrote the first draft of the paper. J.H. reviewed and revised the professional content related to infection. S.T. and J.L., as corresponding authors, controlled the quality of the paper, guided the revision and communicated externally.
Corresponding authors
Ethics declarations
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethical approval
This study was approved by the Affiliated Hospital of Youjiang Medical University for Nationalities.
Declaration of generative AI and AI-assisted technologies in the writing process
No generative AI and AI-assisted technologies were used in the writing process.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Long, C., Lin, Z., Cao, J. et al. CagA-positive Helicobacter pylori may be associated with current infection of clonorchiasis. Sci Rep 15, 39147 (2025). https://doi.org/10.1038/s41598-025-27052-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-27052-3
