
Abstract
Environmental heterogeneity plays a crucial role in shaping the distribution and composition of natural vegetation, including understory ferns. This study investigated the influence of environmental variation on understory fern communities within a one-hectare permanent plot in a low-altitude mountain forest in central Taiwan. Twenty-two environmental factors, including topographic, soil, and biotic variables, were recorded. Multiple regression, cluster (two-way indicator species analysis, TWINSPAN), and ordination (detrended correspondence analysis, DCA, and canonical correspondence analysis, CCA) analyses were conducted. A total of 51 fern species (including Lycophytes) belonging to 20 families and 30 genera were recorded. Among these, 43 were terrestrial, and eight were epiphytic; however, only terrestrial species were analyzed because of the limited representation of epiphytes. Multiple regression analyses revealed that environmental variables significantly affected fern richness, abundance, and community composition. Specifically, stream distance and the importance value (IV) of the saplings significantly influenced fern richness; herb/vine IV affected abundance; and the carbon-to-nitrogen ratio (C/N), manganese concentration (Mn), and herb/vine IV impacted the first axis of the DCA. Furthermore, the elevation, curvature, slope, and topographic wetness index (TWI) significantly influenced the second axis of the DCA. In all the models, topographic variables—particularly stream distance—were one of the most influential drivers. TWINSPAN categorized the ferns into four distinct groups (Diplazium donianum var. donianum [DIPLDO], D. donianum var. aphanoneuron [DIPLAP], Blechnopsis orientalis [BLECOR], and Angiopteris lygodiifolia [ANGILY]), and CCA revealed that environmental factors structured the community compositions in line with the TWINSPAN grouping. The DIPLDO and DIPLAP groups were associated with ridges and upper slope habitats characterized by higher elevations and drier conditions. In contrast, the BLECOR and ANGILY groups were associated with lower elevations, stream proximity, steeper slopes, and higher humidity. This study highlights the role of topographic and soil C/N heterogeneity in structuring fern communities in fine-scale plots for future ecological monitoring in subtropical forest ecosystems.
Similar content being viewed by others


Introduction
Environmental gradients strongly influence the diversity and spatial distribution of plant communities. At regional scales, climate variables such as temperature and precipitation are dominant drivers1,2,3. In contrast, at local scales, fine-scale topography, soil characteristics, and biological interactions play significant roles4,5,6,7,8,9. Among biotic factors, canopy structure and density are widely recognized for their influence on understory plant communities10,11. Ferns, the second largest group of vascular plants, are a dominant component of understory vegetation in tropical and subtropical forests3,12 and are particularly sensitive to environmental heterogeneity.
Topography—such as elevation, slope, aspect, and stream proximity—shapes microhabitats by altering light, temperature, and moisture regimes13,14. Topographic variation at the local scale is known to affect both fern diversity10 and abundance15. Stream proximity, in particular, is a strong predictor of fern assemblages16.
Soil characteristics are also closely tied to fern distribution. Soil moisture (or humidity) is critical for the growth and development of ferns5,17. Variables such as nutrient content (e.g., N, P, K, Ca, and Mg), pH, and organic matter significantly influence fern performance5,16,18. The carbon-to-nitrogen (C/N) ratio, in particular, serves as a proxy for soil fertility and has been linked to fern richness in several tropical studies19,20. However, soil properties are partially influenced by topographic variations21,22, which in turn may affect the distribution of ferns.
Understory ferns depend on canopy-mediated light availability for growth and reproduction23,24. Canopy openness not only alters photosynthetically active radiation but also modulates temperature and humidity in the understory25. Research in Southeast Asia and Taiwan suggests that canopy openness is an important factor influencing fern richness and cover10,26. Furthermore, ferns interact with other plant groups. Dense fern layers can suppress tree seedling recruitment27,28, whereas the diversity of co-occurring understory taxa appears to influence fern richness in varying ways29,30. These biotic interactions may result in mutual inhibition or facilitation depending on local environmental conditions.
Ferns reproduce via spores that are readily dispersed by wind. Despite the fact that many ferns may produce spores capable of travelling long distances, chances of establishing new populations are low31. Allopatric differentiation may be associated with gametophytes that are highly sensitive to microclimatic and edaphic parameters32,33. In addition, environmental factors at the mesoscale—such as soil moisture, humidity, temperature, wind speed, rainfall, vegetation type, and canopy openness—significantly influence fern distribution by affecting sporophytes’ water requirements, temperature tolerance, and photosynthetic capacity5,26,34,35,36. Therefore, the diversity of forest microenvironments varies across regions and is reflected in corresponding differences in fern diversity and composition. We surveyed the relationship between environmental heterogeneity and fern diversity within a one-hectare plot embedded in a broader 25-hectare permanent plot in the Lienhuachih region, central Taiwan. We addressed the following questions: (1) Which environmental factors most strongly influence fern richness, abundance, and composition? (2) How do ferns cluster into ecological groups on the basis of these factors?
Materials and methods
Study area
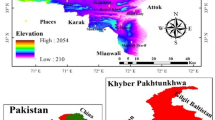
The study site is located in the Lienhuachih Experimental Forest (23°55’N, 120°52’E), which is located in a low-altitude mountainous area of central Taiwan (Fig. 1). A one-hectare permanent plot was established within a natural forest and represents the northwestern section of a broader 25-hectare forest dynamics plot initiated in 2007. The plot encompasses both mid-slope and riparian habitats, with an elevation range spanning from 755 m to 814 m (Table S1). On the basis of earlier tree surveys (DBH ≥ 1 cm), two forest types were identified: one dominated by Diospyros morrisiana and Cryptocarya chinensis and the other by Machilus japonica var. kusanoi and Helicia formosana37. The region experiences a mean annual temperature of 21.2℃ and receives approximately 2,178 mm of precipitation annually, with rainfall concentrated from March to September and a dry season from October to November26. The soil properties of the 25-hectare plot are partially influenced by topography21.

Map of Taiwan (left) and a topographic map (right) of the one-hectare natural forest plot in Lienhuachih within the Houloun stream catchment, a mountainous region in central Taiwan.
Sampling design
The one-hectare plot (100 m × 100 m) was divided into 100 subplots (10 m × 10 m) used as survey units. Vegetation data were collected per subplot, within which all ferns, herbs, vines, and tree saplings (< 1 cm in DBH, > 30 cm in height) were recorded. Fern abundance was assessed by counting individual clumps (treating each clump as one individual) and by estimating percent cover. Epiphytic ferns and vines were measured by the horizontal projection of their canopy cover. Tree data (DBH ≥ 1 cm) were also recorded. Ferns were categorized as either terrestrial or epiphytic, with the former defined as those growing on soil or rocks and the latter as those occurring on tree trunks. Taxonomy follows Volume 6 of the Flora of Taiwan38 and the classification by Kuo et al. (2019)39. Surveys were conducted from July 13 to 15, 2023.
Environmental variables
The topographic variables included elevation, plan curvature, slope, aspect, topographic wetness index (TWI), and distance to the stream. These data were collected from the center of each subplot. Soil variables (data were collected by Chang et al. (2013)40 (2023)41 ) included pH, the carbon-to-nitrogen ratio (C/N), nitrogen, phosphorus, potassium, calcium, magnesium, manganese, zinc, iron, and copper. Soil data were collected at a 20 m × 20 m resolution, with each 10 m × 10 m subplot assigned the values of its nearest soil sample point. Soil moisture was measured at a depth of 5 cm using a RiXEN M-700 S meter between February 26 and March 3, 2024, and the data were averaged from three diagonal points per subplot. Canopy openness was measured from March 15 to 22, 2024, using spherical crown densiometers at a height of 1.3 m at the center of each 10 m × 10 m subplot.
Statistical analysis
Since the soil properties of the 25-hectare plot are influenced by topography, a two-factor Pearson correlation coefficient analysis was conducted to examine the relationships between topography and soil properties in this study plot. Forward stepwise multiple regression (FSMR) with Poisson and linear models was employed to examine the effects of environmental factors on fern diversity. The abundance data for each species were quantified using the importance value (IV), which was calculated as the sum of its relative density (individuals per species/total individuals, unit: %) and relative cover (cover per species/total cover, unit: %). Twenty-two environmental variables—spanning topography (elevation, plan curvature, slope, aspect, TWI, and stream distance), soil properties (pH, C/N, N, P, K, Ca, Mg, Mn, Zn, Fe, Cu, and soil moisture), and biotic factors (canopy openness, tree density, sapling IV, and herb/vine IV)—were included in the analyses (Supplementary Table S1). The dependent variables included fern richness (species count), abundance (IV), and community composition (first two DCA axes). FSMR was performed for model selection using the “MuMIn” package in R. Significant predictors were selected (p < 0.05, chi-square test [Poisson] for fern richness and F test for fern abundance and composition [linear]), and then collinearity was assessed using the variance inflation factor (VIF). Variables with a VIF > 5 were iteratively excluded. Finally, Poisson regression was used to analyze fern richness and its selected predictor variables, whereas linear regression was applied to abundance, composition, and their respective selected predictors.
Community classification was performed using two-way indicator species analysis (TWINSPAN) (dissimilarity metric = total inertia)42. Detrended correspondence analysis (DCA)43 was used to ordinate species and subplots. Canonical correspondence analysis (CCA)44 related species distributions to environmental gradients. The raw data utilized in the aforementioned analysis were derived from the species-subplot matrix, with the data comprising the previously described importance value (IV). All analyses were performed in R v4.3.1.
Results
A total of 51 fern species representing 20 families and 30 genera were recorded within the one-hectare plot. Of these, 43 species were terrestrial, and eight were epiphytic. Diplazium dilatatum was the most abundant species, with 1,011 individuals observed in 98 subplots, followed by Pleocnemia winitii, with 567 individuals in 90 subplots. These two species accounted for 55.8% of the total abundance of terrestrial ferns (Table 1). In contrast, ten terrestrial species were found in only one subplot (Supplementary Table S2), representing 23.3% of the total terrestrial fern richness.
In addition to ferns, 37 herb, 43 vine and 76 sapling species were recorded, resulting in a total of 207 understory species (including 8 epiphytic fern species). Owing to their limited abundance and patchy distribution, epiphytic ferns were excluded from further analyses but are documented in Supplementary Table S2. Among the environmental variables, elevation was most significantly correlated with soil properties (11), followed by slope (nine) and stream distance (seven) (Supplementary Table S3).
The regression models (Table 2) revealed that among the six selected variables, fern richness was significantly influenced by stream distance (negatively) and sapling abundance (positively). Fern abundance was most strongly associated with herb/vine IVs. With respect to fern composition, DCA1 was associated with stream distance, the C/N ratio, manganese, and herb/vine IV—with only stream distance being not significant. The best model of DCA2 included eight variables, among which elevation, curvature, slope, and TWI were significant.
TWINSPAN classified the fern community into four groups (Fig. 2a; Supplementary Figure S1): the Diplazium donianum var. donianum group (DIPLDO; n = 52), the D. donianum var. aphanoneuron group (DIPLAP; n = 21), the Blechnopsis orientalis group (BLECOR; n = 8), and the Angiopteris lygodiifolia group (ANGILY; n = 19). The mean fern richness was lowest in DIPLDO (4.2 ± 1.9) and highest in ANGILY (6.4 ± 2.5) (Supplementary Figure S2). Fern abundance (log-transformed) was positively correlated with richness (r = 0.46, p < 0.001), a pattern that was consistent across groups (Fig. 3). However, significant correlations were observed only in the DIPLDO and ANGILY groups, whereas the other two groups showed no significant correlation.

TWINSPAN and CCA from a one-hectare natural forest plot in the Lienhuachih area of central Taiwan. (a) TWINSPAN identified four fern groups: DIPLDO (□), DIPLAP (△), BLECOR (○), and ANGILY (●). (b) The figure of the first two axes from the CCA; the words beside the lines represent environmental and biological factors, and the direction indicates the trend in which the value increases. (c) The same analysis as in b, with the letters representing the fern species (see Table 1).

Relationships between fern abundance (log-transformed) and richness (r = 0.46, p < 0.001). The Pearson correlation in the four fern groups was r = 0.48 (p < 0.001, DIPLDO), 0.26 (p = 0.264, DIPLAP), 0.50 (p = 0.205, BLECOR), and 0.65 (p = 0.002, ANGILY).
The cumulative explained variance of the first three CCA axes was 9.7%, 16.5%, and 21.2%, respectively. On the first axis of the CCA, herb/vine IV had the highest absolute score (0.80), followed by C/N (0.54), elevation (–0.50), and stream distance (–0.46); on the second axis of the CCA, stream distance (–0.61) had the highest absolute score, followed by Ca (–0.58), elevation (–0.50), and slope (0.43). In addition, herb/vine IV, stream distance, Ca and C/N were the most important determinants of one of the first two CCA axes (Table 3). As shown in Fig. 2b, the BLECOR group is more distinct, whereas the ANGILY group somewhat overlaps with the other groups, and the DIPLDO and DIPLAP groups exhibit greater overlap. Our results showed that these fern groups have adapted to different environments (Fig. 4).

Variation in elevation (a), stream distance (b), slope (c), and C/N (d) among different fern communities. Different letters denote statistically significant differences among the different types of fern vegetation (p < 0.05).
The species ordination (Fig. 2c) revealed dominant ferns (Diplazium dilatatum and Pleocnemia winitii) near the plot center, whereas the species of named TWINSPAN groups aligned with the environmental characteristics of their respective groups. For example, DIPLDO’s D. donianum var. donianum was located in a topographic and edaphic space that is indicative of drier, upland sites.
Discussion
Topographic effects
Topography has long been recognized as a key determinant of forest vegetation patterns8,45,46. In this study, elevation and stream distance emerged as primary predictors of fern richness and composition, despite the modest elevation range (~ 59 m) within the plot. These gradients reflect moisture availability: ridges with higher elevations and well-drained soils tend to be drier, whereas lower streamside zones retain more moisture. The strong correlation between elevation and stream distance (r = 0.40, p < 0.001) reinforces this interpretation. Our findings align with those of previous studies5,10,16 in montane forests where even fine-scale topographic variation influences fern diversity. For the other plant taxa in the 25-ha plot (of which our 1-ha plot was a part), topography was the most important factor affecting the changes in the plant community and species composition37.
Soil effects
Soil properties such as nutrient concentrations and organic matter content often covary with topography because of erosion, leaching, and deposition21,47. While some studies have indicated a positive correlation between soil fertility and fern richness49,50, others have shown that lower fertility results in more fern species15,20. In our plot, C/N was significantly associated with fern composition, particularly along the first two CCA axes. Stream-adjacent soils, which are rich in organic matter and nitrogen, presented elevated carbon-to-nitrogen (C/N) ratios because of the greater accumulation of organic matter than that associated with decomposition in moist areas. Although fern richness was not directly correlated with C/N, its indirect effects via topographic mediation were evident. Calcium and manganese were also included in the regression models, although their contributions were relatively modest.
Water serves as a critical determinant of both fern richness and distribution4,5,24. Water availability, inferred through the topographic wetness index (TWI), slope, and stream distance, likely exerts a dominant control on fern distributions. While soil moisture in the 25-ha plot (6 transects) was an important factor for the understory plants, including fern species30, it did not emerge as a significant predictor in the models in this study plot. The strong influence of hydrologically relevant topographic variables suggests their overriding importance in determining local fern composition.
Biotic influences
Light availability is a well-documented driver of fern performance and affects morphology, abundance, and richness10,24. Although canopy openness was not retained in the final regression models, its significant correlation with tree density (r = − 0.22, p < 0.05) implies indirect effects. Denser tree canopies may reduce understory light, thus constraining fern growth.
Interestingly, this study revealed positive associations between fern richness (or abundance) and both sapling and herb/vine cover, contrary to previous findings that emphasized competitive suppression28,30,48. The factors contributing to this outcome are likely multifaceted. The underlying mechanism may involve moisture availability, which is influenced by proximity to the stream. Although herb and vine cover are strongly correlated with fern abundance and composition, the primary determinant appears to be the distance from streams, as areas closer to streams generally exhibit higher moisture levels. Tuomisto et al. (2002) similarly reported that fern and Melastomataceae diversity co-occurred with tree richness in fertile tropical soils29.
Fern community grouping and habitat differentiation
TWINSPAN and CCA revealed clear compositional differentiation among the four fern groups, corresponding to distinct habitat types. The DIPLDO and DIPLAP groups were associated with ridges and upper slope habitats characterized by higher elevations and drier conditions. In contrast, the BLECOR and ANGILY groups were associated with lower elevations, stream proximity, steeper slopes, and higher humidity. These habitat preferences support the role of environmental filtering in fern assembly and align with prior vegetation classifications within the same forest37.
In terms of the correlation between fern richness and abundance, compared with the DIPLDO group, the ANGILY group exhibited communities with greater evenness. These findings suggest that the environment inhabited by the ANGILY group is more conducive to the survival of a diverse range of ferns.
Conclusion
This study highlights the significant role of environmental heterogeneity in shaping fern diversity and community composition in a low-altitude subtropical forest. Among the examined factors, topographic variables—particularly stream distance—exerted one of the most influential drivers of fern richness, abundance, and species assemblage. Soil properties, especially C/N, further mediated these relationships and reflected microhabitat variation. The classification of ferns into four ecological groups on the basis of environmental gradients highlights the structuring effect of habitat differentiation. The DIPLDO and DIPLAP groups occupied ridges and upper slope habitats characterized by higher elevations and drier conditions, whereas the BLECOR and ANGILY groups were associated with lower elevations, greater proximity to streams, steeper slopes, and higher humidity. This study highlights the role of topographic and soil-related heterogeneity in structuring fern communities in fine-scale plots. Long-term monitoring incorporating both abiotic and biotic variables is essential for understanding how fern communities respond to environmental change and for informing conservation strategies in subtropical forest ecosystems.
Data availability
The datasets utilized and/or analyzed during the current study are available from the first author upon reasonable request.
References
Antonelli, A. et al. Geological and Climatic influences on mountain biodiversity. Nat. Geosci. 11, 718–725 (2018).
Lin, H. Y. et al. Climate-based approach for modeling the distribution of montane forest vegetation in Taiwan. Appl. Veg. Sci. 23, 239–253 (2020).
Suissa, J. S., Sundue, M. A. & Testo, W. L. Mountains, climate and niche heterogeneity explain global patterns of fern diversity. J. Biogeogr. 48, 1296–1308 (2021).
Richard, M., Bernhardt, T. & Bell, G. Environmental heterogeneity and the Spatial structure of fern species diversity in one hectare of old-growth forest. Ecography 23, 231–245 (2000).
Karst, J., Gilbert, B. & Lechowicz, M. J. Fern community assembly: the roles of chance and the environment at local and intermediate scales. Ecology 86, 2473–2486 (2005).
Bohlman, A. S. et al. Importance of soils, topography and geographic distance in structuring central Amazonian tree communities. J. Veg. Sci. 19, 863–874 (2008).
Da Silva, K. E. et al. Tree species community Spatial structure in a Terra firme Amazon forest, Brazil. Bosque (Valdivia). 35, 347–355 (2014).
Zhou, T. et al. Biodiversity of Jinggangshan mountain: the importance of topography and geographical location in supporting higher biodiversity. PLoS ONE. 10, e0120208 (2015).
Zuleta, D. et al. Importance of topography for tree species habitat distributions in a Terra firme forest in the Colombian Amazon. Plant. Soil. 450, 133–149 (2020).
Jones, M. M., Cicuzza, D., Straaten, O., Veldkamp, E. & Kessler, M. Determinants of fern and angiosperm herb community structure in lower montane rainforest in Indonesia. J. Veg. Sci. 25, 1216–1224 (2014).
Mendes, C. N., Diniz, E. S., Terra, M. C. N. S., Jeannot, K. K. & Fontes, M. A. L. Light conditions imposed by canopy: allometric strategies of an understorey palm (Geonoma schottianamart) in Atlantic forest. J. Trop. Sci. 31, 332–342 (2019).
Weigand, A. et al. Global fern and lycophyte richness explained: how regional and local factors shape plot richness. J. Biogeogr. 47, 59–71 (2020).
Chao, K. J., Chao, W. C., Chen, K. M. & Hsieh, C. F. Vegetation dynamics of a lowland rainforest at the Northern border of the paleotropics at Nanjenshan, Southern Taiwan. Taiwan. J. Sci. 25, 29–40 (2010).
Moeslund, J. E., Arge, L., Bøcher, P. K., Dalgaard, T. & Svenning, J. C. Topography as a driver of local terrestrial vascular plant diversity patterns. Nord J. Bot. 31, 129–144 (2013).
Cicuzza, D. & Mammides, C. Soil, topography and forest structure shape the abundance, richness and composition of fern species in the fragmented tropical landscape of Xishuangbanna, Yunnan, China. Forests 13, 1453 (2022).
Bonari, G. et al. Fine-scale fern ecological responses inform on riparian forest habitat conservation status. Biodivers. Conserv. 31, 2141–2161 (2022).
Poulsen, A. D., Tuomisto, H. & Balslev, H. Edaphic and floristic variation within a 1-ha plot of lowland Amazonian rain forest. Biotropica 38, 468–478 (2006).
Viana, J. L., Turner, B. L. & Dalling, J. W. Compositional variation in understorey fern and palm communities along a soil fertility and rainfall gradient in a lower montane tropical forest. J. Veg. Sci. 32, 12947 (2021).
Lwanga, J. S., Balmford, A. & Badaza, R. Assessing fern diversity: relative species richness and its environmental correlates in Uganda. Biodivers. Conserv. 7, 1387–1398 (1998).
Zuquim, G. et al. Predicting environmental gradients with fern species composition in Brazilian Amazonia. J. Veg. Sci. 25, 1195–1207 (2014).
Chang, L. W. et al. Lienhuachih Subtropical Evergreen Broadleaf Forest Dynamics Plot Tree Species Characteristics and Distribution Patterns 334–335 (Taiwan Forestry Research Institute, 2012).
Wei, J. B., Xiao, D. N., Zeng, H. & &Fu, Y. K. Spatial variability of soil properties in relation to land use and topography in a typical small watershed of the black soil region, Northeastern China. Environ. Geol. 53, 1663–1672 (2008).
Messier, C., Parent, S. & Bergeron, Y. Effects of overstory and understory vegetation on the understory light environment in mixed boreal forests. J. Veg. Sci. 9, 511–520 (1998).
Hill, J. D. & Silander, J. A. Distribution and dynamics of two ferns: Dennstaedtia punctilobula (Dennstaedtiaceae) and Thelypteris noveboracensis (Thelypteridaceae) in a Northeast mixed hardwoods–hemlock forest. Amer. J. Bot. 88, 894–902 (2001).
Angelini, A., Corona, P., Chianucci, F. & Portoghesi, L. Structural attributes of stand overstory and light under the canopy. Ann. Silvic Res. 39, 23–31 (2015).
Lee, P. H., Huang, Y. M., Chiou, W. L., Tseng, Y. H. & Tzeng, H. Y. The correlation between fern diversity and environmental factors-case study in a Chinese fir plantation in Lienhuachih, central Taiwan. Taiwan J. For. Sci. 39, 271–289 (2024).
George, L. O. & Bazzaz, F. A. The fern understory as an ecological filter: growth and survival of canopy-tree seedlings. Ecology 80, 846–856 (1999).
Royo, A. A. & Carson, W. P. On the formation of dense understory layers in forests worldwide: consequences and implications for forest dynamics, biodiversity, and succession. Can. J. Res. 36, 1345–1362 (2006).
Tuomisto, H. et al. Distribution and diversity of pteridophytes and Melastomataceae along edaphic gradients in Yasuní National Park, Ecuadorian Amazonia. Biotropica 34, 516–533 (2002).
Chang, L. W., Huang, J. L., Luo, S. F. & Lee, P. H. Understory plant composition and its relations with environmental factors of the Lienhuachih forest dynamics plot at a subtropical evergreen broadleaf forest in central Taiwan. Taiwan J. For. Sci. 30, 245–257 (2015).
Wolf, P. G., Schneider, H. & Ranker, T. A. Geographic distributions of homosporous ferns: does dispersal obscure evidence of vicariance? J. Biogeogr. 28, 263–270 (2001).
Page, C. N. The diversity of ferns: an ecological perspective. In: The Experimental Biology of Ferns (ed. Dyer, A. F.) 10–56 (1979).
Leake, J. R. The biology of myco-heterotrophic (saprophytic) plants. New. Phytol. 127, 171–216 (1994).
Sanginés-Franco, C., Luna-Vega, I., Ayala, O. A. & Contreras-Medina, R. Distributional patterns and biogeographic analysis of ferns in the Sierra madre Oriental, Mexico. Amer Fern J. 101, 81–104 (2011).
Chiu, T. Y. et al. Ecophysiological characteristics of three Cyathea species in Northeastern Taiwan. Taiwan J. For. Sci. 30, 147–155 (2015).
Silva, V. L., Mehltreter, K. & Schmitt, J. L. Ferns as potential ecological indicators of edge effects in two types of Mexican forests. Ecol. Indic. 93, 669–676 (2018).
Chang, L. W. et al. Changes of plant communities classification and species composition along the micro-topography at the Lienhuachih forest dynamics plot in the central Taiwan. Taiwania 57, 359–371 (2012).
Boufford, D. E. et al. A Checklist of the Vascular Plants of Taiwan. in Flora of Taiwan, 2nd edition, Vol. 6 (ed. Editorial Committee of the Flora of Taiwan, S. E.) 15–139 (2003).
Kuo, L. Y. et al. Updating Taiwanese pteridophyte checklist: A new phylogenetic classification. Taiwania 64, 367–395 (2019).
Chang, L. W., Zelený, D., Li, C. F., Chiu, S. T. & Hsieh, C. F. Better environmental data May reverse conclusions about niche- and dispersal-based processes in community assembly. Ecology 94, 2145–2151 (2013).
Chang, L. W., Chiu, S. T. & Hsieh, C. F. Contrasting Spatial distribution of pioneer versus non-pioneer saplings in a Taiwanese forest: A multiple scale approach. Ecol. Res. 38, 604–616 (2023).
Rolecěk, J., Tichy, L., Zeleny, D. & Chytry, M. Modified TWINSPAN classification in which the hierarchy respects cluster heterogeneity. J. Veg. Sci. 20, 596–602 (2009).
Hill, M. O. & Gauch, H. G. Detrended correspondence analysis: an improved ordination technique. Vegetatio 42, 47–58 (1980).
Ter Braak, C. J. F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 67, 1167–1179 (1986).
Itoh, A. et al. Importance of topography and soil texture in the Spatial distribution of two sympatric dipterocarp trees in a Bornean rainforest. Ecol. Res. 18, 307–320 (2003).
Jucker, T. et al. Topography shapes the structure, composition and function of tropical forest landscapes. Ecol. Lett. 21, 989–1000 (2018).
Wilcke, W. et al. Soil properties and tree growth along an altitudinal transect in Ecuadorian tropical montane forest. J. Plant. Nutr. Soil. Sci. 171, 220–230 (2008).
Liang, M. et al. Research progress on the ecology of dense fern understory. J. Trop. Subtrop Bot. 30, 219–300 (2022).
Ruokolainen, K., Tuomisto, H., Macía, M. J., Higgins, M. A. & Yli-Halla, M. Are floristic and edaphic patterns in Amazonian rain forests congruent for trees, pteridophytes and melastomataceae? J. Trop. Ecol. 23, 13–25 (2007).
Jones, M. M., Ruokolainen, K., Martinez, N. C. L. & Tuomisto, H. Differences in topographic and soil habitat specialization between trees and two understorey plant groups in a Costa Rican lowland rain forest. J. Trop. Ecol. 32, 482–497 (2016).
Acknowledgements
We thank the students of the Department of Forestry at National Chung Hsing University for their assistance in the wild investigation.
Author information
Authors and Affiliations
Contributions
Conception or design of the work: Pei-Hsuan Lee, Hsy-Yu Tzeng. Data collection: Pei-Hsuan Lee, Li-Wan Chang, Jian-Hong Yang. Data analysis and interpretation: Pei-Hsuan Lee. Drafting the article: Pei-Hsuan Lee. Critical revision of the article: Wen-Liang Chiou, Yao-Moan Huang, Hsy-Yu Tzeng, Yen-Hsueh Tseng. All the authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lee, PH., Huang, YM., Chang, LW. et al. Environmental heterogeneity and its influence on fern diversity in a low-altitude mountain forest in central Taiwan. Sci Rep 15, 44383 (2025). https://doi.org/10.1038/s41598-025-28048-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-28048-9
