
Abstract
Cervical dystonia (CD) is a rare movement disorder marked by involuntary neck muscle contractions. Although somatosensory dysfunction has been proposed, the underlying pathophysiology remains unclear. Temporal discrimination (TD) deficits are discussed as potential markers of a dystonic endophenotype, possibly linked to proprioceptive impairments. This study aimed to assess TD in the neck region and explore its relationship with proprioception in head movements. We evaluated somatosensory temporal discrimination threshold (STDT) and temporal discrimination movement threshold (TDMT) in 20 CD patients and 20 healthy controls (HC). Both measures were applied directly to the cervical region: STDT via skin stimuli on the lateral neck and TDMT via muscle stimuli to the splenius capitis. Proprioception was assessed through a head rotation task. Clinical severity was measured using the TWSTRS scale. STDT and TDMT thresholds were significantly elevated in CD patients compared to HCs and correlated positively with dystonia severity. However, there were no group differences in proprioceptive performance, nor was proprioception correlated with TD or TWSTRS scores. TD is clearly altered in CD and associated with symptom severity, supporting its role in dystonia pathophysiology. In contrast, no proprioceptive deficits were observed, and no link between TD and proprioception was found. This suggests TD and proprioception may represent distinct dysfunctions rather than connected elements of sensorimotor integration. Further studies are needed to refine proprioceptive testing in CD and clarify its relationship with TD.
Similar content being viewed by others


Introduction
As a clinical symptom, dystonia refers to an involuntary, sometimes painful, contraction of muscle groups. Beyond, the term dystonia describes a heterogeneous group of movement disorders where dystonic postures and/or movements are at the core of the clinical picture. These disorders are classified, inter alia, according to the location of the dystonic symptoms1. Cervical dystonia (CD) is the most common form of focal dystonia. Patients are clinically characterised by unnatural twisting of the neck and movements of the head. In addition, CD patients often suffer from non-motor symptoms such as depression, anxiety and social stigma. The burden of disease in this patient group can be significant2. Patients are primarily treated with selective denervation of the dystonic muscle groups by injection of botulinum neurotoxin (BoNT) or, less commonly, with anticholinergic drugs3,4. For patients who do not respond to these therapies, deep brain stimulation (DBS) of the internal globus pallidus is available as a second-line treatment5,6.
For a long time considered a disease of the basal ganglia, a body of research on the pathophysiology of dystonia now points to a network disorder of the central nervous system including cortical structures, the thalamus and the cerebellum7,8. From a neurophysiological perspective, deficits in three processes are considered to contribute to the development of dystonic phenotypes: altered cortical plasticity9,10, loss of cortical inhibition11,12, and altered sensorimotor integration13,14. In order to support these pathophysiological assumptions, there are increasing efforts to characterise stable biomarkers including the use of neurophysiological paradigms to define so-called endophenotypes. In this context, the term endophenotype refers to any measurable component potentially linking clinical presentation (phenotype) and its genetic basis (genotype).
One such paradigm is Temporal Discrimination (TD), which tests the cognitive ability to distinguish between stimuli presented in rapid succession. TD has already been described to be altered in several forms of dystonia, which is considered a possible correlate of altered sensorimotor integration in dystonia patients15,16. Specifically for CD, the paradigm has often been studied at the extremities17,18,19,20, whereas, to the best of our knowledge, only one study has assessed (somatosensory) TD at the neck region21. This appears surprising as the focal appearance draws natural interest in the physiology of this particular body region.
Assessing TD directly in the neck region offers several advantages: it targets the body region clinically affected by cervical dystonia, reduces potential confounding by peripheral impairments such as neuropathy or radiculopathy, allows the investigation of region-specific alterations, and enables the simultaneous assessment of proprioception in the dystonic region.
In a preliminary study, we discussed a possible association between impaired TD and reduced proprioception in healthy subjects and patients with polyneuropathy22. In dystonia, patients are known to exhibit proprioceptive deficits23,24, which may play a role in linking the endophenotypic significance of TD to the phenotypic presentation, together with other processes such as abnormal basal ganglia and cerebellar function.
Based on these considerations and preliminary examinations, we aimed to test the following three hypotheses: (i) multimodal assessment of TD at the neck region is feasible, (ii) TD at the neck region is reduced in people with CD compared to healthy controls, and (iii) TD is correlated with performance in a proprioception test of head movements.
Materials and methods
The protocol conformed to the tenets of the Declaration of Helsinki and was approved by the Ethics Committee at the Medical Faculty of the University of Würzburg. All participants gave written informed consent to participate in the study.
Participants
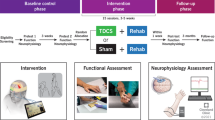
A total of 20 patients with CD, as diagnosed by a movement disorders specialist, were recruited from our outpatient clinic for movement disorders. Clinical assessment included the motor subscale of the Toronto Western Spasmodic Torticollis Rating Scale (TWSTRS)25. Demographic data and clinical characteristics of the patients are summarised in Table 1. Most patients were treated with periodic injections of BoNT A, and only a few received concomitant oral antidystonic therapy or other CNS-active medications (see Table 1 for details). In all cases, dosage had been stable for at least 3 months without subjective side effects. The experiment was conducted at least 10 weeks after the last injection of BoNT A, when both the investigator and the patient judged that there were no or only minor treatment effects remaining. The study excluded patients with clinically relevant psychiatric or neurological comorbidities. All participants were screened for cognitive impairment and depression (MoCA, BDI); only those with normal scores were included. In addition, a control group of 20 healthy volunteers (HC), matched for age and sex, was recruited. All subjects were assessed for clinical deficits in deep sensation and proprioception, testing both pallesthesia and positional sense of the hallux. Individuals with abnormalities were excluded as peripheral afferent dysfunction may interfere with TD assessment22.
Measures of temporal discrimination
Two variants of TD, i.e. somatosensory and kinaesthetic TD, were investigated in this study. The former tests the perception of superficial stimuli on the skin, while the latter refers to the perception of electrically induced muscle movements. Surface electrodes (anode and cathode 1 mm in diameter and 1.5 cm apart) were placed over the lateral neck region to test somatosensory TD thresholds (STDT) (see Fig. 1A). A constant current stimulator (Digitimer, Welwyn Garden City, United Kingdom) delivered pairs of square-wave electrical stimuli of 0.2 ms duration. Stimulation intensity was individually determined by gradually increasing current until participants could clearly perceive stimuli (i.e., 10 out of 10 trials). The stimuli were presented in ascending and descending sequences, each performed twice, starting either from 30 ms or 300 ms, with 5 ms increments. The average was taken for further analysis. STDT was defined as the shortest interstimulus interval (ISI) when participants perceived two separate pulses. For the assessment of kinaesthetic TD, the so-called TD motor threshold (TDMT) was determined. For this purpose, a monopolar electromyography (EMG) needle electrode was inserted into the right splenius capitis muscle (see Fig. 1B). The anode was a surface electrode which was placed above the C7 vertebral body. Using the same stimulation protocol as described above, the TDMT was defined as the shortest ISI at which subjects were able to discriminate clearly (i.e. three times in a row) two separate contractions of the stimulated muscle. Again, the mean of four attempts (i.e. two runs in both ascending and descending order) was used for further analysis.

Schematic illustration of the temporal discrimination measurements: STDT (A) and TDMT (B).
Proprioception task
Proprioception was assessed via a modified head-to-target repositioning task26,27 using a goniometer-based device consisting of a helmet with a smartphone (iPhone 12 Pro, Apple, Los Altos (CA), USA) attached (see Fig. 2A). Head position angles were measured using the iPhone compass application28,29,30. During the test, great care was taken to ensure that the device was positioned horizontally, in order to avoid angular errors. Participants wore an eye mask and performed the tasks without visual feedback. At the start of the procedure, each participant was instructed to adopt a natural and relaxed head position that subjectively corresponded to a neutral gaze direction, without engaging in compensatory muscle activity to correct dystonia. This position was defined as the individual neutral position and referenced as 0 degrees in the compass application. CD patients and HC were made familiar with four different head positions corresponding to rotations of 30 and 60 degrees to the left and right, respectively (see Fig. 2B). The learning process was as follows: Starting from the neutral position, the experimenter passively rotated the participant’s head to the respective position, where it was held for two seconds. This procedure was repeated twice for each of the four positions. The ability to actively reproduce the learned head positions was then assessed. Each trial started from the neutral position, where initially the participant’s head was fixed by the experimenter. Then, the experimenter announced one of the four positions in a pseudorandomised order and released the participant’s head. The participant was instructed to actively move the head to the announced position. When the target position was subjectively reached, the participant signalled this by saying ‘yes’. Before the next position was announced, the experimenter repositioned the participant’s head back to the neutral position. Each position was announced a total of three times. Movements and angles were documented via screen recording of the compass application and subsequently analysed by the examiner. The absolute error between the target position and the achieved position was used as a measure of proprioceptive performance.

Schematic illustration of the proprioception task (modified head-to-target repositioning task): goniometer-based device using the iPhone compass application (A) and assessing predefined head rotations (B).
Statistics
SPSS software (Version 29.0.2.0, IBM, Armonk (NY), USA, https://www.ibm.com/spss) was used for statistical analyses. According to the Shapiro-Wilk test, data were not normally distributed. Therefore, the Mann-Whitney U test was used for group comparisons and the Spearman test for correlations. Statistical significance was set at p < 0.05.
Results
Twenty CD patients and 20 controls (HC) were included, with 13 women and seven men in each group. The mean age was 59.1 ± 9.6 years in the CD and 59.2 ± 9.9 years in the HC group (p = 0.961). In CD patients, the median TWSTRS motor score was 17 [range 6 to 22] points. The average disease duration was 19.8 ± 12.2 years. Eleven patients presented with dystonic head tremor (see Table 1).
In both temporal discrimination paradigms, the threshold levels of the CD patients were significantly higher than those of the HC group: STDT CD group 132.2 ± 48.7 ms vs. STDT HC group 97.5 ± 21.6 ms (p = 0.029; Fig. 3A) and TDMT CD group 117.3 ± 42.6 ms vs. TDMT HC group 83.3 ± 22.7 ms (p = 0.017; Fig. 3B).

Somatosensory temporal discrimination threshold (STDT; A) and temporal discrimination motor threshold (TDMT; B) in cervical dystonia (CD) vs. healthy controls (HC). The asterisk indicates p < 0.05.
Regarding proprioceptive performance, patients with CD and HC showed comparable angular deviation from the target (9.6° ± 4.5° vs. 8.7° ± 4.0°, p = 0.659). A subgroup analysis comparing a CD cohort with (9.9 ± 5.3, p = 0.670) and without tremor (9.2 ± 3.7, p = 0.799) with HC also showed no group differences.
Looking for associations of temporal discrimination performance with clinical and behavioral measures, respectively, higher STDT as well as TDMT levels were positively correlated with the severity of motor symptoms (i.e. higher TWSTRS scores) in CD patients: Spearman Rho STDT 0.539 (p = 0.014; Fig. 4A) and TDMT 0.647 (p = 0.002; Fig. 4B). In contrast, there were no significant correlations of the proprioceptive performance with clinical rating or TD performance.

Correlation test of STDT (A) and TMDT (B) with dystonia clinical symptom severity (measured by TWSTRS).
As an additional finding, STDT performance correlated with age, i.e. older patients had higher thresholds (Spearman Rho 0.541, p = 0.014). However, this association was not observed for TDMT performance. Furthermore, STDT and TDMT did not differ between patients taking CNS-active medications and those who were not.
Discussion
In this study, the temporal discrimination (TD) of somatosensory and kinaesthetic stimuli was assessed in the neck region of patients with cervical dystonia (CD) and matched healthy controls (HC). To the best of our knowledge, this is the first study to record TDMT from this region and to concomitantly assess physiological and behavioral measures of the affected body area in CD patients. We had hypothesized that (i) TD assessment in the neck region is feasible, (ii) TD levels are elevated in CD in comparison to HC and (iii) TD is correlated with pointing errors in a proprioception task involving the head. In brief: both STDT via surface electrodes and TMDT via EMG needle electrodes in the splenius capitis muscle proved technically feasible and produced plausible results, with STDT and TDMT significantly elevated in CD patients compared to HC. In addition, STDT and TDMT levels correlated significantly with the clinical symptom severity of cervical dystonia. However, there was no significant association of STDT or TDMT with proprioceptive performance in the plane of head rotation.
Temporal discrimination and dystonia
Studies of STDT in dystonic patients were first published at the end of the last century and showed elevated thresholds in measurements from the hand affected by dystonia in patients with writer’s cramp31 and other non-focal dystonias that affect the upper limbs32,33. In the course of time, these findings were also extended to body regions not affected by dystonia – for example, to the non-affected hand in patients with hand dystonia34,35 or to the hand in patients with CD36 or blepharospasm37. It has even been shown that asymptomatic carriers of the DYT1-TOR1A mutation present with similar alterations38. In particular, the latter finding can be interpreted as a strong argument for considering elevated STDT levels as a dystonic endophenotype, i.e. a heritable trait that – albeit with incomplete penetrance - can reflect a predisposition for the development of clinical dystonia. For patients with CD, there are a number of other published observations on this topic: Elevated STDT also extends to the skin of the face and neck21, STDT alterations do not change with therapies such as BoNT39 or DBS40, and they do not change spontaneously over time41. Interestingly, however, increased STDTs could also be demonstrated in patients clinically classified as having psychogenic dystonia42. In conclusion, the finding of an altered STDT appears to be quite robust in the context of the previous literature, and our results explicitly confirm the preliminary findings of Scontrini et al. that STDT is also increased in the neck region of CD patients21.
Whereas the STDT tests the processing of somatosensory stimuli, the TDMT primarily tests the perception of kinesthesia43. Compared to STDT, much less data is available on this paradigm. Again, elevated thresholds have been described in focal dystonia, namely writer’s cramp44. Somewhat surprising, one study found TDMT normal in patients with dystonic tremor and elevated in patients with essential tremor, while exactly the opposite was described for STDT45. As to our knowledge, TDMT was recorded either from the flexor carpi radialis or the first dorsal interosseus muscle in all published studies. Overall, scientific evidence for TDMT in dystonia is less clear than for STDT.
The following conceptual hypothesis regarding the neuroanatomical and neurophysiological underpinnings of TD has emerged from the results of various neuroimaging studies (reviewed in15: TD involves a brainstem–cerebellar–basal ganglia network, with the superior colliculus acting as a key sensory input hub. The cerebellum contributes to precise timing and sensorimotor integration, supporting both somatosensory and kinesthetic processing46. Basal ganglia integrity is crucial for integrating sensory input from the superior colliculus, thalamus, and cerebellum47. Modulated by dopamine, this network helps to select stimuli appearing important for behaviour. Additionally, sensory signals reaching the S1 cortex refine perception by inhibitory interneurons, thereby enhancing TD48,49.
Based on such a neural circuit model, the extent to which deficits in TD may underlie the pathophysiology of CD can be speculated about: Dystonia as well as abnormal TD may stem from a common disruption in the midbrain network that controls attentional orienting. Impaired GABAergic inhibition in the superior colliculus causes abnormal burst firing in visual sensory neurons. This abnormal firing leads to prolonged neuron activation, which then induces hyperexcitability in premotor neurons in the deep layers of the superior colliculus. Along subsequent motor pathways, this hyperactivity may result in the muscle spasms seen in CD48. In addition, the cerebellum likely modulates temporal processing within this network, further influencing abnormal TD and motor output in dystonia.
Apart from the midbrain, the classic basal ganglia structures certainly play a role in explaining the link between TD and dystonia. In focal dystonia, structural and functional putaminal alterations have been described. Putaminal enlargement is seen in adult-onset dystonias like blepharospasm50 and is linked to abnormal TD in laryngeal dystonia51. The putamen is involved at early stages of the TD sequence and is activated during stimulus differentiation52, suggesting its enlargement reflects dysfunction in both focal dystonia and temporal processing15.
The observed correlation between TD abnormalities and clinical disease severity in CD may seem at odds with the idea of TD as an endophenotypic marker—that is, a heritable trait largely independent of symptom expression. Tremor could be a potential contributor, but in our cohort, TD thresholds did not differ between patients with and without tremor, making this unlikely. Instead, elevated TD may rather serve as a biomarker of network dysfunction within the basal ganglia–cerebellar–brainstem circuitry, rather than a purely stable endophenotypic trait.
This perspective aligns with ongoing debates about the stability of TD as an endophenotype and highlights the need to consider broader cognitive and non-motor contributions to dystonia. It has been hypothesised that not only TD, but the decision-making process itself is retarded in dystonia patients53. This seems justified given the high prevalence of non-motor symptoms such as depression, anxiety, and cognitive deficits in dystonia54. Of note, however, in a preliminary study addressing this potential confounding factor, we found equal reaction times in CD patients and healthy controls in a simple reaction time paradigm55.
Proprioception and dystonia
The presence of proprioceptive deficits in focal dystonia is widely accepted by the scientific community and has been substantiated by a body of evidence (reviewed in23,56. The deficits in TD and proprioception are each considered to be independent correlates of impaired sensorimotor integration in dystonia (c.f57. for a review). Abnormal processing of sensory information is considered to be in the centre of pathophysiology, resulting in dystonic motor output.
Sensorimotor integration involves the sensory and motor cortices, as well as premotor areas, the basal ganglia and the cerebellum58. Proprioceptive signals reach the somatosensory cortex via the dorsal column–medial lemniscus pathway passing thalamus and the cerebellum via spinocerebellar tracts for coordination and balance. Obviously, there is a significant overlap between TD and proprioception in terms of their shared networks. Recent work has also shown that basal ganglia structures are involved in the processing of proprioceptive stimuli by recording local field potentials in the pallidum of dystonia patients with DBS59,60,61.
Proprioceptive deficits in CD have been described repeatedly. For example, the tonic vibration reflex has been found abnormal in dystonic patients on conventional upper limb testing62,63. Moreover, people with CD showed alterations in head alignment when the vibration test is applied to the neck muscles compared to HC64. Vibro-tactile stimulation procedures are currently even investigated as a therapeutic method65,66,67. Kinematic studies of purposeful arm movements in patients with CD showed discrete abnormalities, with movements being less accurate in CD than those in matched controls68,69. Furthermore, compared to controls, CD patients showed reduced capacity in anticipating the timing of human movements, but not those of objects70. Another study has demonstrated deficits in motion perception in CD patients using a motion platform71. Thus, there is remarkably little evidence of proprioceptive deficits in CD in the dystonic body region. This may simply be due to the fact that proprioception testing of neck and head movements is not trivial.
In an approach similar to ours, Avanzino et al. observed proprioceptive deficits at neck and upper limb muscles only in CD patients presenting with dystonic tremor72. In the same trial STDT of the hands did not correlate with proprioceptive performance of the upper limb, while TD at the neck region was not tested72. Contrary to those findings, we could not demonstrate behavioral proprioceptive differences at all, neither between CD patients and HC, nor beween CD patients with and without tremor. However, this could also be due to methodological differences: While our subjects were asked to turn their head from a neutral position to a previously learned target position, the task in the study of Avanzino and colleagues was to return to the neutral position after a passive head movement. Nevertheless, our study showed that TD and proprioceptive performance do not correlate, even when assessed in the body region affected by CD. This suggests proprioception and TD more likely to be independent variables and does not support our hypothesis of a direct association between defective TD and impaired proprioception. However, it is conceivable that abnormalities in sensorimotor integration, rather than peripheral proprioceptive input itself, are critical, possibly involving a broader brainstem–cerebellar–basal ganglia network.
Limitations of the study
As common for studies in rare disease, the validity of the results is limited by the relatively small number of patients.
Methodologically, to increase comparability, we used the right splenius capitis muscle for TDMT in all patients, even though it was not the muscle most affected by dystonia in several patients. Individual selection of the muscle depending on clinical presentation would have been an alternative approach worth considering for future studies.
Assessment of proprioceptive performance was based on active pointing rather than position estimation. It cannot be excluded that sensitivity of this test was insufficient to detect subclinical deficits in the perception of head position. However, the approach was chosen for two reasons: First, we had found active pointing to be positively associated with TD performance in an earlier study22. Second, movement of the head by the experimenter might have introduced a bias due to tactile input, with possible effects similar to a sensory trick. Regarding the latter, however, our test apparatus comprising a smartphone mounted on a helmet may have represented a sensory bias itself. In future, non-contacting technical options like vision-based motion analysis should be applied.
Clinical symptoms were assessed using the TWSTRS motor score. However, this score does not include an item related to tremor which could have been used to correlate tremor severity with TD and proprioceptive performance. Given the influence of tremor on TD and proprioception identified in earlier studies45,72, the use of a scale capturing tremor may have been useful in disentangling the contribution of TD and proprioception to the dystonic tremor phenotype.
Conclusion
The present study provides further evidence that TD is altered in CD, with elevated thresholds for STDT and TDMT. Notably, TD thresholds are positively correlated with motor symptom severity of dystonia. These findings suggest that TD may serve as a biomarker of network dysfunction underlying cervical dystonia, rather than a stable endophenotypic trait. In contrast to one of our hypotheses, our proprioceptive assessment of head positions did not show differences between CD and HC, and, though assessed in the dystonic body region, was not associated with TD thresholds. Thus, we were not able to establish impaired proprioception as a behavioral correlate of TD dysfunction, although both functions are known to activate an overlapping neural network and appear to be established markers of impaired sensorimotor integration, a core component in the pathophysiology of dystonia. Future studies will need to elucidate further aspects of proprioception in CD, especially in the neck region, e.g. by application of video-based methods. Understanding the interplay of TD, proprioception and clinical phenotype will ultimately pave the way for a comprehensive pathophysiological model of dystonia.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- BoNT:
-
Botulinum neurotoxin
- CD:
-
Cervical dystonia
- DBS:
-
Deep brain stimulation
- HC:
-
Healthy controls
- ISI:
-
Interstimulus intervals
- STDT:
-
Somatosensory temporal discrimation threshold
- TD:
-
Temporal discrimination
- TDMT:
-
Temporal discrimination movement threshold
- TWSTRS:
-
Toronto Western Spasmodic Torticollis Rating Scale
References
Albanese, A. et al. Phenomenology and classification of dystonia: a consensus update. Mov. Disord. 28 (7), 863–873 (2013).
Monaghan, R. et al. Non-motor features of cervical dystonia: Cognition, social cognition, psychological distress and quality of life. Clin. Park Relat. Disord. 4, 100084 (2021).
Adam, O. R. & Jankovic, J. Treatment of dystonia. Parkinsonism Relat. Disord. 13 (Suppl 3), S362–S368 (2007).
Bledsoe, I. O., Viser, A. C. & San Luciano, M. Treatment of dystonia: Medications, Neurotoxins, Neuromodulation, and rehabilitation. Neurotherapeutics 17 (4), 1622–1644 (2020).
Volkmann, J. et al. Pallidal neurostimulation in patients with medication-refractory cervical dystonia: a randomised, sham-controlled trial. Lancet Neurol. 13 (9), 875–884 (2014).
Odorfer, T. M. & Volkmann, J. Deep brain stimulation for focal or segmental craniocervical dystonia in patients who have failed botulinum neurotoxin Therapy-A narrative review of the literature. Toxins (Basel) 15 (10) (2023).
Schirinzi, T. et al. Dystonia as a network disorder: a concept in evolution. Curr. Opin. Neurol. 31 (4), 498–503 (2018).
Battistella, G. & Simonyan, K. Clinical implications of dystonia as a neural network disorder. Adv. Neurobiol. 31, 223–240 (2023).
Kroneberg, D. et al. Motor cortical plasticity relates to symptom severity and clinical benefit from deep brain stimulation in cervical dystonia. Neuromodulation 21 (8), 735–740 (2018).
Gilio, F. et al. Short-term cortical plasticity in patients with dystonia: a study with repetitive transcranial magnetic stimulation. Mov. Disord. 22 (10), 1436–1443 (2007).
Hallett, M. Neurophysiology of dystonia: the role of Inhibition. Neurobiol. Dis. 42 (2), 177–184 (2011).
Di Lazzaro, V. et al. Reduced cerebral cortex Inhibition in dystonia: direct evidence in humans. Clin. Neurophysiol. 120 (4), 834–839 (2009).
Abbruzzese, G. & Berardelli, A. Sensorimotor integration in movement disorders. Mov. Disord. 18 (3), 231–240 (2003).
Zittel, S. et al. Normalization of sensorimotor integration by repetitive transcranial magnetic stimulation in cervical dystonia. J. Neurol. 262 (8), 1883–1889 (2015).
Conte, A. et al. Temporal discrimination: mechanisms and relevance to Adult-Onset dystonia. Front. Neurol. 8, 625 (2017).
De Bartolo, M. I. et al. Botulinum toxin effects on sensorimotor integration in focal dystonias. Toxins (Basel) 12 (5) (2020).
Erro, R. et al. Reversal of Temporal discrimination in cervical dystonia after Low-Frequency sensory stimulation. Mov. Disord. 36 (3), 761–766 (2021).
Junker, J. et al. Temporal discrimination threshold and Blink reflex recovery cycle in cervical dystonia - two sides of the same coin? Parkinsonism Relat. Disord. 68, 4–7 (2019).
Kagi, G. et al. Sensory tricks in primary cervical dystonia depend on visuotactile Temporal discrimination. Mov. Disord. 28 (3), 356–361 (2013).
Conte, A. et al. Abnormal sensory gating in patients with different types of focal dystonias. Mov. Disord. 33 (12), 1910–1917 (2018).
Scontrini, A. et al. Somatosensory Temporal discrimination in patients with primary focal dystonia. J. Neurol. Neurosurg. Psychiatry. 80 (12), 1315–1319 (2009).
Odorfer, T. M., Wind, T. & Zeller, D. Temporal discrimination thresholds and proprioceptive performance: impact of age and nerve conduction. Front. Neurosci. 13, 1241 (2019).
Avanzino, L. & Fiorio, M. Proprioceptive dysfunction in focal dystonia: from experimental evidence to rehabilitation strategies. Front. Hum. Neurosci. 8, 1000 (2014).
Bove, M. et al. Neck proprioception and Spatial orientation in cervical dystonia. Brain 127 (Pt 12), 2764–2778 (2004).
Consky, E. et al. The Toronto Western spasmodic torticollis rating scale (TWSRTS): assessment of validity and inter-rater reliability. Neurology 40, 445 (1990).
Yilmaz, K. et al. Comparison of head posture and neck proprioceptive sense of individuals with chronic neck pain and healthy controls: A cross-sectional study. J. Back Musculoskelet. Rehabil. 37 (6), 1705–1713 (2024).
Cerina, V. et al. Cervical proprioception assessed through targeted head repositioning: validation of a clinical test based on optoelectronic measures. Brain Sci. 13 (4) (2023).
Cruz, J. & Morais, N. Intrarater agreement of elbow extension range of motion in the upper limb neurodynamic test 1 using a smartphone application. Arch. Phys. Med. Rehabil. 97 (11), 1880–1886 (2016).
Furness, J. et al. Reliability and concurrent validity of the iPhone((R)) compass application to measure thoracic rotation range of motion (ROM) in healthy participants. PeerJ 6, e4431 (2018).
Tousignant-Laflamme, Y. et al. Reliability and criterion validity of two applications of the iPhone to measure cervical range of motion in healthy participants. J. Neuroeng. Rehabil. 10 (1), 69 (2013).
Sanger, T. D., Tarsy, D. & Pascual-Leone, A. Abnormalities of Spatial and Temporal sensory discrimination in writer’s cramp. Mov. Disord. 16 (1), 94–99 (2001).
Tinazzi, M. et al. Temporal discrimination of somesthetic stimuli is impaired in dystonic patients. Neuroreport 10 (7), 1547–1550 (1999).
Tinazzi, M. et al. Deficits of Temporal discrimination in dystonia are independent from the Spatial distance between the loci of tactile stimulation. Mov. Disord. 17 (2), 333–338 (2002).
Tinazzi, M. et al. The role of somatosensory feedback in dystonia: a psychophysical [correction of psycophysical] evaluation. Neurol. Sci. 23 (Suppl 2), S113–S114 (2002).
Fiorio, M. et al. Temporal processing of visuotactile and tactile stimuli in writer’s cramp. Ann. Neurol. 53 (5), 630–635 (2003).
Tinazzi, M. et al. Timing of tactile and visuo-tactile events is impaired in patients with cervical dystonia. J. Neurol. 251 (1), 85–90 (2004).
Fiorio, M. et al. Tactile Temporal discrimination in patients with blepharospasm. J. Neurol. Neurosurg. Psychiatry. 79 (7), 796–798 (2008).
Fiorio, M. et al. Defective Temporal processing of sensory stimuli in DYT1 mutation carriers: a new endophenotype of dystonia? Brain 130 (Pt 1), 134–142 (2007).
Scontrini, A. et al. Somatosensory Temporal discrimination tested in patients receiving botulinum toxin injection for cervical dystonia. Mov. Disord. 26 (4), 742–746 (2011).
Sadnicka, A. et al. Pallidal stimulation for cervical dystonia does not correct abnormal Temporal discrimination. Mov. Disord. 28 (13), 1874–1877 (2013).
Conte, A. et al. Does the somatosensory Temporal discrimination threshold change over time in focal dystonia? Neural Plast. 2017, p9848070 (2017).
Morgante, F. et al. Abnormal tactile Temporal discrimination in psychogenic dystonia. Neurology 77 (12), 1191–1197 (2011).
Tinazzi, M. et al. Temporal discrimination of two passive movements in humans: a new psychophysical approach to assessing kinaesthesia. Exp. Brain Res. 166 (2), 184–189 (2005).
Tinazzi, M. et al. Temporal discrimination of two passive movements in writer’s cramp. Mov. Disord. 21 (8), 1131–1135 (2006).
Tinazzi, M. et al. Temporal discrimination in patients with dystonia and tremor and patients with essential tremor. Neurology 80 (1), 76–84 (2013).
Pastor, M. A. et al. The functional neuroanatomy of Temporal discrimination. J. Neurosci. 24 (10), 2585–2591 (2004).
Redgrave, P. et al. Interactions between the midbrain superior colliculus and the basal ganglia. Front. Neuroanat. 4 (2010).
Hutchinson, M. et al. Cervical dystonia: a disorder of the midbrain network for Covert attentional orienting. Front. Neurol. 5, 54 (2014).
Cardin, J. A. Inhibitory interneurons regulate Temporal precision and correlations in cortical circuits. Trends Neurosci. 41 (10), 689–700 (2018).
Etgen, T. et al. Bilateral grey-matter increase in the putamen in primary blepharospasm. J. Neurol. Neurosurg. Psychiatry. 77 (9), 1017–1020 (2006).
Termsarasab, P. et al. Neural correlates of abnormal sensory discrimination in laryngeal dystonia. Neuroimage Clin. 10, 18–26 (2016).
Pastor, M. A. et al. Putaminal activity is related to perceptual certainty. Neuroimage 41 (1), 123–129 (2008).
Sadnicka, A. et al. Mind the gap: Temporal discrimination and dystonia. Eur. J. Neurol. 24 (6), 796–806 (2017).
Kuyper, D. J. et al. Nonmotor manifestations of dystonia: a systematic review. Mov. Disord. 26 (7), 1206–1217 (2011).
Odorfer, T. M. et al. Topological differences and confounders of mental rotation in cervical dystonia and blepharospasm. Sci. Rep. 13 (1), 6026 (2023).
Conte, A. et al. The role of sensory information in the pathophysiology of focal dystonias. Nat. Rev. Neurol. 15 (4), 224–233 (2019).
Avanzino, L. et al. Sensory-motor integration in focal dystonia. Neuropsychologia 79 (Pt B), 288–300 (2015).
Edwards, L. L. et al. Putting the sensory into sensorimotor control: the role of sensorimotor integration in Goal-Directed hand movements after stroke. Front. Integr. Neurosci. 13, 16 (2019).
Sedov, A. et al. The role of pallidum in the neural integrator model of cervical dystonia. Neurobiol. Dis. 125, 45–54 (2019).
Joshi, P. et al. Effects of neck proprioceptive modulation on pallidal network connectivity in dystonia. Int IEEE EMBS Conf Neural Eng, 2023 (2023).
Sedov, A. et al. Proprioceptive modulation of pallidal physiology in cervical dystonia. Mov. Disord. 38 (11), 2094–2102 (2023).
Grunewald, R. A. et al. Idiopathic focal dystonia: a disorder of muscle spindle afferent processing? Brain 120 (Pt 12), 2179–2185 (1997).
Yoneda, Y. et al. Abnormal perception of the tonic vibration reflex in idiopathic focal dystonia. Eur. J. Neurol. 7 (5), 529–533 (2000).
Lekhel, H. et al. Postural responses to vibration of neck muscles in patients with idiopathic torticollis. Brain 120 (Pt 4), 583–591 (1997).
Avanzino, L. et al. Vibro-tactile stimulation of the neck induces head righting in people with cervical dystonia. Parkinsonism Relat. Disord. 132, 107263 (2025).
Buraschi, R. et al. Efficacy of segmental muscle vibration on pain modulation in patients with primary cervical dystonia treated with botulinum Type-A toxin: A protocol for a randomized controlled trial. NeuroSci 6 (2) (2025).
Xu, J. et al. Vibro-tactile stimulation of the neck reduces pain in people with cervical dystonia: a proof-of-concept study. Neurol. Sci. 45 (10), 4847–4856 (2024).
Pelosin, E. et al. Cervical dystonia affects aimed movements of nondystonic segments. Mov. Disord. 24 (13), 1955–1961 (2009).
Marinelli, L. et al. In idiopathic cervical dystonia movement direction is inaccurate when reaching in unusual workspaces. Parkinsonism Relat. Disord. 17 (6), 470–472 (2011).
Martino, D. et al. Temporal processing of perceived body movement in cervical dystonia. Mov. Disord. 30 (7), 1005–1007 (2015).
Agharazi, H. et al. Unraveling the twist: Spatial navigational challenges in cervical dystonia. Mov. Disord. 38 (11), 2116–2121 (2023).
Avanzino, L. et al. Tactile and proprioceptive dysfunction differentiates cervical dystonia with and without tremor. Neurology 94 (6), e639–e650 (2020).
Funding
Open Access funding enabled and organized by Projekt DEAL. This publication was supported by the Open Access Publication Fund of the University of Wuerzburg.
Author information
Authors and Affiliations
Contributions
TO: conceptualization, formal analysis, methodology, project administration, validation, writing- original draft preparation.AJ: formal analysis, investigation, visualization, writing- original draft preparation.JV: resources, supervision, validation, writing – review and editing.DZ: conceptualization, methodology, project administration, resources, supervision, validation.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Odorfer, T.M., Junge, A., Volkmann, J. et al. Temporal discrimination at the neck region is associated with severity of cervical dystonia. Sci Rep 15, 43113 (2025). https://doi.org/10.1038/s41598-025-31139-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-31139-2
