
Abstract
Large cetaceans face several anthropogenic threats. Among these, collisions are a major cause of anthropogenic mortality. Assessing and limiting their impact on populations is essential, as these species play an essential ecological role. All types of vessels, including offshore racing vessels, can collide with cetaceans. When a collision occurs between an offshore racing vessel and a large cetacean, the consequences are severe for both the whale, which is often injured or even killed and the vessel, which can suffer severe damage and be forced to withdraw from the race. Our study aimed to develop an encounter model that takes the characteristics of both cetaceans and racing vessels into account to estimate the number of encounters along vessel routes. The model was applied to three different routes commonly used in offshore racing: the first between Newport, USA and Skagen, Denmark; the second between Dover, England and the Gibraltar Strait; and the third between the Gibraltar Strait and Genoa, Italy. The number of encounters was estimated to be 1.7 for Route 1, 4.1 for Route 2 and 2.6 for Route 3. The model was also used to estimate the impact of routing vessels away from any exclusion zones that may be established in areas of high cetacean abundance. This routing could significantly reduce the number of encounters and offer potential solutions to reduce collisions between cetaceans and all types of vessels. The issue of collisions is becoming increasingly important and requires the development of methods to reduce the number of collisions worldwide.
Similar content being viewed by others


Introduction
Large cetaceans face several anthropogenic threats, including entanglement in fishing gear, noise disturbance, collisions and pollution (e.g.1,2,3). Among these threats, collisions, also known as vessel strikes, have been identified as the primary cause of anthropogenic mortality for fin whales (Balaenoptera physalus), humpback whales (Megaptera novaeangliae) and blue whales (B. musculus;4,5,6), and a major cause of mortality for the other species, with entanglement in fishing gear being the other major cause3,7,8. Large cetaceans are particularly vulnerable to collisions due to their large size, extensive time spent at or close to the surface for, inter alia, resting, breathing, and socialising activities, and their long-distance migratory patterns that cross maritime trade routes9. Collisions with ships are expected to increase due to the increasing number, size and speed of commercial and recreational vessels, but also because several whale populations have increased since the moratorium on commercial whaling. Consequently, it is crucial to assess and limit the impact of collisions on marine megafauna, as these species play an essential ecological role9, contributing nutrient and biomass transfers, carbon sequestration, and promoting primary production (plankton) by releasing excreta to the surface10.
The factors that determine collisions with cetaceans are relatively unknown. One of the key factors is the amount of time the animal spends at or near the surface. This is highly dependent on the species and its daily rhythm. For example, beaked whales spend less than 10% of their time at or near the surface11, while North Atlantic right whales (Eubalaena glacialis) spend around 70% of their time there12. Some whales, such as fin whales, show a change in their diving behaviour at night, with shallower dives and longer distances travelled at night than during the day13,14. Calambokidis et al.13 and Keen et al.14 estimated a vulnerability to collisions twice as high at night as during the day. The age of the animals also seems to influence the risk of collision. Young cetaceans seem to be more vulnerable than adults15. The animals’ response, or lack of response, to the presence of a vessel is also a key factor16. Cetaceans, such as sperm whales and beaked whales, may be less able to react after long periods of diving because they are stretching their physiological capacities to the limit16. Resting animals would show lower reactivity because they are less alert. When they are hunting, feeding, or socialising, cetaceans concentrate on their prey or their conspecifics and not on potential dangers. It is also possible that the animals do not perceive incoming vessels, either because they are silent, because of acoustic masking (natural or man-made, very high ambient noise), or through a phenomenon of noise habituation, in which case the vessel is no longer identified as a threat. While we would expect the animals to flee, some of them, such as dolphins, are more likely to be attracted to vessels, as are young large whales, because they are more curious and less experienced than adults16.
Collisions between vessels and marine megafauna are not always fatal, but the damage is generally severe, and the lethality depends mainly on the size and speed of the vessel17,18,19. At high speed, the impact of the collision is greater, and the risk of blunt trauma higher4,17,18. However, estimating collisions with marine megafauna and their impact on species is challenging16 due to the very low reporting rate of vessel strikes9. Many collisions go unnoticed, especially with larger vessels in bad weather conditions or at night. Animals are sometimes transported on the bulbous bow after a collision and found in ports, but only a few such cases have been recorded, representing a tiny proportion of actual collisions. Animals are generally not found after a collision; they sink, are eaten by scavengers, or drift offshore. If animals drift ashore, they may be found stranded. If the carcasses are fresh, it is sometimes possible to link the animal death to a collision; more often, the animals are too degraded to allow the cause of death to be identified20. The knowledge of collisions is, therefore, limited.
According to the International Whaling Commission (IWC) database21, cetaceans collide with all types of vessels, whether motorised or sailing. Ferries represent the largest proportion of vessels involved in reported collisions, followed by sailing vessels, passenger ships, motor yachts, whale-watching vessels, navy vessels, container ships and cargo ships. Due to the sampling bias of this database, which is based on the voluntary reporting of collisions by vessel crews or passengers, it remains difficult to determine the relative impact of each type of vessel and the total number of collisions.
Collisions with sailboats are poorly documented and certainly underestimated. To date, Ritter22 remains the only study to examine collisions between cetaceans and sailboats. He showed a significant increase in the number of reported collisions between sailboats and whales since 1993. Monohulls were involved in 82.1% of the incidents, catamarans in 10.3% and trimarans in 5.1% (these figures are more representative of the general composition of the leisure boat fleet than of the dangerousness of the different types of vessels). Out of all collisions, 34.2% occurred during offshore racing, with 45.9% involving large whales, 10.8% involving small whales and 3.6% involving dolphins.
Collisions in offshore racing have long been taboo, but there is now a gradual increase in awareness and incidents are more frequently reported, underlining the seriousness of the phenomenon. The impact of a racing vessel colliding with a cetacean poses a double risk. Due to the high speed of the vessels, the collision is often fatal to the animals and can cause costly damage to both the vessels and their crews, stressing the shared need to limit collisions with offshore racing vessels. Unlike large vessels, a collision involving a racing vessel is often recorded, and its location may be known from the vessel’s GPS track. As a result, it should be easier to calibrate the models used to estimate the number of collisions with empirically collected data and then apply the model more widely to shipping to estimate the number of collisions on a global scale.
In addition to being able to estimate the number of collisions, it is also necessary to limit this number. Technological tools exist that could help reduce collisions with cetaceans. These include detection systems using artificial intelligence and thermal cameras, or acoustic scarecrows23. However, the effectiveness of such tools remains unproven and requires further research. At present, routing is probably the most effective method for mitigating collisions. It consists of avoiding areas with high concentrations of cetaceans to minimise encounter risks, either by establishing exclusion zones or by imposing routes or waypoints23. In offshore racing, it is quite easy to route vessels and avoid high-risk areas as much as possible. People involved in offshore racing are aware of marine issues, particularly collisions, as they affect their performance, safety, and media image.
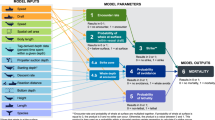
In this study, we aimed to develop an encounter model to estimate the number of encounters between cetaceans and vessels along offshore racing routes. We relied on the model developed by Martin et al.24, which used data on cetacean densities and characteristics of the vessels and species. The model was coupled with weather routing software to predict the routes of the racing vessels depending on their characteristics and weather conditions. We estimated the probability of encountering an animal, i.e. the probability that the animal and the vessel are close enough in time and space to meet each other, rather than the probability of a collision, i.e. the vessel colliding with the animal. The probability that the vessel and the animal avoid each other (the avoidance probability) is complicated to estimate. It appears to be species, age, or individual dependent (e.g.,16,25,26,27,28,29). Similarly, the crew of a sailing vessel may take evasive actions if they detect the presence of a cetacean on the vessel’s trajectory. The likelihood of this detection depends on several factors (vessel speed, sea state, cloud cover, illumination, number of crews on watch, crew experience), and even if detection occurs, the crew may not take evasive action in time to avoid a collision. Therefore, the number of encounters is higher than the number of actual collisions. We then used the encounter model to assess the effect of implementing exclusion zones on the estimated number of encounters. We estimated the effect of routing vessels away from areas known to be hotspots of cetacean density. This routing would offer potential solutions to reduce collisions between cetaceans and all types of vessels.
Results
Regardless of the route considered, the simulated routes depended on the environmental conditions at the start date and in the year considered, and on how these conditions changed along the route. This explains why the simulated routes could differ significantly despite similar starting and finishing points. Two types of vessels were simulated: five vessels of the first type (maximum length of 18 metres and a maximum breadth of 9 metres) on Route 1; four vessels of the first type on Routes 2 and 3; and six vessels of the second type (maximum length of 12 metres and a maximum breadth of 4.5 metres) on Routes 2 and 3.
Route 1 - Newport, USA to Skagen, Denmark
The simulated routes crossed the north-east Atlantic Ocean from Newport, USA, to Skagen, Denmark. Only the first type of vessel was simulated for this route. The routes crossed cetacean concentration areas, particularly east of the USA, and south of Iceland (Fig. 1). Between 60\(^{\circ }\)W and 25\(^{\circ }\)W, the encounter model was not informed by cetacean densities, so the probability of encountering at least one individual was set to zero. In all cases, the encounter probability in the North Sea was low because of the low densities of large cetaceans in this area. In routings 2 to 4, the exclusion zones were defined to avoid the continental shelves off the USA and Canada and the north-west Atlantic Ocean and to progressively avoid the south of Iceland and the north-east of Ireland, as these areas have been identified as high-risk areas with no exclusion zones.
The proposed routing with minimal cetacean encounter probability was routing 4. Instead of bypassing the UK from the north (which is commonly done), this routing required the vessels to navigate the English Channel from west to east. This reduced the probability of encountering at least one individual by 20.6% (from 81.9% to 61.3%) and the mean number of encounters by 0.75 (from 1.7 to 0.95; Table 1) compared to routing 1. Routing 2 slightly decreased, and routing 3 slightly increased the mean number of encounters compared to routing 1, as the routes became increasingly concentrated in the north-west of Ireland, where cetacean densities were the highest.
For most of the proposed routings, based on the cetacean density data, fin and pilot (Globicephala spp.) whales were predicted to be the most likely to be encountered, followed by sperm (Physeter macrocephalus) and minke (Balaenoptera acutorostrata) whales (Table 1). Beaked whales and North Atlantic right whales would be the least likely to be encountered. The North Atlantic right whale would only be encountered in the western North Atlantic Ocean. Routings had different effects on species. The probability of encountering beaked whales increased in routings 2 and 3 but decreased in routing 4 compared to routing 1. For fin and minke whales, the encounter probabilities decreased progressively from routing 1 to 4. For North Atlantic right whales, the encounter probabilities decreased sharply from routings 2 to 4, as the concentration zone in the western Atlantic was avoided. However, there was no difference between these routings. For pilot whales, routings 2 to 4 increased the encounter probability compared to routing 1, and routing 3 even tripled the encounter probability. For sperm whales, the encounter probability was fairly stable between the different routings, although slightly higher for routings 2 to 4.
Route 2 - Dover, England to the Gibraltar Strait
The simulated routes went from Dover, southern England, to the Gibraltar Strait. The two different types of vessels were simulated here. The routes also crossed cetacean concentration areas, particularly in the Bay of Biscay and off Portugal (Fig. 2). Cetacean densities were available throughout the area. All simulated routes crossed the English Channel, where the encounter probability was very low because large cetacean densities are low in this area. The exclusion zones in routings 2 and 3 were identical, but in routing 2, the simulated routes passed mostly to the east of the Bay of Biscay exclusion zone, and in routing 3, they passed to the west.
Regardless of the proposed routing, the probability of encountering at least one individual was estimated to be greater than 95% due to the high density of cetaceans, especially in late spring and early summer in the Bay of Biscay and off the coast of Portugal (Table 2). From the model, the number of encounters was estimated to be between 3.0 and 4.1 individuals. Routing 2 did not change the number of encounters, whereas routing 3 reduced it by 1.1 compared to routing 1, which only included official exclusion zones (e.g., traffic separation schemes, fishing zones, and wind farms).
Based on cetacean density data, there would be more than a 95% chance of encountering at least one pilot whale on all routings, while there would be a 30% chance of encountering at least one beaked whale or sperm whale, and less than a 15% chance of encountering at least one fin and minke whale (Table 2). The probability of encountering beaked whales and sperm whales was the highest in routing 1 and the lowest in routing 2. For minke whales, the probability decreased progressively from routing 1 to 3, while it remained relatively stable for fin whales. For pilot whales, it increased for routing 2 and decreased for routing 3 compared to routing 1.
Route 3 - Gibraltar Strait to Genoa, Italy
The simulated routes in Route 3 went from the Gibraltar Strait to Genoa, Italy (Fig. 3). The two different types of vessels were simulated here. The routes crossed areas with high concentrations of cetaceans in the Alboran Sea (east of the Gibraltar Strait) and in the north-western Mediterranean Sea along the Spanish and French coasts. The encounter probability in the Mediterranean Sea was lower than in the Bay of Biscay, due to lower cetacean densities. Routings 2 and 3 aimed to avoid the centre of the Alboran Sea and the north-western part of the Mediterranean Sea (by enlarging the latter).
The probability of an encounter was high for all three routings (higher than 85%), but was reduced by 4.5% for routing 2 and by 7% for routing 3, compared to routing 1. This resulted in a slight reduction in the mean number of encounters (from 2.6 to 2.0; Table 3).
Based on the cetacean density data, pilot whales would be the species most frequently encountered, followed by fin whales, beaked whales and sperm whales (without exclusion zones; Table 3). For routings 1 to 3, the probability of encountering pilot and fin whales would progressively decrease, with the probability for fin whales falling by almost 73% in routing 3. The probability would remain unchanged for sperm whales and increase for beaked whales.
Discussion
Collisions with vessels are a major cause of death for large cetaceans4,5,6. It is therefore necessary to estimate their impact so that appropriate conservation measures can be taken, in particular, to reduce their frequency. However, this is a complex task because collisions occur offshore and generally go unnoticed. Our study aimed to develop an encounter model that takes the characteristics of both the cetaceans and the offshore racing vessels into account to estimate the number of encounters with cetaceans along the routes taken by the vessels. Our model also allowed us to estimate the effect of routing vessels out of exclusion zones, which can significantly reduce the number of encounters and offer potential solutions for reducing collisions between cetaceans and vessels.
Most studies have implemented the model of Martin et al.24 using vessel densities, based on AIS (Automatic Identification System) data5,14,30,31,32. Here we simulated routes that would be taken by racing vessels based on their characteristics and weather conditions, and thus estimated speeds at each point on the routes. This method has the advantage of being able to deal with vessel-specific routes and reduce the number of encounters based on each vessel’s characteristics.
Cetacean densities must be available on large geographic and temporal scales if a global estimate is to be made. The areas where densities are known are generally limited to those easily accessible by boat or plane, i.e. relatively close to the coast, as surveys are very costly. Densities have been estimated on a large scale on both sides of the Atlantic Ocean33,34,35, but there are still large gaps in the middle of the basin. This is particularly evident in the simulated routes between Newport, USA, and Skagen, Denmark, where densities were not available for a third of the route. Even where surveys were conducted, densities were not available at the same spatial scale (small grid cells for the western and eastern North Atlantic Ocean versus large blocks in the Southern Norwegian Sea) or at all time scales (monthly versus seasonal or annual density predictions), which can greatly bias estimates of the number of encounters, particularly for migratory species. In the north-west and north-east Atlantic Oceans, we used the most recent cetacean data available, estimated by assembling several cetacean surveys. They represent average densities over several years but are available on monthly and seasonal scales. These data need to be regularly updated with new surveys. The density data from Pike et al.36 were quite outdated and only available in blocks, which made it impossible to distinguish between nearby routes because the same densities were applied at each point along the route and between alternative routes nearby. Updated data in the Southern Norwegian Sea would be required.
To develop the model, we set the cetacean length, breadth, density, and surfacing time to species-specific nominal values corresponding to average values found in the literature or estimated from species distribution models. Swimming speed was set to 0 as it was considered negligible compared to vessel speed. The next step in model development will be to incorporate uncertainty in these parameters to account for cetacean behaviour and morphological diversity. The aim will be to construct probability distributions for all parameters and incorporate them into the encounter model. Concerning animal density, the estimate is imperfect and cannot reflect reality in real time. To account for the uncertainty in the densities, constructing appropriate stochastic models with temporal and spatial correlation structures will be required to assess their impact on the estimation of the number of encounters. The final objective will be to derive confidence intervals for the estimated number of encounters that aggregate all sources of uncertainty.
In this study, we fitted an encounter model rather than a collision model because the probabilities of the vessel avoiding the animal and the animal avoiding the vessel were not considered. An encounter results in a collision if the cetacean is in the impact zone (top of the water column) and if neither the cetacean nor the vessel can avoid the collision. Rockwood et al.5 defined the number of collisions as the product of the encounter rate, the probability of the cetacean being in the impact zone, and the probability of not avoiding the collision. The probability of the cetacean being in the impact zone depends largely on the time the animal spends at the surface and the draught of the vessel. The probability of avoidance is very difficult to estimate for either the vessel or the cetacean. For large vessels, the probability of avoidance is often assumed to be zero because the vessels are not very manoeuvrable. The faster the vessel, the more difficult it is to avoid a collision. For cetaceans, the probability of avoidance has not yet been estimated and depends on the characteristics of the vessel, the species, the age, and even the individual16. Rockwood et al.5 have parametrised three scenarios: a decrease in avoidance as vessel speed increases, a constant avoidance of 55% (based on the study of McKenna et al.27) and no avoidance. They showed that without avoidance, the mortality rate was twice as high as with speed-dependent avoidance and an avoidance rate of 55%. Keen et al.14 have proposed a theoretical estimate of the avoidance probability for fin whales. They showed that the risk of collision was influenced by the seasonal variation in the length of the night, the spatial distribution of shipping, the higher rate of traffic at night, the seasonal dynamics and the diurnal patterns of habitats used by cetaceans.
It is also difficult to estimate the probability of avoidance associated with the vessel. This depends on many parameters such as the manoeuvrability, size, and speed of the vessel, but also on the experience and willingness of the captain (probably the most difficult parameter to estimate), the reaction time after detecting an animal, the distance from the animal and whether an alternative route is available. Gende et al.37 have developed a model aimed at reducing collisions through active avoidance of whales, i.e. based on the captain’s operational decisions following the detection of a whale. They used a navigation simulator in which a whale was modelled. They showed that, on average, 23 seconds elapsed between the moment the whale was detected and the command to act. Uncertainty about the whale’s direction of swimming and its speed were the criteria that increased reaction time. If the whale was not detected early enough, the possible actions were very limited. To turn away from the whale by 100 m, a large cruise ship would have to travel 741 m at 10 knots and 1,121 m at 19 knots in simplified and optimal conditions. Knowing the distance, swimming direction, and speed of the whale as soon as it is detected would reduce the latency before taking action. However, we would need to know how the whale behaves underwater, whether it continues a straight course or changes direction. We aim to incorporate the probability of avoidance into future development.
Most studies on collisions focus solely on the impact on cetaceans. To implement mitigation measures, particularly on a voluntary basis, it is necessary to estimate the impact of collisions on vessels and the costs of those collisions and mitigation measures. For example, a change of route could result in additional fuel costs or an arrival delay, which must be considered. Sèbe et al.38 have tried to consider the damage caused to vessels in collisions. They showed that the probability of damage increased with the speed/length ratio of the vessels, but the dataset was too poor to make more detailed estimates. Sèbe et al.39 also assessed the monetary cost of avoiding a whale using an International Maritime Organization tool, the Formal Safety Assessment, which is a rational and systematic process for assessing the risks associated with maritime safety and environmental protection and estimating the costs and benefits of reducing those risks. Taking into account the abundance of the species, its carrying capacity and the estimated population value (based on the estimated willingness to pay per person to conserve the last whale population and the number of inhabitants, as well as an estimated relationship between the willingness to pay and the distance from the site), they estimated the value of avoiding the death of one fin whale in the Mediterranean Sea at US$562,500.
Our ultimate goal is to implement a cost model linked to the encounter model, which would allow the estimation of both the potential financial impact of a collision and the cost of a collision to biodiversity. As mentioned by Cook et al.40 or Sheehy et al.41, the ecosystem services provided by cetaceans are numerous (e.g., provision of food, climate regulation through carbon sequestration, cultural value with whale watching). Consequently, on the one hand, the loss of carbon storage and transformation or the amount of nitrogen recycled through cetacean feeding and excretion, associated with the death of a cetacean due to collision, must be estimated. On the other hand, it is necessary to estimate the cost of damage to the vessel in the event of a collision. This will require data collection. Finally, it should be possible to estimate the benefits of avoiding collisions for both vessels and cetaceans.
Although certainly much smaller than maritime traffic in general, the impact of offshore racing on biodiversity is not negligible as vessels and races become more numerous, larger, and faster. In our study, we estimated an average of 8.4 encounters with cetaceans per year for the 15 simulated vessels across three routes. Given the number of races and training sessions organised, particularly in the North Atlantic Ocean, this suggests that the scale of the collision problem in offshore racing is underestimated. Indeed, Ritter22 identified only 38 collisions with cetaceans during regattas between 1966 and 2010, while, for example, nine collisions with unidentified floating objects (most often animals) were reported during the 2016 edition of the Vendée Globe round-the-world race (collisions with unidentified floating objects retrieved from the https://www.vendeeglobe.org/ website). Skippers are encouraged to report collisions to assess the problem better. Given the results of this study, the implementation of large exclusion zones could help to reduce the number of encounters and thus collisions.
There are currently no general estimates of cetacean mortality associated with offshore racing and, importantly, few countries have regulations requiring teams to report collisions or measures to ensure that organisers of offshore races implement procedures that route vessels through areas with low concentrations of cetaceans. The IWC has a vessel strike strategy which identifies high risk areas and considers potential measures for each of these areas42. Systematic reporting of collisions would allow a general assessment of the impact. Identification of the species struck would allow the identification of the species most vulnerable to collisions, depending on the area crossed by the routes and the racing seasons. We plan to apply our encounter model to future races where cetacean densities are available and compare our estimates with skipper-reported encounters. In offshore racing, it is also possible to route vessels and avoid high-risk areas where possible. People involved in offshore racing are also aware of marine issues and can be directly involved in limiting collisions as they affect their performance, safety and outreach strategies. For all these reasons, offshore racing is a perfect demonstrator for calibrating the encounter model and, we hope, subsequently extending its application to maritime transport in general.
From a species conservation perspective, our encounter model can be used in a risk assessment process to balance the risks associated with shipping43. By establishing exclusion zones, we estimated a reduction in the probability of encountering at least one animal. This was particularly true for Route 1, and less so for Routes 2 and 3, because in the latter two cases, the general route made it difficult to avoid areas where cetaceans are concentrated. Used as part of an overall risk assessment, this method would make it possible to propose specific routes for the vessels, considering their characteristics and the constraints they face, such as ports of call for cargo ships, while trying to reduce the impact on marine megafauna. In this study, our focus was primarily on large cetaceans due to their high vulnerability to collisions4,5,6. However, propeller-driven vessels and sailing vessels may pose a different level of risk to cetaceans. In particular, little data exists on collision risks involving sailing vessels using foils, as these vessels are a relatively recent development. It is generally assumed that small cetacean species take more effective evasive action with regard to oncoming vessels than larger whales, which is reflected in the greater number of reports of large whales being struck. Nevertheless, it is plausible that no cetacean species would be aware of an approaching, high-speed, foiling vessel, meaning any animal at the surface in its path would be unable to take evasive action. Small cetaceans, such as dolphins and porpoises, may be as vulnerable to being struck by high-speed foiling vessels as large whales. These species may therefore need to be included in future risk assessments for offshore racing vessels. As for the 75 species of the marine megafauna identified by Schoeman et al.23, these include small cetaceans, turtles and sunfish (Mola mola), which are also affected by collisions. These species are less studied, probably because their densities are less known, making it difficult to estimate the impact of collisions. Improving knowledge of species distribution is a key step in improving conservation measures for marine megafauna.
Methods
This section provides an overview of the model, parameters, and analyses carried out in the present work.
Encounter model
The present paper relies on a model based on the encounter rate theory44. This theory describes the encounter rate between spatially and temporally mobile agents, considering their abundance, size, speed, and interaction zone31. Applications in environmental sciences include prey-predator interactions, mate search, and pollination. Martin et al.24 used this theory to quantify the risk of collision between vessels and marine mammals. Rockwood et al.5 have extended this model to incorporate vessel characteristics and whale behaviour to quantify collision risk better. Our model was derived from Martin et al.24. In this class of models, the number \(E_\chi\) of encounters associated with a trajectory \(\chi = \{ X(t), t\in [0, T]\}\) follows a Poisson distribution. We denote \(\Lambda _\chi\) the parameter of the Poisson distribution; it corresponds to the expected number of encounters. This section describes the calculation of \(\Lambda _\chi\) and the assumptions involved.
Kinematic model
At a given point along the trajectory, we denote the angle between the vessel (v) and the cetacean (c) routes by \(\phi\), and by \(v_v\) and \(v_c\) the vessel and cetacean velocities. The relative velocity is then \(w(\phi ) = \sqrt{v_v^2 + v_c^2 - 2 v_v v_c \cos \phi }\). Without any prior information about the cetacean’s direction of travel, \(\phi\) is assumed to be uniformly distributed in \([0,2\pi [\), resulting in an averaged relative velocity:
Unlike the vessel velocity, which is known from the trajectory \(\chi\), the cetacean velocity is unknown and assumed to be random with a probability distribution \(f_{V_c}\). Averaging \(V_{re}\) against this distribution gives the mean encounter velocity (function of the vessel velocity only):
Encounter rate
The area covered by a vessel travelling at speed \(v_v\) with an encounter radius \(r_{en}\) in an infinitesimal time interval \(\delta t\) is \(2\, r_{en}\, V_{en}(v_v)\, \delta t\). The encounter radius was originally defined by Martin et al.24 as the sum of the vessel and cetacean encounter radii, \(r_{en} = r_v + r_c\), where the projected area of the vessel or cetacean is assumed to be a disc. So \(\pi r^2 = length*breadth\) and \(r = \sqrt{\frac{length*breadth}{\pi }}\). In our applications to offshore racing boats (this would be incorrect for longer vessels such as cargo ships), we assumed that forward encounters were more likely to occur than side-on encounters since the vessel’s speed is significantly higher than that of the cetaceans. We also assumed a symmetry in the collision, since generally only one side of the boat strikes the animal. Therefore, we based the encounter radius \(r_v\) on the vessel’s breadth \(B_v\) rather than its length and a constant \(c_v\) set to \(c_v={\frac{1}{2\sqrt{\pi }}}\), using the following formula:
For the cetacean encounter radius, one could let \(r_c\) be a function of \(\phi\) since cetaceans are larger in the swimming direction than in the transverse direction. However, accounting for an anisotropic cross-section would lead to a \(\phi\)-dependent encounter radius, which would require averaging the product \(r_{en}(\theta ) w(\phi )\) instead of just the relative velocity in equation (1).
For convenience, we thus considered a fixed encounter radius \(r_c\) taken as:
where \(L_c\) and \(B_c\) are the average length and breadth of the cetaceans.
The effective encounter rate \(\lambda _{en} (x,v_v)\) is finally obtained as:
Where \(0 < T_{surf} \le 1\) is the fraction of time the cetaceans spend close to the sea surface, and \(\delta (x)\) is the cetacean density (number of individuals per unit area).
Expected number of encounters
Assuming the cetacean’s velocity distribution and encounter radius are constant in space and time, and a time-independent density, the expected number of encounters along the trajectory \(\chi\) is:
where \(|dX /dt| = v_v(t)\) is the velocity of the vessel at time t. To better understand this expression, one can consider its slow cetacean/fast vessel limit. When the vessel velocity along the trajectory is significantly higher than the maximal swimming velocity, one can set \(v_c=0\) in the expressions above (that is \(f_{V_c}(v_c)\) is the Dirac mass at \(v_c=0\)). This hypothesis is validated by Glennie et al.45, who have shown through simulations that the biases associated with animal speed are strongly reduced when the speed of the observer (in this case, the vessel) is much greater than the speed of the animal. In this limit, \(w(\phi ) = V_{re} = V_{en}= v_v\), and (6) becomes:
which is \((2 \, T_{surf} \, r_{en})\) times the integral of the population’s density, \(\delta (x)\), along the path of the vessel.
In practice, we discretise the trajectory into a sequence of n successive locations \(X_{i=1,\dots ,n}\) reached at (increasing) times \(t_{i=1,\dots ,n}\). Assuming a sufficiently refined discretisation of \(\chi\), the integral (6) is approximated by:
Finally, the probability of having at least one encounter along \(\chi\) is:
Several populations
The encounter model can also be generalised in the case of \(N_p\) distinct populations of cetaceans, computing for each population an average number of encounters \(\Lambda _{\chi ,p}\) from its characteristics (\(\delta _p, f_{V_c,p}, r_{c,p}, T_{surf,p}\)). The compounded mean number of encounters and probability of at least one encounter along \(\chi\) become:
Putting the model into practice
We used the encounter model to estimate the encounter probability and mean number of encounters along three routes that correspond to previously used race routes. The first route was a transatlantic race from Newport, USA, to Skagen, Denmark. The second route was from the north to the south of Europe, passing through the English Channel and the Bay of Biscay before reaching the Gibraltar Strait. The third route crossed the north-western Mediterranean Sea from the Gibraltar Strait to Genoa, Italy.
Vessel parameters and trajectories
In this study, we simulated two types of vessels, both of which are based on classic offshore racing boats. The first type has a maximum length of 18 metres and a maximum breadth of 9 metres, including appendages. Five vessels were simulated on Route 1 each year, and four on each of the other two routes. The second type has a maximum length of 12 metres and a maximum breadth of 4.5 metres. No vessels of this type were simulated on Route 1. Six vessels were simulated on Routes 2 and 3 each year. We applied the encounter model to routes that were simulated with and without exclusion zones. Encounter predictive assessments were performed here, so the trajectories had to be determined before running the encounter model.
The trajectories of sailing vessels depend primarily on the weather conditions, which affect the routes and the vessel speeds. For a given route, we collected meteorological datasets from 2008 to 2022 to account for meteorological variability. For each year, five datasets were collected to simulate five different start dates within the same month (one day’s delay). The weather data (wind speed, height, and direction) were downloaded from the ERA5 model in the Copernicus Marine Environment Monitoring Service repository (https://cds.climate.copernicus.eu/cdsapp# !/dataset/reanalysis-era5-single-levels?tab=overview) at an hourly resolution on 0.25\(^{\circ }\) or 0.5\(^{\circ }\) resolution grids (depending on the variable).
The qtVlm navigation software (©Meltemus 2017 - 2024, https://www.meltemus.com/) was then used to compute the optimal trajectory that minimises the sailing times to the finish with and without exclusion zones. Exclusion zones affect vessel trajectories. They are related to skipper safety (ice zones) and regulatory requirements (navigation restrictions in high-traffic areas). We also implemented several exclusion zones related to the cetacean presence to assess the reduction of encounters by routing vessels away from areas with high concentrations of cetaceans. For a given vessel type, the optimal trajectory depends on the weather conditions and the sailing characteristics of the vessel. The latter consists of the vessel’s polars, which are look-up tables that predict the vessel speed as a function of the wind speed and angles from the route to the wind. These polar data were simulated for each vessel class and are confidential. Finally, for each vessel and each scenario (three or four scenarios with different cetacean exclusion zones), 75 trajectories were simulated (5 trajectories over 15 years) with one location every 3,600 seconds. For each vessel trajectory, the probability of encountering at least one animal and the mean number of encounters per species and all species (for which density data were available) were estimated at each point on the trajectory and over the entire trajectory. Finally, for each vessel type, we calculated the average encounter probability and the mean number of encounters over all years and extracted the lowest and highest values estimated.
Cetacean densities and characteristics
To run the encounter model, we need an estimate of the cetacean densities along the routes followed by the vessels. To do this, we commonly use species distribution models (SDMs) to relate species observations to characteristics of the environment46. In a previous study, Virgili et al.35, we used SDMs to predict the seasonal distributions and densities of the most common cetacean species in the Northeast Atlantic Ocean and the Mediterranean Sea. We assembled 16 years (from 2005 to 2021) of aerial and shipboard cetacean surveys provided by various organisations. We ran generalised additive models using environmental variables integrated over the water column. Grids of densities (in individuals/km\(^{2}\)) at a spatial resolution of 0.1\(^{\circ }\)x0.1\(^{\circ }\) were used as input data in the encounter model. The grids extended from 35\(^{\circ }\)N to 70\(^{\circ }\)N and from 20\(^{\circ }\)W to 10\(^{\circ }\)E in the Northeast Atlantic Ocean and throughout the Mediterranean Sea (from 34\(^{\circ }\)N to 45\(^{\circ }\)N and from 6\(^{\circ }\)W to 40\(^{\circ }\)E). These prediction grids were used for Routes 2 and 3 and combined with prediction grids provided by other organisations for Route 1.
Indeed, the prediction grids extracted from Virgili et al.35 only covered the Northeast Atlantic Ocean, while Route 1 crossed the entire North Atlantic Ocean. Unfortunately, no density data were available in the centre of the North Atlantic Ocean, but we requested density data in the Southern Norwegian Sea from Pike et al.36 and in the Northwest Atlantic Ocean33,47. In Pike et al.36, cetacean densities were estimated from the Trans-North Atlantic Sightings Survey (T-NASS) carried out in 2007 by using stratified line-transect distance sampling methods. Detection functions were fitted with Hazard Rate and Half Normal key functions, and covariates affecting detection were taken into account. Cetacean densities in individuals/km\(^{2}\) were provided in large blocks. Roberts et al.47 compiled 23 years (from 1992 to 2014) of aerial and shipboard cetacean surveys in the US Atlantic and the Gulf of Mexico and fitted SDMs (generalised additive models) with environmental variables to predict monthly distributions and densities (in individuals/100 km \(^{2}\)) of different cetacean species. Prediction grids at a spatial resolution of 10x10 km were downloaded from the web portal https://seamap.env.duke.edu/models/Duke/EC/. All grids were projected to WGS84.
In this study, we focused on six cetacean species or species groups for which densities were available: fin whales, North Atlantic right whales, sperm whales, beaked whales, pilot whales, and minke whales. Table 4 summarises the characteristics of the six cetacean species used in the encounter model. Lengths, breadths and surfacing times were taken from the literature. These values were used in the initial development of the model, but do not represent the full range of values recorded for these species. Future developments will allow for the integration of these ranges. As explained above, the cetacean swimming speed was set to 0. We considered the mean values of density, length, breadth, and surfacing time for each cetacean species to estimate the number of encounters using the encounter model.

Probability of encountering at least one individual (in %) estimated for all weather routings (2008-2022) along Route 1 between Newport, USA, and Skagen, Denmark, considering different exclusion zones. The scale corresponds to the probability of encountering at least one animal for each vessel location (every 3,600 seconds). The red-hatched polygons represent exclusion zones proposed to reduce the number of encounters along the route. The red cross corresponds to a waypoint, the vessels had to go round to the right. Only the first type of vessel was simulated.

Probability of encountering at least one individual (in %) estimated for all weather routings (2008-2022) along Route 2 between Dover, England and the Gibraltar Strait, considering different exclusion zones. The scale corresponds to the probability of encountering at least one animal for each vessel location (every 3,600 seconds). The red-hatched polygons represent the exclusion zones proposed to reduce the number of encounters along the route. The two types of vessels were simulated.

Probability of encountering at least one individual (in %) estimated for all weather routings (2008-2022) along Route 3 between the Gibraltar Strait to Genoa, Italy, considering different exclusion zones. The scale corresponds to the probability of encountering at least one animal for each vessel location (every 3,600 seconds). The red-hatched polygons represent the exclusion zones proposed to reduce the number of encounters along the route. The two types of vessels were simulated.
Data availability
Vessel tracks were simulated using the qtVlm navigation software (©Meltemus 2017 - 2024, https://www.meltemus.com/), which is an open-access software, but the polars used were provided by Bañulsdesign and are confidential. Cetacean densities in the western Atlantic Ocean are publicly available at https://seamap.env.duke.edu/models/Duke/EC/. Cetacean densities near Iceland are not publicly available and should be requested from the North Atlantic Marine Mammal Commission (NAMMCO). Cetacean densities in the Northeast Atlantic Ocean are not publicly available and should be requested from the Direction Générale de l’Armement Techniques Navales. The corresponding author can provide contact details.
References
Blair, H. B., Merchant, N. D., Friedlaender, A. S., Wiley, D. N. & Parks, S. E. Evidence for ship noise impacts on humpback whale foraging behaviour. Biol. Lett. 12, 20160005. https://doi.org/10.1098/rsbl.2016.0005 (2016).
Unger, B. et al. Large amounts of marine debris found in sperm whales stranded along the North Sea coast in early 2016. Mar. Pollut. Bull. 112, 134–141. https://doi.org/10.1016/j.marpolbul.2016.08.027 (2016).
Moore, M. J. How we can all stop killing whales: a proposal to avoid whale entanglement in fishing gear. ICES J. Mar. Sci. 76, 781–786. https://doi.org/10.1093/icesjms/fsy194 (2019).
Conn, P. B. & Silber, G. K. Vessel speed restrictions reduce risk of collision-related mortality for North Atlantic right whales. Ecosphere 4, art43, https://doi.org/10.1890/ES13-00004.1 (2013).
Rockwood, R. C., Calambokidis, J. & Jahncke, J. High mortality of blue, humpback and fin whales from modeling of vessel collisions on the U.S. west coast suggests population impacts and insufficient protection. PloS one 12, e0183052. https://doi.org/10.1371/journal.pone.0183052 (2017).
Sèbe, M. et al. Estimating the impact of ship strikes on the Mediterranean fin whale subpopulation. Ocean Coast. Manag. 237, 106485. https://doi.org/10.1016/j.ocecoaman.2023.106485 (2023).
Read, A. J., Drinker, P. & Northridge, S. Bycatch of marine mammals in U.S. and global fisheries. Conserv. Biol. 20, 163–169, https://doi.org/10.1111/j.1523-1739.2006.00338.x (2006).
Tulloch, V. et al. Long-term trends and a risk analysis of cetacean entanglements and bycatch in fisheries gear in Australian waters. Biodivers. Conserv. 29, 251–282. https://doi.org/10.1007/s10531-019-01881-x (2020).
Pirotta, V., Grech, A., Jonsen, I. D., Laurance, W. F. & Harcourt, R. G. Consequences of global shipping traffic for marine giants. Front. Ecol. Environ. 17, 39–47. https://doi.org/10.1002/fee.1987 (2019).
Doughty, C. E. et al. Global nutrient transport in a world of giants. Proc. Natl. Acad. Sci. 113, 868–873. https://doi.org/10.1073/pnas.1502549112 (2016).
Wiggins, S. M., Thayre, B. J., Trickey, J. S., Baumann-Pickering, S. & Hildebrand, J. A. Beaked whale passive acoustic tracking offshore of Cape Hatteras 2017. Tech. Rep., Marine Physical Laboratory, Scripps Institution of Oceanography, University of California San Diego, La Jolla, CA, MPL Technical Memorandum 631 October 2018. Submitted to Naval Facilities Engineering Command (NAVFAC) Atlantic, Norfolk, Virginia, under Contract No. N62470-15-D-8006 Subcontract 383-8476 (MSA2015-1176 Task Order 003) issued to HDR, Inc. (2018).
Baumgartner, M. F., Wenzel, F. W., Lysiak, N. S. & Patrician, M. R. North Atlantic right whale foraging ecology and its role in human-caused mortality. Mar. Ecol. Prog. Ser. 581, 165–181. https://doi.org/10.3354/meps12315 (2017).
Calambokidis, J. et al. Differential vulnerability to ship strikes between day and night for blue, fin, and humpback whales based on dive and movement data from medium duration archival tags. Front. Mar. Sci. 6, https://doi.org/10.3389/fmars.2019.00543 (2019).
Keen, E. M. et al. Night and day: Diel differences in ship strike risk for fin whales (Balaenoptera physalus) in the California current system. Front. Mar. Sci. 6, https://doi.org/10.3389/fmars.2019.00730 (2019).
Smith, J. N. et al. Quantifying ship strike risk to breeding whales in a multiple-use marine park: the Great Barrier Reef. Front. Mar. Sci. 7, 67. https://doi.org/10.3389/fmars.2020.00067 (2020).
Ritter, F. & Panigada, S. Chapter 28 - Collisions of vessels with cetaceans – The underestimated threat. In Sheppard, C. (ed.) World Seas: An Environmental Evaluation (Second Edition), 531–547, https://doi.org/10.1016/B978-0-12-805052-1.00026-7 (Academic Press, 2019), second edition edn.
Laist, D. W., Knowlton, A. R., Mead, J. G., Collet, A. S. & Podesta, M. Collisions between ships and whales. Mar. Mammal Sci. 17, 35–75. https://doi.org/10.1111/j.1748-7692.2001.tb00980.x (2001).
Vanderlaan, A. S. M. & Taggart, C. T. Vessel collisions with whales: the probability of injury based on vessel speed. Mar. Mammal Sci. 23, 144–156. https://doi.org/10.1111/j.1748-7692.2006.00098.x (2007).
Garrison, L. P. et al. The effects of vessel speed and size on the lethality of strikes of large whales in us waters. Front. Mar. Sci. 11, 1467387. https://doi.org/10.3389/fmars.2024.1467387 (2025).
Peltier, H. et al. Monitoring of marine mammal strandings along French coasts reveals the importance of ship strikes on large cetaceans: A challenge for the European marine strategy framework directive. Front. Mar. Sci. 6, https://doi.org/10.3389/fmars.2019.00486 (2019).
Winkler, C., Panigada, S., Murphy, S. & Ritter, F. Global numbers of ship strikes: An assessment of collisions between vessels and cetaceans using available data in the IWC ship strike database. IWC B (2020).
Ritter, F. Collisions of sailing vessels with cetaceans worldwide: First insights into a seemingly growing problem. J. Cetacean Res. Manage. 12, 119–127, https://doi.org/10.47536/jcrm.v12i1.598 (2012).
Schoeman, R. P., Patterson-Abrolat, C. & Plön, S. A global review of vessel collisions with marine animals. Front. Mar. Sci. 7, https://doi.org/10.3389/fmars.2020.00292 (2020).
Martin, J. et al. A quantitative framework for investigating risk of deadly collisions between marine wildlife and boats. Methods Ecol. Evol. 7, 42–50. https://doi.org/10.1111/2041-210X.12447 (2015).
Nowacek, D. P., Johnson, M. P. & Tyack, P. L. North Atlantic right whales (Eubalaena glacialis) ignore ships but respond to alerting stimuli. Proc. Royal Soc. London. Ser. B: Biol. Sci. 271, 227–231, https://doi.org/10.1098/rspb.2003.2570 (2004).
Allen, J. K., Peterson, M. L., Sharrard, G. V., Wright, D. L. & Todd, S. K. Radiated noise from commercial ships in the Gulf of Maine: Implications for whale/vessel collisions. J. Acoust. Soc. Am. 132, EL229–35, https://doi.org/10.1121/1.4739251 (2012).
McKenna, M., Calambokidis, J., Oleson, E., Laist, D. & Goldbogen, J. Simultaneous tracking of blue whales and large ships demonstrates limited behavioral responses for avoiding collision. Endangered Species Res. 27, 219–232. https://doi.org/10.3354/esr00666 (2015).
Argüelles, M. B., Coscarella, M., Fiorito, C. & Bertellotti, M. Southern right whales generally appear not to react to transiting research vessels. Mar. Mammal Sci. 38, 6–17. https://doi.org/10.1111/mms.12843 (2022).
Martin, M. J. et al. Exposure and behavioural responses of tagged bowhead whales (Balaena mysticetus) to vessels in the Pacific Arctic. Arctic Science https://doi.org/10.1139/as-2022-0052 (2023).
Crum, N., Gowan, T., Krzystan, A. & Martin, J. Quantifying risk of whale-vessel collisions across space, time, and management policies. Ecosphere 10, e02713. https://doi.org/10.1002/ecs2.2713 (2019).
Udell, B. J. et al. Integrating encounter theory with decision analysis to evaluate collision risk and determine optimal protection zones for wildlife. J. Appl. Ecol. 56, 1050–1062. https://doi.org/10.1111/1365-2664.13290 (2019).
Redfern, J. V., Becker, E. A. & Moore, T. J. Effects of variability in ship traffic and whale distributions on the risk of ships striking whales. Front. Mar. Sci. 6, https://doi.org/10.3389/fmars.2019.00793 (2020).
Roberts, J. J. et al. Habitat-based cetacean density models for the US Atlantic and Gulf of Mexico. Sci. Rep. 6, 22615. https://doi.org/10.1038/srep22615 (2016).
Waggitt, J. J. et al. Distribution maps of cetacean and seabird populations in the North–East Atlantic. J. Appl. Ecol. 57, 253–269. https://doi.org/10.1111/1365-2664.13525 (2020).
Virgili, A. et al. Seasonal distribution of cetaceans in the European Atlantic and Mediterranean waters. Front. Mar. Sci. 11, 1319791. https://doi.org/10.3389/fmars.2024.1319791 (2024).
Pike, D. G. et al. Estimates of the abundance of cetaceans in the Central North Atlantic from the T-NASS Icelandic and Faroese ship surveys conducted in 2007. NAMMCO Sci. Publ. 11, https://doi.org/10.7557/3.5269 (2019).
Gende, S. M. et al. Active whale avoidance by large ships: Components and constraints of a complementary approach to reducing ship strike risk. Front. Mar. Sci. 6, https://doi.org/10.3389/fmars.2019.00592 (2019).
Sèbe, M., Kontovas, C. A. & Pendleton, L. Reducing whale-ship collisions by better estimating damages to ships. Sci. Total. Environ. 713, 136643. https://doi.org/10.1016/j.scitotenv.2020.136643 (2020).
Sèbe, M., Kontovas, C. A., Pendleton, L. & Gourguet, S. Cost-effectiveness of measures to reduce ship strikes: A case study on protecting the Mediterranean fin whale. Sci. Total. Environ. 827, 154236. https://doi.org/10.1016/j.scitotenv.2022.154236 (2022).
Cook, D., Malinauskaite, L., Davísdóttir, B., Ögmundardóttir, H. & Roman, J. Reflections on the ecosystem services of whales and valuing their contribution to human well-being. Ocean. & Coast. Manag. 186, 105100, https://doi.org/10.1016/j.ocecoaman.2020.105100 (2020).
Sheehy, J. M. et al. Review of evaluation and valuation methods for cetacean regulation and maintenance ecosystem services with the Joint Cetacean Protocol data. Front. Mar. Sci. 9, https://doi.org/10.3389/fmars.2022.872679 (2022).
Cates, K. et al. Strategic plan to mitigate the impacts of ship strikes on cetacean populations: 2017–2020 (International Whaling Commission, In IWC Strategic Plan to Mitigate Ship Strikes (Jersey, 2017).
Domene, X., Ramírez, W., Mattana, S., Alcañiz, J. M. & Andrés, P. Ecological risk assessment of organic waste amendments using the species sensitivity distribution from a soil organisms test battery. Environ. Pollut. 155, 227–236. https://doi.org/10.1016/j.envpol.2007.12.001 (2008).
Koopman, B. O. The theory of search I. Kinematic bases. Oper. research 4, 324–346 (1956).
Glennie, R., Buckland, S. T. & Thomas, L. The effect of animal movement on line transect estimates of abundance. PloS one 10, e0121333. https://doi.org/10.1371/journal.pone.0121333 (2015).
Elith, J. & Leathwick, J. R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 40, 677–697. https://doi.org/10.1146/annurev.ecolsys.110308.120159 (2009).
Roberts, J., Yack, T. & Halpin, P. Marine mammal density models for the U.S. navy Atlantic Fleet Training and Testing (AFTT) study area for the phase iv Navy Marine Species Density Database (NMSDD). document version 1.3. Tech. Rep., Report prepared for Naval Facilities Engineering Systems Command, Atlantic by the Duke University Marine Geospatial Ecology Lab, Durham, North Carolina (2023).
Barlow, J., Griffiths, E. T., Klinck, H. & Harris, D. V. Diving behavior of Cuvier’s beaked whales inferred from three-dimensional acoustic localization and tracking using a nested array of drifting hydrophone recorders. J. Acoust. Soc. Am. 144, 2030–2041. https://doi.org/10.1121/1.5055216 (2018).
Würsig, B., Thewissen, J. & Kovacs, K. M. (eds.) Encyclopedia of Marine Mammals (Third Edition) (Elsevier, 2018), academic press edn.
Laran, S. et al. A comprehensive survey of pelagic megafauna: their distribution, densities, and taxonomic richness in the tropical Southwest Indian Ocean. Front. Mar. Sci. 4, 139. https://doi.org/10.3389/fmars.2017.00139 (2017).
Heide-Jørgensen, M. et al. Satellite tracking of minke whales (Balaenoptera acutorostrata) off the coast of northern Norway. J. Cetacean Res. Manage. 3, 175–178 https://doi.org/10.47536/jcrm.v3i2.887 (2001).
Friedlaender, A. et al. Feeding rates and under-ice foraging strategies of the smallest lunge filter feeder, the Antarctic minke whale (Balaenoptera bonaerensis). J. Exp. Biol. 217, 2851–2854. https://doi.org/10.1242/jeb.106682 (2014).
Hain, J. H., Hampp, J. D., McKenney, S. A., Albert, J. A. & Kenney, R. D. Swim speed, behavior, and movement of North Atlantic right whales (Eubalaena glacialis) in coastal waters of northeastern Florida, USA. PloS one 8, e54340. https://doi.org/10.1371/journal.pone.0054340 (2013).
Baird, R. W., Borsani, J. F., Hanson, M. B. & Tyack, P. L. Diving and night-time behavior of long-finned pilot whales in the Ligurian Sea. Mar. Ecol. Prog. Ser. 237, 301–305. https://doi.org/10.3354/meps237301 (2002).
Irvine, L., Palacios, D. M., Urbán, J. & Mate, B. Sperm whale dive behavior characteristics derived from intermediate-duration archival tag data. Ecol. Evol. 7, 7822–7837. https://doi.org/10.1002/ece3.3322 (2017).
Acknowledgements
We are grateful to the many observers who participated in the surveys and collected all the data used to model cetacean densities, as well as to ship captains, crews, and pilots. We thank the Direction Générale de l’Armement Techniques Navales for providing cetacean densities in the Northeast Atlantic Ocean and the Mediterranean Sea. We thank the Duke Marine Geospatial Ecology Laboratory for providing cetacean densities in the Northwest Atlantic Ocean through their web portal (https://seamap.env.duke.edu/models/Duke/EC/). We thank the North Atlantic Marine Mammal Commission (NAMMCO) for providing cetacean densities estimated from the NASS survey.
Funding
The study and open access were funded by Share The Ocean.
Author information
Authors and Affiliations
Contributions
A.V. wrote the main manuscript. A.V., S.F. and M.P. carried out the analyses. V.R., O.L.M. and R.B. contributed their expertise to the study, and R.B. supervised it. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Virgili, A., Fournier, S., Le Maître, O. et al. Assessing cetacean encounter risk in offshore racing. Sci Rep 16, 3921 (2026). https://doi.org/10.1038/s41598-025-33896-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-33896-6
