
Abstract
Soil nutrients and meteorological conditions are pivotal environmental factors influencing plant growth and development. This study systematically analyzes how soil nutrients and meteorological factors influence the phenotypic growth and seed production of wild Elymus nutans in Tibet. These environmental factors are critical ecological determinants, and this research seeks to unveil the complex and diverse ecological adaptation mechanisms of the species. The research encompasses ten counties within the Tibet Autonomous Region, with sampling points distributed across seven distinct ecological environments. By evaluating root soil nutrient levels, collecting meteorological data, and analyzing growth characteristics and seed elemental composition at various sampling points, the study assesses the ecological adaptability of Elymus nutans under diverse soil nutrient and meteorological conditions. Cluster analysis categorized the 25 samples of Elymus nutans into four distinct groups, each exhibiting significant differences in height and leaf length. Correlation analysis revealed a significant negative correlation between sampling point elevation and NH₄⁺, soil total phosphorus, and available phosphorus content, while a significant positive correlation was observed with pH levels. Meteorological variables—including temperature, humidity, and precipitation—significantly influenced plant phenotypes and the elemental composition of seeds. Soil ammonium nitrogen, total phosphorus, and available phosphorus are vital nutrients for the growth of Elymus nutans, and their deficiency has a significant impact on plant phenotypes and overall growth. Environments characterized by high altitude, aridity, and intense radiation constrain plant growth. Climatic conditions influence the growth of Elymus nutans by altering the soil environment, where soil quality plays a critical role in determining growth conditions. These factors interact to collectively impact the ecosystem.
Similar content being viewed by others


Introduction
Soil nutrients and meteorological conditions play critical roles in shaping plant growth and development1,2. These factors have direct effects on plant growth characteristics and indirect impacts on plant reproduction and population dynamics through their influence on seed elemental content and formation3,4. The unique geographical location and climatic features of the Tibetan Plateau make it a crucial area for investigating the interplay between plants and the environment5,6. Elymus nutans, a crucial perennial forage grass extensively found in the Tibetan grasslands, boasts outstanding nutritional value and ecological adaptability. As global climate change accelerates and human activities intensify, grassland ecosystems are confronting escalating pressures7,8. Hence, gaining insights into the adaptive mechanisms and growth performance of Elymus nutans under varying environmental conditions holds paramount importance for the conservation and governance of grassland ecosystems.
The Tibetan Plateau region experiences significant heterogeneity in soil nutrient status and meteorological conditions9,10. The spatial diversity of soil nutrients, combined with the seasonal and yearly fluctuations of meteorological factors, exerts intricate effects on plant growth and development11,12. Essential soil nutrients like nitrogen, phosphorus, and potassium have direct impacts on plant photosynthesis, root development, and biomass13,14, whereas meteorological factors such as temperature, precipitation, and light indirectly govern plant growth traits by influencing physiological processes and metabolic activities15,16. Therefore, investigating the influence of environmental factors on the growth and seed production of Elymus nutans carries substantial scientific importance. Previous research has concentrated on the impact of artificial nutrient supplementation on the ecological adaptability and growth traits of Elymus nutans17,18. Nonetheless, there remains a scarcity of studies that conduct a comprehensive analysis of the interplay between soil nutrient levels, meteorological conditions, growth phenotypes, and seed elemental composition under field conditions. Inadequate soil nutrients have been shown to markedly decrease the growth rate and biomass of Elymus nutans, whereas the addition of artificial nutrients contributes to enhancing its productivity and nutritional content19. Furthermore, variations in meteorological factors like temperature and precipitation exert a substantial influence on the growth season and reproductive cycle of Elymus nutans20,21. Nonetheless, these studies predominantly concentrate on analyzing the influence of individual factors, lacking in-depth investigations into the combined effects of multiple factors on Elymus nutans’ growth and development. This study aims to systematically analyze the influence of soil nutrients and meteorological factors, which are crucial environmental determinants, on the growth phenotype and seed production of wild Elymus nutans in Tibet. This analysis aims to unveil the complex and diverse ecological adaptation mechanisms of the species. Through measuring and examining the nutrient levels in the root soil of Elymus nutans at various sampling locations, along with meteorological data, growth characteristics, and seed elemental composition, we aim to assess the ecological adaptability of Elymus nutans under diverse soil nutrient and meteorological settings. This analysis will help identify environmental factors that restrict the growth and development of wild Elymus nutans on the Tibetan Plateau.
Materials and methods
Study area overview
The research focuses on 10 counties within the Tibet Autonomous Region of China, situated under the cities of Lhasa, Nagqu, Shigatse, Shannan, and Nyingchi. Sampling points are distributed across seven distinct ecological environments: mountainsides, roadsides, alpine grasslands, alpine meadows, alpine meadow grasslands, foothills, and farmland.
Sampling collection and analysis
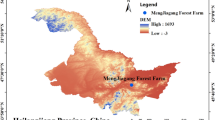
Based on the varied altitude gradients in Tibet ranging from 3015.3 to 4964.0 m and the distribution patterns of wild Elymus nutans, 25 representative sample points were established in diverse habitats located between 28°83′ to 31°85′N and 87°92′ to 94°34′E. During the selection of sample points, priority was given to Elymus nutans exhibiting well-groomed growth and forming large patches. Figure 1 displays all the sample points.

Distribution map of 25 sampling points of Elymus nutans.
Collection and analysis of plant samples
In September 2023, phenotypic data of fully mature Elymus nutans plants were assessed at multiple locations. At each location, a total of 10 plants were selected for sampling. The selection of plant samples was conducted using a randomized sampling method to ensure unbiased representation across the study sites. Mature seeds were also randomly sampled from each plant. The plant characteristics measured included the following: Height (absolute length from plant base to spike top, cm), Ear length (EL, length from spike base to top, cm), Stem length under ear (SLUE, length from base to first internode, cm), Length of Flag leaf (LFL, length of first leaf below spike, mm), Width of Flag leaf (LFW, maximum width of first leaf below spike, mm), Length of seed (LS, length of seed at its longest point, excluding awn length, mm), awn length (AL, mm), and Weight per Thousand Seeds (WTS, determined by the thousand-grain method and averaged over 10 repetitions, g)22,23.
Soil sample collection and analysis
Plant phenotypes were assessed at each sampling point, and seeds were subsequently collected. Rhizosphere soil samples from Elymus nutans were collected and transported to the laboratory for drying. Subsequently, the soil underwent grinding and sieving through a 1 mm sieve to analyze soil nutrients and pH levels. Soil analysis included the determination of various parameters: total nitrogen (soil-TN, %) by the Kjeldahl method, nitrate nitrogen (NO3−, mg/kg) by ultraviolet spectrophotometry, ammonium nitrogen (NH4+, mg/kg) by indophenol blue colorimetry, total phosphorus (soil-TP, g/kg) and available phosphorus (soil-QP, mg/kg) by the molybdenum antimony colorimetric method, total potassium (soil-TK,%) and available potassium (soil-QK, g/kg) by atomic absorption spectrophotometry, organic carbon (soil-OC, %) using the potassium dichromate method (with external heating), and pH by the potentiometric method24,25.
Seed sample collection and analysis
Following the drying of the 25 collected Elymus nutans seed samples in the laboratory, the elemental composition of the seeds was analyzed. The elemental composition of the seeds was analyzed as follows: total nitrogen (seed-TN, g/kg) using a German SEAL Auto Analyzer3 continuous flow analyzer, total phosphorus (seed-TP, g/kg) using flame photometry, total potassium (seed-TK, g/kg) using atomic absorption spectrophotometry, organic carbon (seed-OC, g/kg) through the potassium dichromate volumetric method (with additional heating), total calcium (seed-Ca, g/kg) and total magnesium (seed-Mg, g/kg) by atomic absorption spectrophotometry, and total sulfur (seed-S, g/kg) using the HNO3-HCLO4 digestion method26,27,28.
Seed germination test design
The experiment employed the paper towel germination method. Filter paper soaked in distilled water was placed in a Petri dish with 50 seeds evenly distributed in each dish, constituting one replicate. This procedure was repeated three times for each treatment. The Petri dishes were then placed in a constant temperature incubator at 25℃. Seed germination progress was monitored daily, with distilled water being added as necessary. The artificial climate incubator provided light intensity at 1250 lx, operating on a 12-hour light-dark cycle. Germination was defined as the point when either the root or shoot length equaled or exceeded the predetermined standard length. After 12 days, the number of germinated seeds was tallied to determine the germination rate (G, %). The root length (RL, mm) and shoot length (BL, mm) of the germinated seeds were assessed using a ruler29.
Meteorological data collection and analysis
Meteorological data were obtained from NASA30, covering the period from January 1, 2023, to August 31, 2023. The data analysis encompassed various meteorological parameters: average daily temperature (DAT, ℃), average daily humidity (ADH, %), average daily pressure (ADP, hPa), total precipitation (TW, mm), longitudinal wind speed (EW, m/s), latitudinal wind speed (NW, m/s), terrestrial surface radiation (TSR, W/m²), normal direct irradiance (NDI, W/m²), and scattered radiation (SR, W/m²).
Data analysis
Data were organized using Excel. analysis of variance and stepwise regression analysis were conducted with SPSS31. Correlation analysis, principal component analysis, and cluster analysis were performed using Origin202432. The R language was utilized for Mantel test analysis33 and structural equation modeling (SEM) analysis34.
Results
Descriptive statistics on the phenotype, seed elemental content, and germination indicators of Elymus nutans
Table 1 illustrates the wide range of phenotypic characteristics exhibited by 25 samples of Elymus nutans. Characteristics such as Height, SLUE, LFL, LFW, and AL show high coefficients of variation, ranging from 31.51 to 46.37%. In contrast, EL, LS, and WTS exhibit lower coefficients of variation, ranging from 8.83 to 20.43%. Significant variation is present in the seed content of the 25 Elymus nutans samples, with seed-OC showing a minimal coefficient of variation of 2.87%. In comparison, seed-TN, seed-Mg, and seed-S exhibit relatively lower coefficients of variation, ranging from 14.87 to 18.55%, while seed-TP, seed-TK, and seed-Ca have coefficients of variation exceeding 20%, ranging from 21.50 to 23.72%.
The germination indices of the 25 Elymus nutans seeds present significant variation, with G showing the highest coefficient of variation at 59%, and RL and BL during the seed germination stage exhibiting coefficients of variation of 27.24% and 24.02% respectively. Seed element accumulation and germination indices show a diverse range of variations, offering valuable guidance for seed selection and propagation post-breeding.
Cluster results of 25 samples of Elymus nutans
Cluster analysis of 25 samples of Elymus nutans, as illustrated in Fig. 2A, was performed to uncover patterns in phenotypic traits. The analysis classified the samples into four distinct categories based on Plant phenotype: Class I consists of seven samples characterized by lower WTS, LFW, AL, LS, and EL. Class II comprises a single unique sample with lower height and higher SLUE and LFL. Class III, the largest group with sixteen samples, exhibits lower SLUE, LFL, and height. Class IV also includes a single sample, marked by distinctly higher SLUE, LFL, and height. These groupings underscore the genetic and phenotypic diversity crucial for understanding resource management and adaptability in Elymus nutans.

Cluster analysis results of 25 Elymus nutans (A). Analysis of the correlation between altitude, longitude, latitude, and environmental factors (B).
Correlation analysis between sampling points and environmental factors
According to Fig. 2B, a significant negative correlation exists between the elevation of sampling points and NH4+, soil-TP, and soil-QP content (P < 0.01), along with a significant positive correlation with pH value (P < 0.01). Additionally, the elevation demonstrates significant negative correlations with DAT, ADH, ADP, and TW among the meteorological factors (P < 0.01), a significant negative correlation with NW (P < 0.05), and significantly positive correlations with NDI (P < 0.05) and EW (P < 0.01).
The longitude exhibits significant or very significant positive relationships with NH4+, soil-QP, and total phosphorus in soil nutrients (P < 0.05 or P < 0.01), while showing a significant negative correlation with pH value (P < 0.01). In terms of meteorological factors, longitude is significantly positively correlated with ADP (P < 0.05), very significantly correlated with ADH, TW, and SR (P < 0.01), and very significantly negatively correlated with EW, TSR, and NDI (P < 0.01).
Latitude exhibits a significant negative correlation with soil-TP in soil nutrients (P < 0.01) and a significant positive correlation with pH value (P < 0.01). Moreover, latitude demonstrates very significant negative correlations with DAT, TW, and NW among the meteorological factors (P < 0.01), as well as a very significant positive correlation with EW (P < 0.01).
Correlation analysis between environmental factors and phenotype of Elymus nutans
Analysis of Fig. 3A reveals that the sampling points of Elymus nutans exhibit significant negative correlations in Height, SLUE, and LFL with altitude (P < 0.01), supported by respective R2 fitting equations of 0.76, 0.61, and 0.65. Furthermore, Height and LS show significant positive correlations with longitude (P < 0.05), each with R2 fitting equations of 0.16 and 0.14; SLUE and LFL also demonstrate significant positive correlations with longitude (P < 0.01), accompanied by R2 fitting equations of 0.35 and 0.34. In addition, Height displays a significant negative correlation with latitude (P < 0.01), with an R2 fitting equation of 0.33; SLUE exhibits a significant negative correlation with latitude (P < 0.05), supported by an R2 fitting equation of 0.15. No significant correlations were observed between other phenotypes and altitude, longitude, or latitude. EL and WTS exhibit positive correlations with Height (P < 0.05), indicated by R2 fitting equations of 0.14 and 0.15, respectively; while SLUE and LFL demonstrate significant positive correlations with Height (P < 0.01), supported by R2 fitting equations of 0.61 and 0.48, respectively. Moreover, LFL shows a positive correlation with EL (P < 0.05), with an R2 fitting equation of 0.17; LFW is significantly positively correlated with EL (P < 0.01), supported by an R2 fitting equation of 0.59. Additionally, LFW and WTS show positive correlations with SLUE (P < 0.05), with R2 fitting equations of 0.12 and 0.13; while LFL and LS demonstrate significant positive correlations with SLUE (P < 0.01), supported by R2 fitting equations of 0.67 and 0.26. Lastly, LFL exhibits a significant positive correlation with LFW (P < 0.01), with an R2 fitting equation of 0.33. No significant correlations were identified between the remaining phenotypes.
Examination of Fig. 3B indicates that Height displays significant positive correlations with NH4+ and soil-TP content (P < 0.05), and significant negative correlation with pH (P < 0.01); Height also exhibits a negative correlation with EW (P < 0.05) and positive correlations with DAT, ADP, TW, and NW at significant (P < 0.05) or highly significant levels (P < 0.01); furthermore, Height is significantly (P < 0.05) or highly significantly (P < 0.01) positively associated with seed-TP and seed-TK content. EL demonstrates no significant correlations with soil nutrients, meteorological factors, and seed-TK content, except for a highly significant positive correlation with seed-TK content (P < 0.01). SLUE is highly positively correlated with NH4+, significantly correlated with soil-TP content (P < 0.05), and highly negatively correlated with pH (P < 0.01); SLUE also displays significant (P < 0.05) or highly significant (P < 0.01) positive correlations with DAT, ADH, TW from meteorological factors, and negative correlations with EW, NDI (P < 0.01), while showing a negative correlation with seed-Ca content (P < 0.05). LFL is significantly (P < 0.05) or highly significantly (P < 0.01) positively correlated with NH4+, total phosphorus, and soil-QP content, highly negatively correlated with pH (P < 0.01); LFL shows significant (P < 0.05) or highly significant (P < 0.01) positive correlations with DAT, ADH, ADP, TW, SR from meteorological factors, and negative correlations with EW, TSR, NDI (P < 0.05); LFL is also negatively correlated with seed-Ca content (P < 0.05). LFW demonstrates no significant correlations with soil nutrients and meteorological factors but is significantly positively correlated with seed-TK accumulation (P < 0.05). LS and Seed AL present no significant correlations with soil nutrients, meteorological factors, and seed content. WTS shows no significant correlations with soil nutrients and meteorological factors but exhibits a highly significant positive correlation with seed-TK accumulation (P < 0.05).

Analysis of the correlation matrix of phenotypic traits of Elymus nutans (A). Analysis of the Correlation Between Phenotypic Traits, Environmental Factors, and Seed Elements (B).
Correlation analysis between environmental factors and seed element content of Elymus nutans
In Fig. 4A, it is evident that the Elymus nutans seed-TP content exhibits a significant negative correlation with altitude (P < 0.01), whereas the seed-Ca content shows a significant positive correlation with altitude (P < 0.05). Additionally, the seed-TP content demonstrates a significant negative correlation with latitude (P < 0.05). Furthermore, the seed-TN content is significantly positively correlated with seed-OC accumulation (P < 0.05), while the seed-Ca content shows a significant negative correlation with seed-Ca content (P < 0.05). Moreover, the seed-TP content is significantly positively correlated with seed-TK content (P < 0.05). Notably, among the seed germination indicators, BL shows a significant positive correlation with both seed-TN and seed-TP content (P < 0.05).
Figure 4B demonstrates that the Elymus nutans seed nitrogen content (seed-TN) exhibits a significant positive correlation with soil total phosphorus content (soil-TP) (P < 0.05), and the seed-TN content is significantly positively correlated with soil quick phosphorus content (soil-QP) (P < 0.01). Furthermore, the seed total phosphorus content (seed-TP) shows a significant positive correlation with NH4 and soil-TP content (P < 0.05), but a negative correlation with the pH value (P < 0.05). Additionally, the seed-TP content is significantly positively correlated with NW (P < 0.01) and negatively correlated with EW (P < 0.05). The seed total potassium content (seed-TK) is significantly positively correlated with NW in meteorological factors (P < 0.05). The seed organic carbon (seed-OC) content demonstrates a significant negative correlation with soil quick potassium content (soil-QK) (P < 0.05). Moreover, the seed calcium (seed-Ca) content is significantly negatively correlated with TW (P < 0.05) and positively correlated with EW (P < 0.05). Additionally, the seed sulfur (seed-S) content displays a significant positive correlation with DAT (P < 0.05). No other significant correlations were observed between seed element content and soil nutrients or meteorological factors.

Correlation matrix analysis of seed elemental content in Elymus nutans (A). Correlation analysis between seed elemental content, environmental factors (B).
Multivariate stepwise regression analysis of environmental factors with Elymus nutans height and WTS
Height and WTS serve as crucial indicators of plant growth and reproduction, impacting the stability of population systems, survival success, reproductive rate, genetic diversity, and adaptability. By investigating the influence of these two indicators on population stability, we can further delve into the dynamic changes within ecosystems and the conservation of biodiversity. This study adopts the Height and WTS of Elymus nutans as dependent variables, while considering soil nutrients, meteorological factors, and seed element content as independent variables, utilizing stepwise regression to determine the relationship equation.
Relationship between soil nutrients and Elymus nutans height
Utilizing the stepwise regression method, the relationship Eq. (1) linking Height to soil nutrients is formulated as follows:
The main soil nutrients influencing the Height of Elymus nutans according to Eq. (1) are NO3−, soil-TN, soil-TP, soil-QP, soil-TK, soil-QK, soil-OC, and pH, with NH4+ having a minimal impact. However, due to the standard error test, the regression equation failed to meet the significance level threshold (P < 0.05), suggesting that the impact of soil nutrients on Height as indicated in Eq. (1) is merely indicative. This discrepancy could potentially stem from substantial climate variations across different sampling locations, implying that the influence of soil nutrients on the Height of wild Elymus nutans in Tibet did not achieve statistical significance (P > 0.05).
Relationship between meteorological factors and Elymus nutans height
Utilizing the stepwise regression method, the relationship Eq. (2) linking Height to meteorological factors is formulated as follows:
The relationship between Height and meteorological factors is strongly interconnected, with Eq. (2) displaying a highly significant correlation (P < 0.01). Equation (2) illustrates that all meteorological factors influence the Height of Elymus nutans. Additionally, DAT, NW, and TSR exhibit significant (P < 0.05) or highly significant (P < 0.01) partial correlations with Height. The correlation coefficient in Eq. (2) is 0.9403**, signifying that meteorological factors have a highly significant (P < 0.01) impact on the Height of Elymus nutans in Tibet.
Relationship between soil nutrients and Elymus nutans WTS
Utilizing the stepwise regression method, the relationship Eq. (3) linking WTS to soil nutrients is formulated as follows:
Equation (3) suggests that all soil nutrients have the potential to impact Elymus nutans WTS. However, based on the standard error test, the regression equation failed to meet the significance level criterion (P < 0.05). This could be attributed to the predominant influence of Elymus nutans’ genetic factors on its WTS, suggesting that the effect of soil nutrients on the WTS of wild Elymus nutans in Tibet did not achieve statistical significance (P > 0.05).
Relationship between meteorological factors and Elymus nutans WTS
Utilizing the stepwise regression method, the relationship Eq. (4) linking WTS to meteorological factors is formulated as follows:
Equation (4) indicates that all meteorological factors may influence Elymus nutans WTS. However, the partial correlation coefficients of these factors with WTS, as shown in Eq. (4), did not demonstrate a statistically significant level of correlation. Despite the lack of significance in the impact of meteorological factors on WTS (P < 0.05), these variables significantly influence the height of wild Elymus nutans in Tibet, and a notable correlation exists between height and WTS. Consequently, further analysis is required to investigate the indirect effect of meteorological factors on WTS.
Relationship between seed element content and Elymus nutans WTS
Utilizing the stepwise regression method, the relationship Eq. (5) linking WTS to seed element content is formulated as follows:
Equation (5) reveals that seed-TN, seed-TP, seed-TK, seed-OC, seed-Mg, and seed-S are the primary seed elements influencing the WTS of Elymus nutans, while Ca has only a minimal impact. However, based on the standard error test, the regression equation failed to meet the significance level criterion (P < 0.05), and none of the seed element contents displayed a significant partial correlation coefficient with WTS (P < 0.05). This implies that the influence of seed element contents from wild Elymus nutans in Tibet on WTS did not achieve statistical significance (P < 0.05).
Correlation between phenotype, seed elemental content, and environmental factors in Elymus nutans
Figure 5 illustrates that Mantel test analysis identified a significant positive correlation (P < 0.01) between the phenotype of Elymus nutans plants and NH₄⁺ in soil nutrients, as well as significant positive correlations (P < 0.01) with pH values, and a notable positive correlation (P < 0.05) with soil-TP and soil-QP content. Moreover, Pearson correlation analysis revealed a significant negative correlation (P < 0.01) between NH₄⁺, soil-TP, and soil-QP in soil and pH values. The soil pH plays a crucial role in influencing various biochemical processes and nutrient availability. Typically, a higher pH level indicates a less acidic environment, potentially facilitating the absorption and accessibility of specific nutrients, thereby enhancing plant growth and phenotypic performance. The study indicates that a higher nutrient content in the soil correlates with an elevated pH value, where NH₄⁺, soil-TP, and soil-QP exhibit greater stability in low pH conditions, leading to a negative correlation with pH values.
The Mantel test revealed a significant positive correlation between the phenotype of Elymus nutans and meteorological factors including DAT, ADH, ADP, and TSR (P < 0.05), as well as a highly positive correlation with TW, EW, NDI, and SR (P < 0.01). Additionally, Pearson correlation analysis indicated a high positive correlation between DAT and ADP (P < 0.01); ADH was highly positively correlated with ADP, TW, and SR (P < 0.01), while showing a negative correlation with EW, TSR, and NDI (P < 0.01). Furthermore, ADP exhibited a significant negative correlation with EW (P < 0.01); TW was strongly positively correlated with NW and SR (P < 0.01), but negatively correlated with EW, TSR, and NDI (P < 0.01); EW displayed a strong negative correlation with NW and SR (P < 0.01), and a positive correlation with TSR and NDI (P < 0.01); while TSR was highly positively correlated with NDI (P < 0.01) and negatively correlated with SR (P < 0.01). The meteorological factors DAT, ADH, ADP, and TW all exerted significant positive effects on the phenotype of Elymus nutans, suggesting that appropriate meteorological conditions play a crucial role in enhancing its growth and optimizing phenotype. Moreover, the study revealed a positive correlation between three types of radiation and the phenotype of Elymus nutans, highlighting the significance of light conditions as a vital meteorological factor influencing the growth and expression of Elymus nutans phenotype.
The Mantel test results indicated a lack of significant correlation between the seed elemental content of Elymus nutans and both soil nutrients and meteorological factors. This lack of correlation could be attributed to the specific mechanisms of element accumulation, the intricate nature of environmental factors, cumulative effects, potential measurement errors, and other unmeasured influencing factors. These findings suggest that the seed elemental content is influenced by a multitude of factors, indicating a complex interplay beyond the immediate influence of soil and weather conditions.

Mantel’s test of the plant phenotype, seed elemental content, and environmental factors of Elymus nutans.
Principal component analysis of phenotype, soil nutrients, meteorological factors, and seed element content in Elymus nutans
Examination of Fig. 6A reveals that in the first principal component of the phenotype of Elymus nutans, Height, EL, SLUE, LFL, and LFW exhibit relatively high loadings with positive characteristic vector values, while AL shows a negative characteristic vector value. Additionally, AL demonstrates a negative correlation with Height, EL, SLUE, LFL, and LFW. In the second principal component, LS and AL demonstrate relatively high loadings with positive characteristic vector values, whereas EL and LFW exhibit negative characteristic vector values, showing a negative correlation with AL.
Analysis of Fig. 6B reveals that in the first principal component of the seed elemental content in Elymus nutans, seed-TN, seed-TP, and seed-OC display high loadings with positive characteristic vector values, whereas seed-Ca and seed-Mg demonstrate negative characteristic vector values. Specifically, seed-Ca shows a negative correlation with seed-TN, seed-TP, seed-TK, seed-OC, and seed-S, while seed-Mg is negatively correlated with seed-TN and seed-OC. In the second principal component, seed-TK, seed-Mg, and seed-TP exhibit high loadings with positive characteristic vector values, while seed-TN and seed-OC have negative characteristic vector values. Additionally, seed-TN and seed-OC are inversely correlated with seed-Mg and seed-TK.
Examination of Fig. 6C indicates that in the first principal component of the rhizosphere soil nutrients of Elymus nutans, the characteristic vector values of pH and soil-QK are negative, with pH showing the highest loading. The loadings of soil-TP, soil-QP, and NH4 are relatively high with positive characteristic vector values, while pH and soil-QK exhibit negative correlations with soil-TP, soil-QP, and NH4. In the second principal component, the loadings of soil-TN and soil-OC are relatively high with positive characteristic vector values, whereas the characteristic vector values of soil-QP and NH4 are negative, resulting in negative correlations with soil-TN and soil-OC.
Examination of Fig. 6D reveals that in the first principal component of meteorological factors at sampling points, the loadings of ADH, SR, and TW are relatively high with positive characteristic vector values, while EW, TSR, and NDI exhibit negative characteristic vector values, indicating a negative correlation with ADH, TW, and SR. In the second principal component, DAT and ADP show high loadings with positive characteristic vector values, whereas EW and SR display negative characteristic vector values, and these variables demonstrate a negative correlation with DAT and ADP.

Principal component analysis of the plant phenotype (A) and seed elemental content (B) of Elymus nutans. Principal component analysis of soil nutrients (C) and Principal component analysis of Climate Factors (D).
Relationship among the phenotype of Elymus nutans, soil nutrients, meteorological factors, and seed element content
Figure 7 indicates that elevation and latitude/longitude of sampling points are negatively correlated with plant phenotype, soil nutrients, and meteorological factors, but positively correlated with seed element content. Elevated locations correspond to lower temperatures and air pressure, which may potentially restrict plant photosynthesis and metabolic activity, impacting both plant phenotype and soil microbial activity, leading to a decrease in soil nutrients; seed element accumulation may serve as an adaptation mechanism. Soil nutrients demonstrate positive correlations with plant phenotype and seed element content, since adequate nutrient levels support healthy plant growth and enhance seed element content. Although meteorological factors indicate a negative correlation with plant phenotype, favorable weather conditions contribute to the improvement of soil nutrients and seed element content. Enhanced plant phenotype signifies plants’ enhanced nutrient utilization capacity, thus promoting an increase in seed element content; elevated seed element levels further boost seed germination rate and seedling growth.

Structural equation model analysis of sampling point positions with Elymus nutans plant phenotype, seed elemental content, soil nutrients, and meteorological.
Discussion
The effects of soil nutrients on the phenotype and seed elemental content of Elymus nutans
Soil nutrients serve as the foundation for sustaining plant growth and development35 Soil nitrogen plays a crucial role in influencing plant growth; an adequate nitrogen supply can stimulate plant growth and development, leading to increased biomass and yields. Nitrogen participates in key processes such as protein and chlorophyll synthesis, thereby enhancing plant phenotypic characteristics, including leaf area and plant height36,37,38. NH4+ stands out as one of the primary nitrogen sources essential for plant growth, exerting a significant impact on plant development. An optimal NH4+ concentration can foster root growth and development, enhance nutrient uptake by roots, and facilitate overall plant growth39; however, elevated NH4+ concentrations may impede root development, hamper above-ground plant growth, and suppress plant stature40. In the present study, the demand of Elymus nutans for NH4+ is directly manifested in height, specific leaf uptake efficiency (SLUE), leaf fresh weight per length (LFL) growth, and total phosphorus accumulation in seeds (Fig. 3B), revealing a highly significant (P < 0.01) positive correlation between NH4+ and plant phenotype (Fig. 5). This correlation suggests that the deficiency of NH4+ negatively impacts the growth of Elymus nutans. The concentration of NO3− also influences plant growth41,42; in this study, the NO3− concentration did not directly affect plant phenotype and seed element accumulation in Elymus nutans, possibly due to the non-deficient levels of NO3− content in soil nutrients at the sampling site. Adequate phosphorus is essential for promoting plant root growth, leaf morphological development, influencing flower and fruit growth, nutrient element content in seeds, and the synthesis of fats and proteins, as well as the accumulation of micronutrients43,44. Soil available phosphorus, a form of phosphorus that plants can readily absorb and utilize, directly impacts Elymus nutans’ height, SLUE, LFL growth, seed total nitrogen (seed-TN), and seed-total phosphorus (seed-TP) accumulation in this study. Soil quick phosphorus (soil-QP) affects Elymus nutans by influencing LFL growth and seed-TN accumulation. Additionally, both soil-TP and soil-QP exhibit a highly significant (P < 0.01) positive correlation with plant phenotypes, highlighting the importance of phosphorus as a key nutrient for the growth of Elymus nutans in Tibet, with soil phosphorus content playing a crucial role in the accumulation of seed TN (TP) elements. Maintaining the appropriate soil pH is fundamental for sustaining soil nutrient balance and microbial activity, thereby fostering healthy plant growth45. In this study, pH demonstrates a highly significant (P < 0.01) negative correlation with height, SLUE, LFL, and a significant (P < 0.05) negative correlation with seed total phosphorus content, indicating that a higher pH level limits the growth of Elymus nutans in Tibet. Furthermore, a significant positive correlation between pH and plant phenotype is observed due to the highly significant positive correlation between pH and soil nutrients NH4+, total phosphorus, and soil-QP (P < 0.01). This underscores that despite the enhanced nutrient content in alkaline soil conditions, reduced nutrient availability, compromised plant adaptability, root health issues, and nutritional imbalances resulting from higher pH levels ultimately impede plant growth indicators. This indicates the necessity to maintain a relatively lower pH for grasslands with ample soil nutrients to ensure the normal growth of Elymus nutans46,47. Although the accumulation of seed elements shows a relatively weak correlation with nitrogen, phosphorus, potassium, and other soil elements, soil nutrients exert a positive influence on both plant phenotypes and seed element accumulation. This suggests that soil nutrients indirectly affect seed element accumulation by influencing plant phenotypes. In conclusion, this study reveals that the insufficiency of NH4+, soil-TP, and soil-QP in soil nutrients, coupled with increased pH levels, are the primary factors constraining the growth of Elymus nutans, while the accumulation of seed elements is not directly impacted by soil nutrients. While our study provides important insights into the impact of soil nutrients and pH on Elymus nutans, it is important to acknowledge certain limitations. The study is based on observations from a specific geographic region, which may not fully represent the environmental variability across broader areas. Additionally, the potential confounding effects of unmeasured environmental variables, such as microbial community interactions or specific soil texture aspects, were not explicitly controlled or measured in this study. These factors could introduce biases and limit the generalizability of our findings.
Effects of meteorological factors on the growth and seed formation of Elymus nutans
Meteorological factors play a pivotal role in the growth of plants and seed formation, impacting various aspects of the ecological environment48.DAT is a critical determinant of plant growth season and rate. Different plant species exhibit varying ranges of temperature adaptability, with optimal temperatures enhancing photosynthesis and metabolic activities, while extreme temperature fluctuations can disrupt physiological functions or result in plant mortality49. Variations in DAT directly affect the growth of plant Height, SLUE, LFL, and seed-S content. Changes in air pressure and temperature regulate plant metabolism, growth rate, and water use efficiency by influencing air density, water vapor availability, and airflow patterns50,51. In this study, ADP exhibits a significant positive correlation with ADH (P < 0.01). ADP also influences the growth of SLUE and LFL. The plant phenotype demonstrates a notable positive correlation with DAT and ADP (P < 0.01), suggesting the inhibitory impact of the challenging Tibetan high-altitude environment on the growth of Elymus nutans. Additionally, precipitation directly impacts ADH, underscoring its pivotal role in plant water balance, particularly in transpiration and stomatal regulation52,53. In this study, both TW and ADH have a direct impact on the growth of Height, SLUE, LFL, and LS. The plant phenotype also shows a significant correlation with TW and ADH (P < 0.01), indicating that the low humidity in Tibet’s arid environment significantly constrains the growth of Elymus nutans. The influence of wind on plants extends to transpiration, stomatal regulation, pollination, and seed dispersal54, with a focus on both EW and NW winds. In the Tibetan region, EW wind speed demonstrates a notable positive correlation with Elymus nutans Height, SLUE, LFL, and LS (P < 0.05), while NW winds exhibit a negative correlation with these parameters. Moreover, EW shows a significant positive correlation with the plant phenotype (P < 0.01), whereas NW does not exhibit a correlation due to the beneficial moisture and temperature regulation provided by North-South winds in contrast to the dry air and dust carried by East-West winds55. Radiation plays a crucial role in regulating plant growth by influencing photosynthesis, morphological structure, physiological adaptation, and ecological distribution56. Adequate sunlight radiation generally enhances plant growth and promotes efficient photosynthesis, while insufficient or excessive radiation can trigger corresponding adaptive changes that impact plant growth performance and ecological distribution57. This study revealed a significant (P < 0.05) or highly significant (P < 0.01) positive correlation between all three types of radiation and plant phenotypes. Additionally, perpendicular radiation exhibited a significant (P < 0.05) negative correlation with LFL, while SR showed a significant (P < 0.05) negative correlation with both LFL and SLUE. These findings suggest that favorable lighting conditions can enhance the growth of Elymus nutans plants, whereas excessive light and radiation in the Tibetan region may hinder their growth, particularly affecting LFL and SLUE. While individual meteorological factors may influence the accumulation or composition of certain elements in Elymus nutans seeds, their overall impact on seed element accumulation is minimal. In summary, temperature, precipitation, and light positively influence the phenotypes of Elymus nutans plants, while the high-altitude, arid, and excessively intense radiation environment in Tibet hampers their growth. The influence of meteorological conditions on Elymus nutans extends beyond simple growth measurements. For instance, physiological responses such as stomatal regulation, photosynthesis rates, and stress adaptations play critical roles in how these plants cope with high-altitude conditions. The ecological interactions between Elymus nutans and its environment, including competition with other plant species and mutualistic relationships with soil microorganisms, might also influence growth patterns and should be considered in future studies.
The influence of environmental factors on the growth and seed formation of Elymus nutans
The interaction between the environment and plants forms a complex and mutually dependent system58. Climate variables like temperature, precipitation, and radiation play a direct role in influencing plant growth, photosynthesis, and metabolic processes, while also impacting soil moisture, temperature, and organic matter levels59. Soil serves as the primary medium for plant growth, supplying essential nutrients and water and influencing the development of plant roots and overall health60. Plants uptake water and nutrients from the soil through their roots, simultaneously shaping soil structure and fertility through the decomposition of organic matter and secretion of root exudates61. The overall climate conditions shape the soil environment, wherein soil quality dictates plant growth parameters, with the intricate interplay among these factors establishing the foundation of the ecosystem62. This study highlights the positive impact of favorable meteorological conditions on soil nutrients, thereby fostering the growth and accumulation of seed elements in Elymus nutans. However, the high altitude and latitude of the sampling locations restrict the influence of meteorological factors, leading to an overall inhibition of phenotypic growth in Elymus nutans. Sampling points at high altitudes exhibit the most pronounced inhibitory effect on plant growth, likely due to limited soil nutrients and meteorological factor availability. The study further indicates that the accumulation of elements in Elymus nutans seeds is less influenced by individual soil nutrients and meteorological factors but is significantly impacted by variables such as sampling point altitude, overall soil nutrient levels, and meteorological factors. This could be attributed to the direct influence of soil and meteorological factors on plant phenotypes, indirectly affecting seed element accumulation63. Furthermore, it is plausible that different altitude conditions lead to the interaction between soil nutrients and meteorological factors, influencing the accumulation of seed elements in Elymus nutans64. It is crucial to highlight the limitations associated with the high-altitude sampling locations, as these sites present unique challenges, including limited accessibility and potentially unaccounted meteorological influences, which may not be present in other environments. Moreover, the reliance on ambient environmental conditions as opposed to controlled experiments limits the ability to isolate specific causal relationships. Future research could address these issues by incorporating controlled experimental setups or broader geographical sampling to validate these findings across different ecological contexts.
Conclusions
To conclude, soil ammonium nitrogen (NH4+), soil total phosphorus, and soil available phosphorus emerge as critical nutrients for the growth of Elymus nutans, and their deficiency notably impacts plant phenotype and overall growth. Elevated soil pH levels restrict the uptake and utilization of nutrients by Elymus nutans, thereby impeding its growth. Furthermore, climatic factors like temperature, precipitation, and light exert a positive influence on plant phenotype, whereas the high-altitude, arid, and high-radiation environment in Tibet hinders plant growth. The growth conditions of Elymus nutans are determined by a combination of soil and meteorological factors, with the influence of meteorological factors being restricted by high altitude and latitude, consequently hindering the growth and seed element accumulation of Elymus nutans. Climate conditions contribute to the formation of the soil environment, where soil quality plays a crucial role in shaping the growth conditions of Elymus nutans. These factors interact symbiotically, collectively influencing the ecosystem.
Data availability
The dataset and software will be made available upon request. If anyone needs data from this study, they can contact the first author, Jianting Long.
References
Oldroyd, G. E. D. & Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 368 (6486), eaba0196 (2020).
da Silva, T. M., Cividanes, F. J., Salles, F. A., Pacífico Manfrim Perticarrari, A. L. & Zambon da Cunha, S. B. Monteiro Dos Santos-Cividanes T. Insect pests and natural enemies associated with lettuce Lactuca sativa L. (Asteraceae) in an aquaponics system. Sci. Rep. 14 (1), 14947 (2024).
Mazhar, M. W. et al. Optimizing water relations, gas exchange parameters, biochemical attributes and yield of water-stressed maize plants through seed priming with iron oxide nanoparticles. BMC Plant. Biol. 24 (1), 624 (2024).
Astigarraga, J. et al. Relative decline in density of Northern Hemisphere tree species in warm and arid regions of their climate niches. Proc. Natl. Acad. Sci. U S A. 121 (28), e2314899121 (2024).
Hu, Y. et al. Patterns and driving factors of functional traits of desert species with different elevational distributions in the Tibetan Plateau and adjacent areas. BMC Plant. Biol. 24 (1), 371 (2024).
Zhu, C. et al. Distribution patterns and origins of organophosphate esters in soils from different climate systems on the Tibetan Plateau. Environ. Pollut. 351, 124085 (2024).
Li, J., Li, X., Zhang, C., Zhou, Q. & Chen, S. Phylogeographic analysis reveals extensive genetic variation of native grass Elymus nutans (Poaceae) on the Qinghai-Tibetan plateau. Front. Plant. Sci. 15, 1349641 (2024).
Su, R. et al. Responses of microbial community dynamics, co-occurrences, functional shifts, and natural fermentation profiles of Elymus nutans silage to altitudinal gradients. Microbiol. Spectr. 12 (1), e0251623 (2024).
Liu, J., Pei, X., Zhu, W. & Jiao, J. Water-related ecosystem services interactions and their natural-human activity drivers: implications for ecological protection and restoration. J. Environ. Manage. 352, 120101 (2024).
Guo, X. et al. A field incubation approach to evaluate the depth dependence of soil biogeochemical responses to climate change. Glob Chang. Biol. 29 (3), 909–920 (2023).
Wu, J., Jiao, L., Che, X., Zhu, X. & Yuan, X. Nutrient allocation patterns of Picea crassifolia on the eastern margin of the Qinghai-Tibet Plateau. Int. J. Biometeorol. 68 (6), 1155–1167 (2024).
Zhang, C. et al. Plant diversity is more important than soil microbial diversity in explaining soil multifunctionality in Qinghai-Tibetan plateau wetlands. J. Environ. Manage. 365, 121509 (2024).
Cai, M. et al. Excessive climate warming exacerbates nitrogen limitation on microbial metabolism in an alpine meadow of the Tibetan Plateau: evidence from soil ecoenzymatic stoichiometry. Sci. Total Environ. 930, 172731 (2024).
Li, S. et al. The impact of mixed planting of Poaceae species in the Qinghai-Tibet plateau region on forage yield, soil nutrients, and soil microbial communities. Front. Plant. Sci. 15, 1370593 (2024).
Cheng, X., Luo, M., Chen, K., Sun, J. & Wu, Y. Intra-annual vegetation changes and spatial variation in China over the past two decades based on remote sensing and time-series clustering. Environ. Monit. Assess. 196 (7), 675 (2024).
Lazzarin, M., Driever, S., Wassenaar, M., Marcelis, L. F. M. & van Ieperen, W. Shining light on diurnal variation of non-photochemical quenching: impact of gradual light intensity patterns on short-term NPQ over a day. Physiol. Plant. 176 (4), e14410 (2024 Jul-Aug).
Wan, Z. et al. Effects of water levels on plant traits and nitrogen use efficiency in monoculture and intercropped artificial grasslands. Front. Plant. Sci. 13, 958852 (2022).
Zhao, Q., Guo, J., Shu, M., Wang, P. & Hu, S. Impacts of drought and nitrogen enrichment on leaf nutrient resorption and root nutrient allocation in four tibetan plant species. Sci. Total Environ. 723, 138106 (2020).
Liu, Y. et al. Direct and indirect influences of 8 year of nitrogen and phosphorus fertilization on Glomeromycota in an alpine meadow ecosystem. New. Phytol. 194 (2), 523–535 (2012).
Zhou, Z., Su, P., Yang, J., Shi, R. & Ding, X. Warming affects leaf light use efficiency and functional traits in alpine plants: evidence from a 4-year in-situ field experiment. Front. Plant. Sci. 15, 1353762 (2024).
Liu, Y. Grazing rest during spring regreening period promotes the ecological restoration of degraded alpine meadow vegetation through enhanced plant photosynthesis and respiration. Front. Plant. Sci. 13, 1008550. https://doi.org/10.3389/fpls.2022.1008550 (2022). PMID: 36262656; PMCID: PMC9574263.
Hu, Y. et al. Effects of cd treatment on morphology, chlorophyll content and antioxidant enzyme activity of Elymus nutans Griseb., a native plant in Qinghai-Tibet Plateau. Plant. Signal. Behav. 18 (1), 2187561 (2023).
Jia, Y. et al. QTL mapping of yield-related traits in tetraploid wheat based on Wheat55K SNP array. Plants (Basel). 13 (10), 1285 (2024).
Li, Y. et al. Salinity affects microbial function genes related to nutrient cycling in arid regions. Front. Microbiol. 15, 1407760 (2024).
Zhang, X. et al. Nutrient requirements determined by grain yield and protein content to optimize N, P, and K fertilizer management in China. Sci. Total Environ. 946, 174187 (2024).
Kim, W., Kim, S., Mawhinney, T. P. & Krishnan, H. B. Elemental sulfur concentration can be used as a rapid, reliable, and cost-effective predictor of sulfur amino acid content of soybean seeds. Sci. Rep. 14 (1), 3093 (2024).
Fattobene, M. et al. Distribution of elements in Durum Wheat seed and Milling products: discrimination between cultivation methods through Multivariate Data Analysis. Foods 13 (12), 1924 (2024).
Zhou, J., Guo, W., Hu, Z., Jin, L. & Hu, S. Elemental imaging of fertilized ZnO NP Wheat endosperms using laser ablation-inductively coupled plasma Optical Emission Spectrometry. J. Agric. Food Chem. 71 (49), 19856–19865 (2023).
Long, J., Dong, M., Wang, C. & Miao, Y. Effects of drought and salt stress on seed germination and seedling growth of Elymus nutans. PeerJ 11, e15968 (2023).
Biswas, S. & Biswas, S. Estimation of monthly snowmelt contribution to runoff using gridded meteorological data in SWAT model for Upper Alaknanda River Basin, India. Environ. Monit. Assess. 196 (1), 86 (2023).
Hwang, Y. & Oh, J. Relationship between depression, anxiety, stress, and health-related quality of life in adults with and without chronic diseases: a cross-sectional study. Med. (Baltim). 103 (2), e36967 (2024).
Hu, M. et al. Comprehensive evaluation and selection of 192 maize accessions from different sources. Plants (Basel). 13 (10), 1397 (2024).
Wei, H., You, A., Wang, D. & Zhang, A. Plant-derived essential oil contributes to the reduction of multidrug resistance genes in the sludge composting process. Environ. Int. 190, 108854 (2024).
Khan, S. et al. Predicting the governing factors for the release of colloidal phosphorus using machine learning. Chemosphere 2024 Jun 27:142699 .
Wang, X. et al. Understanding plant-soil interactions underpins enhanced sustainability of crop production. Trends Plant. Sci. Jun. 18 (24), S1360 (2024).
Xu, Y. et al. 2024.The aging of microplastics exacerbates the damage to photosynthetic performance and bioenergy production in microalgae (Chlorella pyrenoidosa). Water Res. Aug 1;259:121841.
Hnizil, O. et al. Integrating NDVI, SPAD, and Canopy Temperature for Strategic Nitrogen and Seeding Rate Management to Enhance Yield, Quality, and Sustainability in Wheat Cultivation. Plants (Basel). Jun 6;13(11):1574. (2024).
Tang, H. et al. Nitrogen availability affects the responses of marsh grass and sedge plants (Phragmites australis and Bolboschoenus Planiculmis) to flooding time. Sci. Total Environ. Jan 15;908: (2023). 168008.168008. Epub 2023 Oct 30.
Wang, S. et al. Effects of excessive nitrogen on nitrogen uptake and transformation in the wetland soils of Liaohe Estuary, northeast China. Sci. Total Environ. Oct. 15, 791:148228 (2021).
Wang, F. et al. Is the NH4+-induced growth inhibition caused by the NH4 + form of the nitrogen source or by soil acidification? Front. Plant. Sci. Sep. 9, 13:968707 (2022).
Duan, Y. et al. Physiological and Morphological Responses of Blackberry Seedlings to Different Nitrogen Forms. Plants (Basel).Mar 28;12(7):1480. (2023).
Saloner, A. & Bernstein, N. Nitrogen source matters: high NH4/NO3 ratio reduces cannabinoids, terpenoids, and yield in Medical Cannabis. Front. Plant. Sci. Jun. 1, 13:830224 (2022).
Zhou, X. et al. Phosphorus addition increases stability and complexity of co-occurrence network of soil microbes in an artificial Leymus chinensis grassland. Front. Microbiol. Mar. 27, 15:1289022 (2024).
Gupta, R. K. et al. Interactive effects of long-term management of crop residue and phosphorus fertilization on wheat productivity and soil health in the rice-wheat. Sci Rep. Jan 16;14(1):1399. (2024).
Hu, Z. et al. 2024.Nutrient-induced acidification modulates soil biodiversity-function relationships. Nat Commun.Apr 3;15(1):2858.
Yang, X. et al. Vegetation communities and soil properties along the restoration process of the Jinqianghe mine site in the Qilian Mountains, China. Front. Plant. Sci. Apr. 22, 15:1358309 (2024).
Yu, L. et al. Cooperation between arbuscular mycorrhizal fungi and plant growth-promoting bacteria and their effects on plant growth and soil quality. PeerJ. Mar 21;10:e13080. (2022).
Ahuja, I., de Vos, R. C., Bones, A. M. & Hall, R. D. Plant molecular stress responses face climate change. Trends Plant. Sci. Dec;15 (12), 664–674 (2010).
Lee, S. et al. Nutrient levels control root growth responses to high ambient temperature in plants. Nat. Commun. Jun 1 (15(1)), 4689 (2024).
Mencuccini, M. et al. A new empirical framework to quantify the hydraulic effects of soil and atmospheric drivers on plant water status. Glob Chang. Biol. Mar. 30 (3), e17222 (2024).
Yang, J. et al. Atmospheric drought dominates changes in global water use efficiency. Sci. Total Environ. Jul. 15, 934:173084 (2024).
Villagra, J., Raggio, J., Alors, D. & Sancho, L. G. Desiccation Tolerance of Epiphytic Macrolichens in an Evergreen Temperate Rain Forest (Alerce Costero National Park, Chile). Plants (Basel). May 31;13(11):1519. (2024).
Wu, Y. et al. Precipitation variation: a key factor regulating plant diversity in semi-arid livestock grazing lands. Front. Plant. Sci. Feb. 28, 15:1294895 (2024).
Bediako-Kyeremeh, B. et al. Effects of wind speed and wind direction on crop yield forecasting using dynamic time warping and an ensembled learning model. PeerJ. Jun 11;12:e16538. (2024).
Chen, S., Liu, W. & Ye, T. Dataset of trend-preserving bias-corrected daily temperature, precipitation and wind from NEX-GDDP and CMIP5 over the Qinghai-Tibet Plateau. Data Brief. May. 21, 31:105733 (2020).
Li, W. et al. Higher Seed Rates Enlarge Effects of Wide-Belt Sowing on Canopy Radiation Capture, Distribution, and Use Efficiency in Winter Wheat. Plants (Basel). Mar 29;13(7):986. (2024).
Neimane-Šroma, S., Durand, M., Lintunen, A., Aalto, J. & Robson, T. M. Shedding light on the increased carbon uptake by a boreal forest under diffuse solar radiation across multiple scales. Glob Chang. Biol. Apr. 30 (4), e17275 (2024).
Li, S. et al. Aridity regulates the impacts of multiple dimensional plant diversity on soil properties in the drylands of northern China. Sci. Total Environ. Jun. 22, 174211 (2024).
Zhang, Y. et al. Global pattern of organic carbon pools in forest soils. Glob Chang. Biol. Jun. 30 (6), e17386 (2024).
Wong, W. S. et al. Limited efficacy of a commercial microbial inoculant for improving growth and physiological performance of native plant species. Conserv Physiol. Jun 18;12(1):coae037. (2024).
Jalloh, A. A., Mutyambai, D. M., Yusuf, A. A., Subramanian, S. & Khamis, F. Maize edible-legumes intercropping systems for enhancing agrobiodiversity and belowground ecosystem services. Sci Rep. Jun 21;14(1):14355. (2024).
Wang, Z. et al. Harvest residue recycling rather than slash-burning results in the enhancement of soil fertility and bacterial community stability in Eucalyptus plantations. Sci. Total Environ. Jun. 18, 945:173850 (2024).
Wang, S. et al. Agronomic traits and ionomics influence on cd accumulation in various sorghum (Sorghum bicolor (L.) Moench) genotypes. Ecotoxicol. Environ. Saf. May. 214, 112019 (2021).
Zhao, Q. Y. et al. Identifying key drivers for geospatial variation of grain micronutrient concentrations in major maize production regions of China. Environ. Pollut. Nov;266 (Pt 2), 115114 (2020).
Acknowledgements
This study was funded by a project chaired by Prof. Yanjun Miao of Xizang Agricultural and Animal Husbandry University National Natural Science Foundation of China (32260342); National Natural Science Foundation of China (U21A20240; U20A2050); and Central Guided Local Science and Technology Development Funds Project (ZYYD2023000147).
Author information
Authors and Affiliations
Contributions
Jianting Long designed the experiment, Xianlei Gao conducted experiments, Jianting Long wrote papers, Yanjun Miao revised the papers.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Long, J., Gao, X. & Miao, Y. Effects of environmental factors on the phenotypic traits and seed element accumulation of wild Elymus nutans in Tibet. Sci Rep 15, 1838 (2025). https://doi.org/10.1038/s41598-025-85415-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-85415-2
