
Abstract
The belief that we can exert an influence in our environment is dependent on distinct components of perceived control. Here, we investigate the neural representations that differentially code for self-efficacy (belief in successfully executing a behavior) and response-efficacy (belief that the behavior leads to an expected outcome) and how such signals may be integrated to inform decision-making. Participants provided confidence ratings related to executing a behavior (self-efficacy), and the potential for a rewarding outcome (response-efficacy). Computational modeling was used to measure the subjective weight of self-efficacy and response-efficacy while making decisions and to examine the neural mechanisms of perceived control computation. While participants factored in both self-efficacy and response-efficacy during decision-making, we observed that integration of these two components was dependent on neural responses within the vmPFC, OFC and striatum. Further, the dlPFC was observed to assign importance to self-efficacy and response-efficacy in specific trials, while dACC computed the trade-off between both components, taking into account individual differences. These findings highlight the contributions of perceived control components in decision-making, and identify key neural pathways involved in computing perceived control.
Similar content being viewed by others

Introduction
Perceived control, which refers to the belief that our ability to exert change in the environment will lead to a desirable outcome, is critical for decision-making. For instance, when COVID-19 vaccines first became available, people often selected vaccination sites based on convenience, perceived safety, or the vaccine brand, with some even traveling farther or waiting longer to book at a preferred site. This demonstrates how perceived control holds inherent value, as individuals are willing to incur costs to exercise control over their vaccination experience1,2,3,4,5. Conversely, lack of control prompts compensatory behaviors, such as perceiving patterns in noise or following superstitions to regain a sense of control6,7. Decades of research have highlighted the importance of perceived control in general well-being8, including increasing one’s ability to cope with stress9,10, exerting positive behavior changes11 and improving cognitive performance12,13. Further, disruptions in perceived control contribute to psychiatric disorders like anxiety and depression14,15.
The core concept of perceived control revolves around the idea that individuals must be able to perceive and identify control within the environment. Researchers utilize diverse methods in experiments, including offering choices16,17, adjusting task difficulty18,19 and controlling performance efficacy20,21. A common observation across such studies is increased neural activation in reward-related regions, including the ventral striatum and ventromedial prefrontal cortex (vmPFC) during anticipation of control or chosen outcomes2,5,19. Furthermore, higher-level integration of control-related reward and cost computation have been suggested to be represented in the dorsal anterior cingulate cortex (dACC) as per the expected value of control theory22.
One limitation of this prior research, however, is that perceived control is often treated as a unitary concept, neglecting its diverse components. The concept of perceived control can be divided into self-efficacy and response-efficacy23. Self-efficacy is the belief in one’s ability to perform actions, while response-efficacy is the belief that certain behaviors lead to desired outcomes. Building on the earlier example of the vaccination experience, individuals’ decisions to get vaccinated involve distinct considerations of self-efficacy and response-efficacy.
They assess their belief in the ability to secure a vaccine appointment (self-efficacy) and whether vaccination would effectively prevent a disease (response-efficacy). Consequently, higher perceptions of both self and response efficacy enhance the likelihood of achieving the desired outcome.
In this paper, we implemented a 3 (self-efficacy) x 3 (response-efficacy) factorial design to quantify these levels and examine their neural correlates. Briefly, while undergoing functional magnetic resonance imaging (fMRI), participants played a series of games where they had to hit a moving target (bar game: measure of self-efficacy) for different probabilities of reward (card game: measure of response-efficacy). Across 3 different tasks, they were then asked to (a) evaluate stimuli that differed as a function of self-efficacy and response-efficacy (Rating task); (b) choose whether to play the trial based on the presented self-efficacy and response-efficacy information (Play task); (c) choose between two different options that differed with respect to self-efficacy and response-efficacy (Choice task). By manipulating self-efficacy and response-efficacy levels, we derived the subjective weight participants assigned to each type of information and their decisions. We hypothesized that neural responses related to self-efficacy might be associated with the subjective value of control (e.g., vmPFC5), while neural responses related to response-efficacy will be more directly associated with reward-related responses (e.g., striatum8). Moreover, given the role of dACC in the computation of control24, we hypothesized that neural responses related to self-efficacy and response-efficacy computation would be represented in the dACC, capturing efficacy-related computations, as participants’ higher level of perceived control calculation in relation to task demands.
Results
The objective of the study was to examine how the integration of self-efficacy and response-efficacy contributes to the formation of perceived control and its impact on decision-making. To accomplish this, we developed a multi-task experimental procedure where participants learned about self-efficacy and response-efficacy separately via training, then executed 3 tasks where self-efficacy and response-efficacy information were presented together (Fig. 1A).

Experimental design. (A) Study Procedure: Participants were instructed to complete the surveys prior to their lab session. Upon arrival at the lab, they proceeded through a series of tasks, including training sessions, an evaluation period, two rating tasks, a play task, and a choice task. (B) Training sessions: The left figure displays three distinct target sizes used in the bar game. Participants were instructed to aim for the target located in the center of the screen using a yellow bar that moved across the screen at a constant speed. During each trial, a specific target size was presented for participants to focus on. The right figure depicts three different card decks, each offering varying probabilities of winning the reward (20%, 50%, 80%), with participants observing the outcome for a specific deck during each trial. (C) Evaluation period: Either the bar game or the card game was displayed on the screen individually, accompanied by a subjective rating of regarding the likelihood of successfully hitting the target (self-efficacy) or the card leading to a reward (response-efficacy). (D) Rating task: Two stimuli were simultaneously presented, and information related to the subsequent question was provided that directed the participant to focus on one particular stimulus (either self-efficacy or response-efficacy). A subjective rating of confidence regarding self-efficacy or response-efficacy was presented afterward similar to the Evaluation period. (E) Play Task: Two stimuli were presented simultaneously similar to the rating task; however, rather than focusing on a specific stimulus (either self-efficacy or response-efficacy), participants were asked to integrate both sources of information followed by questions related to the participants’ choice (whether they would like to play the game) and what they attributed their decisions to (i.e., the bar or the card). (F) Choice Task: Two “Hit the Target” games were presented simultaneously, and participants were asked to choose the game they preferred to play after integrating both sources of information.
Building self-efficacy and response-efficacy—training sessions and evaluation
To understand how participants processed self-efficacy and response-efficacy information, we independently manipulated the likelihood of success of self-efficacy and response-efficacy information in a 3 × 3 factorial design. Two experimental games were created: the “bar game”, which represented self-efficacy, and the “card game”, which represented response-efficacy. In the bar game, using the operational definition of self-efficacy as the belief of an individual in successfully executing a behavior, participants develop their self-efficacy via repeated attempts to hit a target. In the card game, given that response-efficacy was defined as the belief that the behavior would lead to the desired outcome, participants learned to associate earning a card with winning a reward, as the probability of a reward is visually presented in the card (Fig. 1B). The likelihood of success for self-efficacy and response-efficacy information was manipulated with probabilities set at 20%, 50%, and 80%.
To evaluate the effectiveness of the training, participants completed an Evaluation Period in which they provided subjective ratings on the likelihood of successfully hitting the target (Bar game) or winning the reward (Card game) based on the presented stimuli (Fig. 1C). Separate one-way ANOVAs revealed significant differences in participants’ ratings for both self-efficacy (F (2, 76) = 292.83, p < 0.001, η = 0.88) and response-efficacy (F (2, 76) = 160.88, p < 0.001, η = 0.81), indicating that participants correctly evaluated the likelihood of success based on the different levels of self-efficacy and response-efficacy presented (Supplement Figure S2).
Experimental tasks
Following the training (Bar and Card games) and evaluation period, we integrated the bar and card games into a single task, referred to as the “Hit the Target” game, to investigate the neural basis of self-efficacy and response-efficacy during the computation of perceived control via 2 tasks presented in the MRI scanner and 1 task presented outside the MRI. In this interactive setting, participants could only earn a monetary reward if they effectively hit the target with the moving bar (self-efficacy) to win the card, and subsequently, if the card won (response-efficacy). Each of the three tasks differed slightly in the specific questions posed to participants: (1) Rating task - Assessing how self-efficacy and response-efficacy independently influence participants’ perceived confidence in the likelihood of success; (2) Play task - Assessing how integration of self-efficacy and response-efficacy information influences participants’ decision to carry out an action; (3) Choice task - Assessing the trade-offs in terms of self-efficacy and response-efficacy by having participants choose between two different “Hit the Target” games. Taken together, the completion of these tasks allowed for a behavioral and neural measure of how self-efficacy and response-efficacy are independently processed, how they are integrated neurally, and how this integration is considered in the computation of perceived control that influences decision-making.
Behavioral and neural patterns in evaluating self-efficacy and response-efficacy
Subjective ratings of self-efficacy and response-efficacy - rating task
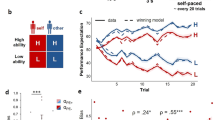
The rating task, where self-efficacy and response-efficacy were presented simultaneously (Fig. 1D), was implemented to assess how participants processed self-efficacy and response-efficacy information. Participants were directed to focus on only one type of information and provide a confidence rating related to either successfully hitting the target (if the cue was the Bar game) or the probability of winning the card (if the cue was the Card game). We observed a significant main effect of self-efficacy on participants’ confidence ratings for successfully hitting the target (F (2, 76) = 273.74, p < 0.001, η = 0.88) and a significant main effect of response-efficacy on participants’ confidence ratings for winning the reward based on the presented card (F (2, 76) = 129.98, p < 0.001, η = 0.776; Fig. 2A). These results support the effectiveness of the experimental design in conveying different levels of self-efficacy and response-efficacy that can impact decision-making.

Results in the rating task: (A) Behavioral: Participants reported greater confidence ratings under the high self-efficacy and response-efficacy levels. In contrast, they reported having the lowest confidence while the self-efficacy and response-efficacy were low. Error bars represent one SEM (***p < 0.001). (B) MVPA: The neural activation in dACC and mPFC exhibit distinct activation patterns associated with participants’ attention shift in self-efficacy and response-efficacy information, guided by the cue information (Question: Target vs. Question: Card). This process involved focusing on one stimulus and evaluating the associated confidence while two stimuli were presented simultaneously.
Neural representation of self-efficacy and response-efficacy
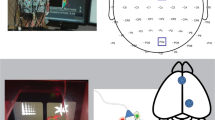
Using multi-voxel pattern analysis (MVPA) as our primary approach, our initial aim was to uncover task-specific neural patterns capable of distinguishing between self-efficacy and response-efficacy conditions during the rating tasks. We observed distinct neural representations in the dACC (Brodmann’s Area (BA) 24/32; peak z-stats = 5.08 at MNI: x, y, z = − 2, 22, 38, cluster-corrected p < 0.05, 677 voxels) and mPFC (BA10/32; peak z-stats = 3.88 at MNI: x, y, z = 2, 46, 12, cluster-corrected p < 0.05, 147 voxels) at the onset of the trial when the stimuli and cue regarding what to focus on were presented. This suggests that at the multi-voxel level, the dACC and mPFC exhibit distinct activation patterns associated with participants’ attention shifts as a function of self-efficacy and response-efficacy information (Fig. 2B).
Following the observation that the dACC and mPFC represented task-specific information, we sought to investigate whether there were brain regions that exhibited linear correlations in a manner similar to participants’ behavior regarding the different levels of self-efficacy and response-efficacy. Given that we hypothesized that neural responses related to self-efficacy might be associated with the subjective value of control, represented in the vmPFC5, while neural responses related to response-efficacy will be more directly associated with a reward response, represented in the striatum8, we performed two GLM analyses (Methods: GLM-1 and GLM-2) to examine the linearity representation in vmPFC and striatum. We conducted region of interest (ROI) analyses focusing on the vmPFC and striatum, utilizing a meta-analytic contrast of brain responses associated with the valuation network and subjective value25. To strengthen the reliability of the findings, we employed a permutation analysis with 50,000 iterations to better assess statistical significance across our selected ROIs. The results revealed that neural activity in the right vmPFC exhibited a significant positive correlation with varying levels of self-efficacy (p = 0.0423). Conversely, the striatum demonstrated a significant positive correlation with response-efficacy information (p = 0.004). These findings suggest a potential distinct representation for self-efficacy and response-efficacy in vmPFC and striatum, respectively.
To rule out the possibility that results from the rating task were solely driven by reward probability, we performed supplementary ROI analyses on the vmPFC and striatum during the evaluation period that occurred during training, where participants assessed the likelihood of success for each piece of presented information. We also employed permutation analysis to ensure the reliability in the findings. In contrast to the rating task, however, no significant effects were observed in the right vmPFC region (self-efficacy: p = 0.6887), nor in the striatum region (response-efficacy: p = 0.2032) during the evaluation period. We then conducted a direct comparison between neural activity in the evaluation and rating tasks across both ROIs. The analyses revealed that vmPFC activity was significantly greater during the rating task compared to the evaluation period with respect to self-efficacy (p = 0.0483). Similarly, we found increased striatum activity during the rating task compared to the evaluation period concerning response-efficacy (p = 0.0156). These results suggest that the activity observed in the rating task cannot be attributed to the likelihood of obtaining a reward, instead representing self-efficacy and response-efficacy information.
Integration of self-efficacy and response-efficacy and the subsequent decision
Behavior - play task and choice task
In light of the result from the rating task, an important remaining question is how the integration of self-efficacy and response-efficacy information contributes to the formation of perceived control and its impact on human decision-making. To characterize the precise nature of self-efficacy and response-efficacy integration, we conducted two tasks to investigate this integration process. First, participants performed the play task, where they were presented with both self-efficacy and response-efficacy information simultaneously and were asked to make decisions on whether they wanted to play the Hit the Target game presented on the screen (Fig. 1E). This required them to integrate both self-efficacy and response-efficacy information. A 3 (self-efficacy) x 3 (response-efficacy) ANOVA revealed significant main effects of both self-efficacy (F (2, 74) = 140.89, p < 0.001, η = 0.79) and response-efficacy (F (2, 74) = 90.176, p < 0.001, η = 0.71; Fig. 3A) and no interaction (F (4, 148) = 1.2388, p = 0.2970, η = 0.03), suggesting that decisions to play were informed by both self-efficacy and response-efficacy information.
In addition to the play task, which assessed participants’ decisions to engage in a specific action (Play or Not Play), participants also performed the choice task where they weighed self-efficacy and response-efficacy information when making decisions. In this task, participants were instructed to choose between two gameplay options, and computational modeling was employed to analyze their subjective weighting on self-efficacy and response-efficacy information. Each option included both a bar and card game that varied with respect to the probability of success, and participants chose the preferred option they would like to execute. Logistic regression analysis and maximum likelihood estimation were conducted on participants’ trial-by-trial choice to examine the subjective weight participants assigned to each piece of information when making decisions (Methods - Behavioral Analyses: Choice task).
We examined participants’ choices using four different models. If participants only consider self-efficacy information (completely ignoring response-efficacy information), the subjective weight participants assign to response-efficacy (\(\:{\omega\:}_{RE}\)) would be 0 (Self-efficacy-only Model). Conversely, if participants completely ignored self-efficacy information (Only considering response-efficacy information), the subjective weight participants assign to self-efficacy (\(\:{\omega\:}_{SE}\)) would be 0 (Response-efficacy-only Model). The integrated model considered both self-efficacy and response-efficacy information, and the Expected Value model further incorporated the expected value of the options into the subjective value estimation. The results indicated that it was the integrated model that considered both self-efficacy and response-efficacy information to predict participants’ decision-making behavior that yielded the best performance (Table 1). However, there was no significant difference found between the weights assigned to self-efficacy and response-efficacy (p = 0.5187). A supplementary analysis that combined the behavioral data from both the play task and the choice task indicated that the subjective value of each option in the play task, estimated using the integrated model, effectively anticipated participants’ decision outcomes (r = 0.353, p < 0.001; Supplement Figure S4). These results suggest that the integration of self-efficacy and response-efficacy had an impact on shaping participants’ perceived control and influenced their decision-making processes.
Decision and attribution dependent neural representation
By utilizing multi-voxel pattern analysis (MVPA), our primary goal within the play task was to reveal two distinct neural patterns. First, we aimed to explore decision-specific neural patterns capable of distinguishing participants’ choices regarding their willingness to engage in the game. This decision-specific signal would pinpoint the brain region responsible for representing the integrated signals that calculate self-efficacy and response-efficacy information for decision-making. Second, we aimed to identify attribution-specific neural patterns that could distinguish participants’ attributions related to their decisions, specifically, which information influenced their choices. This attribution-specific signal would denote the brain regions responsible for representing the control allocation signal, indicating the information participants used in their decision-making.
For the first analysis, we measured the decision signal by examining the neural activity associated with participants’ choices to play or not play the game. We observed decision-dependent neural representations in the vmPFC (BA 25; peak z-stats = 4.12 at MNI: x, y, z = 0, 28, -6, cluster-corrected p < 0.05, 54 voxels), OFC (BA 11; peak z-stats = 3.94 at MNI: x, y, z = − 24, 42, -14, cluster-corrected p < 0.05, 98 voxels), and ventral striatum (peak z-stats = 4.37 at MNI: x, y, z = − 14, 22, -8, cluster-corrected p < 0.05, 66 voxels) during the presentation of stimuli, successfully distinguishing participants’ Yes and No choices (Fig. 3B). For the second analysis, we assessed the attribution signal through participants’ self-reports regarding the factors influencing their decisions. Specifically, after indicating whether they wanted to play, participants explained their reasoning, allowing us to measure how they attributed their choices to information about either the bar or the card. We observed attribution-dependent neural representations in the dlPFC (BA 8/9; peak z-stats = 3.78 at MNI: x, y, z = 26, 36, 42, cluster-corrected p < 0.05, 27 voxels) during the presentation of stimuli. This analysis successfully differentiated participants’ attributions regarding their decisions based on the self-efficacy or response-efficacy information (Fig. 3C). Taken together, the findings suggest that, at the multi-voxel level, the vmPFC, OFC and ventral striatum exhibit distinct activation patterns associated with participants’ decisions to play or abstain from playing the game, while the dlPFC displays distinct activation patterns associated with participants’ attributions of their decisions to self-efficacy and response-efficacy information.

Results in the play task: (A) Behavioral: Participants’ decisions regarding whether to play the game or not were influenced by both self-efficacy and response-efficacy. Error bars represent one SEM. (B) MVPA: The neural activation in vmPFC, OFC and ventral striatum exhibit distinct activation patterns associated with participants’ decision in executing the behavior, the choices to play or not play the game indicated by participants. (C) The neural activation in dlPFC exhibits distinct activation patterns associated with participants’ decision attribution, as indicated by participants’ self-reports regarding the factors driving their choices.
Exploratory analysis: linking participants’ choice behavior with neural representations and individual differences
Building on previous research highlighting the involvement of the dorsal anterior cingulate cortex (dACC) in computing the expected value of control24, we aimed to further investigate how neural activity in the dACC relates to each participant’s decision-making behavior. Here, we distinguish between two forms of control: cognitive control, which encompasses executive functions like attentional and behavioral regulation24, and a broader notion of control related to efficacy—specifically, participants’ beliefs in their influence over outcomes, operationalized as self-efficacy and response-efficacy23. Given the dACC’s established role in the expected value of control computation, we hypothesized that neural responses associated with self-efficacy and response-efficacy computations would localize within the dACC, reflecting participants’ perceived control and adaptive adjustments in line with task demands. To achieve this, we applied our Integrated model during the choice task to probe how individual differences in participant’s achievement motivation26, and trait self-efficacy perceptions27 as well as the neural processing within the dACC during the rating task, contributed to participants’ subjective emphasis on self-efficacy and response-efficacy information.
Using mediation analysis with bootstrapping via the M3 mediation toolbox28,29, we conducted two exploratory analyses to investigate the mechanisms by which individual differences influence participants’ relative subjective weighting of self-efficacy and response-efficacy, mediated by dACC activity during the rating task. With motivation and cognitive control exhibiting a strong correlation30, the initial analysis concentrated on investigating the connection between participants’ achievement motivation26, dACC activity differences estimated from GLM2, and participants’ relative weight assigned to self-efficacy and response-efficacy while making decisions. Recognizing the impact of individual disparities in achievement motivation on decision-making processes is imperative for comprehending how goal orientation influences cognitive strategies, particularly in shaping perceived control. There was no significant association between mastery-approach goal score and dACC activity difference (β = 3.62, p = 0.0987). However, a significant relationship was observed between dACC activity difference and weighting difference (β = 0.02, p = 0.0001). Bootstrapping analysis with 10,000 simulations confirmed that the indirect effect between mastery-approach goal and weighting differences through dACC activity difference was statistically significant (β = 0.07, p = 0.00307; Fig. 4A). No direct effect was found between mastery-approach goal and weighting difference, regardless of whether the dACC activity was included in the model (β = -0.07, p = 0.8293) or not (β = 0.01, p = 0.8549). With mastery-approach goal reflects an individual’s focus on developing competence and mastering new skills or knowledge, high scores indicate a strong orientation towards learning, improvement, and self-development. These findings highlight how mastery-approach goal, which drives individuals towards mastery and improvement, indirectly affects the subjective weighting differences between self-efficacy and response-efficacy during decision-making through the disparity in dACC representation of self-efficacy and response-efficacy information.
Given that overall perceived control may play a part in participants developing context-dependent perceived control8, we proceed to further investigate whether dACC serves as a similar function in mediating the relationship between individuals’ trait self-efficacy27 and participants’ weighting differences. The general self-efficacy measure refers to one’s belief in their capacity to achieve goals and tackle challenges, and understanding how it impacts individuals’ formation of perceiving control on decision-making processes can provide valuable insights into nurturing resilience and effective coping strategies. We identified a noteworthy total effect between general self-efficacy and weighting difference (β = 0.38, p = 0.0033), along with a significant coefficient between dACC activity difference and weighting difference (β = 0.02, p = 0.0001). Adding the dACC activity differences in the model strengthen the direct effect of general self-efficacy on weighting disparities (β = 0.41, p = 0.0011). However, no indirect effect between participants’ general self-efficacy and weighting difference through dACC activity was observed (β = -0.03, p = 0.8731). Subsequently, we conducted a regression analysis to examine the correlation between behavior weighting and the two variables. Three regressors were included: participants’ general self-efficacy scores, dACC activity difference, and an interaction term between perceived self-efficacy trait and brain activity. The robust linear regression revealed significant main effects for both participants’ general self-efficacy (β = 1.2139, SE = 0.5369, t (34) = 2.2608, p = 0.0303; Fig. 4B) and the differences in dACC activity (β = 1.2498, SE = 0.5531, t (34) = 2.2595, p = 0.0304). However, no significant interaction effect was found (β = -0.1034, SE = 0.5515, t (34) = -0.1874, p = 0.8525). These findings suggest that participants’ subjective weight assignment to self-efficacy relative to response-efficacy was primarily influenced by their general self-efficacy and the disparity in dACC activity.

The association between individual variances, encompassing mastery-approach goals and general self-efficacy, and participants’ neural representations alongside their decision-making behavior. (A) Indirect effects model for mastery-approach goal, dACC activity difference and individuals’ relative weight on choice. Participants’ mastery-approach goal score was indirectly related to participants’ relative weight assigned to self-efficacy and response-efficacy during choice through dACC activity differences. (B) Participants’ general self-efficacy was positively correlated with participants’ weighting trade-off while making decisions. Each point represents one participant. The red line represents the regression line and the shaded area represents the 95% confidence interval.
Discussion
In this study, we examined the influence of self-efficacy and response-efficacy on the computation of perceived control and its subsequent impact on decision-making. We created novel tasks to systematically manipulate the levels of self-efficacy and response-efficacy. Participants’ confidence in successfully hitting the target increased with the self-efficacy level, while confidence in winning the reward based on the presented card increased with the response-efficacy level. Thus, participants take into account both self-efficacy and response-efficacy information while making decisions. The results are consistent with prior reports which highlight a preference for control, often referred to as a “control premium”1,2,3,5. The integration of these two types of information involved neural responses within the vmPFC, OFC, and ventral striatum, while the dlPFC was associated with attributing which factor was most important during decision-making. Finally, the dACC was observed to be critical in computing the trade-off between self-efficacy and response-efficacy information after considering individual differences in their achievement motivation.
In the rating task, the vmPFC was engaged when participants reported different levels of self-efficacy, suggesting its involvement in processing self-related beliefs. This is consistent with prior findings implicating the vmPFC in perceiving and exercising control31,32 and computing the subjective value of control5. Our findings support the notion that the representation of controllability in the vmPFC is closely tied to an individual’s internal self-belief, specifically their self-efficacy, which was manipulated by task difficulty in this study. Consistent with previous findings, perceived task difficulty has been shown to negatively correlate with perceived control, whereby increased difficulty reduces self-efficacy by shaping performance expectations18,33. Easier tasks tend to enhance self-efficacy by boosting the likelihood of success, while harder tasks lower it by raising the probability of failure. Given that task difficulty operates as a cost factor within the expected value of control framework24, we structured the design to treat self-efficacy as an independent measure, separate from effort-related considerations. In the task, participants only rated their confidence, without engaging in performance-based responses. This approach allowed us to capture self-efficacy as a self-referential belief, rather than as a function of effort mobilization or aversion—factors typically engaged during active task performance, as discussed in motivational intensity theory34,35. This design minimizes the potential confounding effects of perceived effort costs, focusing specifically on self-efficacy as a standalone construct.
On the other hand, the striatum was found to be engaged when participants processed different levels of response-efficacy, potentially representing the anticipated reward outcomes of the card. This suggests that perceived control, especially response-efficacy, may act as a motivational magnet, with variations in dopaminergic transmission expected to positively correlate with the promotion of approach behavior and response vigor36. Prior studies have provided support for this idea by demonstrating that individuals with a greater sense of control exhibit higher striatal dopaminergic levels37 and engage in more control-seeking behavior. Moreover, corticostriatal circuits have been implicated in the association between performance contingency and perceived control38. These findings highlight the potential significance of dopamine levels and corticostriatal activity in explaining individual differences in control-seeking and control-exerting behaviors. Our study adds to these previous findings and provides further evidence that the representation of controllability in the striatum is closely tied to an individual’s perceived performance contingency, particularly their response-efficacy.
Importantly, our experimental design incorporated two distinct trial types (self-efficacy and response-efficacy) within the rating task. Participants were instructed to concentrate on one attribute during each trial, facilitating the exploration of neural activations linked to shifts in attentional focus on either self-efficacy or response-efficacy. The dACC and mPFC were engaged when participants were directed to different trial types. Specifically, the neural activity pattern observed in the dACC and mPFC effectively discriminates the information participants were prompted to concentrate on, indicating their involvement in processing attention-dependent task signals. This aligns with prior proposals suggesting that the dACC is associated with functions such as conflict monitoring39,40 and, along with the mPFC, attentional control41,42. Our study extends these findings, demonstrating that dACC and mPFC play a role in attentional competition under perceived control context.
We further utilized two new tasks to explore the behavioral and neural representation of integration self-efficacy and response-efficacy and their impact on decision-making. In the play task, participants evaluated one game with independent self-efficacy and response-efficacy levels to determine whether to engage in a specific action (i.e., “play” the game), revealing influences of both self-efficacy and response-efficacy on decisions. Next, in the choice task, participants selected one game to play from two possible options after assessing self-efficacy and response-efficacy information for each option. Computational modeling revealed the integration of self-efficacy and response-efficacy in decision-making. Furthermore, combining data from both tasks showed that the subjective value, estimated using the integrated model, accurately predicted decisions in the play task. Taken together, these findings underscore the critical role of integrating self-efficacy and response-efficacy in shaping perceived control and decision-making processes.
Next, we probed how the brain represents the integration of self-efficacy and response-efficacy information. The play task featured two distinct questions that participants had to assess each trial: 1- play the game or not); 2- specify what they attributed their decision to (self-efficacy or response-efficacy). We observed the involvement of reward-processing regions for executing a behavior (to play or not). Specifically, regions included the vmPFC and OFC, recognized for their role in encoding the subjective value of offered goods43,44,45 and the ventral striatum, associated with a subjective preference for control2. Furthermore, dlPFC is involved when participants attributed their choices to self-efficacy or response-efficacy. Considering the proposed role of the dlPFC in control implementation41 and conflict monitoring theory, this finding posits its involvement in resolving conflicts at the response level46. However, it is important to acknowledge the intricacies of the attribution process, and that participants’ decision-making rationale may be influenced by positive factors supporting their current choice or negative factors guiding them away from alternative options. Hence, while these findings collectively suggest that the decision-making process for executing an action is shaped not only by the reward-processing regions, such as the vmPFC, OFC, and ventral striatum but also by the dlPFC, potentially serving as the control allocation area attributing decisions to either self-efficacy or response-efficacy information, future studies on the attribution process are warranted.
Finally, we performed two exploratory analyses to examine individual differences in the relationship between neural computations and behavioral choices. We considered the established role of the dACC in motivating effortful behavior, behavioral flexibility, and monitoring control signals24,47,48. According to the expected value of control theory, decisions are made by weighing the costs of exerting control against the anticipated rewards22. This theory incorporates perceived control into the analysis of effort allocation, with higher perceived control leading to increased effort49,50,51. This overall value assessment is processed in the dACC, which integrates cost-benefit analyses for control allocation24,50. Based on this theoretical framework, we hypothesized that neural responses in the dACC would reflect a higher-level perceived control calculation responsive to task demands, specifically relating to subjective weight trade-off between self-efficacy and response-efficacy computations. We utilized two individual difference measures—achievement motivation26 and general self-efficacy27—selected from a preliminary set of five questionnaires administered during the pre-session assessment. We selected the measures due to observed high collinearity among the general self-efficacy and other scales, which helps enhance clarity and interpretability while minimizing redundancy. The general self-efficacy was prioritized for further analysis, given its notable correlations with other constructs: the Attentional Control Scale52 (ASC; r = 0.5141, p < 0.0001), the Desirability of Control Scale53 (DCS; r = 0.3891, p = 0.0158), and the Locus of Control Scale54 (LCS; r = 0.3069, p = 0.0609). Therefore, we only selected the achievement motivation scale and general self-efficacy in our exploratory individual differences analysis. We observed an indirect effect of dACC neural representation on the relationship between individuals’ mastery-approach goals and their behavioral trade-offs between self-efficacy and response-efficacy. Additionally, we observed that neural representation in the dACC, and individual differences in the general trait of self-efficacy, effectively predicted participants’ behavioral trade-offs on self-efficacy and response-efficacy. These trade-offs refer to the differential subjective weights participants assigned to their subjective control in the sense of efficacy, self-efficacy and response-efficacy. Given that mastery-approach goals signify a desire to achieve deep understanding and self-improvement, while general self-efficacy indicates confidence in overcoming challenges, these findings align with the expected value of control theory22, suggesting that the dACC is involved in the higher-level computation of control-related rewards, and highlight the significance of individual differences in this process. Although it should be noted that we utilized the ACC area from the automated anatomical atlas (AAL55) as a ROI mask to assess the computation of self-efficacy and response-efficacy which encompasses a broader region of dACC.
Our study emphasizes the importance of the relationship between ACC activation and individual differences in general perceived self-efficacy and mastery-approach goal, influencing participants’ decisions. Specifically, the ACC in general serves as a potential computational signal for processing self-efficacy and response-efficacy information during decision-making after accounting for individual differences. Nonetheless, replication is necessary to confirm these intriguing results.
While the results are promising in terms of further understanding the mechanisms underlying perceived control, there are some limitations to the current investigation that are worth discussing. First, this study manipulated task difficulty to represent the level of self-efficacy and reward contingency to signify the level of response-efficacy and captured these variables using behavioral and fMRI measures. However, the complex nature of perceived control calls for adopting a comprehensive multi-methodological approach, including physiological measures like pupil dilation and real-world behavioral observations such as ecological momentary assessment, to enhance the precision of examining self-efficacy and response-efficacy. Moreover, in simplifying the task, two approaches were employed. First, participants acquired self-efficacy as implicit information, while response-efficacy was provided explicitly. Previous studies have suggested that implicit learning and explicit learning may differ during decision-making processes56,57. Therefore, investigating how the formation of self-efficacy and response-efficacy using both implicit and explicit approaches affects the integration process and subsequent decisions would provide valuable insights for future studies. Second, to mitigate potential confusion and cognitive load for participants, information about the bar and card games were consistently presented in the same location throughout the task. Although the current data cannot assess multivariate patterns related to location, existing studies suggest that spatial attention is primarily represented in parietal regions58,59, rather than in the dACC and vmPFC regions identified in our findings. Future research could address this limitation by varying the spatial locations of the two pieces of information to further explore the impact of location on multivariate patterns in this task.
Finally, our study primarily focused on healthy individuals, but has important implications for clinical populations with prevalence of control loss in psychopathologies like addiction, post-traumatic stress disorder, and depression60,61,62. The distinctive feature of addiction, marked by the inability to resist addictive substances63, emphasizes the need to explore perceived control components in decision-making processes. Further, issues with perceived control are an important characteristic of patients afflicted with depression64. Examining the different components of perceived control and their influence on decision-making provides valuable insights into the mechanisms underlying perceived control and identifies potential avenues for psychotherapeutic intervention in conditions characterized by deficits in this domain. Furthermore, considering the widespread impact of perceived control on general well-being and decision-making, a nuanced understanding of how self-efficacy and response-efficacy interact to shape individuals’ decisions can enhance our comprehension of how perceived control influences choices, coping strategies, and overall psychological health. This knowledge can guide the development of more effective strategies to improve decision-making and adaptive behavior across diverse contexts.
Methods
Participants
A total of 42 healthy (24 female and 18 male; mean age: 24.5; SD: 6.7), right-handed participants were recruited from Rutgers University-Newark and the broader Newark community. Participants received $45 for the fMRI study and a $5 bonus based on performance in the experiment. Participants had no history of psychiatric and neurological illness. Criteria for ineligibility to participate in the experiment included ferromagnetic materials in the body, claustrophobia, and/or suspected pregnancy. All methods were carried out in accordance with relevant guidelines and regulations. All experimental protocols were approved by the Rutgers University Institutional Review Board. Written informed was obtained from all subjects. Two participants were removed from subsequent behavioral and neural analyses due to extreme head movement in MRI. Additionally, one participant was excluded from the play and choice tasks only due to equipment failure. Final analysis was therefore conducted on 39 participants.
Questionnaires
To explore individual differences in controllability and attention, participants completed 5 pre-session questionnaires, including the General Self-Efficacy Scale (GSE27), Locus of Control scale (LCS54), Desirability of Control Scale (DCS53), Attentional Control Scale (ASC52), Achievement Goal Questionnaire (AGQ-R26; GQ65).
Experimental design
To probe how participants integrate self-efficacy and response-efficacy information to shape one’s perception of control and make decisions, two experimental games were designed: the bar game and the card game focused on self-efficacy and response-efficacy, respectively. Participants first underwent two training sessions and one Evaluation Period with the goal of learning self-efficacy and response-efficacy information. Second, they executed 3 tasks where self-efficacy and response-efficacy information was presented together in the same order (1) Rating task; (2) Play task; (3) Choice task.
All computerized tasks were coded and presented using PsychoPy66 and behavioral responses were recorded using either a laptop or an fMRI-compatible button box. Participants received points throughout the experiment as rewards. At the end of the experiment, the points were converted to a monetary reward bonus ($0–5). Below, we discuss the experimental procedures in more detail.
Training sessions and evaluation
Participants first completed two training sessions outside the fMRI scanner to learn self-efficacy and response-efficacy information, respectively. They underwent another two training sessions inside the fMRI scanner to validate the training and an Evaluation period where they assessed training success.
Self-efficacy training - bar game
Rationale: The “bar game” was a visual-motor task designed to manipulate self-efficacy, defined as the belief in successfully executing a behavior. Participants developed their self-efficacy by attempting to hit a target throughout the training session (see Fig. 1B).
Procedure: The game consisted of 60 trials, each presenting a target of varying size in the center of the screen along with a yellow bar moving at a constant speed. Participants were instructed to hit the target with the yellow bar by pressing a button. Successful execution of the behavior would lead to a reward of 5 points. Three different target sizes (10, 20, and 30 pixels) were used during the training session to establish the relationship between self-efficacy and target size. Participants learned that larger target sizes increased the probability of hitting the target, thereby enhancing their confidence in successfully executing the behavior. At the conclusion of this training phase, participants were probed about their estimated likelihood of success for each target size. The position of hits across all training trials was utilized to estimate each participant’s unique motor uncertainty. Based on previous literature, the distribution of participants’ distances from the endpoint to the target approximated an isotropic bivariate Gaussian distribution67. We quantified the level of self-efficacy by estimating the standard deviation (SD) of this distribution for each participant68. In the subsequent tasks, target sizes were adjusted for each participant to achieve specific hit rates (20%, 50%, 80%). To validate this, participants completed a second self-efficacy training consisting of another 60 trials with three adjusted target sizes in the fMRI before scanning.
Response-efficacy training - card game
Rationale: The card game was designed to represent information about response-efficacy, defined as the belief that behavior itself will lead to an expected outcome. The probability of receiving a reward was visually displayed on the presented cards (see Fig. 1B).
Procedure: Participants observed 60 trials of outcomes from three different decks of cards, each deck representing a reward probability of 0.2, 0.5, and 0.8 as shown on the card draw. Each deck was presented in separate blocks. There were 30 trials conducted outside the MRI and another 30 trials inside before the scanning started. In each trial, participants observed the result of a card draw from one of the decks, without making any decisions. They received 5 points if the drawn card indicated a reward; otherwise, they received 0 points. At the conclusion of the response-efficacy training phase, participants were asked to estimate the likelihood of winning for each deck based on the probabilities they observed.
Evaluation period (Fig. 1C)
Rationale: Participants evaluated the likelihood of success associated with a presented bar or card game. Since we manipulate the levels of self-efficacy and response-efficacy with probabilities set at 20%, 50%, and 80% for each type of information, it is essential to differentiate the neural representation of self-efficacy and response-efficacy from the probability of receiving a reward. Therefore, the Evaluation Period was designed as a manipulation check to ensure participants learned self-efficacy and response-efficacy information and a baseline condition that helped distinguish the neural representation of self-efficacy and response-efficacy, which relate more to participants’ beliefs, from the reward probability.
Procedure: The session consisted of 60 trials (evenly split between bar and card games). A typical trial involved a presentation of an item (i.e., bar or card game) for 2 s followed by a 1-3 s fixation period (interstimulus interval [ISI]). Participants were then asked to provide a subjective rating on their evaluation of the likelihood of success of hitting the target (Bar game) or winning the reward (Card game), followed by a 1 s jittered intertrial interval (ITI) showing a fixation cross.
Experimental tasks
Participants completed three separate tasks aimed at examining the behavioral and neural basis of self-efficacy and response-efficacy during the computation of perceived control. The experimental tasks included the “Hit the Target” game, featuring both self-efficacy (Bar game) and response-efficacy (Card game) information. Participants were informed that successfully hitting the target in the bar game would earn them the presented card, which was associated with a monetary reward of 10 points they had a chance to win. Thus, a monetary reward would only be earned if participants successfully hit the target with the moving bar (self-efficacy) and won the card (response-efficacy). To examine the effects of self-efficacy and response-efficacy on perceived control, we independently manipulated the likelihood of self-efficacy and response-efficacy in a 3 × 3 factorial design.
Rating task (Fig. 1D)
Rationale: The rating task was designed to quantify the neural substrates of self-efficacy and response-efficacy representation, respectively.
Procedure: Participants played two sessions of the Rating tasks, each session consisted of 72 trials. Each trial began with two seconds presentation of both self-efficacy (Bar) and response-efficacy (Card) information, with a reminder of what the participants should focus on for that trial. In specific trials, participants were guided to concentrate on self-efficacy information alone, while in different trials, their focus was solely on response-efficacy information. Following a jittered 1-3 s ISI, they were asked to provide a confidence rating regarding successfully hitting the target (if the cue was the Bar game) or the likelihood of winning the card (if the cue was Card game). A 1 s jittered ITI showing a fixation cross was presented to signal the end of each trial.
Play task (Fig. 1E)
Rationale: The Play task was designed to assess how self-efficacy and response-efficacy are integrated. Given that the objective of the study is to examine the integration of self-efficacy and response-efficacy, it is essential not only to investigate the neural substrates of self-efficacy and response-efficacy but also to understand how these types of information are used in decision-making.
Procedure: The design of the Play task was similar to the Rating task, with the key difference being that participants were asked to assess both types of information simultaneously, rather than focusing on just one type of information. A choice to “play” a particular trial indicated whether the presented combination was desirable (e.g., high self-efficacy and high response-efficacy vs. low self-efficacy and low response-efficacy), and the following attribution question provided insight into the reasoning behind their decisions. This session consisted of 72 trials where participants were presented with the Hit the Target game that lasted 2 s and was followed by a jittered 1-3 s ISI. Participants were then given 2 s to make a decision regarding whether they wanted to play the game, followed by a 1 s ISI. Subsequently, a 2 s question was presented asking participants to attribute their decision to either the bar or the card, followed by a 1 s ITI.
Choice task (Fig. 1F)
Rationale: The Choice task was designed to quantify the subjective weight participants assign to self-efficacy and response-efficacy during decision-making through computational modeling in a behavioral task following the fMRI scan. This task probed how participants trade-off these two types of information to form their perceived control and make inferences about their choices.
Procedure: The task consisted of 102 trials, where in each trial, participants faced binary choices between two Hit the Target games. As an incentive control, participants would have a chance to play the game they chose at the end of the experiment for additional monetary compensation.
Data analysis
Behavioral analyses
Training sessions and evaluation
Self-efficacy training - bar game: The hit rates for the adjusted target sizes were assessed across participants to verify that they achieved the target hit rates of 20%, 50%, and 80% as intended by the manipulation. Additionally, a one-way ANOVA was conducted on participants’ single-instance likelihood ratings across different target sizes as a manipulation check (see Supplementary Figure S1A).
Response-efficacy training - card game: A one-way ANOVA was conducted on participants’ single-instance likelihood ratings for different decks of cards as a manipulation check (see Supplementary Figure S1B).
Evaluation period: Two one-way ANOVAs were conducted to examine the effectiveness of our manipulation on participants’ average likelihood ratings. One ANOVA examined the impact of self-efficacy using trials where only the bar game was presented, while the other assessed the effect of response-efficacy using trials where only the card game was presented.
Experimental tasks
Rating task
While participants had simultaneous access to self-efficacy and response-efficacy information, they were directed to focus exclusively on the cue information (Bar or Card). They rated their confidence on a 4-point Likert scale that varied in terms of intensity (Extremely, Slightly) and confidence (Confident, Not Confident). Given the concurrent presentation of self-efficacy and response-efficacy information, two two-way ANOVAs were conducted to determine the effectiveness of converting the likelihood of success into confidence ratings for self-efficacy and response-efficacy information.
Play task
Participants were asked to decide whether they wanted to play the game after considering the presented self-efficacy and response-efficacy information, rating their decision on a 4-point Likert scale that varied in terms of intensity (Definitely, Slightly) and decision (Yes, No). A two-way ANOVA was conducted to examine the influence of self-efficacy and response-efficacy on participants’ decisions to engage in the game. Additionally, participants were asked to attribute their decision to either self-efficacy or response-efficacy information, providing their attribution using a 4-point Likert scale (Definitely Bar, Slightly Bar, Slightly Card, Definitely Card). This attribution question was analyzed using two independent ANOVAs based on participants’ decisions (Yes vs. No) to assess how self-efficacy and response-efficacy influenced their attribution judgments under their decision (see Supplementary Figure S3).
Choice task
Participants were asked to pick between two possible Hit the Target games, and their trial-by-trial data was used to fit into a logistic regression to probe individual choices and estimate the subjective weight participants assigned to each piece of information utilizing four distinct models.
Derivation of the subjective weights: To compare the 2 options, we computed subjective value (SV) for both options by taking into account the probability information of self-efficacy, and the probability information of response-efficacy as follows:
Where PSE is the probability of success in self-efficacy and PRE is the probability of a win in response-efficacy; \(\:{\omega\:}_{SE}\) is the subjective weight participants assign to self-efficacy and \(\:{\omega\:}_{RE}\) is the subjective weight participants assign to response-efficacy. The probability of self-efficacy and response-efficacy (PSE and PRE) was set at 0.2, 0.5, and 0.8.
To probe participants’ choices, we fitted their trial-by-trial choice into a regression. Each choice pair was coded by the SV difference between the two options. Maximum likelihood estimation was used to fit the trial-by-trial choice data of each participant to the following logistic function of the form:
Where P is the probability of choosing the 1st option, SV is the subjective value for each option, b is the slope of the logistic function, and c is a constant.
Model comparison
To investigate participants’ consideration of self-efficacy and response-efficacy in their decision-making process, we conducted a comparative analysis involving four distinct models: the Integrated model, the Self-efficacy-only model, the Response-efficacy-only model, and the Expected Value model.
In the Integrated model, both self-efficacy and response-efficacy information are factored into the decision process, with \(\:{\omega\:}_{SE}\) and \(\:{\omega\:}_{RE}\) representing the subjective weights assigned by participants to each type of information. The Self-efficacy-only model is characterized by fixing \(\:{\omega\:}_{RE}\) to 0, signifying that participants exclusively consider self-efficacy information. Conversely, the Response-efficacy-only model sets \(\:{\omega\:}_{SE}\) to 0, indicating the sole consideration of response-efficacy information. In addition to exploring the integration of self-efficacy and response-efficacy, we examined whether participants incorporated the expected value of options into their decision-making. To do so, we introduced an Expected Value model that factors in the expected value that calculates the anticipated reward from each option as follows:
Where \(\:{\omega\:}_{EV}\) is the subjective weight participants assign to the expected value. The expected value is computed by multiplying PSE, PRE, and the magnitude rewarded, with a fixed value of 10 points in each option. A new weighting parameter was added to reflect the subjective importance participants assign to the expected value of an option.
Bayesian information criterion (BIC) was employed to compare and evaluate the fit of these models since each model had a different number of parameters. Through model comparison across these four models, we aimed to gain insights into participants’ utilization of self-efficacy, response-efficacy, and expected value in their decision-making. Subsequent to the comparison, sign tests were performed to investigate the differences in the weights of participants assigned to self-efficacy and response-efficacy information.
Neuroimaging analysis
Neuroimaging data acquisition and preprocessing
Structural and blood oxygen level-dependent (BOLD) functional images were collected using a 3T PRISMA scanner with a 64-channel head coil at Rutgers University Brain Imaging Center (RUBIC). High-resolution structural images were acquired with a T1-weighted MPRAGE sequence (256 × 256 matrix, FOV 256 mm, 176 1-mm sagittal slices). The blood oxygen level-dependent (BOLD) functional images were acquired with a multi-band echo-planar imaging sequence (TR = 1 s, TE = 30 ms, FOV = 230 mm, flip angle 52, echo spacing = 0.49 ms, multiband acceleration factor 4, 56 axial slices, voxel size 2.4 mm3). A field map sequence acquired prior to functional imaging was used to correct geometrical distortion in the functional images. High-resolution structural images were used to correct geometrical distortion in the functional images. The Fmriprep69 pipeline was used for geometrical distortion correction, head motion and slice-timing correction, and normalization of images to MNI standard space. Motion outliers will be identified using the “non-stdDVARS” and “FramewiseDisplacement” columns from the FMRIPREP confounds file, which are thresholded at the 75th percentile and 1.5 times interquartile range. These outliers will be included as confounded regressors in the first-level general linear model (GLM).
fMRI analysis
Multi-voxel pattern analysis (MVPA)
We employed multivariate pattern analysis (MVPA) to investigate the neural representation of self-efficacy and response-efficacy information. Prior to implementing MVPA, trial-by-trial beta values were estimated from unsmoothed preprocessed fMRI data using the general linear model (GLM) approach70. GLMs included regressors for the trial of interest, same-condition trials, and different-condition trials. convolved with a gamma hemodynamic response function. Motion parameters served as control regressors. These regressors were convolved with a gamma hemodynamic response function, while six motion parameters were included as control regressors. This process was repeated for each trial, resulting in trial-by-trial beta values for all trials in the two conditions. These beta values were then used as observations for the MVPA analysis. We implemented a whole-brain searchlight approach71, where a sphere with a diameter of 5 voxels (10 mm) was formed, and the activity patterns across the voxels within each sphere were used for the MVPA analysis.
In our whole-brain searchlight analysis, we employed a support vector machine (SVM) with a linear kernel to decode different error signals and choices. The SVM implementation was done using the LIBSVM toolbox72. To prevent overfitting, we conducted 3-fold cross-validation, where one-fold served as the testing data, and the other two folds as training data. The training data were used to train the classifier, which was then applied to the testing data to evaluate the classification accuracy. In the linear SVM, the parameter “c” regulates the balance between minimizing training error and maximizing generalization, was optimized within the range [0.001, 0.01, 0.1, 1, 10, 100, 1000]. During the classifier training, the training data were further divided into 3-folds to select the optimal value for the “c” parameter using cross-validation. The optimal value was chosen to maximize the cross-validation accuracy. Once the optimal “c” value was determined, the model was retrained using the entire training data, and the classification accuracy was calculated on the testing data. This procedure was repeated for each of the three folds, and the average classification accuracy was computed. To mitigate the impact of varying numbers of trials in each category on the classification accuracy, we employed balanced accuracy. For balanced accuracy, we calculated the classification accuracy within each category and then averaged the accuracies across all categories. The baseline balanced accuracy was also validated via permutations with 5000 iterations. In each permutation, each trial was randomly assigned to a category with a probability proportional to the number of trials in that category among all the trials. The average balanced accuracy across these iterations served as the baseline accuracy. The baseline accuracy was 50% for the two categories.
Rating tasks
We examined how the multi-voxel neural pattern in the current trial could discriminate the type of information (i.e., self-efficacy vs. response-efficacy) in the rating task. In analyzing this task-dependent signal, we categorized trials into two bins based on task type: self-efficacy trials were labeled as 0, and response-efficacy trials were labeled as 1.
Play task
We examined how the multi-voxel neural pattern in the current trial could distinguish participants’ decisions regarding playing the game (i.e., Yes vs. No) and their attributions for their decision (i.e., self-efficacy (Bar) vs. response-efficacy (Card)) in the Play task. In the analysis concentrating on the decision-dependent signal, we classified trials into two bins according to participants’ decisions: No trials were designated as 0, and Yes trials were designated as 1. In the analysis focusing on the attribution-dependent signal, we categorized trials into two bins based on participants’ attributions: self-efficacy trials were denoted as 0, and response-efficacy trials were denoted as 1.
After obtaining the classification accuracy for each participant, the baseline accuracy was subtracted from the classification accuracy for each participant.
Prior to the group-level analysis, individual accuracy maps were smoothed using a 6 mm FWHM Gaussian kernel. To control false-positive rates73, statistical testing was conducted using one-sample cluster-mass permutation tests with 5000 iterations. A voxel threshold uncorrected p < 0.001 was used to identify clusters, and the corrected cluster p-value (p < 0.05) was estimated for each cluster.
Univariate activity correlated with self-efficacy and response-efficacy
To test our hypotheses regarding the linear representation of self-efficacy and response-efficacy in the vmPFC5 and striatum8, we conducted two GLM analyses. Both GLM analyses were carried out with FMRIB’s Software Library (FSL74) FEAT (FMRI Expert Analysis Tool) Version 6.075.
Evaluation period
GLM-1: The model estimated mean activity for self-efficacy and response-efficacy conditions (3 probability each) at the time of self-efficacy or response-efficacy presentation in the Evaluation period. For each condition separately, we implemented the following regressors: R1) an indicator function with a duration of 2 s. At the time of response, we implemented R7) an indicator function with the duration of subjects’ response time. We set up two contrasts - the contrast [R1 R2 R3] = [-1 0 1] was used to identify the neural correlates of group mean of probability in self-efficacy probability, and contrast [R4 R5 R6] = [-1 0 1] was used to examine the neural correlates of probability in response-efficacy.
Rating task
GLM-2: Univariate analysis in the rating task utilized a run-level GLM with regressors for a 2 s indicator function in each condition (2 Question X 3 Self-efficacy X 3 Response-efficacy) at the time of self-efficacy and response-efficacy representation. At the time of response, we implemented an indicator function with the duration of the subjects’ response time. The same contrasts as GLM-1 were used regarding the type of question asked in the task for self-efficacy and response-efficacy linear representation.
Region of interest (ROI) analysis
The vmPFC and striatum regions of interest were defined by a meta-analytic contrast of brain responses to the valuation network that correlates with subjective value25. To explore potential asymmetric representation, we further segmented the mask regions into left and right vmPFC and striatum. To strengthen the reliability of the finding, we conducted one-sample permutation tests with 50,000 iterations within the rating task to assess the linear representation of self-efficacy and response-efficacy. As a control condition, we repeated the same permutation tests during the evaluation period. Additionally, to examine differences between the ROIs across tasks (rating and evaluation period), we calculated the differences between each ROI’s response in the two tasks and conducted one-sample permutation tests with 50,000 iterations to assess statistical significance. In addition, for the exploration of the dACC region, we utilized the automated anatomical atlas (AAL55), specifically targeting the Anterior Cingulate Cortex (abbreviated as ACC).
Exploratory analysis: combining neural and behavior—mediation analyses
To examine individual differences in the relationship between neural computations and behavioral choices, we employed two measures of individual differences: achievement motivation and general self-efficacy. Achievement motivation26, encompasses mastery-approach, mastery-avoidance, performance-approach, and performance-avoidance goals, providing insight into the strategies participants employ when making decisions. General self-efficacy27 reflects an individual’s belief in their ability to overcome challenges. To understand how achievement motivation and general self-efficacy impact the formation of perceived control in decision-making processes related to neural computations, we utilized participants’ relative subjective weighting of self-efficacy and response-efficacy, calculated from the integrated model in the choice task. Additionally, we examined neural activity in the dorsal anterior cingulate cortex (dACC) during the rating task, given the dACC’s established role in the computation of control-related rewards24. We conducted a linear mixed effects mediation analysis using the MATLAB mediation toolbox28,29, implementing 10,000 bootstrap simulations for robust statistical inference.
Data availability
The behavioral data that support the findings of these studies are available on GitHub (https://github.com/delgado-lab/PerceivedControlIntegration). Thresholded and unthresholded image data are available on NeuroVault (https://neurovault.org/collections/17172/).
References
Bobadilla-Suarez, S., Sunstein, C. R. & Sharot, T. The intrinsic value of choice: the propensity to under-delegate in the face of potential gains and losses. J. Risk Uncertain. 54, 187–202 (2017).
Leotti, L. A. & Delgado, M. R. The inherent reward of choice. Psychol. Sci. 22, 1310–1318 (2011).
Owens, B. D., Grossman, Z. & Fackler, R. The Control Premium: A Preference for Payoff Autonomy Author (s): David Owens, Zachary Grossman and Ryan Fackler Source : American Economic Journal : Microeconomics, November Vol. 6, No. 4 Published by : American Economic Association Stable. 6, 138–161 (2014). (2014).
Wang, K. S., Yang, Y. Y. & Delgado, M. R. How perception of control shapes decision making. Curr. Opin. Behav. Sci. 41, 85–91 (2021).
Wang, K. S. & Delgado, M. R. Corticostriatal circuits encode the subjective value of Perceived Control. Cereb. Cortex. 29, 5049–5060 (2019).
Kay, A. C., Whitson, J. A., Gaucher, D. & Galinsky, A. D. Compensatory control Achieving Order through the mind, our institutions, and the heavens. Curr. Dir. Psychol. Sci. 18, 264–268 (2009).
Whitson, J. A. & Galinsky, A. D. Lacking control increases illusory pattern perception. Science 322, 115–117 (2008).
Ly, V., Wang, K. S., Bhanji, J. & Delgado, M. R. A reward-based framework of perceived control. Front. Neurosci. 13, 1–11 (2019).
Bhanji, J. P., Kim, E. S. & Delgado, M. R. Perceived control alters the effect of acute stress on persistence. J. Exp. Psychol. Gen. 145, 356–365 (2016).
Ronen, T., Hamama, L., Rosenbaum, M. & Mishely-Yarlap, A. Subjective well-being in adolescence: the role of Self-Control, Social Support, Age, gender, and Familial Crisis. J. Happiness Stud. 17, 81–104 (2016).
Schnoll, R. A. et al. Increased self-efficacy to quit and perceived control over withdrawal symptoms predict smoking cessation following nicotine dependence treatment. Addict. Behav. 36, 122–127 (2011).
Armaş, I., Cretu, R. Z. & Ionescu, R. Self-efficacy, stress, and locus of control: the psychology of earthquake risk perception in Bucharest, Romania. Int. J. Disaster Risk Reduct. 22, 71–76 (2017).
Li, J., Lepp, A. & Barkley, J. E. Locus of control and cell phone use: implications for sleep quality, academic performance, and subjective well-being. Comput. Hum. Behav. 52, 450–457 (2015).
Gallagher, M. W., Bentley, K. H. & Barlow, D. H. Perceived Control and vulnerability to anxiety disorders: a Meta-analytic review. Cogn. Ther. Res. 38, 571–584 (2014).
Liu, R. T., Kleiman, E. M., Nestor, B. A. & Cheek, S. M. The hopelessness theory of Depression: a quarter-century in review. Clin. Psychol. Sci. Pract. 22, 345–365 (2015).
Bown, N. J., Read, D. & Summers, B. The lure of choice. J. Behav. Decis. Mak. 16, 297–308 (2003).
Leotti, L. A., Iyengar, S. S. & Ochsner, K. N. Born to choose: the origins and value of the need for control. Trends Cogn. Sci. 14, 457–463 (2010).
Depasque Swanson, S. & Tricomi, E. Goals and task difficulty expectations modulate striatal responses to feedback. Cogn. Affect. Behav. Neurosci. 14, 610–620 (2014).
Stolz, D. S., Müller-Pinzler, L., Krach, S. & Paulus, F. M. Internal control beliefs shape positive affect and associated neural dynamics during outcome valuation. Nat. Commun. 11, 1–13 (2020).
Frömer, R., Lin, H., Wolf, D., Inzlicht, C. K. & Shenhav, A. M. Expectations of reward and efficacy guide cognitive control allocation. Nat. Commun. 12, (2021).
Grahek, I., Frömer, R., Prater Fahey, M. & Shenhav, A. Learning when effort matters: neural dynamics underlying updating and adaptation to changes in performance efficacy. Cereb. Cortex. 33, 2395–2411 (2023).
Shenhav, A., Botvinick, M. M. & Cohen, J. D. The Expected Value of Control: an integrative theory of anterior cingulate cortex function. Neuron 79, 217–240 (2013).
Bandura Self-efficacy: toward a unifying theory of behavior change. Am. Polit Sci. Rev. 1, 139–161 (1978).
Shenhav, A., Cohen, J. D. & Botvinick, M. M. Dorsal anterior cingulate cortex and the value of control. Nat. Neurosci. 19, 1286–1291 (2016).
Bartra, O., McGuire, J. T. & Kable, J. W. The valuation system: a coordinate-based meta-analysis of BOLD fMRI experiments examining neural correlates of subjective value. NeuroImage 76, 412–427 (2013).
Elliot, A. J. & Murayama, K. On the measurement of achievement goals: Critique, Illustration, and application. J. Educ. Psychol. 100, 613–628 (2008).
Schwarzer, R. & Jerusalem, M. General Self-Efficacy Scale I. (1995).
Wager, T. D., Davidson, M. L., Hughes, B. L., Lindquist, M. A. & Ochsner, K. N. Prefrontal-subcortical pathways mediating successful emotion regulation. Neuron 59, 1037–1050 (2008).
Wager, T. D. et al. Brain mediators of cardiovascular responses to social threat. NeuroImage 47, 821–835 (2009).
Botvinick, M. & Braver, T. Motivation and cognitive control: from behavior to neural mechanism. Annu. Rev. Psychol. 66, 83–113 (2015).
Amat, J. et al. Medial prefrontal cortex determines how Stressor controllability affects behavior and dorsal raphe nucleus. Nat. Neurosci. 8, 365–371 (2005).
Christianson, J. P., Thompson, B. M., Watkins, L. R. & Maier, S. F. Medial prefrontal cortical activation modulates the impact of controllable and uncontrollable stressor exposure on a social exploration test of anxiety in the rat. Stress 12, 445–450 (2009).
Stevens, D., Anderson, D. I. & O’Dwyer, N. J. Mark Williams, A. does self-efficacy mediate transfer effects in the learning of easy and difficult motor skills? Conscious. Cogn. 21, 1122–1128 (2012).
Brehm, J. W. & Self, A. THE INTENSITY OF MOTIVATION. (1989).
Richter, M., Gendolla, G. H. E. & Wright, R. A. Three Decades of Research on Motivational Intensity Theory: What We Have Learned About Effort and What We Still Don’t Know. In Advances in Motivation Science (ed. Elliot, A. J.) vol. 3 149–186 (Elsevier, 2016).
Berridge, K. C. From prediction error to incentive salience: mesolimbic computation of reward motivation: from prediction error to incentive salience. Eur. J. Neurosci. 35, 1124–1143 (2012).
Vassena, E., Van Opstal, F., Goethals, I. & Verguts, T. Striatal dopamine D2 binding correlates with locus of control: preliminary evidence from [11 C]raclopride Positron Emission Tomography. Int. J. Psychophysiol. 146, 117–124 (2019).
Liljeholm, M., Tricomi, E., O’Doherty, J. P. & Balleine, B. W. Neural correlates of instrumental contingency learning: Differential effects of action-reward conjunction and disjunction. J. Neurosci. 31, 2474–2480 (2011).
Botvinick, M. M., Carter, C. S., Braver, T. S., Barch, D. M. & Cohen, J. D. Conflict Monitoring and Cognitive Control. (2001).
Kerns, J. G. et al. Anterior cingulate conflict monitoring and adjustments in control. Science 303, 1023–1026 (2004).
MacDonald, A. W., Cohen, J. D., Stenger, V. A. & Carter, C. S. Dissociating the role of the Dorsolateral Prefrontal and Anterior Cingulate Cortex in Cognitive Control. Science 288, 1835–1838 (2000).
Shenhav, A., Straccia, M. A., Musslick, S., Cohen, J. D. & Botvinick, M. M. Dissociable neural mechanisms track evidence accumulation for selection of attention versus action. Nat. Commun. 9, 2485 (2018).
Ballesta, S., Shi, W., Conen, K. E. & Padoa-Schioppa, C. Values encoded in orbitofrontal cortex are causally related to economic choices. Nature 588, 450–453 (2020).
Hare, T. A., O’Doherty, J., Camerer, C. F., Schultz, W. & Rangel, A. Dissociating the role of the Orbitofrontal Cortex and the striatum in the computation of goal values and prediction errors. J. Neurosci. 28, 5623–5630 (2008).
Hare, T. A., Camerer, C. F. & Rangel, A. Self-control in decision-making involves modulation of the vmPFC valuation system. Science 324, 646–648 (2009).
Kim, C., Johnson, N. F., Cilles, S. E. & Gold, B. T. Common and distinct mechanisms of cognitive flexibility in Prefrontal Cortex. J. Neurosci. 31, 4771–4779 (2011).
Kolling, N. et al. Value, search, persistence and model updating in anterior cingulate cortex. Nat. Neurosci. 19, 1280–1285 (2016).
Parvizi, J., Rangarajan, V., Shirer, W. R., Desai, N. & Greicius, M. D. The Will to Persevere Induced by Electrical Stimulation of the human cingulate Gyrus. Neuron 80, 1359–1367 (2013).
Blain, B. & Sharot, T. Intrinsic reward: potential cognitive and neural mechanisms. Curr. Opin. Behav. Sci. 39, 113–118 (2021).
Frömer, R., Lin, H., Dean Wolf, C. K., Inzlicht, M. & Shenhav, A. When effort matters: expectations of reward and efficacy guide cognitive control allocation. Preprint at. https://doi.org/10.1101/2020.05.14.095935 (2020).
Shenhav, A. et al. Toward a rational and mechanistic account of Mental Effort. Annu. Rev. Neurosci. 40, 99–124 (2017).
Derryberry, D. & Reed, M. A. Anxiety-related attentional biases and their regulation by attentional control. J. Abnorm. Psychol. 111, 225–236 (2002).
Burger, J. M. & Cooper, H. M. The Desirability of Control. (1979).
Rotter Rotter’s Locus of Control Scale. 10–12 (1966).
Rolls, E. T., Huang, C. C., Lin, C. P., Feng, J. & Joliot, M. Automated anatomical labelling atlas 3. NeuroImage 206, 116189 (2020).
Loonis, R. F., Brincat, S. L., Antzoulatos, E. G. & Miller, E. K. A Meta-analysis suggests different neural correlates for Implicit and Explicit Learning. Neuron 96, 521–534e7 (2017).
Rüsseler, J., Kuhlicke, D. & Münte, T. F. Human error monitoring during implicit and explicit learning of a sensorimotor sequence. Neurosci. Res. 47, 233–240 (2003).
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y. C. & Yantis, S. Cortical Mechanisms of Cognitive Control for Shifting Attention in Vision and Working Memory. J. Cogn. Neurosci. 23, 2905–2919 (2011).
Lepsien, J. & Nobre, A. C. Cognitive control of attention in the human brain: insights from orienting attention to mental representations. Brain Res. 1105, 20–31 (2006).
Bechara, A. Decision making, impulse control and loss of willpower to resist drugs: a neurocognitive perspective. Nat. Neurosci. 8, 1458–1463 (2005).
Frazier, P., Steward, J. & Mortensen, H. Perceived Control and Adjustment to Trauma: A Comparison Across Events. (2004).
Glass, D. C. & McKnight, J. D. Perceived control, depressive symptomatology, and professional burnout: a review of the evidence. Psychol. Health. 11, 23–48 (1996).
Koob, G. F. & Volkow, N. D. Neurobiology of addiction: a neurocircuitry analysis. Lancet Psychiatry. 3, 760–773 (2016).
Romaniuk, L. et al. The Neurobiology of Personal Control during reward learning and its relationship to Mood. Biol. Psychiatry Cogn. Neurosci. Neuroimaging. 4, 190–199 (2019).
Grant, H. & Dweck, C. S. Clarifying achievement goals and their impact. J. Pers. Soc. Psychol. 85, 541–553 (2003).
Peirce, J. et al. PsychoPy2: experiments in behavior made easy. Behav. Res. Methods. 51, 195–203 (2019).
Trommershäuser, J., Gepshtein, S., Maloney, L. T., Landy, M. S. & Banks, M. S. Optimal compensation for changes in task-relevant movement variability. J. Neurosci. 25, 7169–7178 (2005).
Wu, S. W., Delgado, M. R. & Maloney, L. T. Economic decision-making compared with an equivalent motor task. Proc. Natl. Acad. Sci. U S A. 106, 6088–6093 (2009).
Esteban, O. et al. FMRIPrep: a robust preprocessing pipeline for functional MRI. FMRIPrep Robust. Preprocessing Pipeline Funct. MRI. 16, 306951 (2018).
Mumford, J. A., Turner, B. O., Ashby, F. G. & Poldrack, R. A. Deconvolving BOLD activation in event-related designs for multivoxel pattern classification analyses. NeuroImage 59, 2636–2643 (2012).
Kriegeskorte, N., Goebel, R. & Bandettini, P. Information-based functional brain mapping. Proc. Natl. Acad. Sci. 103, 3863–3868 (2006).
Chang, C. C. & Lin, C. J. LIBSVM: a library for support vector machines. ACM Trans. Intell. Syst. Technol. 2, 1–27 (2011).
Eklund, A., Nichols, T. E., Knutsson, H., Erratum & Academy of Sciences of the United States of America. Cluster failure: Why fMRI inferences for spatial extent have inflated false-positive rates (Proceedings of the National 113 (7900–7905) (2016). https://doi.org/10.1073/pnas.1602413113). Proc. Natl. Acad. Sci. U. S. A. 113, E4929 (2016).
Jenkinson, M., Beckmann, C. F., Behrens, T. E., Woolrich, M. W. & Smith, S. M. Fsl NeuroImage 62, 782–790 (2012).
Smith, S. M. et al. Advances in functional and structural MR image analysis and implementation as FSL. NeuroImage 23, S208–S219 (2004).
Acknowledgements
We would like to acknowledge Alec Karousatos, Melanie Roloff and Mary Mousa for their assistance with data collection and Jamil Bhanji for assistance with data analysis. This work was supported by funding from the McKnight Foundation.
Author information
Authors and Affiliations
Contributions
Y.-Y.Y. and M.R.D. designed the study. Y.-Y.Y. and M.R.D. designed the task. Y.-Y.Y. collected the data. Y.-Y.Y. implemented all the analyses and visualization and drafted the paper. Y.-Y.Y. and M.R.D. interpreted the results and revised the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yang, YY., Delgado, M.R. The integration of self-efficacy and response-efficacy in decision making. Sci Rep 15, 1789 (2025). https://doi.org/10.1038/s41598-025-85577-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-85577-z
Keywords
This article is cited by
-
Childhood Trauma and Social Skills in Residential Care Youth: the Roles of Coping Strategies and Self-efficacy
Journal of Child & Adolescent Trauma (2026)
