
Abstract
Alterations in protein glycosylation are observed in many solid tumor types leading to formation of tumor-associated carbohydrate antigens (TACAs). The most common TACA is the Tn antigen (CD175), which is a mucin-type O-GalNAc-Ser/Thr/Tyr glycan in membrane and secreted glycoproteins. In addition, two other TACAs are CA19-9 (sialyl-Lewis a), which is used as a prognostic serum marker for pancreatic cancer, and its isomer sialyl-Lewis x (SLex, CD15s), which is overexpressed in many cancer types and associated with metastasis. While CD175 and other TACAs may be expressed by many human carcinomas, little is known about their differential expression patterns in tumors, thus limiting their use as tissue biomarkers or therapeutic targets. Here we address the clinicopathological relevance of the expression of CA19-9, CD15s, and CD175 in pancreatic ductal adenocarcinoma (PDAC) tissues. Semi-quantitative IHC staining with well-defined monoclonal antibodies demonstrates that CD175 is expressed in all PDAC specimens analyzed. Unexpectedly, however, these TACAs are differentially expressed within PDAC specimens and their glycoproteins, but not significantly expressed in adjacent normal tissues. These data provide avenues for novel therapeutic approaches that could combine CD175- and CA19-9-targeting therapies for PDAC patients.
Similar content being viewed by others


Introduction
Pancreatic ductal adenocarcinoma (PDAC) is the seventh most common cause of cancer related death worldwide, and the fourth leading cause of death in the U.S.1. Surgical resection provides the only potential curative option, however, less than 20% of patients are eligible for surgical resection at the time of presentation2. Post-surgery, the 5-year overall survival rate is 25–30% of those with node-negative disease and only 10% in those with node-positive disease3. PDAC diagnosis maintains a high rate of lethality, as there are no early clinical diagnostic tests available. Furthermore, there is a lack of effective treatment targeting pancreatic cancer despite advances in diagnostic modalities, chemotherapy, and surgery4. PDAC is well known for its late-stage presentation, aggressive clinical behavior, and resistance to the standard therapies. A major causal factor of the mortality rates in PDAC is associated with the lack of reliable biomarkers for pancreatic cancer, as patients are often asymptomatic until they reach an advanced stage.
Glycosylation is a ubiquitous post-translational modification, leading to formation of aberrant glycan structures termed tumor-associated carbohydrate antigens (TACAs), many of which are used as serum and tissue biomarkers in many cancers5,6,7,8,9. Carbohydrates linked to amino acids in glycoproteins and/or lipid moieties in glycolipids have crucial and fundamental roles in basic biological functions, including protein folding, cell–cell adhesion, signaling pathway, and cellular recognition10,11,12. A common post-translational modification is O-glycosylation and formation of O-GalNAc glycans, which is initiated by the enzymatic formation of the Tn antigen (GalNAcα1-O-Ser/Thr/Tyr) (CD175) in the Golgi apparatus. Normal tissues do not express CD175, as it is elongated to form larger oligosaccharides or glycans on cellular glycoproteins, which are membrane bound and secreted (Fig. 1)13. This aberrant CD175 expression has been reported in 75–90% of patients with PDAC, and up to 67% in precursor lesions, and is associated with metastasis and poor prognosis8,14,15,16. Its expression in PDACs is largely due to epigenetic silencing of the X-linked gene COSMC (C1GALT1C1)17, which encodes the molecular chaperone Cosmc that is required for the extension of normal O-glycans through action of the T-synthase that modifies and masks CD17518.

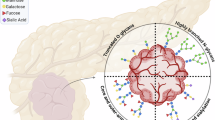
Tumor-associated carbohydrate antigens (TACAs) and their detecting antibodies used in this study. Cartoon showing O-GalNAc glycans including the Tn, CA19-9, and CD15s epitopes and their antibody recognition in red dashed line. pp-GalNAc-Ts, polypeptide-N-acetylgalactosaminyl transferases; T-synthase, core 1 β1,3-galactosyl transferase (C1GalT1); Cosmc, C1GalT1-specific molecular chaperone; C2GnTs, Core 2 β1,6-N-acetylglucosaminyl transferases; GTs, glycosyltransferases. ReBaGs6, anti-Tn antibody; CA19-9, anti-CA19-9 antibody; HECA-452, anti-CD15s antibody. Of note, both CA19-9 and CD15s are also expressed on N-glycans.
Another well-known serum tumor biomarker is CA19-9, also known as sialyl-Lewis a, which can occur in both glycoproteins and glycosphingolipids; its expression, exposure, and secretion are altered in oncogenic cellular transformation19,20,21,22. The CA19-9 epitope can also be present in the normal biliary tree and in acute and chronic biliary tract disease, such as biliary tract obstruction, cholangitis, inflammatory bowel disease, and pancreatitis. On the other hand, an interesting isomer of CA19-9 is sialyl-Lewis x (SLex, CD15s), which occurs normally on leukocytes, but is overexpressed on many cancer types, which commonly promotes cancer metastasis23,24. CD15s is also expressed on vascular endothelial cells and leukocytes only when and where leukocytes extravasate into inflamed tissues25. Both CA19-9 and CD15s are expressed in multiple cancers26,27,28. CA19-9 is clinically used as a serum biomarker for PDAC29,30,31. These TACAs show unique characteristics and have the potential as tissue and serum biomarkers and therapeutics for cancers; however, their clinical application has been limited to therapy response and surveillance for recurrence as opposed to early diagnostics, an area where new tools are greatly needed32,33.
Here, we addressed the clinicopathological expression of CA19-9, CD15s, and CD175 in human PDAC tissues and normal pancreas tissue samples. In this study, CA19-9 and CD15s epitopes were detected with well-characterized antibodies (1116-NS-19-934, and HECA-45235, respectively). Prior investigations of Tn expression in PDAC used lectins such as Vicia villosa agglutinin (VVA) and Helix pomatia agglutinin (HPA) that broadly bind GalNAc, or with antibodies that lack specificity for the CD17515. Some specific anti-Tn antibodies have been identified and confirmed the expression of Tn antigen in other types of carcinomas36,37,38, such as gastric cancers39. Our study utilized a recombinant murine IgM antibody, ReBaGs6 that has high affinity and specificity for CD17540. Semi-quantitative immunohistochemistry (IHC) staining approach demonstrated the significant differential expression patterns for CD175 and other TACAs in PDAC compared to normal pancreas tissue. A major objective of our study was to provide the first detailed molecular understanding of the distribution and expression of these TACAs. We observed that CD175 is expressed in PDAC as well as in premalignant lesions of pancreatic intraepithelial neoplasia (PanIN) and intraductal papillary mucinous neoplasm (IPMN). Interestingly, CD175 and CA19-9 expression differed in location within the same PDAC sample. Overall, our findings provide new insights into TACA expression in PDAC and suggest that CD175 is a potential molecular target for diagnostics and therapy.
Results
Comparative expression patterns of TACAs in PDAC
The O-GalNAc synthetic pathway and formation of the Tn antigen (CD175) is initiated by one of twenty members of the polypeptide N-acetylgalactosaminyltransferases (pp-GalNAc-Ts) (Fig. 1)41. Subsequently, CD175 is modified by T-synthase (C1GalT1), which requires its private chaperone Cosmc (C1GalT1C1) for activity, to form the core 1 O-glycan termed the T (or TF, CD176) antigen, Galβ1-3GalNAcα1-O-Ser/Thr/Tyr)42,43. Core 1 can be further modified to core 2 O-glycans (GlcNAcβ1-6(Galβ1-3)GalNAcα1-O-Ser/Thr/Tyr) by one of three Core 2 GlcNAc transferases (C2GnTs)44,45. Core 2 forms a platform for the elongation of type 1 and 2 chains to polyLacNAc structures (-3Galβ1-3/4GlcNAcβ1-)n, which can carry the fucosylated Lewis antigens46. These Lewis antigens can be sialylated to form sialyl Lewis antigens (i.e. CA19-9, which is Sialyl Lewis a, and CD15s, which is Sialyl Lewis x), expressed in many types of carcinomas. In some cancers, the CD175 is converted by ST6GalNAc-I to the TACA sialyl Tn (STn or CD175s, Neu5Acα2-6GalNAcα1-O-Ser/Thr/Tyr) (Supplementary Fig. S1)7,8.
The immunohistochemical staining (IHC) demonstrated significant expression levels of CD175, CA19-9, and CD15s in PDAC in comparison to normal pancreas (Fig. 2A, B). The majority of PDAC (21/24) had positive CD175 expression (median intracellular H-score = 43 (0–155), extracellular H-score = 44 (0–140)). Intra and extracellular CD175 expression were significantly higher in PDAC when compared to normal pancreas (p < 0.001, p = 0.01). Similar findings were noted with CA19-9 where the majority of PDAC (20/24) had positivity by IHC (median intracellular H-score = 101, extracellular = 66). CD15s demonstrated positivity on IHC (20/24) that was also significant (median intracellular H-score = 58, extracellular = 38) (Fig. 2B, normal, n = 10; PDAC, n = 24).

Comparative expression patterns of TACAs in PDAC. (A) Representative immunohistochemistry (IHC) of benign pancreas, or pancreatic adenocarcinoma (PDAC) stained for Tn, CA19-9, and CD15s antigens. A dark red color indicates the expression of TACAs, and blue represents hematoxylin staining. (B) Comparative H-scores between normal pancreas and PDAC samples for Tn, CA19-9, and CD15s staining (normal, n = 10; PDAC, n = 24, Table 1). P, Kruskal–Wallis one-way analysis of variance (**, p < 0.01). (C) Early pancreatic carcinoma precursor lesions, pancreatic intraepithelial neoplasia (PanIN) and intraductal papillary mucinous neoplasms (IPMN), demonstrate expression of CD175 and CA19-9 (n = 2). All images were captured at 200 × magnification. Scale bars, 100 μm.
We also evaluated the expression of TACAs in PDAC with adjacent normal tissue of the same tissue sections, in which we observed an elevation of those TACAs in PDAC as compared to adjacent normal tissues (Supplementary Fig. S2A and S2B, n = 7, patient-matched). Importantly, the intracellular staining of the CD175 in normal pancreas likely represents immature glycoproteins accumulating during biosynthesis of O-glycans, where CD175 is the normal intermediate biosynthetic structure. We compared the staining patterns between ReBaGs6 and a conventional anti-Tn reagent, the plant lectin VVA, which exhibited a similar staining pattern to ReBaGs6 in PDAC; however, unlike staining with ReBaGs6, we observed some staining of VVA in normal pancreas. This is consistent with VVA cross-reactivity, as it does not specifically bind to CD175 but will bind to any glycan containing terminal GalNAc residues (Supplementary Fig. S3A, normal, n = 10; PDAC, n = 24). There was only low expression of CD175s/STn in PDAC (Supplementary Fig. S3A, normal, n = 10; PDAC, n = 24), which could correlate with higher expression of CA19-9 and CD15s.
Interestingly, there are prior reports of STn expression in PDAC; however, these utilized a different anti-STn antibody (TKH2) from the one used here in our study (B72.3)47. To confirm that the anti-STn antibody used in our study is active and recognizes the CD175s, we performed Western blot analysis using (i) a Cosmc-knockout carcinoma cell line (SimpleCell) that expresses both CD175 and CD175s, (ii) bovine submaxillary mucin (BSM) that expresses both CD175 and CD175s, and (iii) desialylated BSM, which lacks CD175s and expressed more CD175. The results demonstrated the monoclonal antibody B72.3 used in our study actively recognizes CD175s, as it binds as expected to only glycoproteins and cells expressing CD175s (Supplementary Fig. S3B). We further validated the specificity of the B72.3 antibody using a defined glycan microarray, which demonstrated that B72.3 binds only to STn (CD175s) (Supplementary Fig. S3C). Thus, we conclude that there is very low expression of CD175s in PDAC specimens, compared to the high expression of CD175.
Within the tumor specimen samples, we also noted increased expression levels of the CD175 and CA19-9 in precursor lesions such as pancreatic intraepithelial neoplasia (PanIN) and intraductal papillary mucinous neoplasm (IPMN) (Fig. 2C, n = 2, each). Interestingly, the strongest staining for CD175 was observed in early precursor lesions of pancreas, in comparison to the relatively weak staining of CA19-9, suggesting that the CD175 expression may be associated with early tumor progression. In examining the data comparing PDAC tissues from individuals who receive neoadjuvant therapy vs. no therapy (Table 1), in regard to the expression of the Tn, SLea, SLex antigens, we did not observe significant differences.
Unique TACA-carrying glycoprotein profiles in PDAC
To identify whether these TACAs are found in glycoproteins, we used a Western blot approach to analyze the expression profiles of glycoproteins that carry TACAs in PDAC compared to normal pancreas (Fig. 3). In addition, to characterize whether both CA19-9 and CD15s are expressed on N- and/or O-glycans, we treated samples with PNGase F, which releases nearly all N-glycans from glycoproteins, and whose release can be confirmed by the lack of ConA binding which recognizes many types of N-glycans (Supplementary Fig. S4A and S4B). We observed some specific and high molecular weight CD175-carrying glycoproteins in PDAC tissues, but not in normal pancreas (Fig. 3A). Treatment with neuraminidase A, which cleaves all linkages of sialic acids on both N- and O-glycans, did not affect the staining patterns, further confirming that there is little to no expression of STn or CD175s expression, which contains terminal sialic acid and could ‘mask’ the underlying CD175 expression.

Western blots represent unique glycoproteins that carry TACAs in PDAC. Western blots using normal pancreas (24N) and PDAC (39R, 97R, and 103R) tissue extracts with enzymatic treatment with PNGase F (+ PNG), neuraminidase A (+ Neu), or non-treated (NT) analyzed with antibodies to Tn (A), CA19-9 (B), or CD15s (C). Blot images show one representative of two independent experiments with three biological replicates of PDAC (n = 2). See also Supplementary Figure S4 for further information.
For CA19-9 staining, we observed some unique glycoproteins that carry CA19-9 on O-glycans, as its expression was not affected by removal of N-glycans with PNGase F (Fig. 3B). Neuraminidase A treatment eliminated staining of CA19-9, which supports the specificity of anti-CA19-9 antibody for the sialic acid-containing antigen. Lastly, we analyzed the CD15s-carrying glycoproteins in PDAC, and found that some specific bands (− 50 kDa, and − 70 kDa in NT lane in PDAC) carry CD15s epitope on N-glycans, and the high molecular weight bands carry CD15s on both N- and O-glycans, as demonstrated by differential effects of PNGase F treatment on antigen expression (Fig. 3C). No CD15s staining was observed after neuraminidase A treatment, which supports the specificity of anti-CD15s antibody. Thus, we observed that several glycoproteins containing TACAs are overexpressed in PDAC, but not in normal pancreas, suggesting that these antibodies are useful biomarkers for the expression of these glycans in PDAC samples, after fixation, where protein antigens are unaffected, whereas glycolipid and lipid antigens are removed by the fixation process.
Spatial differential expression of TACAs in PDAC with heterogeneous patterns
While we observed the overexpression of TACAs in PDAC, little is known about their differential and spatial expression within the same tumor. From the results in Fig. 3, CA19-9 epitope is expressed on O-GalNAc glycans in glycoproteins, and the staining patterns are strikingly similar to CD175-carrying glycoproteins. We asked, therefore, whether a subset of those glycoproteins express both the CD175 and CA19-9 in a single tumor cell. To address this, we tested for co-staining of these TACAs to determine whether they colocalize within tumor sites. We observed a differential spatial expression between CD175 and CA19-9 within single cells, indicating that cells express one or the other but not both (Fig. 4A, see Supplementary Fig. S5A for further information). It is understandable that expression of CD175, which is a truncated O-glycan, is unlikely to occur in the same cells with extended O-glycans such as CA19-9, based on the biosynthetic pathways shown in Fig. 1. Minimal staining of CD175 and CA19-9 was observed in normal pancreas (Fig. 4B, and Supplementary Fig. S5B). This suggests that a single tumor has a unique set of functional glycosyltransferases to express the distinct TACA epitopes, forming the heterogeneous array of TACAs on each glycoprotein, which presumably could play roles in the different oncogenic properties, and possibly be associated with tumor heterogeneity in PDAC.

Spatial differential expression patterns of TACAs in PDAC. Distinct cellular staining of the CD175 (red), CA19-9 (green), and DAPI (blue) in PDAC (109R, A), or normal pancreas (13N, B) sections by confocal microscope. Both Tn + tumor (#1) and CA19-9 + tumor (#2) as indicated in white dashed box from merge of PDAC staining. See Supplementary Figure S5 for other repetitions. Scale bars, 50 μm.
Discussion
Our studies demonstrate that all PDAC specimens examined express CD175, as well as CA19-9 and CD15s. Importantly, the expression of CD175 and CA19-9 was spatially different within PDACs. Interestingly, both are present largely on O-glycans in multiple tumor-derived glycoproteins that are insensitive to removal by PNGase treatment. These results provide new information about these key TACAs and their potential utility as tissue biomarkers for pancreatic cancer and as therapeutic targets.
CA19-9 (Sialyl Lewis a) is a well-known TACA in gastrointestinal carcinomas as well as for PDACs. However, the expression of CA19-9 is observed in the normal biliary system and in inflammatory diseases such as inflammatory bowel disease and pancreatitis, which limits its utility as a diagnostic serum or tissue biomarker or therapeutic targets in tumors themselves. Additionally, CA19-9 is the sialylated version of the Lewis blood group antigen Lea, but approximately 5–20% of the human population is Lea-negative48, depending on the ethnic group, as their inherited fucosyltransferase (FUT3) lacks activity. In individuals that are Lea-positive, however, elevated serum CA19-9 levels have been observed in late-stage PDAC20,31,49. By contrast, CD175 is widely expressed in many types of human carcinomas, particularly ~ 90% of patients with PDAC consistent with our studies here, and ~ 70% in precursor lesions, but not in normal tissues, as assessed with a variety of anti-Tn/anti-CD175 reagents8. Our study used a specific monoclonal antibody to CD175, but most anti-Tn reagents used historically, such as plant lectins, e.g., VVA, have limited specificity as they cross-react with commonly found terminal GalNAc-expressing glycans, such as blood group A (BGA) antigen, Forssman antigen, and the minor level of Tn antigen sometimes found in glycoforms of human IgA1, potentially creating undesirable false positivity in human carcinomas6,40,50. The antibody ReBaGs6 we used in this study, does not recognize any glycoforms of IgA140.
In our study we addressed the clinicopathological relevance based on TACA expression in PDAC and premalignant lesions (PanIN and IPMN) using well-characterized reagents. All of the TACAs studied were abnormally overexpressed in PDAC, but not in normal/adjacent pancreas (Fig. 2A and B, Supplementary Fig. S2). Surprisingly, CD175 showed higher expression in premalignant lesions of PanIN and IPMN compared to CA19-9 (Fig. 2C), suggesting that anti-CD175 antibody could be useful histologically as a novel early prediction biomarker for PDAC. Our study is not clinical in focus with detailed patient information. Another caveat to the study as it relates to clinical relevance, is that the size and therefore stage of the original tumors was not available. Therefore, we do not have enough information to be able to comment on the staging of the tissues as it relates to the staining performed here, however future studies could incorporate this information.
CA19-9 was originally identified via a unique monoclonal antibody, and was found to be an antigen expressed in colorectal carcinomas and later in pancreatic carcinomas34,51; it, along with CD15s, have been shown to associated with tumor progression in PDAC33,52. Little is known, however, about the differential expression of those TACAs in the same sections in PDAC. In addition, it was unknown whether CA19-9 epitopes are expressed on O-GalNAc glycans in PDAC in which the aberrant CD175 is expressed as well, raising the fundamental question whether these cancer cells express both CA19-9 and the CD175. Therefore, we hypothesized that each single cancer cell expresses either the CD175 or CA19-9, facilitating tumor heterogeneity in PDAC. Our immunohistochemical staining results support this hypothesis where we identified spatial differential expression patterns between CA19-9 and the CD175 within a subset of tumor sites (Fig. 4A, Supplementary Fig. 4A). Furthermore, we found that CD15s in PDACs is expressed on both N- and O-glycans of glycoproteins, whereas CA19-9 is restricted primarily to O-glycans. These data suggest that each cancer cell expresses a rather unique set of glycosyltransferases, playing crucial roles in tumor progression in different ways and potentially influencing the interface with neighboring cancer cells, stromal cells, and immune cells. This finding provides a deeper insight into tumor heterogeneity. The single cell RNA sequence analysis does not account for the spatial expression of TACAs in tissue sections despite laser microdissection to capture CA19-9- and Tn-positive tumor sites. The previous report on exosome sequencing analysis in a whole PDAC tissue showed hypermethylation of the Cosmc gene in ~ 40% of PDAC specimens, which would cause aberrant expression of CD175 in PDAC, which supports our finding17. Another group demonstrated that the diversity of CA19-9 and CD15s expression leads to differentiated tumor malignancy53.
The characterization and identification of cancer specific biomarkers is an unmet need for cancer immunotherapy. A desirable candidate biomarker offers a high specificity, and target abundance in cancers. Several unique carbohydrate antigens including glycoproteins have been developed as biomarkers in the clinical setting, such as CA15-3 (MUC1), CA125 (MUC16), CA72-4 (TAG, or STn), CA19-9 (CA19-9), PSA (prostate specific antigen), CEA (carcinoembryonic antigen), and AFP (alpha fetoprotein), and developing candidates such as Ley, GD2, and GD37,8,9,54,55. In this study, we observed several unique glycoproteins that carry the CD175, CA19-9, and CD15s in PDAC tissues, but not in normal pancreas (Fig. 3). Interestingly, these epitopes are mostly expressed on O-glycans within glycoproteins of high molecular weight, raising the question whether these glycoproteins carry all the TACA epitopes at different positions, or some glycoproteins carry a specific TACA epitope, in a protein-specific manner. The identification of these unique glycoproteins will be further investigated, and the site-specific glycosylation will be elucidated.
It is noteworthy that we detected only low levels of STn (CD175s) relative to Tn (CD175) antigen in the PDAC samples using the immunohistochemical techniques detailed here. The mAb we used for detection of STn was the B72.3 mAb (TAG-72), which we demonstrated binds relatively specifically to the STn antigen on glycan microarrays. In addition, B72.3 mAb bound well to Colo205 cells lacking Cosmc, and thereby expressing both Tn and STn antigens, but did not bind to wild-type Colo205 cells (Figure S3B). We have also used this mAb in multiple studies on colorectal cancer specimens, where we found relatively high expression, along with the Tn antigen56,57. Thus, collectively our results indicate the B72.3 (TAG-72) is specific for STn (CD175s). However, we note that there are other studies which have identified STn expression in PDACs using the B72.3 mAb17,58,59,60, and in some cases using different mAbs61. There are also studies, however, comparing different mAbs to SialylTn, where significant differences are noted in their specificities62. Thus, at this time we conclude that the Tn antigen is highly expressed in PDAC specimens, but the expression of STn antigen needs further study, perhaps using more highly specific reagents.
The data shown here warrants the consideration of these TACAs as multiple targets in PDAC as not only reliable biomarkers but also potential therapeutic targets. While tumor heterogeneity is now well appreciated63, and could cause concern in terms of immunotherapy, various approaches including combination therapies, are being developed to overcome this potential barrier64. The results here suggest that combination therapy with anti-CD175 and anti-CD19-9 might have potential, as well as novel antibodies to be developed that recognize both glycan and adjacent protein epitopes. For CD175 alone this latter possibility has been realized with specific antibodies to mucin polypeptides expressing CD17565, as well as CAR-T type approaches with anti-glycopeptide antibodies66,67,68. In summary, these biomarkers expressed consistently in PDACs provide hope for both diagnostic and therapeutic applications of anti-TACA antibodies.
Materials and methods
Human tissue specimens
Human tissue samples were obtained from the NIH-funded University of Nebraska Medical Center Pancreatic Specialized Program of Research Excellence (SPORE) after receiving institutional IRB approval. Samples included 10 normal pancreas and 25 PDAC fresh frozen tissues, and 10 normal pancreas and 24 PDAC formalin-fixed paraffin-embedded (FFPE) tissue slides (Table 1). These samples were completely de-identified and only the diagnosis of normal tissue, PDAC, metastatic status to the liver, and neoadjuvant chemotherapy were provided as per the Beth Israel Deaconess Medical Center Committee on Clinical Investigations (CCI) Institutional Review Board (IRB)-approved protocols (#2016P000014 and #2018P000381). Informed consent was obtained from all patients as per the IRB protocol at SPORE, and all methods were carried out in accordance with relevant CCI IRB guidelines and regulations.
Immunohistochemistry (IHC)
FFPE tissue sections were deparaffinized by incubation at 60 °C for 45 min followed by using xylene and a descending series of ethanol washes. Antigens were retrieved by boiling in retrieval buffer (10 mM citrate, pH 6.0) under pressurized conditions for 5 min. The subsequent incubations were performed at RT unless otherwise stated. After cooling down, sections were washed in water and treated with 0.3% H2O2 (Cat#H1009, Sigma) in deionized water for 10 min, and blocked with 10% goat serum (Cat#PCN5000, Gibco) in TBS + 0.05% Tween-20 (TBST) for 1 h. Sections were stained with a recombinant mouse anti-Tn (anti-CD175) IgM antibody (ReBaGs6, in house40, diluted to 1 μg/ml), mouse anti-SLea (CA19-9) IgG antibody (11C93, Cat#C0075-03E, VWR, diluted to 2 μg/ml), rat anti-SLex (anti-CD15s) IgM antibody (HECA-452, Cat#555,946, BD Bioscience, diluted to 2 μg/ml) or mouse anti-STn (TAG-72, B72.3) IgG antibody (Cat#sc20042, Santa Cruz Biotechnology, diluted to 2 μg/ml) in TBST overnight at 4 °C. After washing Sections 3 × with TBST for 10 min, horseradish peroxidase (HRP)-labeled goat anti-mouse IgM antibody (Cat#115-035-020, Jackson ImmunoResearch), or goat anti-mouse IgG antibody (Cat#115–035-003, Jackson ImmunoResearch) at 1:200 dilution in TBST were added to sections for 1 h. For CD15s staining, sections were incubated with biotinylated goat anti-rat IgM (Cat#3020-08, Southern Biotech, diluted at 1:200 in TBST) for 1 h, and then incubated with HRP-labeled streptavidin (Cat#SA-5014, Vector Laboratories) at 1:200 dilution in TBST for 1 h. For VVA staining, sections were incubated with biotinylated VVA (Cat#B-1395, Vector labs, diluted to 1 μg/ml in TBST) for 1 h, and then incubated with HRP-labeled streptavidin (Cat#SA-5014, Vector Laboratories) at 1:200 dilution in TBST for 1 h. All staining was visualized using an AEC single solution (Cat#001122, Thermo Fisher Scientific) for 5 min, which develops a deep red deposit where binding occurs in the tissue. All sections were counterstained with hematoxylin (Cat#MHS32, Sigma) for 7 min and incubated in PBS for 15 min, and then cover glass was applied with CLEAR-MOUNT solution (Electron Microscopy Sciences). Isotype mouse IgM (Cat#0101–01, Southern Biotech, diluted to 1 μg/ml), polyclonal mouse IgG (Cat#1033–05, Southern Biotech diluted to 2 μg/ml), or rat IgM (Cat#0120–01, Southern Biotech, diluted to 2 μg/ml) were used as controls. Signals were visualized by incubating sections with aminoethylcarbazole (AEC) substrate solution (Invitrogen), and cell nuclei were counterstained with hematoxylin (Invitrogen). Whole tissue sections were mounted in CLEAR-MOUNT solution.
Imagining and quantification of IHC
Tissue sections were reviewed by microscopy. Slide images were analyzed and photographed using SlideAtlas, a high-performance web-based client–server system for digital pathology at 200 × and 400 × magnification and scored by a clinical pathologist using a combined multiparameter scoring system, the H-score. The H-score is determined by adding the results of multiplication of the percentage of cells with staining intensity (score of 0 for no staining to 3 for strong staining) resulting in a score range from 0 to 300 using the formula: [0 × (% cells 0) + 1 × (% cells 1 +) + 2 × (% cells 2 +) + 3 × (% cells 3 +)]69. Representative images presented at 200 × are shown in Fig. 2 and Supplementary Figures S2 and S3, and representative images at 400 × are show in Supplementary Figure S6.
Tissue homogenization and extraction
Approximately ~ 3 mm3 of fresh frozen PDAC and normal pancreatic tissues stored at -80ºC were thawed on ice, and rinsed 2 × with cold PBS. Tissues were placed in polystyrene round-bottom clear tube, and incubated with 500 μl of cold RIPA lysis buffer (50 mM Tris–HCl, pH 7.4, 150 mM NaCl, 1 mM Na2EDTA, 1 mM EGTA, 1% triton X-100, 0.1% SDS, and 0.5% sodium deoxycholate) with EDTA free protease inhibitor cocktail (Cat#11836170001, Sigma) and DNase I (Cat#10104159001, Sigma, diluted to 10 μg/ml) for 30 min on ice. Tissues were then homogenized using a mechanical homogenizer on ice. Supernatants were collected by centrifugation at 12,000 rpm for 20 min at 4 °C. The concentration of tissue extracts was determined by Pierce BCA protein assay kit (Cat#23225, Thermo Fisher Scientific) with BSA as a standard. Protein extracts were stored in − 80 °C until use.
Enzymatic treatment in tissue extracts
Tissue extracts were incubated with PNGase F (Cat#P0704L) or neuraminidase A (P0722L) following manufacturers’ instruction (New England Biolabs). Briefly, tissue extract (~ 30 μg) was boiled in 1 × Denaturing buffer for 10 min, and incubated with 1 μl of PNGase F in 1 × GlycoBuffer 2 with 1% NP-40, or 1 μl of neuraminidase A in 1 × GlycoBuffer 1 for 4 h at 37 °C.
Western blot
Tissue extract (~ 30 μg/lane) was boiled in Laemmli formulated sample buffer (Cat#1610747, Bio-Rad) containing 2.5% β-mercaptoethanol, and analyzed on SDS-PAGE gel (Cat#M42015, ExpressPlus™ PAGE Gel, 10 × 8, 4–20%, 15 wells, Genscript) at 140 mV for 1 h, and the gel was stained with Coomassie, or transferred to a nitrocellulose membrane (Cat#1704158, Bio-Rad) using Trans-Blot Turbo Transfer System (Bio-Rad). Membranes were blocked with 5% (w/vol) BSA (Cat#BP1600-1, Fraction V, Fisher BioReagents™) in TBST for 1 h at RT. Western blots were analyzed with ReBaGs6 (2 μg/ml), CA19-9 (2 μg/ml), HECA-452 (5 μg/ml), or B72.3 (1 μg/ml) in TBST with 1% BSA as primary staining. Membranes were incubated with HRP-labeled goat anti-mouse IgM, goat anti-mouse IgG, or goat anti-rat IgM antibodies (Cat#3020-05, Southern Biotech) at 1:5,000 dilution in TBST with 0.5% BSA. Signals were developed using SuperSignal™ West Pico Chemiluminescent Substrate (Cat#34578, Thermo Fisher Scientific), then analyzed on an Amersham™ Imager 600 (GE Healthcare Life Sciences).
Confocal microscope imaging
For immunofluorescence with FFPE pancreas tissues, sections were deparaffinized, and antigen-retrieved as described in “Immunohistochemistry”. Sections were treated with 0.3% H2O2 in deionized water for 10 min, and blocked with 10% goat serum in TBST for 1 h at RT. Sections were co-stained with ReBaGs6 (1 μg/ml) and CA19-9 (2 μg/ml) in TBST overnight at 4ºC. After washing sections with 3 × TBST for 10 min, Alexa Fluor 488-labeled goat anti-mouse IgM (Cat#A-21042, Invitrogen, diluted at 1:200 in TBST) and Alexa Fluor 647-labeled goat anti-mouse IgG (Cat#A-21235, Invitrogen, diluted at 1:400 in TBST) were added on sections for 1 h at RT in the dark. Sections were stained with DAPI (0.5 μg/ml) for 10 min at RT in the dark, and then analyzed by confocal microscope (Zeiss; Axioimager Z1). Isotype mouse IgM and IgG (1 and 2 μg/ml, respectively) were used as controls.
Glycan microarray
A CFG glycan microarray (version 5.2) were prepared from amine functionalized glycan structures covalently coupled in microarrays to N-hydroxysuccinimide–derivatized microscope slides as previously described63. Following previously published methods63,64, anti-STn antigen was tested at 5 μg/ml. Binding was detected with Alexa-Fluor 488 labeled anti-mouse IgG secondary antibody at 5 μg/ml. Scanning and quantitation were obtained with ProScanArray scanner (PerkinElmer) and GenePix software. Data are expressed as the mean of relative fluorescence units (RFU).
Statistical analysis
H-scores were analyzed using Kruskal–Wallis, one-way analysis of variance (ANOVA) and Wilcoxon signed-rank test for paired samples. All tests were two-sided, and p-values less than 0.05 were considered statistically significant. All analyses were performed with STATA v.16.1 (College Station, TX).
Statement of significance
Spatial differential expression of CD175 and CA19-9 is seen in pancreatic ductal adenocarcinoma specimens.
Data availability
The datasets used and/or analyzed during the current study are available in the manuscript or from the corresponding author on reasonable request with no restrictions.
References
Stark, A. P. et al. Long-term survival in patients with pancreatic ductal adenocarcinoma. Surgery 159(6), 1520–1527. https://doi.org/10.1016/j.surg.2015.12.024 (2016).
Spanknebel, K. & Conlon, K. C. Advances in the surgical management of pancreatic cancer. Cancer J. 7(4), 312–323 (2001).
Siegel, R. L., Miller, K. D., Fuchs, H. E. & Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 72(1), 7–33. https://doi.org/10.3322/caac.21708 (2022).
Vincent, A., Herman, J., Schulick, R., Hruban, R. H. & Goggins, M. Pancreatic cancer. Lancet 378(9791), 607–620. https://doi.org/10.1016/S0140-6736(10)62307-0 (2011).
Dube, D. H. & Bertozzi, C. R. Glycans in cancer and inflammation–potential for therapeutics and diagnostics. Nat. Rev. Drug Discov. 4(6), 477–488. https://doi.org/10.1038/nrd1751 (2005).
Cervoni, G. E., Cheng, J. J., Stackhouse, K. A., Heimburg-Molinaro, J. & Cummings, R. D. O-glycan recognition and function in mice and human cancers. Biochem. J. 477(8), 1541–1564. https://doi.org/10.1042/BCJ20180103 (2020).
Stowell, S. R., Ju, T. & Cummings, R. D. Protein glycosylation in cancer. Annu. Rev. Pathol. 10, 473–510. https://doi.org/10.1146/annurev-pathol-012414-040438 (2015).
Kudelka, M. R., Ju, T., Heimburg-Molinaro, J. & Cummings, R. D. Simple sugars to complex disease–mucin-type O-glycans in cancer. Adv. Cancer Res. 126, 53–135. https://doi.org/10.1016/bs.acr.2014.11.002 (2015).
Thurin, M. Tumor-associated glycans as targets for immunotherapy: The Wistar Institute experience/legacy. Monoclon. Antib. Immunodiagn. Immunother. 40(3), 89–100. https://doi.org/10.1089/mab.2021.0024 (2021).
Fuster, M. M. & Esko, J. D. The sweet and sour of cancer: Glycans as novel therapeutic targets. Nat. Rev. Cancer 5(7), 526–542. https://doi.org/10.1038/nrc1649 (2005).
Hakomori, S. I. Cell adhesion/recognition and signal transduction through glycosphingolipid microdomain. Glycoconj. J. 17(3–4), 143–151. https://doi.org/10.1023/a:1026524820177 (2000).
Takahashi, M., Kizuka, Y., Ohtsubo, K., Gu, J. & Taniguchi, N. Disease-associated glycans on cell surface proteins. Mol. Aspects Med. 51, 56–70. https://doi.org/10.1016/j.mam.2016.04.008 (2016).
Magalhães, A., Duarte, H. O. & Reis, C. A. The role of O-glycosylation in human disease. Mol. Aspects Med. 79, 100964. https://doi.org/10.1016/j.mam.2021.100964 (2021).
Ju, T. et al. Tn and sialyl-Tn antigens, aberrant O-glycomics as human disease markers. Proteomics Clin Appl. 7(9–10), 618–631. https://doi.org/10.1002/prca.201300024 (2013).
Ju, T., Otto, V. I. & Cummings, R. D. The Tn antigen-structural simplicity and biological complexity. Angew. Chem. Int. Ed. Engl. 50(8), 1770–1791. https://doi.org/10.1002/anie.201002313 (2011).
Schuessler, M. H. et al. Blood group and blood-group-related antigens in normal pancreas and pancreas cancer: Enhanced expression of precursor type 1, Tn and sialyl-Tn in pancreas cancer. Int. J. Cancer 47(2), 180–187. https://doi.org/10.1002/ijc.2910470204 (1991).
Radhakrishnan, P. et al. Immature truncated O-glycophenotype of cancer directly induces oncogenic features. Proc. Natl. Acad. Sci. U. S. A. 111(39), E4066–E4075. https://doi.org/10.1073/pnas.1406619111 (2014).
Ju, T., Aryal, R. P., Kudelka, M. R., Wang, Y. & Cummings, R. D. The Cosmc connection to the Tn antigen in cancer. Cancer Biomark. 14(1), 63–81. https://doi.org/10.3233/CBM-130375 (2014).
Magnani, J. L., Steplewski, Z., Koprowski, H. & Ginsburg, V. Identification of the gastrointestinal and pancreatic cancer-associated antigen detected by monoclonal antibody 19–9 in the sera of patients as a mucin. Cancer Res. 43(11), 5489–5492 (1983).
Goonetilleke, K. S. & Siriwardena, A. K. Systematic review of carbohydrate antigen (CA 19–9) as a biochemical marker in the diagnosis of pancreatic cancer. Eur. J. Surg. Oncol. 33(3), 266–270. https://doi.org/10.1016/j.ejso.2006.10.004 (2007).
Engle, D. D. et al. The glycan CA19-9 promotes pancreatitis and pancreatic cancer in mice. Science 364(6446), 1156–1162. https://doi.org/10.1126/science.aaw3145 (2019).
Magnani, J. L. The discovery, biology, and drug development of sialyl Lea and sialyl Lex. Arch. Biochem. Biophys. 426(2), 122–131. https://doi.org/10.1016/j.abb.2004.04.008 (2004).
Kannagi, R., Izawa, M., Koike, T., Miyazaki, K. & Kimura, N. Carbohydrate-mediated cell adhesion in cancer metastasis and angiogenesis. Cancer Sci. 95(5), 377–384. https://doi.org/10.1111/j.1349-7006.2004.tb03219.x (2004).
Takahashi, S. et al. Overexpression of Sialyl Lewis x antigen is associated with formation of extratumoral venous invasion and predicts postoperative development of massive hepatic metastasis in cases with pancreatic ductal adenocarcinoma. Pathobiology 69(3), 127–135. https://doi.org/10.1159/000048767 (2001).
Lowe, J. B. Selectin ligands, leukocyte trafficking, and fucosyltransferase genes. Kidney Int. 51(5), 1418–1426. https://doi.org/10.1038/ki.1997.194 (1997).
Trinchera, M., Aronica, A. & Dall’Olio, F. Selectin ligands Sialyl-Lewis a and Sialyl-Lewis x in Gastrointestinal Cancers. Biology (Basel) https://doi.org/10.3390/biology6010016 (2017).
Balmaña, M. et al. Identification of potential pancreatic cancer serum markers: Increased Sialyl-Lewis X on ceruloplasmin. Clin. Chim. Acta 442, 56–62. https://doi.org/10.1016/j.cca.2015.01.007 (2015).
Yue, T. et al. Identification of blood-protein carriers of the CA 19–9 antigen and characterization of prevalence in pancreatic diseases. Proteomics 11(18), 3665–3674. https://doi.org/10.1002/pmic.201000827 (2011).
Steinberg, W. The clinical utility of the CA 19–9 tumor-associated antigen. Am. J. Gastroenterol. 85(4), 350–355 (1990).
Lahdenne, P. et al. Tumor markers CA 125 and CA 19–9 in cord blood and during infancy: Developmental changes and use in pediatric germ cell tumors. Pediatr. Res. 38(5), 797–801. https://doi.org/10.1203/00006450-199511000-00026 (1995).
Pour, P. M. et al. Expression of blood group-related antigens ABH, Lewis A, Lewis B, Lewis X, Lewis Y, and CA 19–9 in pancreatic cancer cells in comparison with the patient’s blood group type. Cancer Res. 48(19), 5422–5426 (1988).
Safi, F., Schlosser, W., Kolb, G. & Beger, H. G. Diagnostic value of CA 19–9 in patients with pancreatic cancer and nonspecific gastrointestinal symptoms. J. Gastrointest. Surg. 1(2), 106–112. https://doi.org/10.1016/s1091-255x(97)80097-2 (1997).
Poruk, K. E. et al. The clinical utility of CA 19-9 in pancreatic adenocarcinoma: Diagnostic and prognostic updates. Curr. Mol. Med. 13(3), 340–351. https://doi.org/10.2174/1566524011313030003 (2013).
Koprowski, H. et al. Colorectal carcinoma antigens detected by hybridoma antibodies. Somat. Cell Genet. 5(6), 957–971. https://doi.org/10.1007/BF01542654 (1979).
Duijvestijn, A. M. et al. High endothelial differentiation in human lymphoid and inflammatory tissues defined by monoclonal antibody HECA-452. Am. J. Pathol. 130(1), 147–155 (1988).
Numata, Y. et al. A monoclonal antibody directed to Tn antigen. Biochem. Biophys. Res. Commun. 170(3), 981–985. https://doi.org/10.1016/0006-291x(90)90488-9 (1990).
Takahashi, H. K., Metoki, R. & Hakomori, S. Immunoglobulin G3 monoclonal antibody directed to Tn antigen (tumor-associated alpha-N-acetylgalactosaminyl epitope) that does not cross-react with blood group A antigen. Cancer Res. 48(15), 4361–4367 (1988).
Springer, G. F. T and Tn, general carcinoma autoantigens. Science 224(4654), 1198–1206. https://doi.org/10.1126/science.6729450 (1984).
Persson, N. et al. Epitope mapping of a new anti-Tn antibody detecting gastric cancer cells. Glycobiology 27(7), 635–645. https://doi.org/10.1093/glycob/cwx033 (2017).
Matsumoto, Y. et al. Identification of Tn antigen O-GalNAc-expressing glycoproteins in human carcinomas using novel anti-Tn recombinant antibodies. Glycobiology 30(5), 282–300. https://doi.org/10.1093/glycob/cwz095 (2020).
Bennett, E. P. et al. Control of mucin-type O-glycosylation: A classification of the polypeptide GalNAc-transferase gene family. Glycobiology 22(6), 736–756. https://doi.org/10.1093/glycob/cwr182 (2012).
Ju, T., Brewer, K., D’Souza, A., Cummings, R. D. & Canfield, W. M. Cloning and expression of human core 1 beta1,3-galactosyltransferase. J. Biol. Chem. 277(1), 178–186. https://doi.org/10.1074/jbc.M109060200 (2002).
Ju, T. & Cummings, R. D. A unique molecular chaperone Cosmc required for activity of the mammalian core 1 beta 3-galactosyltransferase. Proc. Natl. Acad. Sci. U. S. A. 99(26), 16613–16618. https://doi.org/10.1073/pnas.262438199 (2002).
Tian, E. & Ten Hagen, K. G. Recent insights into the biological roles of mucin-type O-glycosylation. Glycoconj. J. 26(3), 325–334. https://doi.org/10.1007/s10719-008-9162-4 (2009).
Fukuda, M., Bierhuizen, M. F. & Nakayama, J. Expression cloning of glycosyltransferases. Glycobiology 6(7), 683–689. https://doi.org/10.1093/glycob/6.7.683 (1996).
Kukowska-Latallo, J. F., Larsen, R. D., Nair, R. P. & Lowe, J. B. A cloned human cDNA determines expression of a mouse stage-specific embryonic antigen and the Lewis blood group alpha(1,3/1,4)fucosyltransferase. Genes Dev. 4(8), 1288–1303. https://doi.org/10.1101/gad.4.8.1288 (1990).
Itzkowitz, S. et al. Expression of Tn, sialosyl Tn, and T antigens in human pancreas. Gastroenterology 100(6), 1691–1700. https://doi.org/10.1016/0016-5085(91)90671-7 (1991).
Ramsey, G. et al. The Lewis blood group system in liver transplantation. Transplant. Proc. 19(6), 4591–4594 (1987).
Tempero, M. A. et al. Relationship of carbohydrate antigen 19–9 and Lewis antigens in pancreatic cancer. Cancer Res. 47(20), 5501–5503 (1987).
Matsumoto, Y. Tumor-associated carbohydrate antigens targeting immunotherapy. Trends Glycosci. Glycotechnol. 33(192), E33–E38. https://doi.org/10.4052/tigg.2009.1E (2021).
Koprowski, H., Herlyn, M., Steplewski, Z. & Sears, H. F. Specific antigen in serum of patients with colon carcinoma. Science 212(4490), 53–55. https://doi.org/10.1126/science.6163212 (1981).
Munkley, J. The glycosylation landscape of pancreatic cancer. Oncol. Lett. 17(3), 2569–2575. https://doi.org/10.3892/ol.2019.9885 (2019).
Tang, H. et al. Glycan motif profiling reveals plasma sialyl-lewis x elevations in pancreatic cancers that are negative for sialyl-lewis A. Mol. Cell. Proteom. 14(5), 1323–1333. https://doi.org/10.1074/mcp.M114.047837 (2015).
Yang, J., Tang, A., Ma, J., Sun, X. & Ming, L. The reference intervals for CA125, CA15-3, CA19-9, CA72-4, AFP, CEA, NSE and CYFRA21–1. Scand. J. Clin. Lab. Invest. 79(1–2), 71–74. https://doi.org/10.1080/00365513.2018.1555855 (2019).
Gautam, S. K. et al. Mucins as potential biomarkers for early detection of cancer. Cancers (Basel) https://doi.org/10.3390/cancers15061640 (2023).
Sun, X., Ju, T. & Cummings, R. D. Differential expression of Cosmc, T-synthase and mucins in Tn-positive colorectal cancers. BMC Cancer 18(1), 827. https://doi.org/10.1186/s12885-018-4708-8 (2018).
Dombek, G. E. et al. Immunohistochemical analysis of Tn antigen expression in colorectal adenocarcinoma and precursor lesions. BMC Cancer 22(1), 1281. https://doi.org/10.1186/s12885-022-10376-y (2022).
Abrantes, R. et al. Sialyl-Tn glycan epitope as a target for pancreatic cancer therapies. Front. Oncol. 14, 1466255. https://doi.org/10.3389/fonc.2024.1466255 (2024).
Ching, C. K., Holmes, S. W., Holmes, G. K. & Long, R. G. Comparison of two sialosyl-Tn binding monoclonal antibodies (MLS102 and B72.3) in detecting pancreatic cancer. Gut 34(12), 1722–1725. https://doi.org/10.1136/gut.34.12.1722 (1993).
Rajesh, C. & Radhakrishnan, P. The (Sialyl) Tn antigen: Contributions to immunosuppression in gastrointestinal cancers. Front. Oncol. 12, 1093496. https://doi.org/10.3389/fonc.2022.1093496 (2022).
Romer, T. B. et al. Mapping of truncated O-glycans in cancers of epithelial and non-epithelial origin. Br. J. Cancer 125(9), 1239–1250. https://doi.org/10.1038/s41416-021-01530-7 (2021).
Prendergast, J. M. et al. Novel anti-Sialyl-Tn monoclonal antibodies and antibody-drug conjugates demonstrate tumor specificity and anti-tumor activity. MAbs 9(4), 615–627. https://doi.org/10.1080/19420862.2017.1290752 (2017).
El-Sayes, N., Vito, A. & Mossman, K. Tumor heterogeneity: A great barrier in the age of cancer immunotherapy. Cancers (Basel) https://doi.org/10.3390/cancers13040806 (2021).
Boshuizen, J. & Peeper, D. S. rational cancer treatment combinations: An urgent clinical need. Mol. Cell. 78(6), 1002–1018. https://doi.org/10.1016/j.molcel.2020.05.031 (2020).
Kato, T. et al. A novel Tn antigen epitope-recognizing antibody for MUC1 predicts clinical outcome in patients with primary lung adenocarcinoma. Oncol. Lett. 21(3), 202. https://doi.org/10.3892/ol.2021.12463 (2021).
Ranoa, D. R. E. et al. Single CAR-T cell treatment controls disseminated ovarian cancer in a syngeneic mouse model. J. Immunother. Cancer https://doi.org/10.1136/jitc-2022-006509 (2023).
He, Y. et al. Multiple cancer-specific antigens are targeted by a chimeric antigen receptor on a single cancer cell. JCI Insight https://doi.org/10.1172/jci.insight.130416 (2019).
Posey, A. D. Jr. et al. Engineered CAR T cells targeting the cancer-associated Tn-glycoform of the membrane mucin MUC1 control adenocarcinoma. Immunity 44(6), 1444–1454. https://doi.org/10.1016/j.immuni.2016.05.014 (2016).
Fedchenko, N. & Reifenrath, J. Different approaches for interpretation and reporting of immunohistochemistry analysis results in the bone tissue—a review. Diagn. Pathol. 9, 221. https://doi.org/10.1186/s13000-014-0221-9 (2014).
Acknowledgements
We thank Sandra F. Cummings for technical support.
Funding
The funding was provided by National Heart, Lung, and Blood Institute (No. K12HL14195301, K12HL14195301), and National Institute of General Medical Sciences (No. R24GM137763).
Author information
Authors and Affiliations
Contributions
J.J.C. and Y.M. contributed to the design of the work and executed all experiments.; K.A.S. aided in obtaining the tissue samples; J.N.G. analyzed and provided H-scores on Immunohistochemical staining sections.; J.J.C., Y.M., and R.D.C. wrote the manuscript.; J.H.M. edited the manuscript and reviewed data; G.E.D., K.A.S., A.S.O. contributed to editing of the manuscript. J.J.C. and R.D.C. provided funding. All authors reviewed and approved the manuscript for submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Cheng, J.J., Matsumoto, Y., Dombek, G.E. et al. Differential expression of CD175 and CA19-9 in pancreatic adenocarcinoma. Sci Rep 15, 4177 (2025). https://doi.org/10.1038/s41598-025-86988-8
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-86988-8
