
Abstract
Neonicotinoids are one of the most widely used classes of insecticides in agriculture. They are systemic insecticides mainly used as seed treatments to manage sucking insect pests in a wide range of field crops. Environment and growing conditions can impact the efficacy of neonicotinoid seed treatments. To evaluate this, an experiment was conducted to study the interactive effect of soil, water levels on the neonicotinoid concentration and plant growth in cotton. The experiment was laid out in a split-plot design, with three water levels, 30%, 60%, and 100% (control) of recommended irrigation and four seed treatments, including clothianidin, thiamethoxam, imidacloprid and an untreated control. Cotton seedlings were collected and analyzed for physiological changes and neonicotinoid concentrations at 15, 30 and 45 days after germination (DAG). Data were collected on plant height, fresh biomass and leaf area. The neonicotinoid concentrations were analyzed in leaf tissues using LC-MS/MS. The 30% and 60% of recommended irrigation treatments resulted in significantly taller seedlings with greater biomass and leaf area compared to the 100% water saturation (control) treatment. Thiamethoxam-treated seedlings had greater plant height and shoot fresh mass compared to clothianidin and the untreated control, whereas imidacloprid-treated seedlings had greater leaf area than thiamethoxam, clothianidin, and control at all water levels. 30% recommended irrigation increased neonicotinoid concentrations in leaf tissues, with clothianidin showing higher levels compared to other neonicotinoids at 45 DAG. Neonicotinoid seed treatment efficacy may vary with environmental factors, impacting sustainable pest control.
Similar content being viewed by others

Introduction
Cotton (Gossypium hirsutum L.) holds significant agricultural importance in Texas, particularly in the Texas High Plains (THP) region where up to 40% of the state’s cotton is grown1. Neonicotinoid seed treatments are extensively used against various sucking insects including thrips (Thysanoptera: Thripidae), and aphids (Hemiptera: Aphididae), common early-season pests of cotton2. Neonicotinoids, imidacloprid and thiamethoxam, applied as seed treatments and foliar application are extensively utilized throughout the Texas High Plains3.
These insecticides show moderate persistence in the environment, where in soil, their half-lives could range from a few weeks to several months4. When applied to the seed, a portion of the active neonicotinoid ingredient is transported throughout the entire seedling via vascular tissue (xylem), providing protection against piercing-sucking insects5. The movement of neonicotinoids from the treated seed to the surrounding soil is influenced by irrigation and water flow within the soil5. Seed treatments protect the plant from target pests for approximately 3 to 4 weeks after seedling emergence6,7. Previous studies of neonicotinoid seed treatments in other crops have shown that insecticide concentrations change over time, which could result in changes in efficacy8. As the plant’s biomass increases throughout the growing season, the concentration of neonicotinoids in the plant decreases7. Detection of neonicotinoids active ingredients such as imidacloprid and thiamethoxam in the leaves of plants grown from treated seeds up to 3 to 4 weeks post-planting or beyond suggest that there may be longer term activity and efficacy of these products9. It is also reported that neonicotinoids used as seed treatments persisted in the soil for months post planting and could be present in runoff7.
Neonicotinoids have traditionally been investigated for their efficacy against early-season thrips in cotton fields. However, recent studies have revealed that these substances also exert physiological impacts on plants, influencing their metabolism and structure, thus affecting their overall growth and productivity10. When an agricultural chemical demonstrates such effects on plant metabolism and structure, it falls under the classification of a bio-activator11. Research suggests that neonicotinoid systemic pesticides may bolster plant vigor and offer better protection during drought conditions12. The potential for neonicotinoids to serve as bio-activators has sparked interest in utilizing them, especially to bolster growth during the susceptible stages of seedlings13. For example, thiamethoxam has been observed to function as a bio-activator in rice plants (Oryza sativa L.), influencing photosynthetic pigments, phenylalanine ammonia-lyase activity, root development, and nitrate reductase activity14. Additionally, thiamethoxam seed treatments in soybean (Glycine max L.) enhanced germination under water deficit conditions12. Thiamethoxam and clothianidin effectively mitigated drought stress effects on sugarcane (Saccharum officinarum), resulting in greater total plant weight, increased plant height, and improved photosynthetic activity, along with reduced proline accumulation in drought-stressed plants15. Moreover, thiamethoxam seed treatments boosted root development, protein content, shoot dry matter, and ear dry weight in spring wheat (Triticum aestivum L.) crops10.
Among the most prevalent plant stressors in the Texas High Plains region is moisture stress due to dwindling water resources and increased frequency of drought16. While neonicotinoids are known to increase plant health and vigor, little is known about their persistence and impacts on cotton plant physiology under drought conditions17. Warmer climates constitute the primary regions for cotton cultivation, thereby rendering it susceptible to drought stress at various critical stages throughout its growth cycle18,19. Nevertheless, conditions of water scarcity pose significant threats to sustainable cotton production19. Thus, drought is one of the most detrimental factors constraining plant growth and ultimately crop yield in cotton. Throughout development, growth, and reproduction stages, plants exhibit heightened vulnerability to abiotic stresses20. Cotton has developed diverse morpho-physiological and biochemical mechanisms to counteract drought stress and distinct plant traits serve as key indicators of drought stress tolerance21. Drought stress can impact seedling growth, encompassing parameters such as shoot length, leaf size, leaf area, and dry leaf weight. The reduction in various physiological traits, including photosynthesis and chlorophyll content, becomes evident with increasing plant water stress22,23. The decline in photosynthesis is primarily attributed to leaf relative water content (RWC), and stomatal limitation is also observed20. Consequently, varying moisture levels emerge as primary abiotic stressors defining seedling establishment and early root growth in arid and semi-arid regions across much of the global cotton-growing regions22.
To determine how neonicotinoid seed treatments may act as bioactivators in deficit irrigation, greenhouse studies were conducted to simulate these conditions involving seed treatments and deficit irrigation levels. Neonicotinoid concentrations in cotton leaf tissues were measured in response to varying levels of soil water deficit to determine if neonicotinoids were present and at what concentrations. Plants were also evaluated for growth, biomass, and overall health for better understanding of any bioactivator effects neonicotinoid seed treatments had on cotton seedlings.
Materials and methods
Experimental setup
Staggered replicated experiments were conducted from December 19, 2021, to February 5, 2022 (Trial 1), and from March 3, 2022, to April 20, 2022 (trial 2), at the Texas Tech University Gardens and Greenhouse Complex in Lubbock, Texas, USA (33.583242, -101.882068). Greenhouse conditions were controlled throughout the duration of study and the temperature ranged from 20 to 35°c and relative humidity up to 50%. Supplemental LED lighting was set at 14/10hrs on/off every day. Environmental data including temperature, relative humidity, and photosynthetically active radiation (PAR) were collected periodically throughout the experiments. Both experiments were set up in a split-plot design, with four seed treatments [clothianidin (Poncho Votivo®), imidacloprid (Gaucho®), and thiamethoxam (Cruiser®) and control] and three water deficit levels [no deficit, moderately deficit, and highly deficit corresponding to 100%, 60% and 30% of recommended irrigation rate (250 ml/day)]24. Cotton seeds obtained from Dr. Vyavhare, Deltapine® (DP 1646 B2XF), were treated with each neonicotinoid treatment prior to planting and planted in pots filled with all-purpose soilless media (Berger BM6®). Irrigation in the greenhouse was controlled by a timer to deliver target amount of water per pot at 6-, 8- and 12-hours intervals using 0.5 GPH emitters to maintain adequate soil moisture without any drainage of water or insecticide leaching through the bottom of the pot24. Irrigation timers were tested for 20 min per day based on the needed amount (250, 187 and 125 ml for 100%, 60% and 30%, respectively) to verify that the appropriate amount was being applied per pot. Each treatment had five biological replications seeded, and each trial in the experiment consisted of 180 plants altogether since 60 plants were sampled three times at 15, 30, and 45 days after germination (DAG).
Seed treatments were applied by adding the recommended rate of each chemical to 5 kg of seeds (12.04 mL/kg seed for clothianidin, 9.04 mL/kg seed for imidacloprid, and 8.20 mL/kg seed for thiamethoxam). Seeds were allowed to dry at room temperature for 24 h prior to sowing and one seed per pot was planted.
Plant physiological measurements
Five plants from each treatment were destructively sampled three times at 15 days intervals (15, 30, and 45 DAG), resulting in 60 plants sampled at each sampling date. Plants were cut at the base of the stem, just above the soil. Fresh weight, plant height, and leaf area were measured immediately after cutting each plant. A leaf area scanner (CID Bio-Science, Camas) was used to determine leaf area of all leaves for each plant. Leaf samples were immediately packed in a plastic bag and placed in a small, ice-filled cooler until they could be brought to the laboratory, then frozen at -80 °C until further processing. Plant height was measured by using a standard ruler from the base of the stem to the tip of the apical meristem.
Neonicotinoid concentration analysis
To determine neonicotinoid concentrations in leaves, cotton leaf samples weighing 1 gram each were placed in 50-mL conical tubes, followed by the addition of 25 mL of acetonitrile to each sample. Cotton leaf tissues were homogenized using VWR 200 homogenizer. All samples underwent agitation on a shaker table for 2 h. After this agitation period, the samples were left for 24 h, after which they were centrifuged at 5,000 rpm for 15 min. The decantation process aimed to remove as much acetonitrile as possible from each sample into a 15-mL conical tube, with recorded volumes. Following evaporation to dryness, the samples were reconstituted in one mL of a 1:1 mixture of high-performance liquid chromatography (HPLC)-grade acetonitrile and HPLC-grade water. Quechers salts (Thermo Fisher Scientific Inc., Waltham) were added to sample for extraction as well. The samples were filtered and decanted into syringeless auto-sample vials with a 0.45 μm glass microfiber filter, then subsequently analyzed. Acetamiprid served as a surrogate spike for quality assurance, chosen for its absence in the experiment and, consequently, were not expected in the leaf samples. Each cotton leaf sample underwent spiking with a stock solution of acetamiprid in methanol. Spike-recovery tests conducted on blank plant samples for neonicotinoids indicated that the extraction efficiency was both quantitative and reproducible25. Instrumental analysis was performed using triple-quadrupole liquid chromatography and tandem mass spectrometry [(LC-MS/MS); thermo TSQ Endura: thermo scientific]26.
Statistical analysis
Statistical analysis was performed using JMP Pro 16.0.0 (SAS Institute, Cary, NC) software. Both experiments were analyzed separately because they were conducted at different time of the year in the greenhouse. Data were analyzed for normality using the Shapiro-Wilk test for goodness of fit. When data were normally distributed, analysis of variance (ANOVA) was used to determine significance differences between treatments at p < 0.05. Where significant differences were found, Tukey’s HSD was used to separate treatment means at p < 0.05. Data that were not normally distributed were analyzed using the Kruskal-Wallis Tests to determine significance. When significant differences were found, nonparametric comparisons using the Wilcoxon method were used to compare means. Correlation analysis was performed using Pearson’s pairwise comparison.
Results
Environmental data
In both experiments, there was an increase in temperature over time, with the highest temperatures observed at 45 DAG and the lowest on the 15 DAG (Fig. 1A and B). However, relative humidity levels displayed notable fluctuations throughout the duration of both experiments.

Temperature (°C) profile during the experiments for (A) Trial 1 and (B) Trial 2.
Plant physiological data
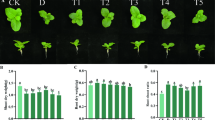
Plant height, shoot biomass, and leaf area increased significantly over time at each sampling day (p < 0.001) in each experiment (Fig. 2). Plant height, shoot biomass, and leaf area were significantly affected by seed treatment and water levels in both trials, however, interaction effects of seed treatment and water levels were not observed for plant height, shoot biomass, and leaf area for both trials (Fig. 2).

Cotton seedling physiological parameters for each trial, irrigation level, sampling day and seed treatment. (A) Plant height at 30% water level; (B) Plant height at 60% water level; (C) Plant height at 100% water level; (D) Plant fresh weight at 30% water level; (E) Plant fresh weight at 60% water level; (F) Plant fresh weight at 100% water; (G) Leaf area at 30% water level ; (H) Leaf area at 60% water level and (I) Leaf area at 100% water level at p ≤ 0.05. Asterisks indicate significant at *p < 0.05, **p < 0.001, ***p ≤ 0.0001. Dashed lines represent trial 1 data and solid lines represent trial 2 data. Error bars represent ± 1 standard error of the mean.
In trial 1, neonicotinoid seed treatments and irrigation levels had no significant effect on plant height at 15 DAG and 30 DAG but the plant height varied with seed treatment x irrigation water interactions at 45 DAG (Fig. 2). At 45 DAG, thiamethoxam and imidacloprid treated seeds produced the tallest seedlings, followed by untreated control and clothianidin treated seeds. Similarly, cotton seedling heights were significantly influenced by seed treatments in 30% and 60% water while plant growth did not vary across seed treatments in 100% moisture regime at 45 DAG (p = 0.02). In trial 2, neonicotinoid seed treatments and water treatments had no significant effect on plant height at 15 DAG and 45 DAG but the height varied at 30 DAG. Clothianidin treated and untreated control plants were taller at 30 DAG followed by imidacloprid and thiamethoxam treated seedlings. Nevertheless, effects of irrigation level were not consistent between trial 1 and trial 2. In trial 2, the 60% irrigation resulted in significantly taller plants than those in 30% irrigation.
Additionally, there were no differences between seed treatments or irrigation levels in fresh plant biomass of seedlings in 15 DAG (trial 1), but irrigation level and seed treatment affected plant biomass at 30 and 45 DAG (Fig. 2). Imidacloprid and thiamethoxam treated seedlings had greater biomass than the clothianidin and control seedlings at 30 DAG and 45 DAG (p ≤ 0.0001 and 0.001, respectively). Interestingly, deficit water level of 30 and 60% of recommended irrigation had significantly greater seedlings biomass at 30 DAG and 45 DAG (p ≤ 0.0001 and 0.001, respectively). In trial 2, the clothianidin treated seedlings were significantly greater in fresh biomass than thiamethoxam after 15 DAG and 30 DAG while there were no significant effects of seed treatments at 45 DAG. Irrigation water level did not exhibit significant treatment effects on seedling biomass in trial 2.
Leaf area was also significantly affected by seed treatments and water level in both trials (Fig. 2). In trial 1, clothianidin treatments had significantly lower leaf area than imidacloprid treated seedlings at 30 and 45 DAG (p = 0.049 and p ≤ 0.0001, respectively). Irrigation treatments also significantly affected seedling leaf area at 30 and 45 DAG, where seedling leaf area increased with increased amount of irrigation water. In trial 2, clothianidin treatments had significantly higher leaf area than thiamethoxam and imidacloprid after 15 and 30 DAG (p = 0.043 and p = 0.003, respectively), while no significant effects of seed treatments or irrigation level were found on leaf area at 45 DAG. In trial 2, there were variable effects of irrigation level on leaf area. At 15 DAG, the 60% water resulted in greater leaf area, followed by 30% and 100% saturation (Fig. 2). However, by 30 DAG the 100% irrigation level treatment resulted in significantly greater leaf area in plant samples. There was no significant effect of irrigation level on leaf area of seedlings at 45 DAG in trial 2.
Neonicotinoid concentrations
Neonicotinoid concentration analysis of cotton seedlings was performed after 15, 30 and 45 DAG for each trial and treatment (Fig. 3).

Neonicotinoid concentrations in cotton seedlings at different sampling dates and water levels in trials 1 and 2. (A) 30% Water level, trial 1; (B) 60% Water level, trial 1; (C) 100% Water level, trial 1; (D) 30% Water level, trial 2; (E) 60% Water level, trial 2; and (F) 100% Water level, trial 2; at p ≤ 0.05. Letters indicate significant differences at *p < 0.05. Error bars represent ± 1 standard error of the mean.
In both trials, the average concentrations of neonicotinoids in cotton leaves varied across water level treatments and sampling dates. Generally, concentrations of all seed treatments declined with time. Data were not normally distributed and therefore, nonparametric analyses were performed. Nonparametric analysis showed no significant differences between neonicotinoid compounds, but there was a significant impact of harvest date on concentrations at 30% irrigation in trial 2. Where 30% irrigation had higher concentrations of neonicotinoids followed by the 60% and 100% treatments (p = 0.008). Overall, clothianidin concentrations were greatest at 15 DAG for 60% irrigation, while concentrations were lower for the 30 and 45 DAG seedlings at 30% and 100% irrigation levels. Thiamethoxam concentration was also greatest in the 15 DAG seedlings for the 30% irrigation level. Notably, the concentrations of neonicotinoids for 30% irrigation were generally higher at 15 DAG compared to that on higher irrigation levels.
Correlation
The relationships between the variables examined in both trials 1 and 2 are clarified by the correlation matrix. irrigation level only significantly affected the final concentration of neonicotinoids in plant tissues, while irrigation level increased neonicotinoid concentration declined in both trials. Furthermore, as temperature increased, the concentration of neonicotinoids decreased. Plant height, fresh weight, and leaf area were positively related to temperature and relative humidity as well as each other in both trials.
Discussion
Environmental data
Environmental variations exert significant impact on the performance of crop chemicals thereby influencing crop’s response to variations in chemical performance. Several contrasting responses observed in our study are attributed to these complex plant-environment relationships. In our study, temperature and relative humidity in the greenhouse exhibited significant fluctuations over the course of both trials, with temperature highest at 45 DAG. The sensitivity of environmental conditions to experiments in greenhouse settings has been well documented27. This highlights the variability in soil responses to experimental conditions and emphasizes the importance of considering site-specific factors in experimental design27,28.
Plant physiological data
Plant height and leaf area are generally reliable indicators of a plant’s health and overall growth29. The delayed growth in cotton plant, reflected by shorter plant height and smaller leaf sizes, due to deficit soil moisture is a direct consequence of the scarcity of irrigation water, which plays a vital role in most metabolic processes essential for the growth and development of plants. Additionally, the lack of water implies a slow import of nutrients necessary for growth30. In this study we saw differing influences of irrigation levels on bioactivator effects of neonicotinoids. This varied by neonicotinoid, irrigation applied, and trials.
The sensitivity of cotton plants to water supply or irrigation volume is well known, where water deficit stress adversely affects various morphological characteristics, including a reduction in cell expansion that impacts leaf and stem growth31. Of the physiological indicators of stress conditions, leaf area is a key component for cotton plants because regulating water balance drives cellular expansion and thus leaf area. Several studies have documented a significant reduction in both leaf area and plant height in cotton plants as a result of water deficit stress32. While there were no significant interactions between seed treatment and irrigation level, each of those factors influenced seedlings separately. Moreover, the leaf area and plant growth parameters varied between the two trials, indicating that the environmental conditions may have influenced the plant parameters, driving the differences between seed treatments and irrigation treatments, also temperature influenced cotton’s water use (evapotranspiration) dynamics33. Variation in fresh shoot biomass and plant height was similar to the trends observed for other parameters as most plant parameters are correlated (Tables 1 and 2).
The three neonicotinoids we tested had a positive impact on cotton plants in terms of plant height, shoot biomass, and leaf area for 45 days, regardless of the amount of soil moisture. Interestingly, stem elongation appears to be more sensitive to the tested neonicotinoids than leaf expansion under water deficit conditions as maximum height was observed at 60% irrigation30,34. However, their significant impact on increasing leaf area was observed only under stressed conditions10. Under optimal irrigation conditions, the neonicotinoids generally increased leaf area and plant vigor at all three sample dates. Specifically, imidacloprid showed growth-promoting effects, such as increased leaf area and plant height followed by clothianidin. However, thiamethoxam was not significantly different from untreated control in influencing plant growth parameters35. Findings of this study are in general agreement with36, who reported similar bioactivator effects of imidacloprid when used as seed treatment cotton seedlings.
Neonicotinoid concentrations
The concentrations of individual neonicotinoids on cotton seedlings were compared across all treatments for all three sample dates. Leaf concentrations of neonicotinoids during the initial sampling surpassed those in subsequent dates, clearly demonstrating the declining amount of neonicotinoid resides on seedling with time. However, it is important to note that concentrations of neonicotinoids were greatest under 30% irrigation level at 15 DAG. This indicates that higher irrigation levels may reduce concentrations and, consequently, the efficacy of neonicotinoid concentrations during early season growth. It is plausible that the availability of sufficient moisture enhanced plant growth and the greater plant growth activity lowered the overall neonicotinoid residues in the tissues. When considering the water level and the duration of neonicotinoid insecticides in the soil-plant system (15, 30 or 45 days after germination), the average concentrations of thiamethoxam tended to be slightly higher in plant tissues than those of imidacloprid and clothianidin. This suggests that thiamethoxam, being more water-soluble, might be more readily translocated through the plant via xylem. However, it is important to acknowledge a substantial amount of sample variation, as evidenced by relatively high standard errors among samples9. Consequently, any interpretation should be approached with awareness of this variability. The concentrations of imidacloprid decreased between 15 DAG and 45 DAG for all treatments. On average, concentrations generally dropped by an order of magnitude to around 250 ppb or lower. At 15 DAG, the 30% irrigation level (high water stress) showed highest concentrations of neonicotinoids. Clothianidin and thiamethoxam was the highest in concentration followed by imidacloprid in both trials7.
When neonicotinoids are administered through seed treatment without additional applications, there is a limited amount of active ingredient for plant uptake. Initially in trial 1, when plants emerged with low biomass in slightly colder environment, the neonicotinoid concentrations were relatively varied in comparison to trial 2. However, as the plants grew and biomass increased, this concentration got diluted or metabolized, leading to a lower concentration of active ingredients in the plant tissue37. The factors affecting neonicotinoid concentrations, like varying water levels and runoff, can vary significantly over time. Given the fluctuating water levels and infrequent major rain events in the Texas High Plains, it would be advantageous to measure neonicotinoid concentrations during both wet and dry seasons and immediately after significant rain events. In a cotton field with neonicotinoid-coated seeds, concentrations are naturally higher in late spring and early summer, corresponding to the planting season and during the winter, when harvesting is complete and fields are inactive, neonicotinoid concentrations would likely be lower7. Seasonal farming practices could also influence the availability of neonicotinoids in runoff. Furthermore, the levels of neonicotinoids in fields can fluctuate from year to year. This variability is likely influenced by the annual weather conditions, such as extremely dry or wet years, which affect runoff and leaching38. Additionally, years with a high prevalence of cotton pests may lead to greater variations in response data. Because of these potential environmental factors, obtaining an accurate representation of neonicotinoid concentrations in cotton leaves is challenging without periodic sampling over time and at different soil types and production regimes.
Correlation analysis
As expected, a strong positive association between plant height and fresh weight indicates that taller plants typically possess higher fresh weights, reflecting robust growth patterns. These findings align with previous studies highlighting the interdependence of plant morphological traits38,39.
Regarding environmental factors and plant responses, positive correlations between temperature and plant height, fresh weight, and leaf area imply that increased temperatures may foster enhanced plant growth and development. This aligns with the physiological responses of plants to temperature variations, where optimal temperatures can promote metabolic processes and biomass accumulation40. Interestingly, the negative correlation observed between the final neonicotinoid concentrations of cotton seedling samples and temperature suggests that higher temperatures may lead to reduced final neonicotinoid concentration levels. This indicates that higher temperatures may accelerate metabolism of these compounds40. Unfortunately, metabolites of the different seed treatments were not examined in this study due to expense and time limitations. However, future research should explore this to further elucidate the complex interactions between environmental factors, plant physiology and the mechanisms underlying temperature-mediated changes in plant metabolism41. In conclusion, the correlation matrix provides valuable insights into the interconnectedness of variables influencing plant growth and environmental responses. These findings contribute to our understanding of plant-environment interactions and have implications for optimizing agricultural practices and ecosystem management strategies.
Conclusion
In this study, we investigated how neonicotinoid insecticides influenced seedling growth, represented by plant height, shoot fresh biomass, and leaf area of cotton seedlings when subjected to water deficit conditions. The results revealed the diverse effects of neonicotinoid seed treatments on the growth and physiology of cotton seedlings. Notably, clothianidin exhibited robust bioactivation effects in trial 2, extending up to 45 days after germination. In trial 1, imidacloprid demonstrated positive effects, although these did not persist until 45 DAG, suggesting potentially long-lasting impacts. Thiamethoxam showed limited effects on growth at 45 DAG, contrasting with findings from other studies12,14,42,43,44. Notably, a 60% irrigation rate had a more pronounced impact on growth parameters, especially leaf area, when coupled with neonicotinoid seed treatments. The observed variability, both within this study and in comparison, to other research, underscores the underexplored nature of this area. Further investigation is warranted to comprehensively elucidate the mechanisms of neonicotinoids as bioactivators under water deficit conditions.
Neonicotinoid seed treatments appear to provide varying durations of pest protection, influenced by water availability and other environmental factors. The discovery of clothianidin and thiamethoxam in the leaves of plants grown from treated seeds, especially under lower water levels for up to 45 days after emergence, indicates successful translocation of the active ingredient through vascular tissues. The claim that seed treatments protect plants for 3 to 4 weeks post-planting seems validated by active ingredient concentrations found in leaf tissues and may even be moderately longer, given the detection of traces 45 days after planting. Reduced water levels may contribute to less leaching of the compounds from the root zone, making neonicotinoid availability higher which could improve the efficacy.
While seed treatments may reduce the total applied active ingredient and mitigate non-target exposure sources like pesticide drift over time in comparison to foliar applications, it is critical to understand that neonicotinoid residues could persist in plant tissues many days after planting. Future research on flowering samples to detect neonicotinoid concentrations in pollen would be useful for assessing whether the recommended application rates of neonicotinoid pesticides as seed treatments genuinely poses a threat to pollinators.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Arnold, M. D., Dever, J. K., Parajulee, M. N., Carroll, S. C. & Flippin, H. D. Simple and effective method for evaluating cotton seedlings for resistance to thrips in a greenhouse, and a thrips species composition on the Texas high plains. Southwest. Entomolog. 37(3), 305–313 (2012).
Elbert, A., Haas, M., Springer, B., Thielert, W. & Nauen, R. Applied aspects of neonicotinoid uses in crop protection. Pest Manag Sci. 64(11), 1099–1105. https://doi.org/10.1002/ps.1616 (2008).
Vyavhare, S. et al. Insect pest management in Texas High Plains cotton. Preprint at (2018). https://esa.confex.com/esa/2018swb/meetingapp.cgi/Paper/129727
Sharma, S. & Singh, B. Metabolism and persistence of Imidacloprid in different types of soils under laboratory conditions. Int. J. Environ. Anal. Chem. 94(11), 1100–1112. https://doi.org/10.1080/03067319.2014.940341 (2014).
Goulson, D. An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 50(4), 977–987 (2013).
Maienfisch, P. et al. Chemistry and biology of Thiamethoxam: A second generation neonicotinoid. Pest Manag Sci. 57(10), 906–913 (2001).
Kohl, K. L. et al. Tracking neonicotinoids following their use as cotton seed treatments. PeerJ 7, 1–16 (2019).
Whalen, A. et al. Temporal profile of neonicotinoid concentrations in cotton, corn, and soybean resulting from insecticidal seed treatments. Agron 11(6), 1200 (2021).
Greenberg, S. M., Liu, T. X., Adamczyk, J. J. & Thrips Thysanoptera: Thripidae) on cotton in the lower Rio Grande Valley of Texas: Species composition, seasonal abundance, damage, and control. Southwest. Entomolog. 34(4), 417–430 (2009).
Macedo, W. R., Castro, P. R. D. C. & Thiamethoxam Molecule moderator of growth, metabolism and production of spring wheat. Pestic Biochem. Physiol. 100, 299–304 (2011).
Castro, P. R. D. C., Serciloto, C. M., Pereira, M. A., Rodrigues, J. L. M. & Rossi, G. Agroquímicos de controle hormonal, Fosfitos e potencial de aplicação de aminoácidos Na agricultura tropical. Piracicaba: Série Produtor Rural 83 (2009).
Cataneo, A. C. et al. Improved germination of soybean seed treated with Thiamethoxam under drought conditions. Seed Sci. Technol. 38(1), 248–251 (2010).
Pynenburg, G. M., Sikkema, P. H. & Gillard, C. L. Agronomic and economic assessment of intensive pest management of dry bean (Phaseolus vulgaris). Crop Prot. 30, 340–348 (2011).
Macedo, W. R., Fernandes, G. M., Possenti, R. A., Lambais, G. R. & Castro, P. R. D.C. Responses in root growth, nitrogen metabolism and nutritional quality in Brachiaria with the use of Thiamethoxam. Acta Physiol. Plant. 35, 205–211 (2013).
Endres, L. et al. Morphological and physiological response of sugarcane under abiotic stress to neonicotinoid insecticides. Theor. Exp. Plant. Physiol. 28, 347–355 (2016).
Bai, Y. et al. Melatonin improves the germination rate of cotton seeds under drought stress by opening pores in the seed coat. PeerJ 8, e9450 (2020).
Ul-Allah, S., Rehman, A., Hussain, M. & Farooq, M. Fiber yield and quality in cotton under drought: Effects and management. Agric. Water Manag 255, 106994 (2021).
Ray, R. L., Fares, A. & Risch, E. Effects of drought on crop production and cropping areas in Texas. Agric. Environ. Lett. 3(1), 170037 (2018).
El-Basyouni, S. A., Massoud, A. H., El-Zahi, E. Z. S. & Al-Kady, M. F. Neonicotinoids insecticides for enhancing cotton plant tolerance against water-deficit stress. Fresenius Environ. Bull. 30(6), 399–409 (2021).
Chastain, D. R. et al. Water deficit in field-grown Gossypium hirsutum primarily limits net photosynthesis by decreasing stomatal conductance, increasing photorespiration, and increasing the ratio of dark respiration to gross photosynthesis. J. Plant. Physiol. 171(17), 1576–1585 (2014).
Lawlor, D. W. & Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant. Cell. Environ. 25(2), 275–294 (2002).
Zhang, L. et al. Effect of drought stress on lipid peroxidation and proline content in cotton roots. J. Anim. Plant. Sci. 24(6), (2014).
Iqbal, M., Khan, M. A., Chattha, W. S., Abdullah, K. & Majeed, A. Comparative evaluation of Gossypium arboreum L. and Gossypium hirsutum L. genotypes for drought tolerance. Plant. Genet. Resour. 17(6), 506–513 (2019).
D’Ambrosio, D. A., Huseth, A. S. & Kennedy, G. G. Temporal efficacy of neonicotinoid seed treatments against Frankliniella fusca on cotton. Pest Manag Sci. 74(9), 2110–2115. https://doi.org/10.1002/ps.4907 (2018).
Nauen, R., Ebbinghaus-Kintscher, U., Salgado, V. L. & Kaussmann, M. Thiamethoxam is a neonicotinoid precursor converted to Clothianidin in insects and plants. Pestic Biochem. Physiol. 76(2), 55–69 (2003).
Peterson, E. M. et al. Agrochemical mixtures detected on wildflowers near cattle feed yards. Environ. Sci. Technol. Lett. 4(6), 216–220. https://doi.org/10.1021/acs.estlett.7b00123 (2017).
Shamshiri, R. R. et al. Review of optimum temperature, humidity, and vapour pressure deficit for microclimate evaluation and control in greenhouse cultivation of tomato: a review. Int. Agrophys. 32, 287–302 (2018).
Hobbs, T., Lynch, J. & Kolka, R. Site-specific critical acid load estimates for forest soils in the Osborn Creek watershed, Michigan. Gen. Tech. Rep. NRS-171. Newtown Square, PA: US Department of Agriculture, Forest Service, Northern Research Station. 171, 1–49 (2017).
Pavlovic, D. et al. Chlorophyll as a measure of plant health: Agroecological aspects. Pestic Fitomed. 29(1), 21–34 (2014).
Gonias, E. D., Oosterhuis, D. M. & Bibi, A. C. Physiologic response of cotton to the insecticide Imidacloprid under high-temperature stress. J. Plant. Growth Regul. 27(1), 77–82. https://doi.org/10.1007/s00344-007-9033-4 (2008).
Jaleel, C. A. et al. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 11, 100–105 (2009).
Shao, H. B., Chu, L. Y., Jaleel, C. A. & Zhao, C. X. Water-deficit stress-induced anatomical changes in higher plants. C R Biol. 331(3), 215–225 (2008).
Li, X. et al. Impacts of growth temperature, water deficit and heatwaves on carbon assimilation and growth of cotton plants (Gossypium hirsutum L). Environ. Exp. Bot. 179, 104204. https://doi.org/10.22541/au.159103674.47131634 (2020).
Smith, E. A. Identifying weed species hosts for onion thrips (Thrips Tabaci Lindeman) and their potential as sources of Iris yellow spot virus (Bunyaviridae: Tospovirus) in New York onion fields. Preprint at (2010). https://ecommons.cornell.edu/bitstreams/d9132c91-3576-40a3-8145-7e44817d5507/download
Preetha, G. & Stanley, J. Influence of neonicotinoid insecticides on the plant growth attributes of cotton and Okra. J. Plant. Nutr. 35(8), 1234–1245. https://doi.org/10.1080/01904167.2012.676134 (2012).
Sehrish, A. et al. Effects of neonicotinoid seed treatments on cotton seedling physiology, nutrition, and growth. Agron 14(4), 799 (2024).
Thompson, D. A. et al. A critical review on the potential impacts of neonicotinoid insecticide use: Current knowledge of environmental fate, toxicity, and implications for human health. Environ. Sci. Process. Impacts 22(6), 1315–1346 (2020).
Wang, Y. et al. Occurrence, Spatial variation, seasonal difference, and risk assessment of neonicotinoid insecticides, selected agriculture fungicides, and their transformation products in the Yangtze river, China: From the upper to lower reaches. Water Res. 247, 120724 (2023).
Zhang, L., Greenberg, S. M., Zhang, Y. & Liu, T. X. Effectiveness of Thiamethoxam and Imidacloprid seed treatments against Bemisia tabaci (Hemiptera: Aleyrodidae) on cotton. Pest Manag Sci. 67(2), 226–232 (2011).
Hilton, M. J., Emburey, S. N., Edwards, P. A., Dougan, C. & Ricketts, D. C. The route and rate of Thiamethoxam soil degradation in laboratory and outdoor incubated tests, and field studies following seed treatments or spray application. Pest Manag Sci. 75(1), 63–78 (2019).
Maurya, J. P. & Bhalerao, R. P. Photoperiod-and temperature-mediated control of growth cessation and dormancy in trees: A molecular perspective. Ann. Bot. 120, 351–360 (2017).
Godói, C. T. D. et al. Thiamethoxam in soybean seed treatment: plant bioactivation and hormesis, besides whitefly control? Sci. Total Environ. 857, 159443 (2023).
Lauxen, L. R., Almeida, A. S., Deuner, C., Meneghello, G. E. & Villela, F. A. Physiological response of cotton seeds treated with Thiamethoxam under heat stress. J. Seed Sci. 38(2), 140–147. https://doi.org/10.1590/2317-1545v38n2159076 (2016).
Macedo, W. R., Araújo, D. K. & de Castro, P. R. C. E. Unravelling the physiologic and metabolic action of Thiamethoxam on rice plants. Pestic Biochem. Physiol. 107(2), 244–249 (2013).
Acknowledgements
This research was supported by a grant through Cotton Incorporated; Texas State Support project no. 20-534TX. The authors would like to acknowledge Carolina Cognasi, Sebastian Espinosa Velasco, Jeremy Mindieta and He Liu for their help during these studies.
Funding
This research was supported by a grant through Cotton Incorporated; Texas State Support project no. 20-534TX.
Author information
Authors and Affiliations
Contributions
Conceptualization, A.S. and C.S.; methodology, C.S., A.S., S.V. and M.G.; formal analysis, A.S., C.C. and C.S.; supervision, C.S.; investigation, A.S., C.S., S.V.; data curation, A.S., and C.S.; writing—original draft preparation, A.S., C.S.; writing—review and editing, A.S., L.H., C.C, S.V., M.P., and C.S; funding acquisition, C.S., M.P. and S.V. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sehrish, A., Vyavhare, S., Parajulee, M. et al. Longevity of neonicotinoid seed treatments in cotton seedlings under various deficit irrigation levels. Sci Rep 15, 6576 (2025). https://doi.org/10.1038/s41598-025-91350-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-91350-z
