
Abstract
Establishing perennial cultivated grasslands on the Qinghai-Tibet Plateau helps address the seasonal imbalance of forage resources and supports the restoration of degraded grasslands. The most common planting patterns—monocropping and mixed cropping—are well-studied in terms of vegetation structure, productivity, and soil nutrients. Despite their significance, the influence of prolonged planting practices on underground soil microbial communities and metabolites has often been neglected. In this study, two characteristic plants, Festuca sinensis ‘Qinghai’ and Poa pratensis ‘Qinghai’, from the area around Qinghai Lake were selected as the experimental subjects by employing 16 S and ITS sequencing methods in conjunction with non-targeted metabolomics analysis. The effects of planting patterns (monocropping and mixed cropping) on rhizosphere soil characteristics, metabolites and microbial community structure were examined. The results showed that compared with monocropping, mixed cropping significantly increased the contents of soil nutrients and key metabolites. In addition, it had a greater impact on fungal diversity than bacterial diversity, particularly in terms of β-diversity. While microbial α-diversity and dominant phyla remained stable, soil fungi were more responsive to changes in soil properties and metabolites. These results show that the new niche differentiation between different species in mixed grassland stimulates the secretion of trehalose and valine, which further affects the fungal community structure and enhances the soil nutrients and ecological functions of degraded grasslands. These findings will guide the restoration of degraded grasslands around Qinghai Lake and the selection of planting strategies to improve local sustainable grassland productivity.
Similar content being viewed by others


Introduction
A complex biological relationship exists between root-zone microorganisms and root cells, which are crucial microhabitats in soil1. Soil, as an indispensable resource influencing plant community growth and development, directly affects plant growth2. The effect of planting patterns on soil nutrients can be attributed to the interaction (competition intensity) between plant species3. With low competition intensity, plant interactions promote the improvement of soil nutrient content mainly through plant litter and debris nutrient return and root exudate deposition. Nevertheless, with increasing competition intensity, the competitive absorption of soil nutrients by plant roots and the negative influence of interspecific root interaction on soil physical and chemical properties gradually offset the increase of soil nutrient content4. Microorganisms play a vital role in matter and energy flow, significantly influencing soil functions and ecosystem services5. In grassland ecosystems, soil microorganisms, mainly bacteria and fungi, are highly sensitive to changes in soil nutrients and are essential for maintaining underground ecosystem functionality6. Studies suggest that planting patterns are key to controlling soil microbial communities and impact their structure, composition, diversity, activity, and symbiotic relationships, and shapes microbial community structure by influencing core microorganisms7,8. Planting patterns had significant influence on microbe structure and composition of the rhizosphere soil9. Compared to monocropping, mixed cropping conditions result in significant changes in rhizosphere bacterial populations10. Zhao et al.11found that the soil bacterial Chao1 and ACE diversity in mixed sowing was significantly lower than in monoculture grassland. Soil bacterial species network analysis revealed a higher complexity and stable of the mixed plot than the monoculture11,12. Zhang et al.12revealed mixed planting significantly increased the rhizosphere microbial diversity and the bacteria abundance related to nitrogen metabolism, and promoted the growth of functional denitrifying flora. The changes of soil physical and chemical properties caused by different planting patterns will significantly affect the soil microbial community13. Wang et al.14indicated that variations in soil bacterial and fungal community compositions across different patterns were attributed to differences in soluble organic carbon and soil bulk density. The production of root metabolites is an important feature of plant adaptation and a key factor in regulating the aggregation of root microbiota, especially through secondary metabolites with biological activity. Numerous root metabolites are released into the rhizosphere, altering the physical and chemical characteristics of the soil, stimulate microbial activity, and bolster the vitality and functionality of microbial communities15. Changes in planting patterns in cultivated grasslands can alter plant coexistence, resulting in differences in soil metabolites, and further affect soil microbial communities11. Duan et al.16 found that the biosynthesis of amino acids, flavonoid, and the flavone and flavonol was significantly affected by intercropping. The expression of metabolites related to amino acid metabolism were up-regulated. Ma et al.17 indicated that cultivation patterns could affect plant flavonoid biosynthesis, while inappropriate cultivation patterns induced stress in plants, prompting the activation of antioxidant mechanisms for flavonoid synthesis as a defence strategy via indirect pathways. Che et al.18 found that under a mixed sowing treatment, the relative abundance of acidobacterium increased significantly due to the increase of organic acid secretion. Lu et al.19 showed that higher carbohydrate content (such as trehalose, mannopyranose, and sucrose) induced the decrease of Chloroflexi and Actinobacteria abundance.
Although metabolomics has been widely applied to crops like wheat, corn, and rice, and research on plant rhizosphere metabolites and their effects on soil nutrient cycling is increasing20, rhizosphere metabolites serve various ecological functions depending on plant-soil interactions in different ecosystems21. Because of the differences of plant species, mixed cropping and monocropping grassland may cause variations in rhizosphere metabolites22, which may play a key role in shaping the construction of rhizosphere microbial community. However, research on how soil metabolites in cultivated grasslands respond to planting patterns and regulate rhizosphere microbial communities remains limited. Thus, it is crucial to examine the effects of planting patterns on soil properties, metabolites, and microbial characteristics in these grasslands.
The area around Qinghai Lake is located in the ecotone between agriculture and animal husbandry in the alpine region of Qinghai-Tibet Plateau in China and belongs to a fragile ecosystem23. Alpine grassland, as the most important ecological function area in this locality, is also a representative geographical unit in this region. Recently, alpine grasslands in Qinghai have experienced severe degradation due to natural and human factors, leading to the “black beach” phenomenon, which has led to relatively low grassland productivity in this area and restricted the development of grassland animal husbandry24. Perennial grassland cultivation has emerged as the primary method for rapidly and effectively restoring these degraded areas and solving the contradiction between grass and livestock25. The most common planting patterns employed are monocropping and mixed cropping26. At present, the research on planting patterns of perennial cultivated grassland mainly focuses on grasses and legumes with distinct niche differentiation10. However, due to the climate in this area, the adaptability of most perennial Leguminosae plants is compromised, leading to slow renewal and easy degradation of perennial grass-legume mixed cropping grasslands27. Therefore, through field experiments, we explored the feasibility of establishing mixed grasslands in this area using two types of grasses with insignificant niche differentiation28. Research on mixed cropping of different grasses has primarily focused on vegetation community structure, productivity, and soil nutrients29. Mixed cropping, compared to monocropping, significantly enhances vegetation yield and biodiversity through resource complementarity and positive species interactions10. However, restoring degraded grasslands requires attention not only to vegetation but also to the soil’s ecological functions11. However, the impact of different grass planting patterns on soil bacterial and fungal communities remains underexplored, and existing findings are inconsistent28,30. Identifying key drivers of microbial community changes requires considering various factors. Therefore, it is necessary to determine the rhizosphere soil properties and metabolites of alpine grassland under different planting patterns, assess their effects on soil microbial communities, and clarify which planting pattern is most helpful in solving the seasonal imbalance of forage resources and restoring degraded grassland.
In this study, perennial grasslands with different planting patterns were selected from the area around Qinghai Lake. 16 S rRNA and ITS gene sequencing and LC-MS analysis were performed. The effects of monocropping and mixed cropping on rhizosphere soil properties, microbial communities, metabolites and their interactions were systematically studied. We hypothesized that planting patterns significantly influence soil properties, metabolites, and microbial communities, and that changes in soil nutrient content and metabolite composition drive shifts in microbial community structure. The study aims to: (1) assess the effects of planting patterns on soil properties and rhizosphere metabolites; (2) compare the impacts of these patterns on bacterial and fungal diversity and community composition; and (3) elucidate the relationships between soil microbial community structure, soil properties, and metabolites under different planting patterns.
Materials and methods
Experiment design
The study was conducted at Bakatai Farm and Pasture (101°5’E, 36°15’N) in Qinghai Province, China, at an average altitude of 3300 m. The region experiences a plateau continental climate with distinct cold and warm seasons and no absolute frost-free period. The growing season is short and warm, while the dormant season is long and cold. Annual sunshine ranges from 2670.4 to 3036 h, with average temperatures between − 5 °C and 5 °C. Precipitation averages about 338 mm per year, with over 75% falling during the warm season. Dominant native vegetation includes Stipa purpurea, Kobresia capillifolia, and Potentilla nivea. The soil type at the site is kastanozems. The experimental plot was established in 2019 using 12 species of perennial grass, and it consisted of 12 monocropping treatments and 6 mixed cropping treatments. Three replicates were performed per plot, and a completely random design was adopted, resulting in 54 experimental plots covering an area of approximately 18 ha. The research objectives of this experiment is only a part of the existing experiments, which are two monocropping treatments and one mixed cropping treatment: Festuca sinensis ‘Qinghai’ monocropping (F), with an area of approximately 1.09 ha and a sowing density of 30.00 kg·ha−1; Poa pratensis ‘Qinghai’ monocropping (P), with an area of approximately 1.00 ha and a sowing density of 11.25 kg·ha−1; and Festuca sinensis ‘Qinghai’ + Poa pratensis ‘Qinghai’ mixed cropping (FP), at a mixed cropping ratio of 1:1, with an area of approximately 0.91 ha and a sowing density of 21.00 kg·ha−1. Mechanical strip sowing was carried out with a row spacing of 15 cm and a sowing depth of 3–5 cm (Fig. S1). Urea and diammonium phosphate were mixed and applied at 75 kg·ha−1 each. Urea topdressing was carried out in May every year, with a fertilization amount of 300 kg·ha−1. The cultivated grassland was not grazed throughout the year. The forage seeds were obtained from the Qinghai Academy of Animal and Veterinary Sciences.
Soil sampling
In August 2022, soil samples were gathered from the pasture during its growing season using a random sampling technique. A thorough excavation of the root system was performed to collect rhizosphere soil, which was defined as soil attached to roots after shaking (rhizosphere soil < 2 mm). The roots of perennial cultivated grassland vegetation are mainly concentrated in the soil surface layer, so the sampling depth was 0–20 cm. After removing loose gravel and plant debris, the samples were processed in three ways: one portion was sieved through a 2 mm mesh and frozen at −80 °C for microbial community and metabolite analysis; another was air-dried for physicochemical testing; and the third was kept at 4 °C for assessing soil ammonium nitrogen (AN), nitrate nitrogen (NN) levels, and microbial biomass.
Determination of soil physicochemical properties
Soil pH was determined by mixing soil with distilled water at a 1:5 ratio, shaking for 35 min, and measuring with a pH meter (METTER TOLEDO)31. Total carbon (TC) and total nitrogen (TN) were assessed using an elemental analyzer (FLASHSMART, Germany)32. Soil organic carbon (SOC) was measured via the K2Cr2O7 oxidation method with FeSO4titration. Ammonium nitrogen (AN) and nitrate nitrogen (NN) were extracted using potassium chloride33. Total phosphorus (TP) was evaluated with the molybdenum-antimony colorimetric method, while available phosphorus (AP) was quantified through sodium bicarbonate extraction followed by molybdenum-antimony colorimetry34. Soil microbial biomass, including carbon (MBC), nitrogen (MBN), and phosphorus (MBP), was measured using the chloroform fumigation-extraction technique35.
High-throughput sequencing
Total DNA from the microbial community was obtained through the Fecal Genome DNA Extraction Kit (DP712-02, TIANGEN, China) according to the manufacturer’s instruction manual36. DNA quality was evaluated through agarose gel electrophoresis, and its concentration was measured with an ultraviolet spectrophotometer. For amplification, the bacterial 16 S rRNA gene’s V3-V4 region was targeted using primers 341 F (5’-CCTACGGGNGGCWGCAG-3’) and 805R (5’-GACTACHVGGGTATCTAATCC-3’), while the fungal ITS2 region was amplified using primers ITS1FI2 (5’-GTGARTCATCGAATCTTTG-3’) and ITS2 (5’-TCCTCCGCTTATTGATATGC-3’)37. The PCR products were purified by AMPure XT beads (Beckman Coulter Genomics, Danvers, MA, USA) and quantified by Qubit (Invitrogen, USA). The amplicon pools were prepared for sequencing and the size and quantity of the amplicon library were assessed on Agilent 2100 Bioanalyzer (Agilent, USA) and with the Library Quantification Kit for Illumina (Kapa Biosciences, Woburn, MA, USA), respectively. The libraries were sequenced on NovaSeq PE250 platform. Paired-end reads was assigned to samples based on their unique barcode and truncated by cutting off the barcode and primer sequence. Paired-end reads were merged using FLASH. Quality filtering on the raw reads were performed under specific filtering conditions to obtain the high-quality clean tags according to the fqtrim (v0.94). The ASV feature sequence and the ASV abundance table were obtained by removing all singleton ASVs. Alpha and Beta diversity analyses were performed using the obtained feature sequences and feature abundance tables of the ASVs. Blast was used for sequence alignment, and the feature sequences were annotated with SILVA database for each representative sequence.
Rhizosphere metabolites analysis
To prepare the samples, 100 mg of soil was weighed and ground in liquid nitrogen. Metabolites were then extracted with a 50% methanol solution. After adding 1 ml of chilled 50% methanol, the mixture was shaken at room temperature for 10 min and stored at − 20 °C overnight. Following centrifugation at 4000 g for 20 min, the supernatant was transferred to a new 96-well plate for analysis. Samples not tested immediately were kept at − 80 °C until LC-MS analysis. Additionally, 10 µl from each sample was combined to form a QC sample. All samples underwent LC-MS non-targeted metabolomics analysis (Appendix S2). Sequencing for both microbial and metabolome analyses was performed by LC-Bio Technology Co., Ltd., located in Hangzhou, Zhejiang Province, China.
Statistical analyses
A one-way ANOVA was used to evaluate differences in soil physicochemical properties, taxonomic abundances, α-diversity indices, and rhizosphere metabolites among different planting patterns (P < 0.05). To explore the relationships between varying metabolites and soil properties, Pearson correlation analysis was employed. Metabolite profiles were examined using Partial Least Squares Discriminant Analysis (PLS-DA). Non-metric Multidimensional Scaling (NMDS) based on Bray-Curtis distance was utilized to assess microbial β-diversity differences across treatments. PERMANOVA statistical model was used to test the microbial community structure under different planting methods. Redundancy Analysis (RDA) with the “envfit” function from the ‘vegan’ package identified key environmental factors affecting microbial community changes. The contributions of soil nutrients, metabolites, and their interactions with shifts in microbial community structure were analyzed using a linear mixed-effects model based on REML from the “glmm.hp” package. All statistical computations were carried out with R software (version 4.0.2).
Results
Impact of planting patterns on soil properties and metabolites
Planting patterns had a significant impact on soil chemical properties (Fig. 1). Specifically, mixed cropping notably reduced TC content (P < 0.05), with decreases of 4.41% and 3.09% compared to monocultures of F and P, respectively (Fig. 1B). Conversely, mixed cropping significantly increased levels of TP, AN, NN, SOC, AP, MBC, MBN, and MBP (P < 0.01). TP ranged from 0.74 to 0.84, with increases of 12.73% and 12.05% compared to monocultures of F and P, respectively (P < 0.01) (Fig. 1D). AN ranged from 23.27 to 26.52, with increases of 13.95% and 11.73% compared to monocultures of F and P, respectively (P < 0.01) (Fig. 1E). NN ranged from 13.33 to 13.97, with increases of 4.75% and 4.10% compared to monocultures of F and P, respectively (P < 0.01) (Fig. 1F). SOC ranged from 12.46 to 14.19, with increases of 13.88% and 11.67% compared to monocultures of F and P, respectively (P < 0.01) (Fig. 1G). AP ranged from 2.46 to 3.14, with increases of 28.07% and 23.20% compared to monocultures of F and P, respectively (P < 0.01) (Fig. 1H). MBC ranged from 86.94 to 99.07, with increases of 13.95% and 11.72% compared to monocultures of F and P, respectively (P < 0.01) (Fig. 1I). MBN ranged from 3.98 to 4.54, with increases of 13.95% and 11.72% compared to monocultures of F and P, respectively (P < 0.01) (Fig. 1J). MBP ranged from 1.39 to 1.58, with increases of 13.64% and 11.47% compared to monocultures of F and P, respectively (P < 0.01) (Fig. 1K). Mixed cropping had minimal effects on soil pH and TN (P > 0.05) (Fig. 1A and C).

Impact of planting patterns on soil properties. Different lowercase letters indicated significant differences between different treatments. One-way analysis of variance was used for significance labeling results between different treatments. (A) pH: pH contents; (B) TC: total carbon; (C) TN: total nitrogen; (D) TP: total phosphorus; (E) AN: ammonia nitrogen; (F) NN: nitrate nitrogen; (G) SOC: soil organic carbon; (H) AP: available phosphorus; (I) MBC: microbial biomass carbon; (J) MBN: microbial biomass nitrogen; (K) MBP: microbial biomass phosphorus.
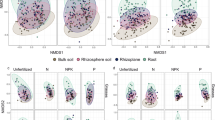
Planting patterns had a significant impact on metabolites. In the rhizosphere of perennially cultivated grasslands, 11,157 metabolites were identified, with 286 classified as secondary metabolites. These secondary metabolites were categorized into 44 groups. The categories with a total proportion greater than 1% included: fatty acyls (20.4%); carboxylic acids and derivatives (15.3%); organooxygen compounds (12.8%); benzene and its derivatives (9.8%); glycerophospholipids (6%); prenol lipids (6%); imidazopyrimidines (3.4%); organonitrogen compounds (3%); organic phosphoric acids and derivatives (2.1%); phenols (1.7%); and undefined (19.6%) (Fig. 2A). NMDS analysis of the perennial grasslands showed significant differences in rhizosphere metabolites across different planting patterns (Adonis: R² = 0.19, P < 0.01; ANOSIM: R² = 0.32, P < 0.01; Fig. 2B).

Soil metabolomics analysis under different planting patterns. (A) The classification profile of 286 metabolites; (B) NMDS analysis of metabolites in perennial cultivated grassland.
PLS-DA analysis indicated a clear separation of metabolites between treatments (R²X = 0.555, R²Y = 0.984, Q² = 0.422) (Fig. S2). A one-way ANOVA of the top 20 rhizosphere metabolites showed that planting patterns significantly affected lower-content metabolites but had minimal impact on the top 20, only altering the levels of Trehalose, Valine, and Phenylglyoxylic acid (Fig. S3). Specifically, mixed cropping significantly increased Trehalose and Valine levels (P < 0.05). Trehalose content increased by 67.71% and 40.68% compared to monocultures of F and P, respectively (Fig. 3A). Valine content increased by 46.38% and 20.22% compared to monocultures of F and P, respectively (Fig. 3B). In contrast, mixed cropping significantly decreased Phenylglyoxylic acid content by 2.14% and 4.29% compared to monocultures of F and P, respectively (Fig. 3C).

Impact of planting patterns on metabolites. F: Festuca sinensis ‘Qinghai’ monocropping; P: Poa pratensis ‘Qinghai’ monocropping; FP: Festuca sinensis ‘Qinghai’ + Poa pratensis ‘Qinghai’ mixed cropping cultivated grassland.
Impact of planting patterns on the correlation between soil properties and differential metabolites
A correlation analysis between soil physicochemical properties and differential metabolites (Fig. 4) revealed that Valine, Trehalose, and soil nutrients (TP, AN, NN, SOC, AP, MBC, MBN, and MBP) exhibited significant positive correlations (P < 0.01), while Valine and TC showed a negative correlation (P < 0.05). Phenylglyoxylic acid, however, did not exhibit a significant correlation with soil physicochemical properties.

Impact of planting patterns on the correlation between soil properties and differential metabolites. * Correlation is significant at the 0.05 level; ** Correlation is significant at the 0.01 level; *** Correlation is significant at the 0.001 level. pH: pH contents; TC: total carbon; TN: total nitrogen; TP: total phosphorus; AN: ammonia nitrogen; NN: nitrate nitrogen; SOC: soil organic carbon; AP: available phosphorus; MBC: microbial biomass carbon; MBN: microbial biomass nitrogen; MBP: microbial biomass phosphorus.
Impact of planting patterns on microbial communities
Microbial diversity under different planting patterns was assessed using ASV levels. The results showed that α diversity (observed_species, Shannon, and Chao1 indices) of soil bacteria and fungi was not significantly influenced by planting patterns (Fig. 5). NMDS analysis results showed that the planting patterns had inconsistent effects on microbial β-diversity. The bacterial community structure did not show significant differences under different planting patterns, while the fungal community structure had significant differences (Fig. 6).

Impact of planting patterns on α diversity of microorganisms (bacterial and fungal). F: Festuca sinensis ‘Qinghai’ monocropping; P: Poa pratensis ‘Qinghai’ monocropping; FP: Festuca sinensis ‘Qinghai’ + Poa pratensis ‘Qinghai’ mixed cropping cultivated grassland.

Impact of planting patterns on β diversity of microorganisms (bacterial and fungal).
The planting patterns had no impact on the microbial community composition of the perennial cultivated grassland. At the phylum level, the bacterial community was classified into 42 phyla (Fig. S4), with Actinobacteria, Proteobacteria, Acidobacteria, Chloroflexi, and Gemmatimonadetes being the dominant groups, comprising over 80% of the total sequences. The relative abundances of these five bacterial phyla remained consistent across different planting patterns. The fungal community was classified into 13 phyla, with Ascomycota, Basidiomycota, Zygomycota, Fungi_unclassified, Glomeromycota, and Chytridiomycota being the dominant phyla, accounting for over 98% of the total relative abundance. Similarly, the relative abundances of these dominant fungal phyla were stable across planting patterns.
Impact of planting patterns on correlation of soil properties, metabolites and microbial communities structure
RDA revealed that bacterial and fungal community structures responded differently to changes in soil properties and rhizosphere metabolites (Table S1, Fig. 7). The first two axes of the RDA accounted for 47.20% of the total variation in bacterial communities (PC1 = 30.40%, PC2 = 16.80%). The bacterial community structure was influenced by soil properties such as TP, SOC, AN, and AP, as well as metabolites like Trehalose and Valine. For fungal communities, the first axis of the RDA explained 20.29% of the total variation, while the second axis explained 17.40%. Soil properties including TC, TP, SOC, AN, and AP, along with metabolites such as Trehalose, Valine, and Phenylglyoxylic acid, were significant environmental factors affecting fungal communities (P < 0.05, Fig. 7).

Redundancy analysis was performed using the aggregated data of bacterial and fungal communities and abiotic and biotic variables (arrows). pH: pH contents; TC: total carbon; TN: total nitrogen; TP: total phosphorus; AN: Available nitrogen; SOC: soil organic carbon; AP: available phosphorus.
The mixed-effects model further showed that soil properties and metabolites have a similar impact on bacterial community structure. The influence of soil properties on bacterial community accounted for 30.51%, and the influence of metabolites on bacterial community accounted for 34.40%. The interaction between soil properties and metabolites explained 35.1% of the differences in bacterial community structure. In contrast, for fungal community structure, changes are primarily influenced by metabolites rather than soil properties. The effect of soil properties on fungal community accounted for 73.24%, and the effect of metabolites on fungal community accounted for 8.90%. The interaction between soil properties and metabolites explained 17.86% of the differences in fungal community structure (Fig. 8).

The contribution of soil properties, metabolites, and their interaction to the changes in microbial community structure.
Discussion
Impact of planting patterns on soil properties and metabolites
Soil physical and chemical properties are key indicators of soil quality and essential for maintaining plant productivity. A decline in these properties is characteristic of degraded grasslands. However, cultivating grasslands in such areas can effectively restore soil conditions and support sustainable production38. Research indicates that different vegetation restoration methods have a significant impact on soil quality, with cultivated grasslands consistently showing better soil quality compared to naturally restored areas39. Different planting patterns have different ecological impacts on soil40. Mixed cropping, compared to monocropping, results in more complex canopy structures that better resist erosive rainfall41. Jiang and Niu28found that forage mixed cropping reduced soil pH and TN but increased TP. Our study found that while different planting patterns did not significantly alter soil pH and TN, they notably affected SOC, TP, AN, NN, AP, MBC, MBN, and MBP. Mixed cropping demonstrated greater benefits for soil quality compared to monocropping. In mixed cropping grasslands, plant root systems may occupy different soil depths and zones, reducing competition and improving nutrient uptake. Higher root biomass in mixed cropping systems contributes more nutrients to the soil, and increased plant diversity enhances nutrient accumulation through litter and detritus decomposition11,42. These findings contrast with previous studies, which may be influenced by factors such as grass varieties, planting duration, and regional conditions43.
Rhizosphere metabolites are of great significance in regulating the interaction between plants and microorganisms44. Plants produce unique metabolites45, and their rhizospheres often adjust metabolite production in response to environmental changes, leading to species-specific interactions46. These metabolites, key indicators of biochemical activity, include Trehalose, a stress-related disaccharide that provides energy and protection to plant cells under adverse conditions such as extreme cold and high UV radiation47. Valine, another important metabolite, acts as a signaling molecule that supports plant growth and regulates nitrogen balance48. Recent studies highlight significant differences in root metabolite profiles across various planting patterns. Li et al.49 discovered that continuous planting alters the accumulation of soil metabolites, significantly impacting lipids, organooxygen compounds, and carboxylic acids. Similarly, Feng et al.50 observed that planting patterns influence the buildup of highly interconnected amino acids and lipid metabolites within symbiotic networks.
Our study found that planting patterns significantly influence rhizosphere metabolites, with mixed cropping markedly increasing Trehalose and Valine compared to monocropping. This increase is likely due to changes in carbohydrate, amino acid, and organic acid content caused by interactions among plant roots. Higher SOC in mixed cropping may explain the elevated levels of Trehalose and Valine, as carbon compounds enhance microbial activity, leading to greater production of microbial metabolites and polysaccharides51. Metabolites not only supply nutrients to microorganisms but also alter rhizosphere properties, impacting nutrient status52. Additionally, plant-secreted metabolites influence soil microbial communities and nutrient status53. Lin54found significant correlations between metabolites and soil SOC, TC, TP, AN, NN, and AP, but not soil pH, while Ma55 noted a strong association between root-secreted substances and TN, TC, MBC, and MBP. Peng et al.56reported that SOM, pH, NN, and AP significantly affected Ginkgo biloba’s secondary metabolites. Our study, however, found Trehalose and Valine content to be significantly correlated with TP, SOC, AN, NN, and AP, which may be due to differences in study locations and plant species57.
Impact of planting patterns on microbial communities
Soil microorganisms are vital indicators of soil health and play a key role in biogeochemical cycles, organic matter decomposition, and plant development8. Studies have shown that soil management practices and plant species have a significant impact on microbial community structures. Planting patterns influence microbial communities differently, with varying sensitivities to these patterns18. For instance, mixed cropping with grasses and legumes has been found to enhance soil bacterial diversity and abundance compared to monoculture systems10. Chen et al.58reported increased microbial diversity in mixed cropping systems, though short-term studies suggest minimal changes in bacterial diversity across planting patterns30. In alpine grasslands on the Qinghai-Tibet Plateau, mixed cropping may reduce microbial diversity, particularly affecting bacterial richness without substantially altering fungal diversity. Jiang and Niu28 observed that seeding methods influence the abundance of key soil bacteria, with mixed cropping decreasing Acidobacteria and Bacteroidetes while increasing Actinomycetes and Proteobacteria, although fungal group abundance remained relatively stable. In contrast, Zhang et al.59found no significant differences in bacterial communities or the abundance of dominant bacterial and fungal phyla between monoculture and mixed cropping of perennial grasses. Plant traits and soil conditions create specific niches that shape microbial communities60. The similar traits and small niche differences among dominant grass species in this study might suggest that mixed cropping effects are more about species richness than distinct species variations61. Although soil pH—a key factor affecting microbial communities62—was not significantly influenced by planting methods in our study, indicating a complex interplay of pH and nutrient effects63, our results still highlighted significant impacts of planting patterns on microbial community structures. Consistent with previous research, different planting patterns notably affect soil bacterial and fungal communities40. Chen et al.58found that mixed cropping systems led to higher total, bacterial, and fungal biomass, along with increased below-ground biomass and root activity, which enhances soil rhizobium deposition and carbon flux. This can improve soil nutrient availability and exchange capacity. The increased plant diversity in mixed cropping systems potentially boosts plant productivity and influences soil microbial communities64. Furthermore, plant-produced metabolites can affect interspecific competition and microbial community composition65,66. Despite variations in microbial community composition across planting patterns, dominant soil bacterial and fungal species showed minimal differences between mixed and monocropping. It appears that plant diversity impacts rare microbial species more than dominant ones, suggesting that mixed cropping may alter microbial community structure by affecting the abundance of certain rare bacterial and fungal taxa28.
Impact of planting patterns on correlation of soil properties, metabolites and microbial community structure
There is a complex interplay between plant rhizosphere microecology and the evolving dynamics of soil microorganisms and their metabolites. Rhizosphere microbial communities are shaped by various ecological factors, including soil pH, moisture, salt concentrations, soil type, and metabolites. These metabolites, originating from both microorganisms and plants, are crucial in influencing the soil microbiome by affecting microbial food webs, soil chemistry, gene expression, and interactions among microbes67. Soil type significantly determines microbial communities, with soil AN and plant aboveground biomass being key drivers for bacterial communities, while plant TP, AP, and the MBC to MBN ratio primarily affect fungal communities68. Although Che et al.18 found a notable correlation between SOM and TN with bacterial community composition, no significant correlation was observed with fungal communities.
Different planting patterns modify soil physicochemical properties, which in turn impact microbial community structure. Our findings revealed significant responses of soil bacterial and fungal β-diversity to soil nutrients (TP, SOC, AN, NN, AP) and variations in microbial β-diversity across planting patterns, aligning with earlier studies18,33,68. The diversity and abundance of rhizosphere microbes are directly affected by root metabolites, which vary with plant species, genotypes, and environmental conditions69. For instance, metabolites from Mikania micrantha and Centaurea diffusa roots impact microbial diversity by altering soil microbial communities70. Drought and heat stress (DHS) stimulate corn roots to secrete L-valine, which promotes the recruitment of Gemmatimonadota and enhances root uptake of nitrate and ammonium under DHS71, This is different from our research results, because these plants are vine and knapweed, and our research object is perennial grass. Lopes and Schachtman72noted that sugars and gibberellins are critical in shaping rhizosphere bacterial communities, especially during early plant growth. Due to niche differentiation among different species in mixed cropping grasslands, the niche of the same species may vary between mixed cropping and monocropping, leading to differences in rhizosphere metabolites and further influencing the structure of the soil microbial community. Notably, the relationship between fungal communities and differential metabolites in perennial cultivated grasslands is stronger than that of bacterial communities. We speculate that the metabolites in cultivated grasslands have a greater impact on fungal communities than on bacterial ones. Fungal and bacterial communities engage in continuous chemical dialogue, which prompts bacteria to produce more diverse and heterogeneous metabolites, thereby altering the overall metabolic profile of the rhizosphere soil. Alternatively, it may be that these metabolites recruit specific bacteria, whose chemical signals in turn have a more significant effect on the structure and diversity of fungal communities. These hypotheses point to the interaction mechanisms between bacteria, fungi, and plants, which require further validation through more precise experiments73.
Amino acids and sugars, which are rich in carbon and nitrogen, greatly influence soil microbial community structure. For example, Blastococcus and Nocardioides, as well as metabolites like Valine, positively correlate with soil health and ecosystem functionality, while Trehalose and Valine are vital for plant growth15. Overall, planting patterns have a lesser effect on soil bacterial β-diversity compared to fungal β-diversity, with varying responses driven by metabolites. The influence of planting patterns on soil microorganisms may involve direct interactions or less understood pathways, potentially through soil enzyme activities and bacterial-feeding nematodes. Future studies should investigate these indirect mechanisms and explore the interactions among plants, microbes, and soil nematodes under different environmental conditions.
Conclusions
This study provides a comprehensive analysis of how different planting patterns affect soil properties, microbial communities, and metabolites in cultivated grasslands of alpine regions, offering new theoretical insights. The results demonstrate that mixed cropping significantly enhances rhizosphere soil nutrients and metabolites compared to monocropping, with a strong correlation between soil properties and metabolites. Changes in TP, SOC, AN, AP, Trehalose, and Valine are key predictors of bacterial community structure, while TC, TP, SOC, AN, AP, Trehalose, Valine, and Phenylglyoxylic acid are crucial for predicting fungal community structure. The mixed-effects model further reveals that fungal community responses to planting patterns are more dependent on metabolites. Overall, this research quantifies the efficacy of different planting patterns on soil ecosystems and emphasizes that mixed cropping is more effective than monocropping. Therefore, we suggest prioritizing mixed cropping patterns in ecological restoration projects for degraded grasslands.
Data availability
The datasets of soil properties and metabolites generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. The raw sequence data of microorganisms reported in this paper have been deposited in the Genome Sequence Archive (GSA) database under accession numbers CRA016422 and CRA016429.
References
Kuzyakov, Y. & Razavi, B. S. Rhizosphere size and shape: Temporal dynamics and Spatial stationarity. Soil Biol. Biochem. 135, 343–360 (2019).
Guo, J. H. et al. Species mixing improves soil properties and enzymatic activities in Chinese Fir plantations: A meta-analysis. CATENA 220, 106723 (2023).
Sun, Y. et al. Plant intraspecific competition and growth stage alter carbon and nitrogen mineralization in the rhizosphere. Plant. Cell. Environ. 44 (4), 1231–1242 (2021).
Xiao, L., Min, X. X., Liu, G. B., Li, P. & Sha, X. Effect of plant–plant interactions and drought stress on the response of soil nutrient contents, enzyme activities and microbial metabolic limitations. Appl. Soil. Ecol. 181, 104666 (2023).
Rillig, M. C. et al. The role of multiple global change factors in driving soil functions and microbial biodiversity. Science 366 (6467), 886–890 (2019).
Li, D. et al. Imidacloprid-induced stress affects the growth of pepper plants by disrupting rhizosphere-plant microbial and metabolite composition. Sci. Total Environ. 898, 165395 (2023).
Zhou, B. et al. Effects of phthalate esters on soil microbial community under different planting patterns in Northern China: case study of Hebei Province. Chemosphere 307 (2), 135882 (2022).
Dang, K. et al. Distinct planting patterns exert legacy effects on the networks and assembly of root-associated microbiomes in subsequent crops. Sci. Total Environ. 946, 174276 (2024).
Ma, W. Y., Liao, X. L., Wang, C. & Zhang, Y. Effects of four cropping patterns of lilium Brownii on rhizosphere Microbiome structure and replant disease. Plants (Basel). 11 (6), 824 (2022).
Yan, H. L. et al. Grass-legume mixed cropping enhance forage production via the bacterial community. Agric. Ecosyst. Environ. 338 (6), 108087 (2022).
Zhao, W., Yin, Y. L., Song, J. Q. & Li, S. X. Mixed sowing improves plant and soil bacterial community restoration in the degraded alpine meadow. Plant. Soil. 499, 379–392 (2024).
Zhang, Y. et al. Phytoremediation performance of mixed planting patterns and the associated rhizosphere microbial community in pilot-scale constructed wetlands. Chemosphere 361, 142482 (2024).
Ali, A. et al. Different cropping systems regulate the metabolic capabilities and potential ecological functions altered by soil Microbiome structure in the plastic shed mono-cropped cucumber rhizosphere. Agric. Ecosyst. Environ. 318, 107486 (2021).
Wang, X. et al. Effects of different artificial vegetation restoration modes on soil microbial community structuree in the soil erosion area of Southern China. CATENA 237, 107803 (2024).
Huang, A. C. et al. A specialized metabolic network selectively modulates Arabidopsis root microbiota. Science 364 (6440), eaau6389 (2019).
Duan, Y. et al. The effects of tea plants-soybean intercropping on the secondary metabolites of tea plants by metabolomics analysis. BMC Plant Biol. 21 (1), 482 (2021).
Ma, D. et al. Accumulation characteristics of plant flavonoids and effects of cultivation measures on their biosynthesis: A review. Plant Physiol. Biochem. 215, 108960 (2024).
Che, T. et al. Mixed planting reduces the shaping ability of legume cover crop on soil microbial community structure. Appl. Soil. Ecol. 178, 104581 (2022).
Lu, S. et al. Co-analysis of rhizosphere metabolomics and bacterial community structures to unfold soil ecosystem health in camellia Oleifera land under long-term cultivation. Appl. Soil. Ecol. 171 (2), 104336 (2022).
Brown, R. W. et al. Soil metabolomics-current challenges and future perspectives. Soil Biol. Biochem. 193, 109382 (2024).
Bi, B. Y. et al. Rhizosphere soil metabolites mediated microbial community changes of Pinus sylvestris Var. Mongolica across stand ages in the mu Us desert. Appl. Soil. Ecol. 169, 104222 (2022).
Su, Y. et al. Dynamic metabolites: A Bridge between plants and microbes. Sci. Total Environ. 899, 165612 (2023).
Jia, J. et al. Characteristics of microbial communities and antibiotic resistance genes in typical rivers of the Western Qinghai lake basin. Water Biology Secur. 3 (2), 100249 (2024).
Li, J. C., Wang, Y., Zhang, L. S., Han, L. Y. & Hu, G. Y. Aeolian desertification in China’s Northeastern Tibetan plateau: Understanding the present through the past. CATENA 172, 764–769 (2019).
Chen, X. et al. Reasonable management of perennial planting grassland contributes to positive succession of soil microbial community in Sanjiangyuan of Qinghai-Tibetan plateau, China. J. Plant. Ecol. 15 (2), 359–371 (2021).
Maiti, P. et al. Landscape level ecological assessment and eco-restoration strategies for alpine and sub-alpine regions of the central himalaya. Ecol. Engineering: J. Ecotechnology. 180, 106674 (2022).
Li, S. D. et al. The impact of mixed planting of Poaceae species in the Qinghai-Tibet plateau region on forage yield, soil nutrients, and soil microbial communities. Front. Plant Sci. 15 https://doi.org/10.3389/fpls.2024.1370593 (2024).
Jiang, X. & Niu, K. C. Effects of grass mixed-sowing on soil microbial diversity on the Qingzang (Tibetan) plateau. Chin. J. Plant. Ecol. 45 (5), 539–551 (2021).
Guan, Q. Q. et al. Analyzing the Spatial heterogeneity of number of plant individuals in grassland community by using power law model. Ecol. Model. 320, 316–321 (2016).
Zhang, J. X. et al. Responses of soil microbial communities of sown perennial grassland in apline region to different sowing ways and growth years. ACTA AGRESTIA SINICA. 29 (2), 270–280 (2021).
Dong, R. Z. et al. Differences in soil microbial communities with successional stage depend on vegetation coverage and soil substrates in alpine desert shrublands. Plant. Soil. 485, 549–568 (2023).
Tong, Y. S. et al. Nitrogen application increases the productivity of perennial alpine cultivated grassland by improving soil physicochemical properties and microbial community characteristics. Plant. Soil. 505, 559–579 (2024).
Xie, L. L., Ma, Y. S., Wang, Y. L., Ma, Y. & Wang, X. L. Changes in soil bacterial and fungal community composition and functional groups during the artificial restoration of degraded grassland of Black-Soil mountain. Ecol. Evol. 14 (10), e70361 (2024).
Huang, X. M. et al. Changes of soil microbial biomass carbon and community composition through mixing nitrogen-fixing species with Eucalyptus urophylla in subtropical China. Soil Biol. Biochem. 73, 42–48 (2014).
Yin, Y. L. et al. Soil microbial character response to plant community variation after grazing prohibition for 10 years in a Qinghai-Tibetan alpine meadow. Plant. Soil. 458, 175–189 (2021).
Logue, J. B. et al. Experimental insights into the importance of aquatic bacterial community composition to the degradation of dissolved organic matter. ISME J. 10 (3), 533–545 (2016).
Walters, W. et al. Improved bacterial 16S rRNA gene (V4 and V4-5) and fungal internal transcribed spacer marker gene primers for microbial community surveys. mSystems 1 (1), e00009–15 (2016).
Bai, X., Jia, X. U., Zhao, C. L. & Shao, M. A. Artificial forest conversion into grassland alleviates deep-soil desiccation in typical grass zone on China’s loess plateau: regional modeling. Agric. Ecosyst. Environ. 320, 107608 (2021).
Liu, J. R. et al. Mixed planting mode is the best measure to restore soil quality in alpine mines. Soil Tillage. Res. 244, 106209 (2024).
Zhang, M. M. et al. Responses of soil microbial community structure, potential ecological functions, and soil physicochemical properties to different cultivation patterns in Cucumber. Geoderma. 429 (11), 116237 (2022).
Wen, B. J. et al. Response relationship between vegetation structure and runoff-sediment yield in the hilly and gully area of the Loess Plateau, China. CATENA. 227, 107107 (2023).
Hector, A. et al. General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding. Ecology 91 (8), 2213–2220 (2010).
Jing, X. et al. Soil microbial carbon and nutrient constraints are driven more by climate and soil physicochemical properties than by nutrient addition in forest ecosystems. Soil Biol. Biochem. 141, 107657 (2020).
Hong, Y. C., Zhou, Q., Hao, Y. Q. & Huang, A. C. Crafting plant root metabolome for improved microbe-assisted stress resilience. New Phytol. 234 (6), 1945–1950 (2021).
Nayyar, A. et al. Soil microbial quality associated with yield reduction in continuous-pea. Appl. Soil. Ecol. 43 (1), 115–121 (2009).
Lu, S. et al. Long-term rice-rice-rape rotation optimizes 1,2-benzenediol concentration in rhizosphere soil and improves nitrogen-use efficiency and rice growth. Plant. Soil. 445, 23–37 (2019).
Kaya, C., Ashraf, M., Alyemeni, M. N., Rinklebe, J. & Ahmad, P. Trehalose and NO work together to alleviate cd toxicity in pepper (Capsicum annuum L.) plants by regulating cadmium sequestration and distribution within cells and the antioxidant defense system. Sci. Hort. 314 (1), 111948 (2023).
Smirnova, I., Sadanov, A., Baimakhanova, G., Faizulina, E. & Tatarkina, L. Metabolic interaction at the level of extracellular amino acids between plant growth-promoting rhizobacteria and plants of alfalfa (medicago sativa l). Rhizosphere 21 (3), 100477 (2022).
Li, W. G. et al. Untargeted metabolomics to study changes in soil microbial community in response to tillage practices. Appl. Soil. Ecol. 199, 105409 (2024).
Feng, K. et al. Biodiversity and species competition regulate the resilience of microbial biofilm community. Mol. Ecol. 26 (21), 6170–6182 (2017).
Wu, B. H. et al. Miscanthus Sp. root exudate alters rhizosphere microbial community to drive soil aggregation for heavy metal immobilization. Sci. Total Environ. 949, 175009 (2024).
Fisk, L. M., Barton, L., Jones, D. L., Glanville, H. C. & Murphy, D. V. Root exudate carbon mitigates nitrogen loss in a semi-arid soil. Soil Biol. Biochem. 88, 380–389 (2015).
Jin, J. et al. Integrated microbiology and metabolomics analysis reveal patterns and mechanisms of improvement the achnatherum inebrians adaptability to N addition by endophytic fungus Epichloë gansuensis. Environ. Exp. Bot. 213, 105421 (2023).
Lin, W. H. Study on the Ecological Characteristics of Epichloë Endophyte-Festuca Sinensis Associations (DOCTOR OF PHILOSOPHY, 2020).
Ma, Y. Study on Rhizosphere Nutrient Regulation Mechanism of Dominant Plants in Degraded Alpine Meadow on the Eastern Edge of Qilian Mountains (DOCTOR OF PHILOSOPHY, 2020).
Peng, M. W. et al. Morphological, physiological and metabolomic analysis to unravel the adaptive relationship between root growth of ephemeral plants and different soil habitats. Plant Physiol. Biochem. 202 (1), 107986 (2023).
Chen, P. et al. Converting leguminous green manure into Biochar: changes in chemical composition and C and N mineralization. Geoderma 232–234, 581–588 (2014).
Chen, C., Chen, H. Y., Chen, X. L. & Huang, Z. Q. Meta-analysis shows positive effects of plant diversity on microbial biomass and respiration. Nat. Commun. 10, 1332 (2019).
Zhang, G. Y., Shen, Z. X. & Fu, G. Function diversity of soil fungal community has little exclusive effects on the response of aboveground plant production to experimental warming in alpine grasslands. Appl. Soil. Ecol. 168, 104153 (2021).
Pot, V. et al. Understanding the joint impacts of soil architecture and microbial dynamics on soil functions: insights derived from microscale models. Eur. J. Soil. Sci. 73 (3), e13256 https://doi.org/10.1111/ejss.13256 (2022).
Wang, W. Y. et al. Root exudates enhanced rhizobacteria complexity and microbial carbon metabolism of toxic plants. iScience 25 (10), 105243 (2022).
Bai, Y. X., Wei, H., Ming, A. G., Shu, W. W. & Shen, W. J. Tree species mixing begets admixture of soil microbial communities: variations along bulk soil, rhizosphere soil and root tissue. Geoderma 438 (2), 116638 (2023).
Hallam, S. J. & McCutcheon, J. P. Microbes don’t play solitaire: how Cooperation Trumps isolation in the microbial world. Environ. Microbiol. Rep. 7 (1), 26–28 (2015).
Stefanowicz, A. M. et al. Herbaceous plant species and their combinations positively affect soil microorganisms and processes and modify soil physicochemical properties in a mesocosm experiment. For. Ecol. Manag. 532 (10), 120826 (2023).
Gong, X. W. et al. Linkages of microbial community structure and root exudates: evidence from microbial nitrogen limitation in soils of crop families. Sci. Total Environ. 881, 163536 (2023).
Ridgeway, J., Morrissey, E. M. & Brzostek, E. R. Plant litter traits control microbial decomposition and drive soil carbon stabilization. Soil Biol. Biochem. 175 (1), 108857 (2022).
Liu, K. H., Ding, X. W. & Wang, J. J. Soil metabolome correlates with bacterial diversity and co-occurrence patterns in root-associated soils on the Tibetan plateau. Sci. Total Environ. 735, 139572 (2020).
Bahram, M. et al. Structure and function of the global topsoil Microbiome. Nature 560, 233–237 (2018).
Zhao, M. L. et al. Root exudates drive soil-microbe-nutrient feedbacks in response to plant growth. Plant. Cell. Environ. 44 (2), 613–628 (2020).
Gao, L. et al. Effect of altitude on the release of metabolites in the tobacco rhizosphere soil. Eurasian Soil. Sci. 55, 1786–1793 (2022).
Yuan, A. et al. Dynamic interplay among soil nutrients, rhizosphere metabolites, and microbes shape drought and heat stress responses in summer maize. Soil Biol. Biochem. 191, 109357 (2024).
Lopes, L. D. & Schachtman, D. P. Rhizosphere and bulk soil bacterial community succession is influenced more by changes in soil properties than in rhizosphere metabolites across a maize growing season. Appl. Soil. Ecol. 189 (4), 104960 (2023).
Liu, J. P. et al. Integrated Microbiome and metabolomics analysis reveal a closer relationship between soil metabolites and bacterial community than fungal community in Pecan plantations. Land. Degrad. Dev. 34 (10), 2812–2824 (2023).
Funding
This study was financially supported by the National Key R&D Program of China (2022YFD1302104).
Author information
Authors and Affiliations
Contributions
QuanMin Dong, ChunPing Zhang conceived the ideas and designed the study; XiaoFang Zhang, Lian Huo, Xue Zhang, Zehang Yu, Xiaoxia Yang, Quan Cao, Zengzeng Yang collected the data; XiaoFang Zhang, Yongshang Tong, Yuzhen Liu analyzed the data; XiaoFang Zhang, ChunPing Zhang, Yuzhen Liu wrote and revised the draft; All authors contributed to the article and approved the submitted version.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, X., Zhang, C., Liu, Y. et al. Effects of planting patterns on physicochemical properties, metabolites and microbial community structure of rhizosphere soil in perennial cultivated grassland. Sci Rep 15, 12047 (2025). https://doi.org/10.1038/s41598-025-94366-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-94366-7
