
Abstract
Susceptibility of bats to white-nose syndrome (WNS), a lethal disease caused by the fungus Pseudogymnoascus destructans (Pd), may be influenced by the amount of activity outside hibernacula during the winter. We tested the effects of hibernaculum type (aboveground or subterranean) and Pd status (positive or negative) on winter activity of tricolored bats (Perimyotis subflavus) in the southeastern USA along with the effects of ambient temperature, precipitation, and stage of hibernation. We placed acoustic detectors at the entrances of 13 hibernacula (4 aboveground and Pd-positive, 4 aboveground and Pd-negative, 4 subterranean and Pd-positive, and 1 subterranean and Pd-negative) during winter 2020–21 and 2021–22. While neither hibernaculum type nor Pd status alone predicted probability of activity or levels of activity, these factors interacted with temperature, precipitation, and stage of the hibernation period. Activity increased at a greater rate with temperature and time since the onset of hibernation in aboveground and Pd-negative sites and decreased at a faster rate in response to precipitation. Our results suggest that tricolored bats using aboveground hibernacula such as culverts or bridges may be less susceptible to WNS due to greater nighttime activity. However, use of these structures may have other costs such as higher freezing and predation risks.
Similar content being viewed by others

Introduction
Many species use extended torpor or hibernation during winter as a means to manage energetic demands resulting from cold temperatures and reduced food availability1. During these periods, animals allow their body temperatures to drop towards ambient, thus decreasing their metabolic costs. Bouts of torpor are interrupted by episodic arousals that serve many purposes including ridding the body of accumulated wastes, reactivating the immune system, and alleviating dehydration and sleep deprivation2,3. Among bats, these arousal periods are also sometimes characterized by activity outside the hibernaculum to switch roosts, drink, and occasionally forage4,5,6,7. Little is known about the behavior of bats outside hibernacula during winter, but this information is essential for effective management of winter habitats as well as understanding bats’ responses to stressors such as climate change and disease7,8. For example, increased activity during winter due to warming temperatures may represent an ecological trap if resources such as insects or water are not available9.
White-nose syndrome (WNS) is a fungal disease that affects bats during winter hibernation10. The fungus invades bats’ epidermis, resulting in a series of physiological responses, including increased arousals from torpor, increased blood CO2levels, water and electrolyte loss, and hyperventilation11. The combined effect of these responses is an increase in energy use and consequently, the depletion of bats’ fat reserves and in many cases, death. The disease was first documented in upstate New York, USA in 200612and was most likely introduced from Europe13,14. Since its introduction, the fungus that causes WNS (Pseudogymnoascus destructans; hereafter referred to as Pd) has spread widely throughout much of the Midwestern, northeastern, and southern regions of the U.S., as well as parts of the western U.S. and up through much of southern Canada (https://whitenosesyndrome.org/where-is-wns). Mortality in some species has resulted in > 90% declines in populations within the affected area15.
Most of the mortalities from WNS have occurred within four species: northern long-eared bats (Myotis septentrionalis), little brown bats (M. lucifugus), Indiana bats (M. sodalis), and tricolored bats (Perimyotis subflavus)15. Pd has been found on many other species that do not show diagnostic symptoms of the disease16, whereas other species that are infected appear to be far less susceptible to its effects. For example, big brown bats (Eptesicus fuscus) show diagnostic signs of the disease but have not exhibited significant population declines15,17. Decreased susceptibility of big brown bats to WNS compared to species such as little brown bats may be due to thermoregulatory responses18, larger body size and fat reserves19, differences in skin fatty acid composition and higher concentrations of fatty acids with anti-fungal properties20,21,22, and higher wing acidity23. Rafinesque’s big-eared bats (Corynorhinus rafinesquii) and southeastern myotis (M. austroriparius) also do not appear to be susceptible to WNS, perhaps due to their shallow torpor patterns and frequent arousals from winter torpor resulting in higher immune function24,25. Because Pd does not grow when temperatures are > 19.0 °C26, bats that exhibit shallow torpor and frequent nighttime activity may be less susceptible to WNS than those that go into prolonged periods of deep torpor because their body temperatures rise above the growth threshold for Pd and their immune systems may be more active27,28,29,30. Thus, knowledge of the torpor and activity patterns of bats throughout winter may help predict the response of bats to Pd once the fungus has been transferred to an area.
Tricolored bats are found throughout the eastern USA and are one of the species most severely impacted by WNS15. Across much of their range tricolored bats hibernate in subterranean structures such as caves, mines, and tunnels31. However, in areas where these structures are not available, such as much of the southern U.S., they roost in culverts, bridges, and trees during winter32,33,34,35,36. Tricolored bats that use these aboveground structures can go into deep torpor with torpor bouts lasting up to 15.5 days but they may also leave the roost at night to forage or switch roosts28. Arousals from torpor in aboveground hibernacula usually occur around dusk. In contrast, tricolored bats that hibernate in caves and tunnels arouse randomly throughout day and night although there is little evidence that tricolored bats in belowground leave the hibernacula37.
While Pd and WNS are prevalent in much of the southeastern U.S., the fungus has not been detected in many areas, particularly along the Atlantic and Gulf Coasts and in Florida (https://whitenosesyndrome.org/where-is-wns). To better understand the potential for morbidity and mortality if the fungus arrives in these areas, we studied activity outside the roost in tricolored bats roosting in subterranean and aboveground hibernacula in four southeastern U.S. states (Fig. 1). Because greater activity, particularly on warmer nights when insects may be active, may decrease bats’ susceptibility to WNS, and bats in aboveground hibernacula may be more active those in subterranean hibernacula, our objective was to test the effects of hibernaculum type (aboveground or subterranean) on activity of tricolored bats outside hibernacula during winter. Although WNS causes bats to arouse more frequently during hibernation and often leave the hibernacula5,38, arousals return to pre-WNS frequency during the endemic phase of the disease39. Thus, activity outside hibernacula in the endemic phase of the disease likely represents behaviors such as foraging, drinking, or movement among hibernacula. We also examined the effects of Pd status (positive or negative), environmental conditions (ambient temperature and precipitation) and stage of the hibernation period. We hypothesized that nighttime activity would vary with hibernaculum type, Pd status, and environmental conditions. We predicted that activity outside the hibernaculum would be greater in aboveground hibernacula because bats would be more responsive to outside environmental conditions and that bats in Pd-negative sites would be more active than those in Pd-positive sites because bats infected with Pd need to conserve energy40. We also predicted that activity would be positively associated with ambient temperature and negatively associated with nighttime precipitation and be highest during the early and late parts of the hibernation season.

Location of tricolored bat (Perimyotis subflavus) hibernacula monitored for winter activity across the southeastern USA.
Methods
Study sites
We monitored nighttime activity at 13 sites across Alabama, Florida, Georgia, and South Carolina, USA over two winters although one site (Bullock Creek Culvert, Alabama), was only monitored during 2021–22 (Fig. 1). Eight of the sites were aboveground structures (six culverts and two bridges) and five were subterranean (two caves, two incomplete railroad tunnels, and one abandoned gold mine; Table 1). Prior to and during the study, all sites were monitored for the presence of Pd and WNS using standard protocols as outlined by the USGS National Wildlife Health Center (NWHC; https://d9-wret.s3.us-west-2.amazonaws.com/assets/palladium/production/s3fs-public/media/files/NWHC%20Winter%202023-2024%20Bat%20Submission%20Guide_v01022024.pdf). Samples taken prior to our study were analyzed by various members of the White-Nose Syndrome Diagnostic Laboratory Network, and samples taken during this study were processed by the laboratory at Kennesaw State University, one of the members of the Network. Following the guidelines of the NWHC, we considered a site to be Pd-positive if at least some samples had a Ct ≤ 37. Pd was documented in the four Pd-positive subterranean sites and three of the four aboveground Pd-positive sites prior to the onset of the study (Table 1). One site was first tested in the first year of our study and was Pd-positive. However, none of the bats in the aboveground Pd-positive sites showed signs of WNS. One of the sites, Black Diamond Tunnel in northwestern Georgia received an anti-fungal treatment from 2016 to 202341. However, a high proportion of bats tested in this hibernaculum still had Ct < 37 during this study42. Due to the progression of the disease from its origin in New York (https://whitenosesyndrome.org/where-is-wns), Pd-positive sites were in the northern and higher elevation portions of the study area, whereas Pd-negative sites were in the southern and lower elevation portions (Fig. 1). The number of tricolored bats inhabiting the sites ranged from 3 to 654 (Table 1).
Acoustic sampling and analysis
Because tricolored bats in the northern hibernacula in our study area enter hibernation in late September or early October and emerge in early to mid-March43, our goal was to sample October through March. However, sampling started later in 2020–21 due to delays related to the SARS-COV2 pandemic and we were only able to monitor bat activity at each site from late November or early December 2020 through mid- to late March 2021. In the second year we were able to monitor activity from early October 2021 through mid- to late March 2022. We placed Anabat Express bat detectors (Titley Scientific, Columbia, MO) at the entrance of each hibernaculum; when there were two entrances such as in the case of culverts, bridges, and one cave, we placed a detector at each entrance. Detectors were placed on trees or poles approximately 2–3 m above ground and approximately 5–20 m from the entrance depending on the hibernaculum. Although some activity can occur during the day in WNS-affected bats5, most studies have shown that activity of bats during winter occurs primarily within the early part of the night44,45,46. Thus, we programmed detectors to run from 1800 to 2400 h each night.
We used Kaleidoscope Pro version 5.4.3 (Wildlife Acoustics, Maynard, MA) to identify call files to species. We modified the potential species lists for each site so that only species that were likely to be in the area based on range maps were included for each site. To minimize misclassification, we used the Conservative setting and required at least five pulses be present. We did not manually vet all tricolored bat call files but spot-checked files from each site to verify correct identifications, particularly on nights when high numbers of tricolored bat calls were recorded. In most cases we deemed that the identifications were correct. In those few cases where we did not agree with the auto-identifications, we did not include those files in the analyses. For those sites where two detectors were placed, we averaged the nightly call files from the two detectors.
Weather data
We obtained minimum, maximum, and mean daily temperatures and hourly precipitation data from the nearest weather station to each site. Data were obtained either from Remote Automated Weather Stations (RAWS; https://raws.dri.edu/) or through Mesowest (https://mesowest.utah.edu/) when RAWS stations were farther away. We averaged hourly temperatures to obtain daily mean temperature and totaled the rainfall for 1700–2400 h (i.e., the recording period plus the hour prior).
Statistical analysis
We used R version 4.4.0 for all analyses. We tested for collinearity among our independent variables and found that mean, maximum, and minimum temperatures were highly correlated (r ≥ 0.90). Therefore, we only used mean daily temperature in our analyses.
We used linear mixed models (Package lme4)47 with site as the random effect to test for differences in temperature and precipitation between years using the full dataset (October–March) and December–March data only due to the shortened field season in 2020–21. We also used a chi-square analysis (Package gmodels; https://cran.r-project.org/web/packages/gmodels/index.html) to test whether the proportion of nights during which rainfall occurred differed between years.
Because data were collected late-November through March in 2020–21and for the entire winter season (October through March) in 2021–22, we analyzed the two years of acoustic data separately. To account for repeated measures (i.e., multiple nights) at each site as well as Type and Status of each site, we used Site within Type and Status as a random effect in the models which adjusted for the repeated observations within each site. We used generalized mixed models (Package lme4) to test the effects of hibernaculum type (Type), Pd status (Status), ambient temperature (Temperature), presence or amount of precipitation (Precipitation), and stage within the hibernation period (Date) on tricolored bat activity. We considered including latitude and elevation as independent variables, but they were synonymous with site. We first tested whether the presence of activity during the night (0 or 1) was affected by these factors using a binomial distribution and then we tested whether the level of activity (number of call files per night) was affected by these factors using a Poisson distribution. Because we averaged the number of calls at sites with two entrances, we used the round function in R to meet the assumptions of a Poisson distribution for those sites where the number of call files per night was averaged. Further, the number of bats in a structure likely influenced the level of activity at a site (i.e., the number of call files was likely related to the number of bats in a site). Therefore, we used the number of bats in each hibernaculum as an offset in our activity models. Tricolored bats enter hibernation in late September at our northern sites43, so we used days from October 1 (Date) as our measure of hibernation stage and also included its quadratic term (Date2) to account for the differences between the onset and end of hibernation and its middle. Because our goal was to examine the effects of hibernaculum type and Pd status on tricolored bat activity as well as the interactions of temperature, period of the hibernation period, and precipitation with hibernaculum type and Pd status, we built one model for each year and response type (presence or level of activity) to test these effects. Unfortunately, some interactions were not estimable and could not be included in some models. When this occurred, we included those interactions that allowed us to look at the most interactions. Further, using the amount of precipitation per night resulted in non-convergence of our binomial models so we used the presence of rain during the night as our Precipitation variable in those models. We scaled all continuous independent variables to a mean of 0 and a standard deviation of 1 before analysis and used α ≤ 0.05 to denote significant effects.
Results
We monitored bat activity over 3,742 detector nights: 539 at subterranean hibernacula and 1,005 at aboveground hibernacula in 2020–21 and 718 at subterranean hibernacula and 1,480 at aboveground hibernacula in 2021–22. We recorded 204,145 files containing bat calls of which 27,007 were identified as tricolored bat calls (946 at subterranean sites and 6,066 at aboveground sites in 2020–21, and 3,631 at subterranean sites and 16,364 at aboveground sites in 2021–22). Other species identified were big brown bats, eastern red bats/Seminole bats (Lasiurus borealis or L. seminolus), hoary bats (L. cinereus), silver-haired bats (Lasionycteris noctivagans), southeastern myotis, gray bats (M. grisescens), eastern small-footed bats (M. leibii), little brown bats, northern long-eared bats, evening bats (Nycticeius humeralis), and Brazilian free-tailed bats (Tadarida brasiliensis). Tricolored bats were active during some nights at all hibernacula with the greatest level of activity (i.e., mean number of files/night) occurring at one of the bridges (Table 1). The proportion of nights that bats were active, as well as the level of activity, varied considerably among sites and between years at some hibernacula but not at others (Table 1).
The number of nights during which rain occurred did not vary significantly between years when data for all months (October through March) were considered (χ2 = 2.06, d.f. = 1, P = 0.15) or just December through March (χ2 = 0.04, d.f. = 1, P = 0.84). Further, the amount of rain that fell did not differ between years when all months were considered (F = 1.15, d.f. = 1, 2555, P = 0.28) or when only December through March were considered (F = 2.61, d.f. = 1, 1864, P = 0.11). In contrast, temperatures were significantly warmer in 2021–22 than in 2020–21 when all months were considered and for December–March only (F = 159.59, d.f. = 1, 2800, P < 0.0001 and F = 38.15, d.f. = 1, 2133, P < 0.0001, respectively). Mean daily temperatures were 9.6 ± 0.58 °C in December–March 2020–2021. In 2021–22, mean daily temperatures were 12.2 ± 0.50 °C from October through March and 11.0 ± 0.58 °C from December through March.
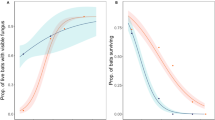
Probability of activity did not vary with Type in either year but varied with Status in 2020–21 (Table 2); the interaction between Type and Status was not significant in either year. The probability of activity was significantly greater in Pd-negative sites (0.65 ± 0.12) than Pd-positive sites (0.07 ± 0.03) in 2020–21 (Fig. 2 A). In 2021–22, probability of activity was 0.68 ± 0.13 in Pd-negative sites and 0.42 ± 0.11 in Pd-positive sites (Fig. 2B). Temperature, Precipitation, and Date were significant factors in both years (Table 2). Probability of activity was positively related to temperature and days since the onset of hibernation, whereas the relationship between probability of activity and rain during the night was negative (Table 2). However, ambient temperature and days since the onset of hibernation sometimes interacted with Type and Status making interpretation more complex. In 2020–21, the interaction between Temperature and Status was significant, and the interactions between the quadratic term of Date and Type and quadratic term of Date and Status were significant (Table 2). The probability of activity increased with ambient temperature in both Pd-negative and Pd-positive sites but increased at a greater rate in Pd-negative sites than Pd-positive sites (Fig. 3 A). The probability of activity increased linearly with Date in subterranean sites and in Pd-negative sites but was greater at the beginning and end of the hibernation period in aboveground sites and Pd-positive sites (Fig. 4 A and B). In 2021–22, the interaction between Status and Date was significant (Table 2). The probability of activity increased at a greater rate in Pd-negative sites as time since the onset of hibernation progressed (Fig. 4D). In 2021–22 the effects of Precipitation did not vary with either Type or Status (Table 2); these interactions could not be tested for 2020–21. Probability of activity was lower during nights with rain in both years (0.09 ± 0.03 in 2020–21 and 0.35 ± 0.09 in 2021–22) versus no rain (0.19 ± 0.05 in 2020–2021 and 0.57 ± 0.09) in both hibernaculum types and Pd-positive and Pd-negative sites.

Mean (± 1 S.E.) probability of tricolored bat (Perimyotis subflavus) activity (A and B) and mean (± 1 S.E.) number of calls per night (C and D) in aboveground and subterranean hibernacula in the southeastern USA during winter 2020–21 and 2021–22.

Mean number of calls per night in aboveground and subterranean hibernacula in the southeastern USA in relation to ambient temperature and hibernacula type (A and C) and Pd status (B and D) during winter 2020–21 and 2021–22. Shaded areas represent 95% confidence limits.

Probability of tricolored bat (Perimyotis subflavus) activity (A-D) and mean number of calls per night (E-H) in aboveground and subterranean hibernacula in the southeastern USA in relation to stage of hibernation (days since October 1) and hibernacula type and Pd status during winter 2020–21 and 2021–22. Shaded areas represent 95% confidence limits.

Mean number of calls per night in relation to the amount of precipitation during the night in aboveground and subterranean hibernacula in the southeastern USA during winter 2020–21 and 2021–22. Shaded areas represent 95% confidence limits.
Neither Type nor Status were significant predictors of activity level in either year (Table 3). However, the interaction between Type and Status was significant in 2020–21 (Table 3). The difference between activity levels in Pd-negative and Pd-positive sites was greater in aboveground hibernacula (14.2 ± 11.8 calls/night versus 0.08 ± 0.06 calls/night) than in subterranean hibernacula (2.3 ± 3.3 versus 0.5 ± 0.4; Fig. 2 C). Activity levels were positively associated with Temperature and Date in both years and negatively associated with Precipitation (Table 3). However, most interactions between these factors and Type and Status were significant. Activity increased at a greater rate with Temperature and Date in Pd-negative sites than Pd-positive sites in 2021–22 and 2020–21 respectively (Figs. 3 C and D and 4 F H) and decreased more rapidly with Precipitation in 2021–22 (Fig. 5D); this relationship was not significant in 2020–21 but followed a similar pattern (Fig. 5B). In both years, activity increased more rapidly with Temperature (Fig. 3E and G) and Date (Fig. 4E and G) in aboveground hibernacula than subterranean hibernacula. In contrast, activity decreased more rapidly in both years with Precipitation in aboveground sites than subterranean sites (Fig. 5 A and C).
Discussion
Tricolored bats were active throughout the winter across our 13 sites, similar to other studies conducted on winter bat activity in the south27,48,49. However, the probability of tricolored bat activity and the level of activity varied considerably across sites. We found environmental factors and stage of the hibernation season were more important predictors of this variation than hibernaculum type or Pd status, likely due to bats’ having evolved their torpor and activity patterns to environmental conditions. However, bats in aboveground and subterranean structures and in structures that were either Pd-positive or Pd-negative, responded to these environmental factors in somewhat different ways. This suggests that even though hibernaculum type and Pd status were not significant factors alone, these factors may contribute to tricolored bats’ winter activity outside hibernacula and thus, their susceptibility to the effects of WNS. For example, although tricolored bats in Stumphouse and Black Diamond Tunnels, both subterranean Pd-positive sites, experienced high mortality after the arrival of Pd41,50, no evidence of mortality and only one incidence of diagnostic signs of WNS have been found in Pd-positive culverts (Katrina Morris, Georgia Department of Natural Resources, pers. comm.). While we were not able to include latitude or elevation in our models, these factors may have also contributed to our results due to their influence on ambient temperature.
As we predicted, ambient temperature was a strong predictor of both the probability of bat activity on a given night as well as the level of activity (i.e., number of call files), and both the probability of activity and the level of activity increased with ambient temperature. Almost all other studies examining bat winter activity have found a similar relationship between bat activity and temperature5,29,44,51. Although there were a few instances of tricolored bat activity occurring when average daily temperatures were < 0 °C, the probability of activity began to increase between approximately 5 °C and 10 °C; the level of activity began to increase when average daily temperatures ranged between 10 °C and 20 °C (Fig. 3). In managed pine stands of east Texas and Louisiana in the Southern Coastal Plains, tricolored bats were not detected at temperatures < 4.5 °C and had a mean threshold temperature for activity of 13.7 °C52. This threshold was relatively high compared to other species in the area, suggesting that tricolored bats are more likely to stay in torpor than other species, even on warmer nights. Tricolored bats are more active away from their roosts when nighttime temperatures rise above 10 °C28, which may be related to increased availability of insects53, decreased energetic costs of flight at warmer temperatures54, or both. Further, bats roosting in bridges and tree foliage are more likely to be active than bats roosting in more thermally stable tree cavities28. The highest activity that we observed in the current study was for bats in one of these bridges.
Although activity increased with temperature in both above- and belowground sites and at Pd-positive and Pd-negative sites, there were some differences in the rates of those increases. For example, probability of activity increased far more rapidly with temperature in Pd-negative sites than Pd-positive sites in 2020–21 (Fig. 3 A), and the level of activity increased at a greater rate in Pd-negative sites in that year (Fig. 3C). Similarly, the level of activity increased more rapidly with temperature in aboveground hibernacula than subterranean ones in both years. Tricolored bats in aboveground hibernacula are more likely to arouse near dusk28, whereas tricolored bats in subterranean sites arouse randomly throughout the day37. Thus, tricolored bats in aboveground sites may be better able to assess nighttime ambient conditions and respond more quickly when conditions are favorable for outside activity. Bats in Pd-positive sites may need to conserve more energy than those in Pd-negative sites due to the effects of the disease40, and therefore, may be less likely to leave the roost unless conditions are highly favorable55.
Tricolored bat activity was negatively associated with the presence and amount of precipitation during the night. Further, the decrease in activity with rainfall was steeper in aboveground hibernacula and Pd-negative sites. Few studies have examined the effects of precipitation on bat activity; however, in Portugal winter bat activity was negatively associated with precipitation56. Torpor bout lengths of tricolored bats using aboveground roosts in winter are positively associated with precipitation in South Carolina28, and the probability of summer torpor in little brown bats increases with the presence of rain57. When rain results in wet pelage and subsequent increased evaporative cooling, energetic costs increase significantly possibly due to increased metabolic rates58. Wet fur may also affect flight dynamics and rain may affect sensory capabilities such as echolocation. All these factors may contribute to the negative association of outside bat activity with rain. The more rapid shutdown of activity by tricolored bats in aboveground hibernacula in response to rain may be due to their increased ability to monitor environmental conditions such as the presence of rain due to the more exposed nature of the roost59. In contrast, bats in subterranean hibernacula may need to exit the site to assess conditions.
We predicted that activity to be greatest at the beginning and end of the hibernation season, as observed in other bat communities29,60,61. However, this response only occurred for the probability of activity in 2020–21 and only in aboveground structures and at Pd-positive sites (Fig. 4). Instead, activity tended to increase as the hibernation period progressed. The presence of a curvilinear response in 2020–21 and not in 2021–22 is particularly curious since we did not start monitoring activity until late November or early December in 2020–21, after the period that we predicted that activity would be high. Increased activity at the end of the season is likely related to bats starting to emerge from hibernation43. Low activity at the beginning of the season may reflect bats entering the hibernacula to begin hibernation for the winter. For example, two tricolored bats that were radio-tagged outside Stumphouse Tunnel (one of the sites in this study) in late September-early October roosted in trees for a few days and then entered the tunnel and did not leave for over a month62. One of these bats was observed in the tunnel throughout the winter. Outside activity of tricolored bats was also minimal between mid-October and February or March at two caves in Indiana63,64. We observed that tricolored bat activity increased at a greater rate at the end of the season in aboveground hibernacula and Pd-negative sites. Tricolored bats in aboveground hibernacula may be more responsive to ambient conditions and may leave the hibernacula earlier to move to summer habitats than those in subterranean structures although, we are unaware of any studies that have examined this. Little brown bats with WNS choose roosts that allow them to use daily torpor after emergence from hibernacula to conserve energy65. Similarly, tricolored bats in Pd-positive sites in our study may have stayed in hibernacula longer to conserve energy, resulting in lower activity at the end of hibernation season compared to bats in Pd-negative sites.
Winter temperatures are projected to increase over the next several decades in the southeast66, which suggests that activity of tricolored bats outside hibernacula may also increase, particularly in aboveground hibernacula. However, more frequent arousals and activity outside hibernacula may also lead to increased predation, increased risk of fat depletion and dehydration, and phenological mismatches such as lack of insect availability during activity periods, particularly for bats that hibernate in subterranean hibernacula and more northern latitudes9. For example, tricolored bats that hibernate in caves in Florida, which has a subtropical climate, have declined by 70% in the absence of WNS67. The decline is closely related to increasing temperatures and may be due to increased energy use related to hibernating at warmer temperatures. Therefore, even though outside activity may be higher in warmer winters, potentially helping to mitigate the effects of WNS, increasingly warm winters may have long-term effects on tricolored bat populations.
Once Pd and WNS were introduced to North America in New York it spread north, south, and west12with the spread south occurring primarily down the Appalachian Mountains68. Once Pd arrives in an area it usually becomes widespread (https://whitenosesyndrome.org/where-is-wns). Hence, it was often not possible to find Pd-negative sites near Pd-positive sites. Consequently, three of our five Pd-negative sites were considerably farther south than our Pd-positive sites (Fig. 1). Thus, some of our results related to the effects of Pd status may have been related to geographical effects. While we were not able to account for all of these effects, we did account for temperature which is often related to latitude and elevation. We were also only able to include one belowground Pd-negative site in our study which may have affected our ability to detect the effects of hibernaculum type independent of Pd status and environmental conditions. We encourage more research on activity of tricolored bats as well as other species in Pd-negative belowground hibernacula to better assess their susceptibility to WNS should Pd arrive.
Our results suggest that tricolored bats using aboveground hibernacula such as culverts or bridges, may be less susceptible to the effects of WNS compared to those using subterranean sites due to increased activity and, in some areas, may not even be hibernating48. However, use of aboveground hibernacula may have some costs. For example, temperatures within aboveground hibernacula are far more variable than those in subterranean hibernacula42. Thus, bats using these structures may be subject to freezing events69. These bats may also be subject to greater predation rates because the sites are usually smaller and more easily accessible than subterranean sites70. Further, noise associated with traffic above bridges and culverts may decrease bats’ foraging efficiency71,72. Finally, the stream and river systems associated with bridges and culverts and used by bats as travel and foraging corridors, may also serve as a transmission corridor for Pd36. In the southeastern USA, this may result in Pd being transmitted to caves in karst regions of Florida and coastal Alabama that are currently Pd free. Tricolored bats in these caves will likely be susceptible to similar morbidity and mortality from WNS as tricolored bats in northwestern South Carolina, which have experienced 90% declines, due to similar torpor patterns37,50. However, some bats in subtropical caves, such as those in Florida, may leave these caves and move to aboveground hibernacula67. Thus, predicting the susceptibility of tricolored bats to the effects of Pd and WNS will require far more information on their behavior and torpor patterns during winter across the WNS-free zone.
Data availability
Data collected during and analyzed during the current study are available from the correspondingauthor on reasonable request.
References
Geiser, F. Hibernation eLS at (2011). https://doi.org/10.1002/9780470015902.a0003215.pub2
Humphries, M. M., Thomas, D. W. & Kramer, D. L. The role of energy availability in mammalian hibernation: A cost-benefit approach. Physiol. Biochem. Zool. 76, 165–179 (2003).
Prendergast, B. J., Freeman, D. A., Zucker, I. & Nelson, R. J. Periodic arousal from hibernation is necessary for initiation of immune responses in ground squirrels. Am. J. Physiol. - Regul. Integr. Comp. Physiol. 282, 1054–1062 (2002).
Lausen, C. L. & Barclay, R. M. R. Winter bat activity in the Canadian prairies. Can. J. Zool. 84, 1079–1086 (2006).
Bernard, R. F. & McCracken, G. F. Winter behavior of bats and the progression of white-nose syndrome in the southeastern United States. Ecol. Evol. 7, 1487–1496. https://doi.org/10.1002/ece3.2772 (2017).
Bernard, R. F., Willcox, E. V., Jackson, R. T., Brown, V. A. & McCracken, G. F. Feasting, not fasting: winter diets of cave hibernating bats in the United States. Front. Zool. 18, 1–13. https://doi.org/10.1186/s12983-021-00434-9 (2021).
Boyles, J. G., Dunbar, M. B. & WhitakerJr. J. O. Activity following arousal in winter in North America. Mamm. Rev. 36, 267–280 (2006).
Weller, T. J., Cryan, P. M. & O’Shea, T. J. Broadening the focus of bat conservation and research in the USA for the 21st century. Endanger. Species Res. 8, 129–145. https://doi.org/10.3354/esr00149 (2009).
Festa, F. et al. Bat responses to climate change: a systematic review. Biol. Rev. 98, 19–33. https://doi.org/10.1111/brv.12893 (2023).
Blehert, D. S. et al. Bat white-nose syndrome: an emerging fungal pathogen? Science 323, 227 (2009).
Verant, M. L. et al. White-nose syndrome initiates a cascade of physiologic disturbances in the hibernating bat host. BMC Physiol. 14 https://doi.org/10.1186/s12899-014-0010-4 (2014).
Coleman, J. T. H. & Reichard, J. D. Bat white-nose syndrome in 2014: a brief assessment seven years after discovery of a virulent fungal pathogen in North America. Outlooks Pest Manag. 25, 374–377. https://doi.org/10.1564/v25_dec_08 (2014).
Drees, K. P. et al. Phylogenetics of a fungal invasion: origins and widespread dispersal of white-nose syndrome. MBio 8, e01941–e01917. https://doi.org/10.1128/mBio.01941-17 (2017).
Leopardi, S., Blake, D. & Puechmaille, S. J. White-nose syndrome fungus introduced from Europe to North America. Curr. Biol. 25, R217–R219 (2015).
Cheng, T. L. et al. The scope and severity of white-nose syndrome on hibernating bats in North America. Conserv. Biol. 35, 1586–1597. https://doi.org/10.1111/cobi.13739 (2021).
Bernard, R. F., Foster, J. T., Wilcox, E. V., Parise, K. L. & McCracken, G. F. Molecular detection of the causative agent of white-nose syndrome on Rafinesque’s big-eared bats (Corynorhinus rafinesquii) and two species of migratory bats in the southeastern USA. J. Wildl. Dis. 51, 519–522. https://doi.org/10.7589/2014-08-202 (2015).
Udell, B. J. et al. Using mobile acoustic monitoring and false-positive N-mixture models to estimate bat abundance and population trends. Ecol. Monogr. 94, e1617. https://doi.org/10.1002/ecm.1617 (2024).
Moore, M. S. et al. Energy conserving thermoregulatory patterns and lower disease severity in a bat resistant to the impacts of white-nose syndrome. J. Comp. Physiol. B. 188, 163–176. https://doi.org/10.1007/s00360-017-1109-2 (2018).
Frank, C. L. et al. The resistance of a North American Bat species (Eptesicus fuscus) to white-nose syndrome (WNS). PLoS One. 9, e113958. https://doi.org/10.1371/journal.pone.0113958 (2014).
Ingala, M. R., Ravenelle, R. E., Monro, J. J. & Frank, C. L. The effects of epidermal fatty acid profiles, 1-oleoglycerol, and triacylglycerols on the susceptibility of hibernating bats to Pseudogymnoascus destructans. PLoS One. 12, e0187195. https://doi.org/10.1371/journal.pone.0187195 (2017).
Frank, C. L. et al. The effects of cutaneous fatty acids on the growth of Pseudogymnoascus destructans, the etiological agent of white-nose syndrome (WNS). PLoS One. 11, e0153535. https://doi.org/10.1371/journal.pone.0153535 (2016).
Frank, C. L., Sitler-Elbel, K. G., Hudson, A. J. & Ingala, M. R. The antifungal properties of epidermal fatty acid esters: insights from white-nose syndrome (WNS) in bats. Molecules 23 https://doi.org/10.3390/molecules23081986 (2018).
Vanderwolf, K. J., Kyle, C. J., Faure, P. A., Mcalpine, D. F. & Davy, C. M. Skin pH varies among bat species and seasons and between wild and captive bats. Conserv. Physiol. 9, coab088. https://doi.org/10.1093/conphys/coab088 (2021).
Johnson, J. S., Lacki, M. J., Thomas, S. C. & Grider, J. F. Frequent arousals from winter torpor in Rafinesque’s big-eared Bat (Corynorhinus rafinesquii). PLoS One. 7, e49754. https://doi.org/10.1371/journal.pone.0049754 (2012).
Johnson, J. S., Sharp, N. W., Monarchino, M. N., Lilley, T. M. & Edelman, A. J. No sign of infection in free-ranging Myotis austroriparius hibernating in the presence of Pseudogymnoascus destructans in Alabama. Southeast. Nat. 20, 20–28. https://doi.org/10.1656/058.020.0102 (2021).
Verant, M. L., Boyles, J. G., Waldrep, W. Jr., Wibbelt, G. & Blehert, D. S. Temperature-dependent growth of Geomyces destructans, the fungus that causes bat white-nose syndrome. PLoS One. 7, e46280. https://doi.org/10.1371/journal.pone.0046280 (2012).
Grider, J. F., Larsen, A. L., Homyack, J. A. & Kalcounis-Rueppell, M. C. Winter activity of coastal plain populations of bat species affected by white-nose syndrome and wind energy facilities. PLoS One. 11, e0166512. https://doi.org/10.1371/journal.pone.0166512 (2016).
Newman, B. A., Loeb, S. C. & Jachowski, D. S. Thermally unstable roosts influence winter torpor patterns in a threatened bat species. Conserv. Physiol. 12, coae014. https://doi.org/10.1093/conphys/coae014 (2024).
Reynolds, D. S., Shoemaker, K., von Oettingen, S. & Najjar, S. High rates of winter activity and arousals in two New England bat species: implications for a reduced white-nose syndrome impact? Northeast Nat. 24, B188–B208. https://doi.org/10.1656/045.024.s720 (2017).
Jackson, R. T., Willcox, E. V. & Bernard, R. F. Winter torpor expression varies in four bat species with differential susceptibility to white–nose syndrome. Sci. Rep. 12, 5688. https://doi.org/10.1038/s41598-022-09692-x (2022).
Fujita, M. S. & Kunz, T. H. Pipistrellus subflavus. Mamm. Species. 228, 1–6 (1984).
Ferrara, F. J. & Leberg, P. L. Characteristics of positions selected by day-roosting bats under bridges in Louisiana. J. Mammal. 86, 729–735. https://doi.org/10.1644/1545-1542(2005)086[0729:COPSBD]2.0.CO;2 (2005).
Newman, B. A., Loeb, S. C. & Jachowski, D. S. Winter roosting ecology of tricolored bats (Perimyotis subflavus) in trees and bridges. J. Mammal. 102, 1331–1341. https://doi.org/10.1093/jmammal/gyab080 (2021).
Sandel, J. K. et al. Use and selection of winter hibernacula by the eastern pipestrelle (Pipistrellus subflavus) in Texas. J. Mammal. 82, 173–178. http://dx.doi.org/10.1644/1545-1542(2001)082<0173:UASOWH>2.0.CO;2 (2001).
Meierhofer, M. B. et al. Structural and environmental predictors of presence and abundance of tri-colored bats in Texas culverts. J. Mammal. 99, 1274–1281. https://doi.org/10.1093/jmammal/gyz099 (2019).
Lutsch, K. E., McDonald, A. G., Gabriel, K. T. & Cornelison, C. T. Roadway-associated culverts may serve as a transmission corridor for Pseudogymnoascus destructans and white-nose syndrome in the coastal plains and coastal region of Georgia, USA. J. Wildl. Dis. 58, 322–332. https://doi.org/10.7589/JWD-D-21-00069 (2022).
Sirajuddin, P., Loeb, S. C., Britzke, E. R., Peoples, B. K. & Jachowski, D. S. Winter torpor patterns of tricolored bats (Perimyotis subflavus) in the southeastern U.S. J. Mammal. 106, 469-479. (2025). https://doi.org/10.1093/jmammal/gyae112
Reeder, D. M. et al. Frequent arousal from hibernation linked to severity of infection and mortality in bats with white-nose syndrome. PLoS One. 7, e38920. https://doi.org/10.1371/journal.pone.0038920 (2012).
Lilley, T. M. et al. White-nose syndrome survivors do not exhibit frequent arousals associated with Pseudogymnoascus destructans infection. Front. Zool. 13, (2016). https://doi.org/10.1186/s12983-016-0143-3
McGuire, L. P., Mayberry, H. W. & Willis, C. K. R. White-nose syndrome increases torpid metabolic rate and evaporative water loss in hibernating bats. Am. J. Physiol. Integr. Comp. Physiol. 313, R680–R686. https://doi.org/10.1152/ajpregu.00058.2017 (2017).
Gabriel, K. T. et al. Development of a multi-year white-nose syndrome mitigation strategy using antifungal volatile organic compounds. PLoS One. 17, e0278603. https://doi.org/10.1371/journal.pone.0278603 (2022).
Ferrari, M. Investigating biometrics of Perimyotis subflavus in traditional and nontraditional hibernacula in the Southeastern United States as it relates to susceptibility to white-nose syndrome. M.S. Thesis, Kennesaw State University (2022).
Brown, R. L. Hibernating tricolored bats at a southeastern recreation site : Microsite use and public perceptions. M.S. Thesis, Clemson University (2023).
Arndt, R. J. & Lima, S. L. Landscape-wide flight activity by wintering bats predictably follows pulses of warmth in the Midwestern United States. J. Mammal. 101, 1489–1501. https://doi.org/10.1093/jmammal/gyaa088 (2020).
Turbill, C. Winter activity of Australian tree-roosting bats: influence of temperature and climatic patterns. J. Zool. 276, 285–290. https://doi.org/10.1111/j.1469-7998.2008.00487.x (2008).
Park, K. J., Jones, G. & Ransome, R. D. Winter activity of a population of greater horseshoe bats (Rhinolophus ferrumequinum). J. Zool. 248, 419–427 (1999).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. https://doi.org/10.18637/jss.v067.i01 (2015).
Stevens, R. D. Winter activity of Perimyotis subflavus outside culverts in East Texas. West. North. Am. Nat. 84, 67–71. https://doi.org/10.3398/064.084.0106 (2024).
Shute, K. E., Loeb, S. C. & Jachowski, D. S. Seasonal shifts in nocturnal habitat use by coastal bat species. J. Wildl. Manage. 85, 964–978. https://doi.org/10.1002/jwmg.22060 (2021).
Loeb, S. C. & Winters, E. A. Changes in hibernating tricolored bat (Perimyotis subflavus) roosting behavior in response to white- nose syndrome. Ecol. Evol. 12, e9045. https://doi.org/10.1002/ece3.9045 (2022).
Schwab, N. A. & Mabee, T. J. Winter acoustic activity of bats in Montana. Northwest. Nat. 95, 13–27. https://doi.org/10.1898/NWN13-03.1 (2014).
Andersen, B. R., Stevens, R. D., Grimshaw, J. R. & Mcguire, L. P. Interspecific variation in lower temperature thresholds of an assemblage of wintering bats. J. Mammal. 105, 98–106. https://doi.org/10.1093/jmammal/gyad103 (2024).
Taylor, L. R. Analysis of the effect of temperature on insects in flight. J. Anim. Ecol. 32, 99–117 (1963).
Klüg-Baerwald, B. J., Gower, L. E., Lausen, C. L. & Brigham, R. M. Environmental correlates and energetics of winter flight by bats in southern Alberta, Canada. Can. J. Zool. 94, 829–836. https://doi.org/10.1139/cjz-2016-0055 (2016).
Czenze, Z. J. & Willis, C. K. R. Warming up and shipping out: arousal and emergence timing in hibernating little brown bats (Myotis lucifugus). J. Comp. Physiol. B. 185, 575–586. https://doi.org/10.1086/692623 (2015).
Barros, P. A., Ribeiro, C. & Cabral, J. A. Winter activity of bats in mediterranean peri-urban deciduous forests. Acta Chiropterologica. 19, 367–377. https://doi.org/10.3161/15081109ACC2017.19.2.013 (2017).
Besler, N. K. & Broders, H. G. Combinations of reproductive, individual, and weather effects best explain torpor patterns among female little brown bats (Myotis lucifugus). Ecol. Evol. 9, 5158–5171. https://doi.org/10.1002/ece3.5091 (2019).
Voigt, C. C., Schneeberger, K., Voigt-Heucke, S. L. & Lewanzik, D. Rain increases the energy cost of bat flight. Biol. Lett. 7, 793–795. https://doi.org/10.1098/rsbl.2011.0313 (2011).
Geipel, I., Smeekes, M. J., Halfwerk, W. & Page, R. A. Noise as an informational cue for decision-making: the sound of rain delays bat emergence. J. Exp. Biol. 222, jeb192005. https://doi.org/10.1242/jeb.192005 (2019).
Zahn, A. & Kriner, E. Winter foraging activity of central European vespertilionid bats. Mamm. Biol. 81, 40–45. https://doi.org/10.1016/j.mambio.2014.10.005 (2016).
Whiting, J. C., Doering, B., Aho, K. & Rich, J. Long–term patterns of cave–exiting activity of hibernating bats in western North America. Sci. Rep. 11, 8175. https://doi.org/10.1038/s41598-021-87605-0 (2021).
Rosales, E. & Thesis, M. S. Tri-colored bat habitat use and selection in northwestern South Carolina. Clemson University. (2024).
Whitaker, J. O. Jr. & Rissler, L. J. Winter activity of bats at a mine entrance in Vermillion County, Indiana. Am. Midl. Nat. 127, 52–59 (1992).
Whitaker, J. O. Jr. & Rissler, L. J. Seasonal activity of bats at Copperhead cave. Proc. Indiana Acad. Sci. 101, 127–134 (1992).
Fuller, N. W. et al. Disease recovery in bats affected by white-nose syndrome. J. Exp. Biol. 223, jeb211912. https://doi.org/10.1242/jeb.211912 (2020).
Liu, L. et al. Analyzing projected changes and trends of temperature and precipitation in the southern USA from 16 downscaled global climate models. Theor. Appl. Climatol. 109, 345–360. https://doi.org/10.1007/s00704-011-0567-9 (2012).
Smith, L. M., Sylvia, A., Doonan, T. J. & Gore, J. A. Decline of tricolored bats (Perimyotis subflavus) in Florida caves in winter. Southeast. Nat. 22, 136–153. https://doi.org/10.1656/058.022.0202 (2023).
Hoyt, J. R., Kilpatrick, A. M. & Langwig, K. E. Ecology and impacts of white-nose syndrome on bats. Nat. Rev. Microbiol. 19 https://doi.org/10.1894/0038-4909-66.4.333 (2021).
McSweeny, T. & Brooks, D. M. Some observations of severe weather events on a large urban population of free-tailed bats (Tadarida brasiliensis). Southwest. Nat. 66, 333–338. https://doi.org/10.1894/0038-4909-66.4.333 (2022).
McSweeny, T. & Brooks, D. M. Predation at an urban free-tailed bat (Tadarida brasiliensis) colony by night Herons. Southwest. Nat. 64, 60–63. https://doi.org/10.1894/0038-4909-64-1-60 (2019).
Siemers, B. M. & Schaub, A. Hunting at the highway: traffic noise reduces foraging efficiency in acoustic predators. Proc. R. Soc. B 278, 1646–1652 (2011). https://doi.org/10.1098/rspb.2010.2262
Bunkley, J. P. & Barber, J. R. Noise reduces foraging efficiency in pallid bats (Antrozous pallidus). Ethology 121, 1116–1121. https://doi.org/10.1111/eth.12428 (2015).
Acknowledgements
This study was funded in part by the U.S. Fish and Wildlife Service (Grant No. F20 AP10196 - 00) and the USDA Forest Service, Southern Research Station. We thank K. Morris, E. Ferrall, P. Pattavina, A. Pitt, N. Sharp, and Georgia-Alabama Land Trust for assistance with field work and providing access to field sites. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Author information
Authors and Affiliations
Contributions
S.C.L., C.T.C., T.M., and A.J.E. designed the study and obtained funding. S.C.L., E.A.W., R.G.B., J.R.A., M.F., J.U., and L.S. conducted field work. S.C.L. and W.C.B. analyzed data. S.C.L. wrote initial manuscript and all authors assisted with revisions.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics statement
We followed the U.S. Fish and Wildlife Service decontamination protocols at all sites and methods were approved by the USDA Forest Service Institutional Animal Use and Care Committee (#2021 - 002), the Kennesaw State University Institutional Animal Use and Care Committee (#21 − 002), and the University of West Georgia Institutional Animal Use and Care Committee (#1005). Methods were consistent with the ARRIVE guidelines and were conducted under state permits of Alabama (2019097049268680, 2021085096868680, 2022097843868680) and South Carolina (SC- 12 - 2020, SC- 12 - 2021, SC- 21 - 2022); work in Georgia and Florida was done under the supervision of Georgia Department of Natural Resources and Florida Fish and Wildlife Conservation Commission, respectively.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Loeb, S.C., Bridges, W.C., Winters, E.A. et al. Winter activity of tricolored bats in aboveground and subterranean hibernacula in the southeastern USA. Sci Rep 15, 13644 (2025). https://doi.org/10.1038/s41598-025-97703-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-97703-y
