
Abstract
According to the 2021 KDIGO guidelines, hydroxychloroquine (HCQ) had been recommended for the treatment of IgA nephropathy (IgAN). However, the precise mechanisms by which HCQ ameliorated proteinuria in IgAN were not fully understood. This study investigated the potential mechanisms of HCQ in reducing proteinuria in IgAN using network pharmacology and molecular docking approaches. Targets associated with IgAN, proteinuria, and HCQ were identified from databases including DisGeNET, GeneCards, OMIM, and PharmMapper. GO and KEGG analyses were conducted using the DAVID platform. Protein–protein interaction (PPI) networks were constructed using the STRING database, and hub genes were identified using Cytoscape software. The selection of hub genes was corroborated with data from the GEO database and validated through molecular docking. Additionally, miRNAs were predicted using NetworkAnalyst. A total of 48 genes were identified as being associated with the reduction of proteinuria in IgAN. The findings suggested that HCQ’s mechanism of action in mitigating proteinuria in IgAN primarily involved pathways related to inflammation. Furthermore, this mechanism was linked to the regulatory effects of miR-130b-3p on the expression of genes such as MMP2, IGF1, and PPARG. HCQ targeted miR-130b-3p, thereby influencing the TLR/MyD88/NF-κB signaling pathway and modulating the expression of MMP2, IGF1, and PPARG. This action may have been responsible for the observed reduction in proteinuria associated with IgAN.
Similar content being viewed by others

Introduction
IgA nephropathy (IgAN) is a common form of primary glomerulonephritis worldwide and a major cause of renal failure1. Proteinuria is a risk factor for impaired renal function in nephropathies and is associated with poor prognostic outcomes in IgAN2. Short-term reduction of proteinuria has been shown to significantly decrease the risk of disease progression in IgAN. In clinical practice, proteinuria reduction is often used as a surrogate outcome measure to evaluate the efficacy of IgAN treatments3. Angiotensin-converting enzyme inhibitors (ACEIs) and angiotensin receptor blockers (ARBs) are the mainstay of therapy for reducing proteinuria in IgAN. However, despite standard treatment with ACEIs/ARBs, more than 50% of patients still present with proteinuria levels exceeding 1 g/d4. Even after treatment with glucocorticoids, immunosuppressants, and biologics, proteinuria in some patients remains unimproved5. Therefore, it is essential to explore additional treatment strategies that can effectively and safely reduce proteinuria in IgAN patients.
Hydroxychloroquine (HCQ), a well-established antimalarial drug, has gained widespread application in the treatment of autoimmune diseases, exerting its effects through anti-inflammatory and immunomodulatory mechanisms6. Recent studies have demonstrated that HCQ could significantly reduce proteinuria in patients with IgAN7,8,9. Although its efficacy in reducing proteinuria might not have been as potent as that of glucocorticoids, immunosuppressants, and biologics, its favorable safety profile, characterized by fewer adverse reactions, had garnered ongoing interest in the treatment of IgAN10. According to the 2021 KDIGO guidelines, HCQ is recommended for Chinese IgAN patients at high risk of progressing to chronic kidney disease (CKD), even with adequate supportive management11. However, the specific mechanism by which HCQ reduces proteinuria in IgAN remains unclear. However, the specific mechanism by which HCQ reduced proteinuria in IgAN had remained unclear.
MicroRNA (miRNA) is identified as a key factor in the gene expression regulatory network. It regulate the expression of multiple genes, thereby influencing cell function and the occurrence and development of diseases12. The expression level of miRNA can be detected in blood and urine, which exhibited high stability and resistance to degradation. Consequently, miRNA is considered a potential biomarker for disease and a candidate for drug targeting13. MiRNA plays a significant regulatory role in the pathogenesis, progression, diagnosis, and management of IgAN14. Network pharmacology (NP), an integrative analytical approach bases on the interplay of disease, gene, drug, and target protein interaction networks, is employed to elucidate the mechanisms of drug action by integrating systems biology and computational biology with drug target-related “omics” data15. Several studies had applied NP to dissect the mechanisms of drug action in renal diseases, including IgAN and diabetic nephropathy (DN)16,17. Therefore, to uncover the molecular mechanisms by which HCQ reduced proteinuria in IgAN and to lay a theoretical groundwork for subsequent investigations, a NP analysis was performed in this study (Fig. 1).

A flowchart illustrating the process of HCQ alleviating proteinuria in IgAN.
Materials and methods
IgAN and albuminuria targets prediction
The DisGeNET18, GeneCard19, and Online Mendelian Inheritance in Man (OMIM)20 are powerful databases of human genes. With “IgA nephropathy” and “albuminuria” as keywords, the DisGeNET, GeneCard, and OMIM databases were searched for disease-related genes.
HCQ targets prediction and common targets intersection
The PubChem database contains data on over 100 million chemical substances, encompassing various types such as organic small molecules and bioactive molecules21. PharmMapper database is a highly valuable bioinformatics tool that focuses on serving the fields of drug development and molecular mechanism exploration. It relies on a massive built-in pharmacophore model to quickly and accurately predict potential drug target proteins that small molecules may act on22. UniProt, with its vast and authoritative protein database resources, collectes and organizes standard gene name information across a wide range of species, as well as corresponding detailed gene annotations23. The 2D and 3D chemical structures of HCQ were downloaded from the PubChem database, and the PharmMapper database was utilized for uploading the data (Fig. 2). The maximum conformation generation number in PharmMapper was set to 300. UniProt was employed to convert target gene identifiers into official names. A Venn diagram was constructed to identify the shared target genes between diseases and drugs24.

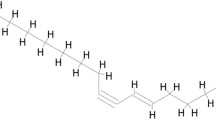
Chemical structure of HCQ.
GO and KEGG pathway analyses
The DAVID database is a powerful assistant in the field of bioinformatics, focusing on gene and protein information processing, including gene function annotation and gene pathway enrichment analysis25. To determine the key pathways influencing the reduction of albuminuria in IgAN by HCQ, the DAVID database was utilized for GO and KEGG analyses26. The results with P < 0.05 were evaluated using the Hiplot online platform and subsequently used for further analyses27.
Construction of a PPI network
The STRING database is a protein interaction network database widely used in the field of bioinformatics, encompassing a broad spectrum of data sources that includes experimentally validated interactions, computationally predicted interactions, literature mining, and information from other databases28. The targets were submitted to the STRING database to construct a protein–protein interaction (PPI) network. Data visualization and network analysis were conducted using Cytoscape 3.9.1 software, followed by the use of the cytoHubba plugin to identify core proteins.
Verification of the GEO dataset
The GEO database is a public database established and maintained by the National Center for Biotechnology Information (NCBI), playing a significant role in bioinformatics research. The database contains a wide range of data sources, including high-throughput gene expression data submitted by research institutions globally29. The dataset of human samples was searched using the keyword “IgA nephropathy” in the GEO database. Differential expression analysis was conducted using the GEO2R online tool, with the application of screening criteria “P < 0.05” and “|log2FC| > 2” to identify differentially expressed genes (DEGs). Subsequent to this analysis, a volcano plot was generated for annotating the core genes.
Molecular docking
Molecular docking (MD) is a computational technique used to predict interaction and binding patterns between molecules, with the aim of studying and optimizing the binding modes and mechanical interactions between compounds and proteins30. Autodocktools 1.5.7 was utilized to assess the interactions between the key targets (receptors) and HCQ (ligand), and PyMOL was employed for visualization purposes. The HCQ structure was extracted from the PubChem database, while the target structure was sourced from the Protein Data Bank (PDB)31. A Lamarckian genetic algorithm search was performed to execute 50 runs. A binding energy less than − 5 kcal/mol was indicative of good docking activity32, and proteins that stably bound to HCQ were selected as potential targets for their role in mitigating albuminuria in IgAN.
Construction of MiRNA networks
NetworkAnalyst integrates a substantial body of experimentally validated or computationally predicted miRNA-gene interaction data and employes corresponding algorithms to analyze input gene and miRNA datasets, thereby identifying potential interaction relationships33. In an effort to further elucidate the biological markers and miRNA regulatory mechanisms associated with HCQ’s role in reducing proteinuria in IgAN, genes that stably docked with HCQ were imported into the NetworkAnalyst database for gene-miRNA network analysis (Table 1). The prediction of miRNAs through genes is founded on sequence complementarity, which involves forecasting interactions between miRNAs and their target gene mRNAs based on this complementarity. Such complementary pairing may result in mRNA degradation or translation inhibition, thus modulating gene expression. Numerous studies had previously employed gene-miRNA network analysis to forecast biological markers and target miRNAs for conditions such as polycystic ovary syndrome34, kidney stones35, systemic lupus erythematosus36, and Parkinson’s disease37. Visualization and comprehensive analysis of the gene-miRNA network were performed using Cytoscape 3.9.1 software to identify candidate miRNAs.
Results
Preparation of the target
After duplicates were eliminated, 1,277 and 268 targets associated with IgAN and albuminuria were obtained from the OMIM, DisGeNET, and GeneCards databases, respectively. Following confirmation of the results with the UniProt database, 287 HCQ targets were identified based on the PharmMapper database. Diseases and drug targets were ascertained using the Venn platform, which disclosed 48 common targets (Fig. 3).

Venn diagram depicting overlapping genes associated with HCQ, IgAN, and Albuminuria”.
Enrichment of GO and KEGG
48 overlapping targets were imported into the DAVID database for subsequent GO and KEGG analyses. With a threshold set at P < 0.05, 163 biological processes (BP), 23 cellular components (CC), 46 molecular functions (MF), and 67 signaling pathways were identified. The top 30 GO terms and 20 KEGG terms were selected for further analysis (Figs. 4 and 5). The results indicated that HCQ reduced IgAN proteinuria through involvement in several pathways related to inflammation and oxidative stress, including the AGE-RAGE pathway, IL-17 pathway, and HIF-1 pathway.

GO functional enrichment analyses of common targets in HCQ-IgAN-albuminuria. BP: biological process; CC: cellular component; MF: molecular function.

KEGG pathway enrichment analyses of shared targets. Visualization by adjusted P-value color scale and genecount dot size.
PPI network establishment and analysis
To elucidate the protein–protein interaction (PPI) interactions and the underlying mechanisms, a PPI network was constructed from the STRING database and used to analyze the 48 common targets (Fig. 6). Among these, the top 10 targets—ALB, MMP9, CASP3, IGF1, SRC, PPARG, ACE, CCL5, NOS3, and MMP2—were selected for further investigation (Table 2).

PPI network of 47 shared targets. Nodes represent proteins. Edges represent the intersection between proteins. The colors and node sizes in the network reflected the integrated target number (degree).
Verification of the GEO dataset
Differential gene expression analysis was performed on the dataset GSE73953 obtained from the GEO database, comprising 15 samples of IgAN and 16 samples of normal kidney tissues. With P < 0.05 and |log2FC| > 2 as the selection criteria, the core genes—CASP3, IGF1, SRC, PPARG, NOS3, and MMP2—were identified as being downregulated in the differential expression analysis (Fig. 7).

Volcano plot of differentially expressed genes (DEGs) in the GEO dataset GSE73953. Blue represents downregulated DEGs, red represents upregulated DEGs, and grey represents not DEGs.
Verification via molecular docking
To further verify the results, molecular docking (MD) of the core targets and HCQ was conducted (Table 3). The findings disclosed stable binding structures between HCQ and MMP2, NOS3, PPARG, IGF1, and CASP3, with binding energies below − 5 kcal/mol (Figs. 8 and 9). The interactions of HCQ with these proteins suggested its potential for mitigating albuminuria in individuals with IgAN.

2D molecular docking structures of HCQ with core targets. (A): PPARG, (B): MMP2, (C): NOS3, (D): CASP3, (E): IGF1. The purple structure represents HCQ.

2D molecular docking structures of HCQ with core targets. (A): PPARG, (B): MMP2, (C): NOS3, (D): CASP3, (E): IGF1. Green represents the chemical structure of HCQ, blue represents the protein, and yellow dashed lines represent the hydrogen bonds formed between HCQ and the protein.
Construction of the MiRNA networks
Gene-miRNA network analysis disclosed a network comprising 171 nodes and 182 interaction lines, encompassing 5 genes and 166 predicted miRNAs. Notably, miR-130b-3p was the sole miRNA found to regulate the expression of more than two genes concurrently, indicating that HCQ may have mitigated albuminuria in IgAN by modulating the expression of MMP2, IGF1, and PPARG via miR-130b-3p (Fig. 10).

Gene-miRNA network. Orange circular nodes represent genes, and green triangular nodes represent miRNAs.
Discussion
Even low-risk IgAN patients were observed to progress to kidney failure within a decade38. The KDIGO guidelines highlighted that the management of IgAN predominantly focused on optimizing supportive care; however, this approach may not completely hinder disease progression. Furthermore, they recommended the use of HCQ for Chinese IgAN patients at high risk of progression39. While the mechanism by which HCQ reduces proteinuria in IgAN required further exploration to enhance clinical application. Our study employed network pharmacology (NP) and molecular docking (MD) techniques, revealing that HCQ might mitigate proteinuria in IgAN by targeting miR-130b-3p, thereby regulating the expression of MMP2, IGF1, and PPARG.
HCQ is an immunomodulator commonly used to treat autoimmune diseases and is also widely used in lupus nephritis (LN)40. The KDIGO guidelines recommended that all systemic lupus erythematosus (SLE) and LN patients should receive HCQ unless there were contraindications41. Based on the significant therapeutic effect of HCQ in LN, many researchers turned to IgAN as a treatment target for HCQ in recent years. A randomized controlled study found that the addition of HCQ to RAAS inhibition significantly reduced proteinuria in IgAN patients over a 6-month continuous treatment period7. A retrospective study with a 2-year follow-up period found that long-term use of HCQ significantly reduced proteinuria and was safer with fewer side effects compared to systemic corticosteroid therapy42. HCQ interfered with the body’s pH value, inhibited the activation of Toll-like receptors (TLRs), and alleviated the inflammatory response by suppressing the TLR/MyD88/NF-κB signaling pathway, thereby diminishing proteinuria in LN40. Similarly, as an immune-related disease, the TLR/MyD88/NF-κB signaling pathway also played an important role in the occurrence and development of IgAN43.
MiR-130b-3p, located within the human chromosomal region 22q11.22, was originally identified in tumor-related investigations44. In the context of kidney diseases, it was determined that miR-130b-3p plays a significant regulatory role in the onset and progression of LN, but there are no studies on miR-130b-3p in IgAN. A study found that miR-130b-3p levels were significantly higher in patients with early-stage LN compared to healthy controls and further elevated in advanced LN. The expression of miR-130b-3p in early LN positively correlated with the renal chronicity index and 24-h proteinuria levels. Additionally, transfection of miR-130b-3p inhibitor increased E-cadherin expression, decreased α-SMA expression, inhibited epithelial-mesenchymal transition, and ameliorated renal fibrosis45. Previous studies demonstrated that the activation of the TLR/MyD88/NF-κB signaling pathway was intimately associated with the development of renal fibrosis, and that renal fibrosis and damage in the LN mouse model could be mitigated by inhibiting the TLR9/MyD88/NF-κB signaling pathway46. Synthesizing these findings with prior research, the TLR/MyD88/NF-κB signaling pathway was identified as pivotal in the pathogenesis of IgAN. Our study discovered that miR-130b-3p played a crucial role in IgAN. Consequently, we hypothesized that miR-130b-3p might be involved in the pathogenesis and progression of IgAN by modulating the TLR/MyD88/NF-κB signaling pathway.
Our research revealed that HCQ might mitigate proteinuria in IgAN by modulating the expression of MMP2, IGF1, and PPARG via miR-130b-3p. Additionally, the TLR/MyD88/NF-κB signaling pathway was identified to influence the expression of MMP2, IGF1, and PPARG, thereby regulating cell proliferation and fibrosis in IgAN. In IgAN, MMP2 participated in the damage and repair processes of the glomerulus and tubulointerstitium, encompassing the degradation of extracellular matrix (ECM) and the synthesis of new matrix, which impacted the proliferation and fibrosis of renal cells47. Previous studies had demonstrated that miR-130b-3p could modulate the expression of MMP2, thereby regulating the proliferation, migration, and invasion of bladder cancer cells48. IGF-1 was shown to influence the infiltration of renal interstitial inflammatory cells and the accumulation of fibroblasts, contributing to the development of renal interstitial fibrosis49. MiR-130b-3p was found to regulate the secretion of IGF-1, thus affecting fibroblast activity50. The activation of PPARG was suggested to protect renal function by enhancing energy metabolism and reducing oxidative stress in IgAN51, and several studies had identified that miR-130b-3p could directly target and modulate PPARG expression52. Synthesizing the literature with our study findings, we hypothesized that miR-130b-3p impacts the TLR/MyD88/NF-κB signaling pathway, thereby regulating the expression of MMP2, IGF1, and PPARG.
In conclusion, our study revealed that miR-130b-3p was a key intersecting miRNA in the pharmacological mechanism of HCQ and the pathogenesis of IgAN. Integrating our findings with prior discussions, we hypothesized that HCQ might exert therapeutic effects on IgAN by modulating the levels of miR-130b-3p. Synthesizing our results with previous studies, we postulated that HCQ targeted miR-130b-3p, thereby influencing the TLR/MyD88/NF-κB signaling pathway and regulating the expression of MMP2, IGF1, and PPARG, which could contribute to the reduction of IgAN proteinuria.
Nevertheless, our study had limitations, as cell and animal experiments were not conducted due to budgetary constraints. Further experimental validation was necessary to ascertain whether HCQ modulated the expression of MMP2, IGF1, and PPARG genes via miR-130b-3p, thereby influencing the TLR4/NF-κB/MAPK signaling pathway and potentially mitigating albuminuria in IgAN. Moreover, the potential of miR-130b-3p as a novel non-invasive biomarker for assessing disease progression and prognosis needed to be validated.
Conclusion
HCQ targeted miR-130b-3p, thereby influencing the TLR/MyD88/NF-κB signaling pathway and regulating the expression of MMP2, IGF1, and PPARG, which contributed to the reduction of IgAN proteinuria.
Data availability
All data generated or analysed during this study are included in this article.
References
Chen, T. et al. Prediction and risk stratification of kidney outcomes in IgA nephropathy. Am. J. Kidney Dis. 74, 300–309. https://doi.org/10.1053/j.ajkd.2019.02.016 (2019).
Wong, M. G. et al. The therapeutic evaluation of steroids in IgA nephropathy global (TESTING) study: Trial design and baseline characteristics. Am. J. Nephrol. 52, 827–836. https://doi.org/10.1159/000519812 (2021).
Canney, M. et al. Quantifying duration of proteinuria remission and association with clinical outcome in IgA nephropathy. J. Am. Soc. Nephrol. 32, 436–447. https://doi.org/10.1681/asn.2020030349 (2021).
Barratt, J. et al. IgA nephropathy patient baseline characteristics in the sparsentan PROTECT study. Kidney Int. Rep. 8, 1043–1056. https://doi.org/10.1016/j.ekir.2023.02.1086 (2023).
Sarafidis, P. A., Memmos, E., Alexandrou, M. E. & Papagianni, A. Mineralocorticoid receptor antagonists for nephroprotection: Current evidence and future perspectives. Curr. Pharm. Des. 24, 5528–5536. https://doi.org/10.2174/1381612825666190306162658 (2018).
Desmarais, J. & Link, M. S. Hydroxychloroquine Not a heart breaker! Arthritis Care Res. (Hoboken) 73, 770–771. https://doi.org/10.1002/acr.24493 (2021).
Liu, L. J. et al. Effects of hydroxychloroquine on proteinuria in IgA nephropathy: A randomized controlled trial. Am. J. Kidney Dis. 74, 15–22. https://doi.org/10.1053/j.ajkd.2019.01.026 (2019).
Tang, C. et al. Long-term safety and efficacy of hydroxychloroquine in patients with IgA nephropathy: A single-center experience. J. Nephrol. 35, 429–440. https://doi.org/10.1007/s40620-021-00988-1 (2022).
Tang, C. et al. The efficacy and safety of hydroxychloroquine in pregnant patients with IgA nephropathy: A retrospective cohort study. Nephrol. (Carlton) 27, 155–161. https://doi.org/10.1111/nep.13991 (2022).
Petrou, D., Kalogeropoulos, P., Liapis, G. & Lionaki, S. IgA nephropathy: Current treatment and new insights. Antibodies (Basel). https://doi.org/10.3390/antib12020040 (2023).
Rovin, B. H. et al. Executive summary of the KDIGO 2021 guideline for the management of glomerular diseases. Kidney Int. 100, 753–779. https://doi.org/10.1016/j.kint.2021.05.015 (2021).
Genedy, H. H. et al. MicroRNA-targeting nanomedicines for the treatment of intervertebral disc degeneration. Adv. Drug Deliv Rev. 207, 115214. https://doi.org/10.1016/j.addr.2024.115214 (2024).
Ho, P. T. B., Clark, I. M. & Le, L. T. T. MicroRNA-based diagnosis and therapy. Int. J. Mol. Sci. https://doi.org/10.3390/ijms23137167 (2022).
Yao, X. et al. MicroRNAs in IgA nephropathy. Ren. Fail. 43, 1298–1310. https://doi.org/10.1080/0886022x.2021.1977320 (2021).
Nogales, C. et al. Network pharmacology: Curing causal mechanisms instead of treating symptoms. Trends Pharmacol. Sci. 43, 136–150. https://doi.org/10.1016/j.tips.2021.11.004 (2022).
Chen, Y., Lu, M., Lin, M. & Gao, Q. Network pharmacology and molecular docking to elucidate the common mechanism of hydroxychloroquine treatment in lupus nephritis and IgA nephropathy. Lupus 33, 347–356. https://doi.org/10.1177/09612033241230377 (2024).
Lu, M., Ou, J., Deng, X., Chen, Y. & Gao, Q. Exploring the Pharmacological mechanisms of tripterygium wilfordii against diabetic kidney disease using network pharmacology and molecular docking. Heliyon 9, e17550. https://doi.org/10.1016/j.heliyon.2023.e17550 (2023).
Pinero, J. et al. The disgenet knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 48, D845–D855. https://doi.org/10.1093/nar/gkz1021 (2020).
Lin, Y. & Hu, Z. Bioinformatics analysis of candidate genes involved in ethanol-induced microtia pathogenesis based on a human genome database: genecards. Int. J. Pediatr. Otorhinolaryngol. 142, 110595. https://doi.org/10.1016/j.ijporl.2020.110595 (2021).
Hamosh, A., Amberger, J. S., Bocchini, C., Scott, A. F. & Rasmussen, S. A. Online Mendelian inheritance in man (OMIM®): Victor McKusick’s magnum opus. Am. J. Med. Genet. A 185, 3259–3265. https://doi.org/10.1002/ajmg.a.62407 (2021).
Kim, S. et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 49, D1388–D1395. https://doi.org/10.1093/nar/gkaa971 (2021).
Wang, X. et al. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 45, W356–W360. https://doi.org/10.1093/nar/gkx374 (2017).
UniProt. The universal protein knowledgebase in 2021. Nucleic Acids Res. 49, D480–D489. https://doi.org/10.1093/nar/gkaa1100 (2021).
Jia, A., Xu, L. & Wang, Y. Venn diagrams in bioinformatics. Brief. Bioinform. 22, bbab108. https://doi.org/10.1093/bib/bbab108 (2021).
Sherman, B. T. et al. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 50, W216–w221. https://doi.org/10.1093/nar/gkac194 (2022).
Kanehisa, M., Furumichi, M., Tanabe, M., Sato, Y. & Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 45, D353–d361. https://doi.org/10.1093/nar/gkw1092 (2017).
Li, J. et al. Hiplot: A comprehensive and easy-to-use web service for boosting publication-ready biomedical data visualization. Brief. Bioinform. https://doi.org/10.1093/bib/bbac261 (2022).
Szklarczyk, D. et al. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 49, D605–d612. https://doi.org/10.1093/nar/gkaa1074 (2021).
Wang, Y. et al. Mechanisms underlying the therapeutic effects of Qingfeiyin in treating acute lung injury based on GEO datasets, network pharmacology and molecular docking. Comput. Biol. Med. 145, 105454. https://doi.org/10.1016/j.compbiomed.2022.105454 (2022).
Stanzione, F., Giangreco, I. & Cole, J. C. Use of molecular docking computational tools in drug discovery. Prog Med. Chem. 60, 273–343. https://doi.org/10.1016/bs.pmch.2021.01.004 (2021).
Velankar, S., Burley, S. K., Kurisu, G., Hoch, J. C. & Markley, J. L. The protein data bank archive. Methods Mol. Biol. 2305, 3–21. https://doi.org/10.1007/978-1-0716-1406-8_1 (2021).
Li, B., Rui, J., Ding, X. & Yang, X. Exploring the multicomponent synergy mechanism of Banxia Xiexin decoction on irritable bowel syndrome by a systems pharmacology strategy. J. Ethnopharmacol. 233, 158–168. https://doi.org/10.1016/j.jep.2018.12.033 (2019).
Burley, S. K. et al. Protein data Bank: A Comprehensive Review of 3D Structure Holdings and Worldwide Utilization by Researchers, Educators, and Students. Biomolecules. https://doi.org/10.3390/biom12101425 (2022).
Heidarzadehpilehrood, R., Pirhoushiaran, M., Binti Osman, M., Ling, K. H. & Abdul Hamid, H. Unveiling key biomarkers and therapeutic drugs in polycystic ovary syndrome (PCOS) through pathway enrichment analysis and hub Gene-miRNA networks. Iran. J. Pharm. Res. 22, e139985. https://doi.org/10.5812/ijpr-139985 (2023).
Gao, Y. et al. Identification of biomarkers and potential therapeutic targets of kidney stone disease using bioinformatics. World J. Urol. https://doi.org/10.1007/s00345-023-04704-5 (2024).
Peng, Z. et al. Exploration of the molecular mechanisms, shared gene signatures, and MicroRNAs between systemic lupus erythematosus and diffuse large B cell lymphoma by bioinformatics analysis. Lupus 31, 1317–1327. https://doi.org/10.1177/09612033221114578 (2022).
Zhu, L., Chen, D., Wang, X. & He, C. Identification and validation of oxidative stress-related hub genes in Parkinson’s disease. Mol. Neurobiol. https://doi.org/10.1007/s12035-024-04622-6 (2024).
Pitcher, D. et al. Long-term outcomes in IgA nephropathy. Clin. J. Am. Soc. Nephrol. 18, 727–738. https://doi.org/10.2215/cjn.0000000000000135 (2023).
Kidney disease: Improving global outcomes glomerular diseases work, G. KDIGO 2021 Clinical Practice Guideline for the Management of Glomerular Diseases. Kidney Int 100, S1–S276. https://doi.org/10.1016/j.kint.2021.05.021 (2021).
Rao, I. R. et al. Hydroxychloroquine in nephrology: Current status and future directions. J. Nephrol. 36, 2191–2208. https://doi.org/10.1007/s40620-023-01733-6 (2023).
KDIGO 2024 Clinical Practice Guideline for the Evaluation and Management of Chronic Kidney Disease. Kidney Int. 105, S117–S314. https://doi.org/10.1016/j.kint.2023.10.018 (2024).
Lei, S. F. et al. Comparison between hydroxychloroquine and systemic corticosteroids in IgA nephropathy: A 2-year follow-up study. BMC Nephrol. 24 175 (2023).
Ciferska, H. et al. Does the renal expression of Toll-like receptors play a role in patients with IgA nephropathy? J. Nephrol. 33, 307–316. https://doi.org/10.1007/s40620-019-00640-z (2020).
Lv, M. et al. Exosomal miR-130b-3p suppresses metastasis of non-small cell lung cancer cells by targeting DEPDC1 via TGF-β signaling pathway. Int. J. Biol. Macromol. 275, 133594. https://doi.org/10.1016/j.ijbiomac.2024.133594 (2024).
Wang, W. et al. Up-regulation of serum MiR-130b-3p level is associated with renal damage in early lupus nephritis. Sci. Rep. 5, 12644. https://doi.org/10.1038/srep12644 (2015).
Yu, T. et al. Extracellular HMGB1 induced glomerular endothelial cell injury via TLR4/MyD88 signaling pathway in lupus nephritis. Mediators Inflamm. 2021 9993971. https://doi.org/10.1155/2021/9993971 (2021).
Danilewicz, M. & Wagrowska-Danilewicz, M. Differential glomerular Immunoexpression of matrix metalloproteinases MMP-2 and MMP-9 in idiopathic IgA nephropathy and Schoenlein–Henoch nephritis. Folia Histochem. Cytobiol. 48, 63–67. https://doi.org/10.2478/v10042-008-0086-4 (2010).
Yu, X. et al. LncRNA CASC15 functions as an oncogene by sponging miR-130b-3p in bladder cancer. Eur. Rev. Med. Pharmacol. Sci. 24, 7203. https://doi.org/10.26355/eurrev_202007_21853 (2020).
Wei, L. et al. Rs1520220 and Rs2195239 polymorphisms of IGF-1 gene associated with histopathological grades in IgA nephropathy in Northwestern Chinese Han population. Kidney Blood Press. Res. 43, 80–87. https://doi.org/10.1159/000486914 (2018).
Expression of Concern. miR-130b-3p modulates Epithelial-Mesenchymal crosstalk in lung fibrosis by targeting IGF-1. PLoS One 17, e0263701. https://doi.org/10.1371/journal.pone.0263701 (2022).
Song, J. et al. Peroxisome proliferator-activated receptor gamma C161T polymorphisms and survival of Japanese patients with Immunoglobulin A nephropathy. Clin. Genet. 64, 398–403. https://doi.org/10.1034/j.1399-0004.2003.00154.x (2003).
Zhang, J. et al. miR-130b-3p involved in the pathogenesis of age-related hearing loss via targeting PPARγ and autophagy. Hear. Res. 449, 109029. https://doi.org/10.1016/j.heares.2024.109029 (2024).
Acknowledgements
We are appreciated for all the public databases and websites applied in the present study. This work was supported by the Fujian Provincial Nature and Science Foundation (Grant No. 2025J01120421) and the Zhongshan Hospital of Xiamen University Discipline Reserve Talent Training Program 2024.
Author information
Authors and Affiliations
Contributions
Qing Gao conceived the experiments. Yuxin Wang and Zheng Wan designed the study. Meiqi Lu performed the experiments and wrote the paper. Yixuan Chen and Mengshu Lin analyzed and interpreted the data. Xiaoqi Deng, and Xiaoxia Su contributed materials and analysis tools.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lu, M., Wang, Y., Wan, Z. et al. Exploring the mechanism of proteinuria reduction by hydroxychloroquine in IgA nephropathy using network pharmacology and molecular mocking. Sci Rep 15, 20836 (2025). https://doi.org/10.1038/s41598-025-97950-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-97950-z
