
Abstract
Documentation of biodiversity and its geographical distribution is necessary to understand the processes and drivers of evolutionary diversification as well as to guide conservation and management initiatives. Among the most emblematic patterns of biodiversity in the world’s oceans is the Coral Triangle (Indo-Australian Archipelago), widely recognized to be the center of species richness for a variety of marine life forms. The distribution of biodiversity remains incompletely documented, however, for a majority of reef-associated invertebrate taxa, including the zooxanthellate soft corals (Octocorallia) that dominate hard substrate on many Indo-Pacific reefs. We used a genetic approach to document the diversity of Indo-Pacific soft corals, sequencing two single-locus barcoding markers for > 4400 soft coral specimens and assigning individuals to molecular operational taxonomic units as proxies of species. We document two centers of species richness for zooxanthellate soft corals, one in the Indo-Australian Archipelago and a second, equally diverse center in the Western Indian Ocean. Centers of endemicity for soft corals are coincident with these centers of species richness, although the peripheral Red Sea and Hawaii also support high proportions of endemic taxa. The patterns documented here suggest that biogeographic distributions of soft coral families may be driven in part by larval dispersal potential: taxa with benthic larvae are absent from most oceanic islands of the central Pacific and are represented by higher proportions of endemic taxa in other geographic regions. Our findings demonstrate the distinct biogeographic patterns among reef taxa and underscore the need to document and analyze species distributions of more reef-associated invertebrate groups to derive a complete picture of reef biogeography.
Similar content being viewed by others


Introduction
As the impacts of anthropogenic activity on the world’s oceans alter the abundance and distribution of species, it has become more critical than ever to document both regional and global patterns of biodiversity. Recognition of those patterns is necessary in order to begin to understand the underlying drivers of biodiversity1,2, how those patterns may shift in the future with ongoing environmental change3, and to establish priorities for conservation and management4,5,6. To date, patterns of marine biodiversity and designations of biodiversity hotspots (i.e. areas where high biodiversity coincides with high anthropogenic threats4) have been established based largely on studies of a few well-known taxa such as the shallow-water reef-building scleractinian corals7,8,9, coral reef-associated fish10,11,12,13, some mobile macroinvertebrates4,14,15, and macroalgae16,17. For the vast majority (70–90%) of marine taxa, however, global patterns and the underlying drivers of biodiversity remain unassessed2. Both lack of sampling and a lack of the taxonomic expertise necessary to identify taxa precisely across all geographic regions are major impediments to the documentation of marine biodiversity18,19.
Quantifying patterns of biodiversity necessitates discriminating species (i.e., independently evolving lineages20) and accurately assessing their geographic ranges. With the application of genetic tools to assess species boundaries it has become clear that many “well known” species assumed to occupy wide geographic ranges are in fact complexes of species that may or may not be sister lineages, some with far more restricted ranges than previously believed21,22,23. As a result of these revised understandings of species boundaries and the accompanying recognition that morphological characters formerly assumed to be species-diagnostic may not be, historical records of species occurrences are an increasingly unreliable source of information for assessing species richness and geographic ranges23. For example, a search for records of the reef-building scleractinian Acropora tenuis would suggest that this species has a pan-Indo-Pacific distribution from the Red Sea to French Polynesia24, whereas a recent revision of the species supported by molecular evidence suggests that A. tenuis is narrowly restricted to the SW Pacific23. Assessment of biodiversity and biogeography based on the distribution of genetic lineages in addition to historically recognized morphospecies concepts is therefore necessary to better our understanding of the coral reef ecosystem.
Octocorals (soft corals, sea fans, whips and sea pens) are among the most common and conspicuous sessile invertebrate macrofauna in shallow coral reef communities. A majority of the octocoral species that dominate these communities are zooxanthellate, engaging in photosymbioses with the same clades of symbiodiniacean dinoflagellates found in scleractinian corals25,26, therefore playing similar ecological roles and subject to the same environmental stressors27,28. Octocorals may compete for space with scleractinians29 while also providing food, habitat complexity and shelter to numerous other taxa30,31,32. Some octocorals engineer massive reef structures33 or contribute to reef accretion34,35. There is growing evidence that octocoral-dominated communities may represent an alternative state to scleractinian-dominated reefs and that shifts to octocoral-dominated communities are increasing in frequency32,36.
On many shallow-water reefs throughout the Indo-Pacific, large, fleshy soft corals (i.e. octocorals that lack a skeletal axis) are among the most abundant space-occupying taxa, particularly in certain inshore, nutrient-rich communities37,38. Despite their ubiquity and ecological importance, zooxanthellate soft corals have been much less well studied than scleractinians, and both their overall diversity and their geographic distribution are poorly understood39. Species delimitation in these soft coral taxa is especially challenging40,41. They exhibit environmental plasticity in colony growth form42; little variation in soft tissue characters such as polyp structure; and do not form a rigid skeleton39. Inadequately or inaccurately illustrated descriptions in the historical literature, missing type specimens, and the need for taxonomic revisions of most major taxa represent additional challenges to reliable identification and discrimination of species39,40,43.
Over the past two decades, application of genetic tools to the study of octocorals has changed our understanding of phylogenetic relationships at all taxonomic levels44. Numerous cryptic species complexes have been identified within taxa previously considered to have widespread geographic distributions45,46,47. Genetic studies have also revealed the need to synonymize some taxa that have been erroneously split based on unreliable or variable morphological characters40. As a result of these advances in our understanding of species boundaries, existing information on the number and geographic distributions of taxa from morphological identifications alone requires revision48. Here, we used a molecular approach to identify genetic lineages (i.e. molecular operational taxonomic units, or MOTUs) as proxies of nominal species. Using sequences from two DNA barcode markers (mtMutS, 28S rDNA), we assigned approximately 4400 specimens to MOTUs in order to assess for the first time the overall biodiversity and biogeographic distribution of shallow-water, zooxanthellate soft corals across the Indo-Pacific.
Methods
Collections
Soft corals were collected during biodiversity surveys conducted throughout the Indo-Pacific during ~ 1998–2023 (Supplementary Table S1), using SCUBA to depths of ~ 30 m. Colonies were collected haphazardly, without quantifying relative abundance, but attempts were made to sample representatives of all distinguishable morphospecies. Only reef-dwelling, zooxanthellate soft coral taxa were included in these analyses, i.e. species belonging to families Capnellidae, Cladiellidae, Lemnaliidae, Sarcophytidae, Sinulariidae, Xeniidae, and the genus Litophyton (Nephtheidae) (Supplementary Table S2). Whole colonies or colony fragments were preserved in > 70% ethanol; for more recent collections (since ~ 2005), small subsamples of tissue for DNA analysis were also preserved separately in > 95% ethanol. All collections have been deposited in museums (Supplementary Table S2); some of the collections and taxa included in our analyses have been the focus of previous regional and taxonomic publications (Supplementary Table S1).
DNA barcoding and species delimitation
DNA was extracted from preserved tissues using a Qiagen DNeasy Blood & Tissue kit. Fragments of the mitochondrial gene mtMutS and nuclear 28S ribosomal DNA—both standard DNA barcodes used for octocorals—were amplified by polymerase chain reaction (PCR) with previously published primers and protocols49 and sent for Sanger-sequencing (Retrogen Inc., San Diego). Edited contigs were aligned using MAFFT v.550 with the FFT-NS-i method. For many specimens, only one of the two barcoding markers was sequenced successfully. Consequently, alignments for mtMutS and 28S were analyzed separately for each taxon.
Following previous analyses of octocoral biodiversity48,49, mothur v. 1.4851 was used to define molecular operational taxonomic units (MOTUs) (considered here as proxies of species) based on average genetic distance thresholds of 0.003 (mtMutS) and 0.005 (28S rDNA). In previous studies, a threshold of 0.003 applied to mtMutS yielded the highest concordance between MOTUs and morphospecies identifications; for 28S rDNA, a more conservative threshold of 0.005 yielded comparable concordance and minimized false positives49. Pairwise genetic distances (uncorrected p) among sequences were calculated using dist.seqs (calc = onegap, countends = F, cutoff = 0.1, output = lt), partitioned into MOTUs using cluster (method = average, precision = 1000), and bin.seqs was then used to output a fasta file with sequences labeled by MOTU identity.
Species richness and phylogenetic diversity
Collection locations (Fig. 1) were assigned to marine provinces52 for analysis of patterns of biodiversity. Occupancy matrices were constructed to reflect the numbers and presence or absence of each MOTU in each province. For each genus or family, we constructed separate matrices for MOTUs defined using mtMutS or 28S rDNA. All taxa were also combined to generate occupancy matrices reflecting the total number of MOTUs of all zooxanthellate soft corals recorded in all provinces. The number of MOTUs (i.e. species) detected is a function of sampling effort53. Because sample sizes differed by as much as two orders of magnitude among provinces (Table 1, Supplementary Fig. S1), we standardized estimates of species richness using a coverage-based rarefaction and extrapolation method53 as implemented in iNEXT Online (v. March 2024)54,55. Species richness was standardized by sample coverage (SC) where 1 – SC is the probability that the next individual sampled from a community will be a species that has not yet been found53. Base sample coverage (i.e., expected SC at 2X the actual sample size) was estimated for each province, and a combination of rarefaction and extrapolation (R/E) was used to compare species richness at the lowest base sample coverage value observed among the majority of provinces (Table 1). Species richness per province was also standardized separately for taxa that brood larvae (Capnellidae, Lemnaliidae, Xeniidae, Litophyton) and those that broadcast spawn (Cladiellidae, Sarcophytidae, Sinulariidae) based on published information56.

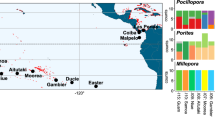
Map of locations at which zooxanthellate soft corals were sampled. Circles are scaled to indicate the number of specimens sequenced from each location (see Table 1). Locations are listed in Supplementary Table S1. Colors and numbers denote different marine provinces52: 09 – Warm Temperate NW Pacific; 18 – Red Sea and Gulf of Aden; 19 – Somali/Arabian; 20 – Western Indian Ocean; 25 – South China Sea; 26 – Sunda Shelf; 27 – Java Transitional; 28 – South Kuroshio; 29 – Tropical NW Pacific; 30 – Western Coral Triangle; 31– Eastern Coral Triangle; 32 – Sahul Shelf; 33 – NE Australian Shelf; 34 – NW Australian Shelf; 35 – Tropical SW Pacific; 36 – Lord Howe and Norfolk Islands; 37 – Hawaii; 38 – Marshall, Gilbert and Ellis Islands; 39 – Central Polynesia; 40 – SE Polynesia. Broken lines delineate marine realms52.
Phylogenetic species variability (PSV), a measure of phylogenetic disparity (or lack of evolutionary redundancy) among species57, was calculated for each province using ‘picante’58,59. For each barcode marker, a phylogenetic tree that included a single representative of each MOTU was constructed using maximum likelihood in IQTree v. 2.1.260 with an edge-linked partition model61 (Supplementary Figs. S1, S2). The optimal model of evolution for each gene (mtMutS: HKY + F + I + G4; 28S: GTR + F + R4) was found using ModelFinder62.
Compositional betadiversity
To estimate β-diversity among provinces the ‘betapart’ package58,63 was used to calculate pairwise values of Sørensen’s dissimilarity index (βSOR) based on MOTU composition in each province. β-diversity was further partitioned into components of nestedness (βSNE) and turnover (βSIM), where nestedness accounts for differences in species composition resulting from one community having a subset of the same species found in another, while turnover reflects the occurrence of different species between communities. Pairwise dissimilarities among provinces (βSOR) were used to create non-metric multidimensional-scaling (nMDS) ordination plots in Primer E v 664. Hierarchical clustering with group average linking was also conducted and the resulting dissimilarity groupings from the dendrograms were overlaid on the respective nMDS plots.
Results
A total of 3597 mtMutS sequences (~ 735 bp) and 3765 28S rDNA sequences (~ 710 bp) were obtained and analyzed from 4441 specimens (Supplementary Table S2). Numbers of specimens sequenced for each barcode marker varied greatly among taxa and marine provinces52, reflecting both natural variation in diversity and abundance as well as differences in collection effort (Fig. 2a,b; Supplementary Table S3). Family Sarcophytidae (i.e., genera Lobophytum, Sarcophyton, Sclerophytum) was the most sampled, with a total of 2120 mtMutS and 1994 28S sequences, while only 86 mtMutS and 60 28S sequences were obtained for Capnellidae (Capnella). For Cladiellidae only the 28S marker was sequenced due to the relative invariance of mtMutS among species within the family65. Numbers of sequences recovered per marine province ranged from 501 (mtMutS) and 498 (28S) from the Western Indian Ocean (province 20) to only one (mtMutS) and two (28S) sequences from Hawaii (province 37) (Table 1). In general, sampling effort and number of sequences obtained were lower for the easternmost Indo-Pacific provinces (37–40), Coral Triangle (provinces 30–31) and SW Pacific (provinces 35–36) as most specimens came from older collections available in museum repositories (Supplementary Table S2). No collections were available for analysis from several provinces in the central and northern Indian Ocean (provinces 21–24).

Number of sequences obtained per octocoral family and marine province for (a) 28S rDNA and (b) mtMutS barcoding markers. Number of molecular operational taxonomic units (MOTUs) delineated per province based on species delimitation using (c) 28S rDNA or (d) mtMutS. No mtMutS sequences were obtained for Cladiellidae. (See also Supplementary Fig. S3 for sampling curves relating number of MOTUs detected to number of sequences obtained.)
Species richness and phylogenetic diversity
Numbers of MOTUs estimated using the two different markers differed, with mtMutS generally yielding more conservative estimates than 28S rDNA (Fig. 2c,d; Table 1). Total number of MOTUs of all families estimated with 28S was 471; excluding Cladiellidae, it was 426 using 28S and 295 using mtMutS. Estimates of numbers of MOTUs per family were highest for Sarcophytidae (mtMutS = 159; 28S = 188) and lowest for Sinulariidae (mtMutS = 8; 28S = 10) (Fig. 2c, d; Supplementary Table S3). The greatest discrepancy in numbers of MOTUs estimated using different markers was for Litophyton, for which values ranged from 8 (mtMutS) to 46 (28S).
Species richness per province standardized by rarefaction/extrapolation at a sample coverage of 0.85 ranged from 2 to 99 (Fig. 3a,f; Table 1; Supplementary Fig. S4). The highest species richness estimated using both 28S and mtMutS was found in the Western Indian Ocean, NE Australian Shelf and Western Coral Triangle (Fig. 3a,f). Provinces with significantly lower species richness (based on non-overlapping 95% confidence intervals48) included the Red Sea and NW Indian Ocean; Sunda Shelf; and Warm Temperate NW Pacific (Supplementary Fig. S4). Species richness was lowest in several of the easternmost provinces (Hawaii, SE Polynesia). Estimates of species richness for Central Polynesia, Lord Howe Is. and the Marshall Islands were also low but could not be extrapolated to SC = 0.85 due to low sample coverage (SC < 0.72). As a result, the species richness estimates for those provinces have high uncertainty (Supplementary Fig. S4) and should be interpreted with caution.

Distribution of zooxanthellate soft corals by province52. Panels (a–e): MOTUs determined using 28S rDNA; (f–j) MOTUs determined using mtMutS (a,f) Species richness estimated by rarefaction/extrapolation to a standard sample coverage of 0.8555; (b,g) Endemicity (proportion of MOTUs recorded in a single province); (c,h) PSV, phylogenetic species variability57; standardized species richness of (d,i) brooding and (e,j) broadcast-spawning clades.
Phylogenetic species variability (PSV)59 was also lowest in the easternmost Indo-Pacific (provinces 38–40) and Java Transitional (PSV < 0.5), reflecting the absence of several families from those regions (Fig. 3c,h; Table 1; Supplementary Fig. S4). PSV was also relatively low in the South China Sea (PSV < 0.5) where octocoral communities tended to be dominated disproportionately by Sarcophytidae (Fig. 2). Among all other provinces, PSV was uniformly quite high (PSV = 0.5–0.750) with overlapping 95% confidence intervals (Supplementary Fig. S4), reflecting the diversity of families and genera found at most locations.
Low sample coverage for some taxa precluded using rarefaction/extrapolation to compare standardized species richness of individual families. Observed numbers of MOTUs of each family varied greatly among provinces, although all families exhibited high richness of MOTUs in the Western Indian Ocean and NE Australian Shelf (Fig. 2c,d; Supplementary Table S3). Sarcophytidae was the most species-rich family overall and the only family to be sampled in every province. In addition to the Western Indian Ocean and NE Australian Shelf, many Sarcophytidae MOTUs were sampled in the Tropical NW Pacific, South China Sea, and South Kuroshio (Fig. 2c,d). Xeniidae was the next most species-rich family but was not collected from the eastern Indo-Pacific regions (provinces 38–40). Observed richness of xeniid MOTUs was highest in the Western Indian Ocean, NE Australian Shelf, Western Coral Triangle, and South Kuroshio. Cladiellidae was sampled in all provinces except Hawaii and the Marshall Islands, but overall richness was lower than in Sarcophytidae and Xeniidae. The largest numbers of MOTUs of Cladiellidae were found in the Western Indian Ocean, NE Australian Shelf, South China Sea, South Kuroshio and Warm Temperate NW Pacific (Fig. 2c,d). Other clades (Capnellidae, Lemnaliidae, Sinulariidae, Litophyton) were not found in any of the four easternmost provinces sampled (provinces 37–40). Lemnaliidae and Litophyton were both most species-rich in the Western Indian Ocean and NE Australian Shelf but were either absent from or not diverse in most other provinces. Capnellidae and Sinulariidae were the least diverse families, with just one or no MOTUs found in most provinces. The highest numbers recorded for these taxa were six MOTUs of Capnellidae in the NE Australian Shelf and five MOTUs of Sinulariidae in the Arabian Sea (province 19). Neither of those families were sampled from the Red Sea.
The distribution of soft coral diversity across the Indo-Pacific differed strongly between clades that are known to brood their planula-larvae and those that broadcast spawn56, with the latter extending into more remote oceanic areas. Species richness of brooders (standardized by R/E) was highest in the Western Indian Ocean, NE Australian Shelf and Western Coral Triangle (Fig. 3d,i). Brooders were absent (or not sampled) from most provinces in the eastern Indo-Pacific (38–40) with the exception of a xeniid that is endemic to Hawaii (Table 1). In addition, the species richness of brooding taxa was relatively low in the northern and eastern Indian Ocean including the Red Sea (Fig. 3d,i). In contrast, species richness of broadcast spawning taxa was relatively uniform throughout the Central Indo-Pacific provinces and Western Indian Ocean but with notable peaks in the South China Sea and Tropical NW Pacific (Fig. 3e,j). Broadcast spawners were found in every province, although their species richness declined eastward across the Indo-Pacific.
Endemism
Regional endemism of MOTUs was high, with 66.8% of MOTUs found in just one province and 45.3% of MOTUs represented by a single sequence (Fig. 4). Excluding singletons and considering only those MOTUs that were sampled more than once, 39.3% were found in just one province. Endemism varied greatly among clades, ranging from 55% in Sarcophytidae to 89% in Litophyton (MOTUs delineated with 28S rDNA), and was generally higher in brooding taxa (Xeniidae, Lemnaliidae, Capnellidae and Litophyton) than in broadcast-spawning families (Fig. 5). Among all clades, only 9.6% of MOTUs were found in more than five provinces (Fig. 4).

Frequency distribution of number of marine provinces in which MOTUs of zooxanthellate soft corals were found. MOTUs defined using (a) 28S rDNA or (b) mtMutS barcodes. Families are indicated by color. No mtMutS sequences were obtained for Cladiellidae.

Percent of MOTUs in a family that were endemic (green: sampled in only one province); non-endemic (dark blue: sampled in 2–9 provinces); or widespread (light blue: sampled in 10 or more provinces). MOTUs delineated by (a) 28S rDNA or (b) mtMutS barcode. No mtMutS sequences were obtained for Cladiellidae. Numbers = total number of MOTUs per family.
The proportions of MOTUs found within a province that were endemic to that province were 0–50% (Figs. 3b,g, 6; Table 1). Endemism was highest in the Western Indian Ocean (45.5%), NE Australian Shelf (40.9%), Red Sea (40.7%) and Hawaii (50%) (Fig. 3b,g). Percentages of endemic species were lowest in the Tropical SW Pacific, SE Polynesia, Sunda Shelf and Java Transitional. Estimates of endemism based on MOTUs delineated using mtMutS were in general lower than those delineated with 28S (Figs. 5, 6; Table 1). In contrast to 28S, with mtMutS no endemic taxa were recorded in the Sunda Shelf, Hawaii, the Marshall Islands, or SE Polynesia, and only one at Lord Howe Island. These provinces with no recorded mtMutS endemics include four of the six easternmost provinces sampled.

Percent of MOTUs in a province that were endemic (green: sampled in only one province); non-endemic (dark blue: sampled in 2–9 provinces); or widespread (light blue: sampled in 10 or more provinces). MOTUs delineated by (a) 28S rDNA or (b) mtMutS barcode. No mtMutS sequences were obtained for Cladiellidae.
Only 16 of 471 MOTUs (3.4%) were found in 10 or more of the 20 provinces surveyed (18 of 295 (6.1%) based on mtMutS) (Fig. 4). Ten of these 16 most geographically widespread MOTUs belonged to Sarcophytidae, three to Cladiellidae, two to Sinulariidae and one to Xeniidae. Fifteen had distributions spanning the Western Indian Ocean to the eastern Indo-Pacific; one widespread Sclerophytum MOTU was absent from the Western Indian Ocean. As a percentage of total MOTUs, these geographically widespread taxa accounted for 14–30% of the richness in most provinces (Fig. 6). They were, however, overrepresented in the easternmost provinces (37–40) as well as in the Warm Temperate NW Pacific and Sunda Shelf, where they accounted for 35–80% of MOTUs.
Community composition
Betadiversity indices reflected the high levels of regional endemism of MOTUs. Baselga’s multiple-site dissimilarity index57 for all taxa (28S MOTUs) was βSOR = 0.931 with βSIM = 0.859 suggesting that most of the dissimilarity in species composition among provinces was due to turnover rather than nestedness (βSNE = 0.072) (Supplementary Table S4). Among the easternmost provinces (37–40), however, the nestedness component of betadiversity was high: 19 of the 20 highest pairwise comparisons (top 10%, βSNE > 0.48) involved those four provinces (Supplementary Table S5). Conversely, 15 of the 20 lowest pairwise values of the turnover component of betadiversity (bottom 10%, βSIM < 0.35) were observed among provinces 37–40. These trends support the observations that MOTUs with wide geographic distributions were overrepresented and endemics rare in the relatively species-depauperate oceanic provinces in the eastern Indo-Pacific (Fig. 6). Among families, multiple-site values of βSOR and βSIM calculated for MOTUs delineated by 28S were highest for Xeniidae (βSOR = 0.963, βSIM = 0.911) followed by Sarcophytidae (βSOR = 0.910, βSIM = 0.804), Lemnaliidae (βSOR = 0.905, βSIM = 0.828) and Cladiellidae (βSOR = 0.896, βSIM = 0.813) (Supplementary Table S4). While pairwise betadiversity values for MOTUs delineated using mtMutS were generally lower than those found with 28S, the same trends and patterns were observed (Supplementary Table S5). Among families, however, multiple-site βSOR was higher for Lemnaliidae (βSOR = 0.955, βSIM = 0.779) than for Xeniidae (βSOR = 0.945, βSIM = 0.845) and Sarcophytidae (βSOR = 0.900, βSIM = 0.783), and nestedness (βSNE) values were higher for all taxa (Supplementary Table S4).
MDS plots of dissimilarity (βSOR) suggested that community composition of MOTUs was similar among most provinces with the notable exceptions of the easternmost provinces (37–40), which formed a distinct cluster in both 28S and mtMutS plots (70% dissimilarity, Fig. 7), and Lord Howe Is. (36), which was distinct from all other provinces. In both analyses, provinces in the Western and NW Indian Ocean (18–20) were also somewhat distinct from those in the Eastern Indian Ocean and central Indo-Pacific.

MDS plot of compositional betadiversity (βSOR) of zooxanthellate soft corals among marine provinces. MOTUs delineated with (a) 28S rDNA or (b) mtMutS. Solid lines: 80% dissimilarity clusters; dashed lines: 70% dissimilarity clusters. Red, western Indo-Pacific; blue, central Indo-Pacific; pink, eastern Indo-Pacific; yellow, Warm Temperate NW Pacific.
Discussion
The patterns of biodiversity we document here for genetic lineages (proxies of species) of zooxanthellate soft corals differed from the two most studied and emblematic groups of Indo-Pacific reef-dwelling organisms, the reef-building scleractinian corals and coral reef-associated fish. In both of those groups, as well as some other mobile macroinvertebrates14,15, species richness is highest in the Indo-Australian Archipelago region of the central Indo-Pacific and decreases longitudinally to both the east and west4,6,8,9,11,12. Centers of endemism for these taxa are not concordant with centers of species richness, with most studies suggesting that the highest levels of endemism for both scleractinians and reef fish occur in peripheral areas of relatively lower species richness such as the SW Indian Ocean, Red Sea, Hawaii, and the easternmost regions of the Indo-Pacific (e.g. Marquesas, Easter I.)1,7,10,12,13,66,67.
In contrast to these well-documented biogeographic patterns—but concordant with some recent studies of scleractinians68,69—we find multiple centers of diversity for reef-dwelling zooxanthellate soft corals, with both richness and phylogenetic diversity of MOTUs in the Western Indian Ocean (WIO) similar to that of the Western Coral Triangle and NE Australian Shelf. Moreover, centers of endemism for soft corals are coincident with these areas of high species richness, in particular in the WIO. With the exception of Hawaii, peripheral areas with low species richness such as the oceanic islands of the eastern Indo-Pacific have proportionately fewer endemic lineages and an overrepresentation of lineages with pan-Indo-Pacific geographic ranges. As in scleractinians, reef fish and some mobile macroinvertebrates67, the marginal Red Sea also supports a very high percentage of endemic lineages of soft corals despite its lower species richness.
The only previous study of the biogeographic distribution of Indo-Pacific octocorals focused on the speciose genus Sclerophytum (formerly Sinularia; family Sarcophytidae) and estimated species’ ranges based on records of species names from the literature derived from classical taxonomic identification methods70. That study concluded that species richness of Sclerophytum was highest in the WIO and Red Sea but warned that the pattern could have been an artifact of the relatively greater number of studies that had been conducted in those regions at the time. Indeed, our results indicate that species richness of Sarcophytidae peaks in the Tropical NW Pacific and is lower in the WIO and Red Sea. The species richness of zooxanthellate soft corals as a group is, however, very high in the WIO. That pattern is not driven by the species richness of Sclerophytum, but rather by the high diversity of families found in the WIO and by the high species richness of some of those families (e.g., Lemnaliidae, Xeniidae) that are absent from or not diverse in the easternmost Indo-Pacific provinces.
Sampling bias is also very likely to have influenced the patterns documented here69. Estimates of sample coverage (a measure of how likely it is that all species have been found53) were highest in the WIO, NW Pacific and NE Australia, where multiple octocoral-specific biodiversity surveys have been carried out by taxonomic specialists. Sample coverage was lower for other areas, in particular the Eastern Coral Triangle and all provinces in the oceanic eastern Indo-Pacific (37–40). Most of the DNA barcode data for these geographic areas was obtained from specimens in historical museum collections, often collected by non-specialists during general biodiversity surveys. In addition to lower sampling effort for soft corals in these regions, sequencing success rate was often much lower for older material. Combined, those factors contributed to smaller sample sizes and lower sample coverage estimates. Increased sampling of the Eastern Coral Triangle province in particular will likely increase species richness estimates for the Indo-Australian Archipelago region relative to the well-sampled WIO. Extensive sampling in the central Indian Ocean and Indian subcontinent—regions for which we lack any genetic data—are also necessary to confirm the possible faunal break between the WIO and central Indo-Pacific provinces. Despite these limitations, we have now documented for the first time biogeographical patterns that will facilitate further exploration of the drivers of diversity in this ecologically important group of foundation species.
The observed distribution of soft coral lineages suggests a strong role for larval dispersal in their biogeography. As far as is known, all species of the family Xeniidae brood larvae, a mode of reproduction that is generally assumed to result in lower dispersal capability56. The very few species of Lemnaliidae, Capnellidae and Litophyton whose reproductive biology has been studied are also internal or external brooders56. The planula larvae released by brooding octocorals typically have no pelagic phase and settle to the substrate immediately after release from the maternal colony71. The near absence of these presumably brooding taxa from the eastern Indo-Pacific provinces may be explained by the inability of their benthic larvae to disperse across large expanses of open ocean, limiting their ability to colonize remote islands. In contrast, all members of Sarcophytidae, Cladiellidae and Sinulariidae that have been studied so far are broadcast spawners that release eggs and sperm into the water column, necessitating a pelagic embryonic stage with a greater potential for dispersal56. The diversity of these broadcast spawning taxa peaks not in the SW Indian Ocean and Coral Triangle, but instead in the South China Sea and oceanic islands of Micronesia. These families were also found in the remote provinces of the eastern Indo-Pacific; moreover, the taxa sampled from those provinces disproportionately belonged to lineages documented to have pan-Indo-Pacific geographic ranges.
Despite what would appear to be similar larval stages and dispersal capabilities, species richness of zooxanthellate soft corals declines much more steeply across the eastern Indo-Pacific than it does for scleractinians56. While numerous taxa of scleractinians have crossed the Pacific, as evidenced by their occurrence in the Eastern Tropical Pacific, no zooxanthellate soft corals have been documented to occur east of SE Polynesia (Pitcairn Is., C. McFadden, pers. obs.). As a group, zooxanthellate soft corals are also absent from the tropical Atlantic Ocean, with the exception of species that have been introduced recently via anthropogenic activities72,73. Some taxa of scleractinians, including brooders, exhibit long pelagic larval duration (PLD) that may facilitate long-distance dispersal74. We currently lack explicit information on PLD for any octocorals to support the hypothesis that their larval dispersal capabilities are inherently lower than those of scleractinians. We have also not yet explored the role of other environmental drivers (i.e. environmental filtering) or geological history that might contribute to the distribution of biodiversity in zooxanthellate soft corals in Indo-Pacific coral reef ecosystems9.
Limitations of a single-locus genetic approach to species delimitation
The single-locus molecular markers we have used to delineate lineages (MOTUs) have known limitations that may introduce biases into the patterns of species richness and endemism documented here. The mitochondrial genomes of most anthozoan corals evolve very slowly relative to other metazoans75, with the result that not all species of octocorals can be discriminated reliably using mitochondrial markers such as mtMutS40,49. Although more reliable for species discrimination in some taxa, 28S rDNA shares many of the same limitations41,49. In Cladiellidae and Nephtheidae in particular, rates of evolution of mitochondrial genes appear to be very slow, and there is little sequence differentiation among most species65,76. In other families, however, there is generally good congruence between the numbers of MOTUs estimated using mtMutS and the nuclear 28S rDNA49. Previous publications have shown that both of these barcode markers recover relationships among these focal genera and families that are congruent with phylogenomic analyses41,44.
As a result of the limitations of these molecular markers, estimated numbers of MOTUs are likely to be conservative and true species richness in some families may be higher than these analyses suggest. For example, neither mtMutS nor 28S rDNA alone discriminate all species of Ovabunda in the Red Sea40 or Sclerophytum in Dongsha Atoll, South China Sea41. We have undoubtedly underestimated numbers of species for these taxa. If recently diverged species with non-overlapping ranges have inadvertently been lumped into a single MOTU in our analyses, geographic ranges of species will also have been overestimated and endemism underestimated21,23. We consider this possibility more likely than the over-splitting of species (and consequent underestimation of geographic ranges and overestimation of endemism). The limitations of the markers may also bias or obscure the geographic patterns of endemism documented in the current study. We recorded few endemic lineages in peripheral provinces, but if species have colonized those regions and diverged only recently, these slowly evolving markers may not yet distinguish them from ancestral populations. Despite their limitations, the use of single-locus molecular markers to support and revise morphospecies concepts in octocorals has greatly improved our understanding of species boundaries40,41,45,46,47 and species ranges48. Use of more sensitive genomic approaches to delimit species23,41, combined with detailed morphological studies, will likely lead to further refinement of those species boundaries and the biogeographical patterns documented in this study.
Conclusions and future work
Studies of scleractinian corals and other taxa with rich fossil records suggest that contemporary distributions of biodiversity have largely been driven by historical geological processes rather than by environmental variables or taxon-specific traits9,68,77. While most studies have focused on the origin and geological drivers of the present-day peak of biodiversity in the tectonically active Indo-Australian Archipelago78, there is growing recognition of the importance of the Western Indian Ocean as a second peak of marine biodiversity, driven in part by the survival of relictual Tethyan lineages in a geologically stable environment68,69,77. The patterns we document for zooxanthellate soft corals support the WIO as a center of biodiversity and endemism that may be commensurate with the IAA. While the lack of a fossil record for soft corals presents challenges to reconstructing historical patterns of biodiversity, additional sampling of these understudied taxa and regions of the ocean coupled with integrative approaches to species delimitation will continue to further our understanding of the distribution of biodiversity in the Indo-Pacific and yield new insights into the history and drivers of diversification.
Data availability
Sequence data that support the findings of this study have been deposited in GenBank. All accession numbers are listed in Supplementary Table S2. Previously unpublished 28S rDNA sequences have been deposited as a Targeted Locus Study (TLS), BioProject PRJNA1171007, accession KIWD01000000.
References
Huang, D., Goldberg, E. E., Chou, L. M. & Roy, K. The origin and evolution of coral species richness in a marine biodiversity hotspot. Evolution 72, 288–302 (2018).
Danovaro, R. Understanding marine biodiversity patterns and drivers: The fall of Icarus. Mar. Ecol. https://doi.org/10.1111/maec.12814 (2024).
Worm, B. & Lotze, H. K. Marine biodiversity and climate change. In Climate Change: Observed Impacts on Planet Earth, 3rd ed. (ed Letcher, T. M.) 445–464 (Elsevier, 2021). https://doi.org/10.1016/B978-0-12-821575-3.00021-9.
Roberts, C. M. et al. Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 295, 1280–1284 (2002).
Selig, E. R. et al. Global priorities for marine biodiversity conservation. PLoS ONE 9, e82898 (2014).
Rogers, A. D. et al. Critical habitats and biodiversity: Inventory, thresholds and governance. In The Blue Compendium: From Knowledge to Action for a Sustainable Ocean Economy (eds Lubchenco, J. & Haugan, P. M.) 333–391 (Springer, 2020).
Hughes, T. P., Bellwood, D. R. & Connelly, S. R. Biodiversity hotspots, centres of endemicity, and the conservation of coral reefs. Ecol. Lett. 5, 775–784 (2002).
Hoeksema, B. W. Delineation of the Indo-Malayan centre of maximum marine biodiversity: The Coral Triangle. In Biogeography, Time, and Place: Distributions, Barriers, and Islands (ed. Renema, Willem) 117–178 (Springer Netherlands, Dordrecht, 2007). https://doi.org/10.1007/978-1-4020-6374-9_5.
Keith, S. A., Baird, A. H., Hughes, T. P., Madin, J. S. & Connolly, S. R. Faunal breaks and species composition of Indo-Pacific corals: The role of plate tectonics, environment and habitat distribution. Proc. R. Soc. B 280, 20130818 (2013).
Allen, G. R. Conservation hotspots of biodiversity and endemism for Indo-Pacific coral reef fishes. Aquat. Conserv. 18, 541–556 (2008).
Kulbicki, M. et al. Global biogeography of reef fishes: A hierarchical quantitative delineation of regions. PLoS ONE 8, e81847 (2013).
Cowman, P. F. Historical factors that have shaped the evolution of tropical reef fishes: A review of phylogenies, biogeography, and remaining questions. Front. Genet. 5, 394 (2014).
Cowman, P. F., Parravicini, V., Kulbicki, M. & Floeter, S. R. The biogeography of tropical reef fishes: Endemism and provinciality through time. Biol. Rev. 92, 2112–2130 (2017).
Bribiesca-Contraras, G., Verbruggen, H., Hugall, A. F. & O’Hara, T. D. Global biogeographic structuring of tropical shallow-water brittle stars. J. Biogeogr. 46, 1287–1299 (2019).
Rosa, R. et al. Global patterns of species richness in coastal cephalopods. Front. Mar. Sci. 6, 469 (2019).
Kerswell, A. P. Global biodiversity patterns of benthic marine algae. Ecology 87, 2479–2488 (2006).
Yip, Z. T., Quek, R. Z. B. & Huang, D. Historical biogeography of the widespread macroalga Sargassum (Fucales, Phaeophyceae). J. Phycol. 56, 300–309 (2020).
Hebert, P. D. N., Cywinska, A., Ball, S. L. & de Waard, J. R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B 270, 313–321 (2003).
Fukami, H. Regional specific approach is a next step for setting evolutionary-based conservation priorities in the scleractinian corals. Anim. Cons. 18, 318–319 (2015).
DeQuieroz, K. Species concepts and species delimitation. Syst. Biol. 56, 879–886 (2007).
Paulay, G. & Meyer, C. Dispersal and divergence across the greatest ocean region: Do larvae matter? Integr. Comp. Biol. 46, 269–281 (2006).
Keshavmurthy, S. et al. DNA barcoding reveals the coral “laboratory rat”, Stylophora pistillata, encompasses multiple identities. Sci. Rep. 3, 1520 (2013).
Bridge, T. C. L. et al. A tenuis relationship: Traditional taxonomy obscures systematics and biogeography of the ‘Acropora tenuis’ (Scleractinia: Acroporidae) species complex. Zool. J. Linn. Soc. https://doi.org/10.1093/zoolinnean/zlad187 (2023).
OBIS Ocean Biodiversity Information System. Intergovernmental Oceanographic Commission of UNESCO. https://obis.org. (2024)
Goulet, T. L., Simmons, C. & Goulet, D. Worldwide biogeography of Symbiodinium in tropical octocorals. Mar. Ecol. Prog. Ser. 355, 45–58 (2008).
Liberman, R., Benayahu, Y. & Huchon, D. Octocorals in the Gulf of Aqaba exhibit high photosymbiont fidelity. Front. Microbiol. 13, 1005471. https://doi.org/10.3389/fmicb.2022.1005471 (2022).
Goulet, T. L., LaJeunesse, T. C. & Fabricius, K. E. Symbiont specificity and bleaching susceptibility among soft corals in the 1998 Great Barrier Reef mass coral bleaching event. Mar. Biol. 154, 795–804 (2008).
Coffroth, M. A. et al. What makes a winner? Symbiont and host dynamics determine Caribbean octocoral resilience to bleaching. Sci. Adv. 9, eadj6788 (2023).
Rodriguez, M. V. B., Segumalian, C. S., Lalas, J. A. A. & Maningas, J. M. C. Octocorals outcompete scleractinian corals in a degraded reef. IOP Conf. Ser. Earth Environ. Sci. 420, 012027 (2019).
Garra, S., Hall, A. & Kingsford, M. J. The effects of predation on the condition of soft corals. Coral Reefs 39, 1329–1343 (2020).
Tsounis, G., Steele, M. A. & Edmunds, P. J. Elevated feeding rates of fishes within octocoral canopies on Caribbean reefs. Coral Reefs 39, 1299–1311 (2020).
Lasker, H. R., Bramanti, L., Tsounis, G. & Edmunds, P. J. The rise of octocoral forests on Caribbean reefs. Adv. Mar. Biol. 87, 361–410 (2020).
Reimer, J. D. et al. A large Heliopora-dominated reef off the east coast of Okinawajima Island Japan. Fauna Ryukyuana 69, 19–23 (2024).
Jeng, M.-S., Huang, H.-D., Dai, C.-F., Hsiao, Y.-C. & Benayahu, Y. Sclerite calcification and reef-building in the fleshy octocoral genus Sinularia (Octocorallia: Alcyonacea). Coral Reefs 30, 925–933 (2011).
Shoham, E., Prohaska, T., Barkay, Z., Zitek, A. & Benayahu, Y. Soft corals form aragonite-precipitated columnar spiculite in mesophotic reefs. Sci. Rep. 9, 1241. https://doi.org/10.1038/s41598-018-37696-z (2019).
Reverter, M., Helber, S. B., Rohde, S., de Goeij, J. M. & Schupp, P. J. Coral reef benthic community changes in the Anthropocene: Biogeographic heterogeneity, overlooked configurations, and methodology. Global Change Biol. 28, 1956–1971 (2021).
Fabricius, K. Reef invasion by soft corals: Which taxa and which habitats? in Proceedings of the Australian Coral Reef Society 75th Anniversary Conference, Heron Island, October, 1997 (eds. Greenwood, J. G., & Hall, N. J.) 78–90 (University of Queensland, 1998).
Lalas, J. A. A., Jamodiong, E. A. & Reimer, J. D. Spatial patterns of soft coral (Octocorallia) assemblages in the shallow coral reefs of Okinawa Island, Ryukyu Archipelago, Japan: Dominance on highly disturbed reefs. Reg. Stud. Mar. Sci. 71, 103405. https://doi.org/10.1016/j.rsma.2024.103405 (2024).
Fabricius, K. & Alderslade, P. Soft Corals and Sea Fans: A Comprehensive Guide to the Tropical Shallow-Water Genera of the Central West-Pacific, the Indian Ocean and the Red Sea (Australian Institute of Marine Science, 2001).
McFadden, C. S. et al. Species boundaries in the absence of morphological, ecological or geographical differentiation in the Red Sea octocoral genus Ovabunda (Alcyonacea: Xeniidae). Mol. Phylogen. Evol. 112, 174–184 (2017).
Quattrini, A. M. et al. A next generation approach to species delimitation reveals the role of hybridization in a cryptic species complex of corals. BMC Evol. Biol. 19, 116 (2019).
West, J. M., Harvell, C. D. & Walls, A.-M. Morphological plasticity in a gorgonian coral (Briareum asbestinum) over a depth cline. Mar. Ecol. Prog. Ser. 94, 61–69 (1993).
Halász, A., McFadden, C. S., Toonen, R. J. & Benayahu, Y. Re-description of type material of Xenia Lamarck, 1816. Zootaxa 4652, 201–239 (2019).
McFadden, C. S., van Ofwegen, L. P. & Quattrini, A. M. Revisionary systematics of Octocorallia (Cnidaria: Anthozoa) guided by phylogenomics. Bull. Soc. Syst. Biol. 1(8735), 1–79 (2022).
van Ofwegen, L. P., Benayahu, Y. & McFadden, C. S. Sinularia leptoclados (Ehrenberg, 1834) (Cnidaria: Octocorallia) re-examined. ZooKeys 272, 29–59 (2013).
van Ofwegen, L. P., McFadden, C. S. & Benayahu, Y. Sinularia polydactyla (Ehrenberg, 1834) (Cnidaria, Octocorallia) re-examined, with description of a new species. ZooKeys 581, 71–126 (2016).
Benayahu, Y., Ekins, M. & McFadden, C. S. Overview of the genus Sympodium Ehrenberg, 1834 (Octocorallia, Alcyonacea, Xeniidae), with the description of new species, revealing regional endemism. Zootaxa 5072, 324–350 (2021).
McFadden, C. S. et al. Molecular operational taxonomic units reveal regional endemism in the Indo-Pacific octocoral family Xeniidae. J. Biogeogr. 46, 992–1006 (2019).
McFadden, C. S., Brown, A. S., Brayton, C., Hunt, C. B. & van Ofwegen, L. P. Application of DNA barcoding to biodiversity studies of shallow-water octocorals: Molecular proxies agree with morphological estimates of species richness in Palau. Coral Reefs 33, 275–286 (2014).
Katoh, K., Kuma, K., Toh, H. & Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucl. Acids Res. 33, 511–513 (2005).
Schloss, P. D. et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541 (2009).
Spalding, M. D. et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. Bioscience 57, 573–583 (2007).
Chao, A. & Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 93, 2533–2547 (2012).
Chao, A. et al. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 84, 45–67 (2014).
Chao, A., Ma, K. H. & Hsieh, T. C. iNEXT Online: Software for Interpolation and Extrapolation of Species Diversity. Program and User’s Guide http://chao.stat.nthu.edu.tw/wordpress/software_download/inext-online/ (2016).
Kahng, S. E., Benayahu, Y. & Lasker, H. R. Sexual reproduction in octocorals. Mar. Ecol. Prog. Ser. 443, 265–283 (2011).
Helmus, M. R., Bland, T. J., Williams, C. K. & Ives, A. R. Phylogenetic measures of biodiversity. Am. Nat. 169, E68–E83 (2007).
R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/ (2024).
Kembel, S. An introduction to the picante package. (2010)
Minh, B. Q. et al. IQTree-2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 37, 1530–1534 (2020).
Chernomor, O., von Haeseler, A. & Minh, B. Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 65, 997–1008 (2016).
Kalyaanamoorthy, S., Minh, B. Q., Wong, T. K. F., von Haeseler, A. & Jermiin, L. S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 14, 587–589 (2017).
Baselga, A. The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Global Ecol. Biogeogr. 21, 1223–1232 (2012).
Clarke, K. R., & Gorley, R. N. PRIMER v7: User manual/tutorial. (PRIMER-E-LTD, 2015).
Benayahu, Y. et al. Diversity and distribution of octocorals (Coelenterata: Anthozoa) on the coral reefs of Penghu Taiwan. Zool. Stud. 51, 1529–1548 (2012).
DiBattista, J. D. et al. On the origin of endemic species in the Red Sea. J. Biogeogr. 43, 13–30 (2016).
DiBattista, J. D. et al. A review of contemporary patterns of endemism for shallow water reef fauna in the Red Sea. J. Biogeogr. 43, 423–439 (2016).
Obura, D. An Indian Ocean centre of origin revisited: Palaeogene and Neogene influences defining a biogeographic realm. J. Biogeogr. 43, 229–242 (2016).
Kusumoto, B. et al. Global distribution of coral diversity: Biodiversity knowledge gradients related to spatial resolution. Ecol. Res. 35, 315–326 (2020).
van Ofwegen, L. P. Status of knowledge of the Indo-Pacific soft coral genus Sinularia May, 1898 (Anthozoa: Octocorallia). In Proceedings of the Ninth International Coral Reef Symposium vol 1, pp 167–171 (2002).
Benayahu, Y. & Loya, Y. Substratum preferences and planulae settling of two Red Sea alcyonaceans: Xenia macrospiculata Gohar and Parerythropodium fulvum fulvum (Forskål). J. Exp. Mar. Biol. Ecol. 83, 249–261 (1984).
Mantelatto, M. C., da Silva, A. G., Louzada, T. S., McFadden, C. S. & Creed, J. C. Invasion of aquarium origin soft corals on a tropical rocky reef in the southwest Atlantic, Brazil. Mar. Pollut. Bull. 130, 84–94 (2018).
Ruiz Allais, J. P., Benayahu, Y. & Lasso-Alcala, O. M. The invasive Unomia stolonifera (Alcyonacea, Xeniidae) is dominating the benthos in the Southeastern Caribbean Sea. Mem. Fund. La Salle Cienc. Nat. 79, 63–80 (2021).
Harrison, P. L. & Wallace, C. C. Reproduction, dispersal and recruitment of scleractinian corals. In Ecosystems of the World 25: Coral Reefs (ed. Dubinsky, Z.) 133–207 (Elsevier, 2011).
Shearer, T. L. & Coffroth, M. A. Barcoding corals: Limited by interspecific divergence not intraspecific variation. Mol. Ecol. Res. 8, 247–255 (2008).
Cordeiro, R. T. S. et al. Neospongodes atlantica, a potential case of an early biological introduction in the Southwestern Atlantic. PeerJ 10, e14347. https://doi.org/10.7717/peerj.14347 (2022).
Yasuhara, M. et al. Hotspots of Cenozoic tropical marine biodiversity. Oceanogr. Mar. Biol. Ann. Rev. 60, 243–300 (2022).
Renema, W. et al. Hopping hotspots: Global shifts in marine biodiversity. Science 321, 654–657 (2008).
Acknowledgements
We thank the collections staff at the California Academy of Sciences for access to specimens collected by the Coral Reef Research Foundation under contracts to the U.S. National Cancer Institute; Dr. Zafrir Kuplik (SMNHTAU) for curatorial assistance; and E. Hou, L. West, B. Bell, and J. Najera for laboratory assistance. We also thank the many people and organizations who have supported field collections over the years, including: Dr. S. Arakaki (Amakusa Station, Kyushu University) and Dr. Y.W. Lau (U. Ryukyus) for field survey help in S. Kuroshio and Okinawa; E. Tessier, director “Parc marin de La Réunion”, C. Bourmaud, J. P. Quod and M. Aknin for support of collections in La Réunion and Madagascar; Ministry of Livestock and Fisheries Development, Fisheries Department, Republic of Kenya for issuing collecting permits; Ming-Shiou Jeng (BRCAS, Academia Sinica), Chang-Feng Dai (National Taiwan University) and Keryea Song (National Sun Yat-sen University, Kaohsiung) for enthusiastic support of collections in Taiwan; Mr. W.S. Tsai, director of the Penghu Marine Biology Research Center, Fisheries Research Institute Penghu, for providing facilities and support in Penghu; staff members of Dongsha Atoll National Park, Dongsha Atoll Research Station (DARS), and Biodiversity Research Center, Academia Sinica (BRCAS) for support of collections on Dongsha Atoll; staff at the Inter University Institute for Marine Sciences in Eilat (IUI) for their support throughout the study period; the Israel Nature and National Parks Protection Authority for issuing collection permits; The Kimberley Woodside Collection Project; The Census of Marine Life (Coral Reef Program), CREEFs; M. Claereboudt, S. Dobertsov (Sultan Qaboos University), S. Wilson, and O. Taylor (Five Oceans Environmental Services LLC) for their support of collections in Oman; the Environment Authority of Oman for granting collection permits; The Richard Lounsbery Foundation, J.H. Ausubel (Rockefeller University) and L. Brown (Lounsbery Foundation) for support of research at Naturalis and their continued support and encouragement to K.S.-N; collaborators at Vamizi Lodge, AndBeyond, the Environmental Research and Conservation Center of Lúrio University, and the Wildlife Conservation Society (WCS) for assistance with collections in Mozambique; and R.M.A. Luciano and C.S. Segumalian of the Marine Science Institute of the University of the Philippines-Diliman for collecting and preparing part of the collections from the West Philippine Sea. The expeditions in the Philippines were also made possible with the logistical support provided by the Philippine Navy, Philippine Coast Guard, and the Department of Agriculture-Bureau of Fisheries and Aquatic Resources. Collection and export permits were obtained from the relevant agencies and jurisdictions for all material collected and are available from the authors upon request.
Funding
Funding for this research was provided by US National Science Foundation awards DEB-1929319 (C.S.M., A.M.Q.) and Binational Science Foundation BSF-2019624 (Y.B.). Collections in Oman were funded by NSF DEB-1856245 (G.P., C.S.M.). Collections in S. Kuroshio and Okinawa were funded by a TBRC ORCHIDS grant to C.S.M. and J.D.R. Collections in Reunion Is. were made possible by grants to Y.B. from the “Conseil régional de La Réunion” and “Association Parc marin de La Réunion”. Collections in Kenya and Tanzania were funded by PharmaMar, Spain. Collection trips to Penghu Archipelago were supported by a grant to Y.B. from the National Science Council of Taiwan. Collection trips to Dongsha Atoll, Taiwan were made possible by a grant to Y.B. from the Taiwanese Ministry of Science (MOST), and collections in Green Is., Kenting National Park and northern Taiwan were supported by grants to Y.B. from the National Science Council, R.O.C.; Taiwanese Ministry of Science (MOST); and the Biodiversity Research Center, Academia Sinica (BRCAS). The Dalio Foundation (Singapore) Limited, GSK-EDB Trust Fund, HSBC, Shell Eastern Trading (Pte) Ltd and ExxonMobil Asia Pacific generously contributed in support of the Singapore Comprehensive Marine Biodiversity Survey II. Collections in the Philippines were funded by DENR-Biodiversity Management Bureau (DENR-BMB), National Security Council (NSC) and UP Office of the Vice President for Academic Affairs-Enhanced Creative Work Grant (ECWRG) to M.V.B.-R. Work in Saudi Arabia was supported by KAUST baseline funds to F.B. Australian collections by M.E. were made as part of the Census of Marine Life (CReefs Program) funded by the Australian Government under the auspices of the Australian Institute of Marine Science, the Great Barrier Reef Research Foundation and BHP Billiton, and Woodside Energy, as a part of the Kimberley Woodside Collection Project (2008–2011). B.N. was supported by the EAF-Nansen Programme and the FAO Education Programme. R.P. was supported by a Rasmussen Summer Research award from the Hixon Center for Climate and the Environment, Harvey Mudd College.
Author information
Authors and Affiliations
Contributions
C.S.M. wrote the main manuscript text with contributions from A.M.Q., Y.B., G.P., D.H., F.B., J.D.R. and M.E. C.S.M., A.L., A.M.Q. and K.S-N. conducted analyses and prepared figures. C.S.M., B.N., J.A.A.L., L.M., Y.K., K.S-N., G.P., F.B., J.D.R., D.H., M.V.B-R., M.E. and Y.B. organized and participated in field collections. C.S.M., K.L.E., A.L., B.N., S.A., S.K.D., K.M.D., N.P.M., M.M-P., J.N., A.P., S.P., R.P., R.P-R., K.E.S. and T.W. conducted laboratory work. C.S.M., Y.B. and A.M.Q. conceived the study. All authors reviewed and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
McFadden, C.S., Erickson, K.L., Lane, A. et al. Biodiversity and biogeography of zooxanthellate soft corals across the Indo-Pacific. Sci Rep 15, 15461 (2025). https://doi.org/10.1038/s41598-025-98790-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-98790-7
Keywords
This article is cited by
-
Exploring the diversity and distribution of Mozambique’s soft corals (Octocorallia) through an integrative approach
Marine Biodiversity (2026)
-
The hidden diversity of Saudi Arabian Red Sea octocorals revealed through a morpho-molecular assessment across bathymetric and latitudinal gradients
Scientific Reports (2025)
