
Abstract
Mungbean (Vigna radiata (L.) R. Wilczek var. radiata) is a highly nutritious legume grown in diverse cropping systems. However, its productivity is often hindered by abiotic stresses, particularly drought, which significantly impacts yield during critical growth stages such as flowering and pod filling. With climate change projected to increase drought frequency, leguminous crops are expected to be more vulnerable, exacerbating food security concerns. This study aimed to identify drought-tolerant mungbean genotypes by evaluating 122 genotypes (116 minicore accessions and six check varieties) under well-watered (WW) and drought-stressed (DS) conditions in a semi-controlled pot culture setup. Drought tolerance was assessed using multiple stress indices (e.g., STI, GMP), principal component analysis (PCA), and the multi-trait genotype-ideotype distance index (MGIDI). Significant genetic variability was observed across both WW and DS conditions, with drought stress leading to reductions in grain yield (21.1%), leaf area (35.4%), canopy greenness (32.1%), and photosynthetic rate (51.8%). Traits such as plant height, number of branches, pods per plant, seeds per pod, transpiration rate, and grain yield showed high heritability and genetic advance, indicating strong breeding potential. PCA analysis identified major traits contributing to drought tolerance, including yield-related traits and physiological responses, such as transpiration rate, stomatal conductance, and photosynthetic efficiency. MGIDI effectively identified 18 genotypes based on multi-trait performance. Among them, genotypes VI003685AG, VI002051BG, VI000852AG, VI002402BG, and VI003957AG consistently ranked as top performers across both traditional indices and MGIDI, demonstrating preliminary drought tolerance. The integration of stress indices, PCA, and MGIDI provided a comprehensive approach to genotype selection, offering potentially valuable genetic resources for future mungbean breeding programs targeting drought-prone environments. However, these findings are preliminary and require multi-location field validation before breeding use.
Similar content being viewed by others
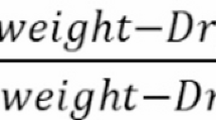

Introduction
Mungbean (Vigna radiata (L.) R. Wilczek var. radiata) is a vital pulse crop cultivated across India, Asia, Africa, South America, and Australia. It is valued not only for its high protein content (20–25%) and essential nutrients but also for its role in improving soil fertility through biological nitrogen fixation1,2. In India, mungbean ranks as the third most important legume crop after chickpea and pigeonpea, occupying nearly 5.51 million hectares and producing around 4 million tonnes annually, with an average yield of 570 kg/ha3. Being largely rainfed, mungbean is particularly vulnerable to drought, which is the most severe constraint limiting productivity, especially during flowering and pod-filling stages. Drought-induced yield losses can range from 51 to 85%, depending on genotype, stress intensity, and environmental conditions4,5,6. Climate change is expected to exacerbate these challenges by increasing the frequency and severity of drought events, threatening global food security and the achievement of Sustainable Development Goal 2: Zero Hunger7,8.
Although agronomic interventions such as mulching, conservation tillage, and plant growth-promoting rhizobacteria (PGPR) can mitigate drought effects, their impact is often limited, site-specific, and economically constrained9. Developing genetically drought-resilient cultivars offers a more sustainable and cost-effective solution. However, drought tolerance is a complex, polygenic trait influenced by genotype × environment interactions, stress intensity, and timing. Physiological and biochemical traits, such as canopy temperature, relative water content (RWC), stomatal conductance, and net CO2 assimilation, are commonly used to characterise genotypic responses under water-limited conditions6.
Further, several stress indices, including Tolerance Index (TOL), Stress Tolerance Index (STI), Yield Index (YI), Yield Stability Index (YSI), Stress Susceptibility Index (SSI), Mean Productivity (MP), and Geometric Mean Productivity (GMP), have been widely applied to evaluate drought effects on yield10,11,12. However, most studies focus on single traits or stress indices in isolation, which may fail to capture the complex, integrated responses of genotypes to drought and often yield inconsistent selection outcomes13,14. Multivariate approaches, such as principal component analysis (PCA) and the Multi-trait Genotype-Ideotype Distance Index (MGIDI), allow simultaneous evaluation of multiple traits, account for trait correlations, and improve selection accuracy15,16. Despite their potential, these approaches remain underutilised in mungbean breeding programs, and limited studies have explored the integration of physiological, yield, and stress indices to identify drought-resilient genotypes.
Against this background, we hypothesise that integrating morphophysiological traits with drought stress indices using multivariate approaches (PCA and MGIDI) can effectively identify mungbean genotypes with superior, balanced drought tolerance under controlled water-limited conditions, providing insights for targeted breeding. Accordingly, the present study had two broad objectives: (i) to evaluate the performance and drought resilience of diverse mungbean genotypes under well-watered and drought-stressed conditions by integrating morpho-physiological traits and stress indices, and (ii) to explore genetic variability and trait associations using multivariate analyses to identify promising genotypes for future breeding programs. By combining physiological measurements, drought indices, and advanced multivariate approaches, this study provides a comprehensive framework for assessing drought tolerance. However, it is important to note that these findings are preliminary and require multi-location field validation before breeding use. This approach addresses a critical knowledge gap in multi-trait selection in mungbean.
Materials and methods
Site description
The experiment was conducted in 2024 at a controlled-environment greenhouse facility (Allice Biotechnology®) of the ICAR–National Institute of Abiotic Stress Management (NIASM), Baramati, Maharashtra, India (18°09′30.62″ N, 74°30′03.08″ E). The semi-automated greenhouse maintained average daily conditions of 26 ± 2.5 °C temperature, 68.2 ± 10.8% relative humidity, and photosynthetically active radiation (PAR) ranging between 450 and 750 μmol m⁻2 s⁻1.
Plant materials
The plant materials used in this study consisted of a mungbean minicore collection developed by the World Vegetable Centre17 and six local and national check varieties (IPM-205-7, Vaibhav, NM-94, PAU-911, PDM-139, and EC-0398949). Among these, EC-0398949 was registered explicitly with NBPGR, India, for drought-adaptive traits. The original minicore collection comprised 296 genetically diverse genotypes representing broad agroecological and geographical origins. However, due to poor germination and susceptibility to powdery mildew, only 116 genotypes were deemed viable for drought screening. Including the six check varieties, a total of 122 accessions were selected for experimental trials and subsequent data analysis. Detailed information on the selected genotypes and check varieties is provided in Supplementary Table 1.
Preparation of pots, fertiliser application, and seed sowing
Mungbean genotypes were cultivated in plastic pots, each measuring 15 cm in height and 12 cm in diameter, with three drainage holes at the base to ensure adequate water percolation. Prior to potting, the soil was sterilised to eliminate soilborne pathogens and ensure uniform microbial conditions. Each pot was filled with 8 kg of a homogenised mixture of black soil and farmyard manure (FYM) at a volumetric ratio of 50:1 (V/V). Macronutrients, viz., nitrogen (N), phosphorus (P), and potassium (K), were applied at the recommended agronomic dose of 10:45:10 kg ha⁻1, respectively. The required NPK quantities were calculated based on the total soil weight per pot and uniformly incorporated into the soil matrix. After mixing, each pot was adjusted to precisely 8 kg to standardise soil volume and initial moisture content across all treatments. Each genotype was represented by six pots: three allocated to well-watered (WW) conditions and three to drought-stress (DS) treatment. Three healthy seeds were manually sown per pot at a depth of 15–18 mm. Following germination, thinning was performed to retain one vigorous seedling per pot, thereby minimising intra-specific competition. The experiment was laid out using a completely randomised block design (CRBD) with three biological replicates per treatment to ensure statistical robustness.
Soil moisture assessment and stress imposition
Soil field capacity (FC) and wilting point (WP) were determined gravimetrically, with the soil exhibiting an FC of 30–35%, a WP of 12–14%, and an available water range of 16–23%. To determine FC, finely ground, oven-dried soil was placed in perforated pots, saturated overnight by capillary action, drained to remove excess water, and weighed to calculate moisture content relative to dry weight. During the experiment, soil moisture in each pot was monitored daily using a precision digital balance, and the exact amount of water lost was restored to maintain the target moisture range for each treatment. Under well-watered (WW) conditions, moisture was kept at 28–31% (near FC), while drought-stress (DS) pots were maintained at 14–18% by supplying only the water required to keep moisture within this narrow range from bud initiation to physiological maturity. This gravimetric approach ensured consistent stress levels across replicates and minimised variation caused by differential evapotranspiration. Although pot experiments ensure uniform stress imposition, they inherently restrict root volume, which may lead to pot-bound stress that could interact with drought responses. While we used appropriately sized pots and ensured uniform seedling establishment to minimise these effects, this limitation remains when extrapolating the results to field conditions.
Trait measurement
Growth, morphological, and yield-related traits were assessed for all plants under WW and DS conditions in each replication. The measured traits included: Plant height (PH; cm), number of primary branches per plant (NPB), number of secondary branches per plant (NSB), number of pods per plant (PP), pod length (PL; cm), number of seeds per pod (NSP), test weight (TW; g) and grain yield per plant (GY; g),
Physiological parameters
Physiological parameters were assessed during peak stress periods to evaluate plant responses under well-watered and drought-stress conditions.
Leaf area (LA; cm2 per leaf) was measured non-destructively using a handheld leaf area meter (LI-3000C®, LI-COR, USA). Canopy greenness was quantified using the Normalised Difference Vegetation Index (NDVI), recorded with a GreenSeeker® handheld sensor (Trimble, USA) positioned 1.0 m above the plant canopy, following the method of18. NDVI was calculated as:
where NIR is near-infrared reflectance, and IR is infrared reflectance. Chlorophyll fluorescence (Qmax = Fv/Fm) was determined using a real-time chlorophyll fluorescence imaging system (FluorCam FC 1000-H®, Photon Systems Instruments) according to the protocol described by19, where Fv = Fm-F0. Additionally, gas exchange parameters including leaf temperature (Tleaf; °C), vapor pressure deficit (VPD; Pa/kPa), stomatal conductance (GH2O; mmol m⁻2 s⁻1), transpiration rate (E; mmol m⁻2 s⁻1), and net CO2 assimilation rate (A; µmol m⁻2 s⁻1)were measured using a portable gas exchange system (GFS-3000®, WALZ, Germany) between 09:00 and 11:30 AM, following the method described by20.
Data analysis
Descriptive statistical analyses were performed for all measured morpho-physiological traits under both well-watered (WW) and drought-stressed (DS) conditions to evaluate phenotypic variability among genotypes. Parameters such as mean, minimum, and maximum values were computed using R software (version 4.5.1). To visualise the distribution, symmetry, and spread of trait data under the two water regimes, violin plots were generated using PAST software version 4.0321.
Analysis of variance
The data collected on growth, yield, and physiological traits were first subjected to the Shapiro–Wilk test to assess normality and Levene’s test to evaluate homogeneity of variances. As the data met the assumptions of normal distribution and equal variance, no transformation was applied. A two-way analysis of variance (ANOVA) was conducted separately for each trait to determine the effects of genotype (G), water regime (W), and their interaction (G × W). The following linear model was used:
where Yijk is the observed value of the trait for the k-th replicate of the i-th genotype under the j-th water regime. μ: overall mean, \(Gi\) is the effect of the i-th genotype, \(Wj\) is the effect of the j-th water regime, \((G\times W)ij\) is the interaction effect between genotype and water regime, εijk is the random error term, εijk ~ N(0, σ2).
The significance of main and interaction effects was tested using F-tests. Where significant differences were observed, mean comparisons were performed using Tukey’s Honest Significant Difference (HSD) test at a 5% significance level. All statistical analyses were conducted in R (version 4.5.1) using the packages metan, lme4, and agricolae.
Genetic parameters, including the genotypic coefficient of variation (GCV; %), phenotypic coefficient of variation (PCV; %), heritability in the broad sense (h2bs; %), genetic advance (GA), and genetic advance as a percentage of the mean (GAM), were calculated following the methodology outlined by22.
Correlation studies: Pearson’s correlation coefficients were calculated for all morphophysiological traits under both WW and DS conditions to assess the strength and direction of trait associations. Correlation analysis was performed using R software version 4.5.123, and the resulting correlation matrices were visualised using a correlogram.
Principal Component Analysis (PCA): Principal Component Analysis (PCA) was performed to examine the multivariate structure of the trait data and identify key traits contributing to genotypic variation under each water regime. PCA was conducted on standardised trait values, and biplots were generated to visualise the relationships between genotypes and traits in the reduced dimensions of the first two principal components (PC1 and PC2).
Multi-trait Genotype–Ideotype Distance Index (MGIDI): To identify drought-tolerant genotypes based on multiple traits, the Multi-trait Genotype–Ideotype Distance Index (MGIDI) was computed following the method of24. This index integrates multiple traits by calculating the Euclidean distance between each genotype and an ideotype (ideal genotype). Traits were grouped based on their contributions to the principal components, and factor analysis was used to address multicollinearity. The ideotype was defined to reflect breeding goals: higher values were desirable for PH, NPB, NSB, PP, NSP, PL, TW, GY, A, E, GH2O, NDVI, LA, and Qmax, while lower values were preferred for VPD and Tleaf. All traits were standardised and direction-adjusted so that lower MGIDI scores indicated closer similarity to the ideotype and thus greater desirability. Analyses were conducted in R using the metan package. Selection differentials and contributions of individual traits to the MGIDI were also evaluated to determine key drivers of genotype performance under drought.
Stress tolerance indices and ranking of genotypes
Various stress tolerance indices were calculated based on yield performance under both WW and DS conditions, as detailed in Table 1. These indices were used to rank genotypes according to their relative tolerance to drought stress.
Ranking of genotypes
To identify the most drought-tolerant genotypes, the rank sum (RS) was calculated across all stress indices. Genotypes were ranked from 1 to 122 based on their grain yield under both non-stress (Yp) and stress (Ys) conditions, with the highest-yielding genotypes receiving rank 1. Stress tolerance indices, including the Stress Tolerance Index (STI), Yield Stability Index (YSI), Geometric Mean Productivity (GMP), Mean Productivity (MP), and Yield Index (YI), which are positively correlated with drought tolerance, were ranked in descending order. Conversely, stress-sensitivity indices, such as Tolerance (TOL) and Stress Susceptibility Index (SSI), were ranked in ascending order, with lower values indicating greater tolerance. The RS for each genotype was obtained by summing the ranks from all indices. Genotypes with the lowest RS were considered the most drought-tolerant and received the highest final ranking, and vice versa12.
Results
Analysis of variance (ANOVA)
The analysis of variance (ANOVA) revealed highly significant effects (p < 0.01) for treatment, genotype, and the genotype × treatment (G × E) interaction across all evaluated traits (Tables 2 and 3). The significant G × E interaction indicated that genotypes responded differently to water stress. Genotypes VI003685AG, VI001408BG, and VI001435AG exhibited stable performance across both WW and DS conditions for yield and physiological traits, indicating robustness. In contrast, genotypes like VI001514AG and VI000938AG demonstrated greater sensitivity, showing significant declines in performance under drought conditions. These findings underscore the potential to identify genotypes with consistent performance across environments to develop drought-resilient cultivars.
Growth, morphological, physiological, and yield attributes of mungbean genotypes
Mungbean genotypes exhibited substantial variation under both well-watered (WW) and drought-stressed (DS) conditions. Violin plots (Fig. 1) revealed distinct shifts in trait distributions, with most traits showing significant declines under drought stress. Plant height (PH), primary branches (NPB), secondary branches (NSB), pods per plant (PP), seeds per pod (SPP), pod length (PL), test weight (TW), grain yield (GY), and leaf area (LA) all decreased significantly under drought stress, indicating the negative impact of water limitation on growth, reproduction, and photosynthetic potential.

Violin plot illustrating the genotypic variability in morpho-physiological traits of mungbean under well-watered (WW) and drought-stressed (DS) conditions. The plot visualizes the distribution, density, and spread of trait values across different genotypes, highlighting the effects of drought stress on trait expression. Significance was tested at P < 0.05%. Note: PH: Plant height, NDVI: Normalised Difference Vegetation Index, Qmax: Photosystem II efficiency, VPD: Vapour Pressure Deficit, GH2O: Stomatal Conductance.
Physiological traits showed the most drastic changes. Net CO2 assimilation (A), stomatal conductance (GH2O), and transpiration rate (E) dropped significantly (− 51.8%, − 49.3%, and − 48.8%, respectively) under drought stress. Conversely, leaf temperature (Tleaf) and vapour pressure deficit (VPD) increased by 8.1% and 27.5%, respectively, reflecting a greater evaporative demand. Chlorophyll fluorescence (Qmax) declined slightly by 12%, indicating a relative stability of PSII under drought conditions.
The magnitude of drought-induced reductions varied across traits (Table 4). For example, PH decreased by 28.5%, NPB by 33.2%, and NSB by 24.9%, while PP showed a moderate decline of 7%, and PL remained stable. Grain yield demonstrated considerable genotypic variation, with VI003685AG achieving the highest yield under both WW (5.52 g) and DS (5.27 g) conditions, whereas VI001514AG produced the lowest yields (0.58 g WW; 0.43 g DS).
Genotypic variability was observed in both morphological and physiological traits. For instance, PH ranged from 18.7 to 62.3 cm under WW and 12.11 to 46.51 cm under DS. NPB ranged from 3 to 10 (WW) and 2 to 8.15 (DS), while NSB varied from 3 to 13.5 (WW) and 2 to 10 (DS). PP ranged from 6.01 to 41 (WW) and 4 to 38.01 (DS), while PL remained stable at 4.26 to 12.36 cm across both conditions. TW showed a slight decrease of 4.8%, and LA ranged from 11.62 to 48.02 cm2 under WW and 3.18 to 39.29 cm2 under DS. NDVI was highest in Vaibhav (0.74) under WW and VI003685AG (0.74) under DS. The photosynthetic rate (A) peaked in VI003685AG under both conditions, while VI002739AG had the lowest rate under DS. Stomatal conductance and transpiration varied significantly, with VI003685AG consistently performing best.
Overall, the data show that drought stress adversely affected most traits, but the extent of reduction varied widely among genotypes. Genotype VI003685AG exhibited superior performance across morphological, physiological, and yield parameters, indicating its strong potential for drought tolerance.
Assessment of genetic variability in mungbean genotypes under WW and DS conditions
Genetic variability parameters, including phenotypic coefficient of variation (PCV), genotypic coefficient of variation (GCV), broad-sense heritability (h2bs), and genetic advance as a percentage of the mean (GAM), are summarised in Table 5. For most traits, PCV values were only slightly higher than their corresponding GCV values. Traits such as PH, PB, NSB, PP, NSP, TW, GY, and key physiological parameters (A, E, and gH2O) showed high GCV and PCV values (> 20%) under both WW and DS conditions. In contrast, Qmax, NDVI, and VPD displayed low to moderate variability, suggesting that these traits are more stable across genotypes.
Heritability estimates were high for most traits (> 0.60), indicating that the traits are predominantly under genetic control and thus responsive to selection. High GAM values, especially for yield-related and physiological traits, further underscore the potential for significant genetic improvement through breeding.
Trait associations between grain yield and morpho-physiological traits
Pearson’s correlation analysis revealed significant positive associations between grain yield (GY) and several morphological and physiological traits under both WW and DS conditions (Figs. 2 and 3). Under WW conditions, GY was positively correlated with PH (r = 0.35), NPB (r = 0.45), NSB (r = 0.51), PP (r = 0.55), NSP (r = 0.50), PL (r = 0.42), NDVI (r = 0.31), E (r = 0.30), GH2O (r = 0.19), LA (r = 0.40), and A (r = 0.27). TW and Qmax also showed positive, though non-significant, associations.

Association of morpho-physiological traits with yield and yield-related attributes of mungbean genotypes under well-watered (WW) conditions. The figure illustrates the relationships between key physiological parameters and yield components, providing insights into trait contributions to productivity under WW conditions. WW: Well Watered, PH: Plant height, NPB: Number of Primary Branches, NSB: Number of Secondary Branches, PP: Pods per Plant, PL: Pod Length, TW: Test Weight, NDVI: Normalized Difference Vegetation Index, Tleaf: Leaf Temperature, E: Transpiration rate, A: Net CO2 Assimilation rate, VPD: Vapour Pressure Deficit, GH2O: Stomatal Conductance, LA: Leaf area per leaf, GY: Grain yield per plant.

Association of morpho-physiological traits with yield and yield-related attributes of mungbean genotypes under drought-stressed (DS) conditions. The figure highlights the relationships between key physiological traits and yield components, offering insights into trait contributions to productivity under drought stress conditions. DS: Drought Stress, PH: Plant height, NPB: Number of Primary Branches, NSB: Number of Secondary Branches, PP: Pods per Plant, PL: Pod Length, TW: Test Weight, NDVI: Normalized Difference Vegetation Index, Tleaf: Leaf Temperature, E: Transpiration rate, A: Net CO2 Assimilation rate, VPD: Vapour Pressure Deficit, GH2O: Stomatal Conductance, LA: Leaf area per leaf, GY: Grain yield per plant.
Under DS conditions, these relationships were strengthened. GY exhibited strong positive correlations with PH (r = 0.67), NPB (r = 0.58), NSB (r = 0.62), PP (r = 0.53), NSP (r = 0.51), PL (r = 0.38), TW (r = 0.18), NDVI (r = 0.52), E (r = 0.57), gH2O (r = 0.38), LA (r = 0.60), Qmax (r = 0.41), and A (r = 0.63). These strong correlations highlight the importance of canopy development, reproductive efficiency, and photosynthetic performance in maintaining productivity under drought. Conversely, Tleaf (r = − 0.18) and VPD (r = − 0.30) were negatively correlated with GY under DS, indicating that increased heat load and evaporative demand contribute to yield penalties.
Stress tolerance indices and genotype ranking
Yield-based drought tolerance indices (Table 6) provided additional insights into genotype performance across WW and DS environments. VI003685AG maintained the highest grain yield in both conditions, while VI001514AG consistently produced the lowest. Tolerance index (TOL) values varied widely, with VI001435AG (0.001) and VI001974BG (0.005) showing minimal yield reductions under DS, indicating superior stability. In contrast, the check variety PDM-139 recorded the highest TOL (0.54), reflecting high sensitivity. At the same time, mean productivity (MP) and geometric mean productivity (GMP) identified VI001191BG as the top performer, whereas VI001514AG ranked lowest. The stress susceptibility index (SSI) further highlighted VI001435AG (0.001) as the most tolerant, while VI000938AG (0.48) was the most susceptible. The stress tolerance index (STI) placed VI001191BG highest (4.21), and the yield index (YI) and yield stability index (YSI) favoured VI003957AG and V001435AG, respectively.
Additionally, to integrate performance across all indices, a cumulative ranking approach was used. Genotypes VI003685AG, VI002051BG, VI004811BG, and VI000852AG emerged as the most drought-tolerant and stable performers, whereas check varieties such as Vaibhav, PAU-911, NM-94, EC-0398949, and IPM-205-7 ranked substantially lower than those genotypes.
Principal component analysis (PCA) and multi-trait genotype-ideotype distance index (MGIDI)
Principal Component Analysis (PCA) was conducted to examine the multivariate structure of trait variation across the mungbean genotypes. The first two principal components (PC1 and PC2) explained 30.7% of the total variability (Fig. 4). PC1 (17.3%) was primarily associated with yield-related traits, including NSP, PL, NPB, NSB, and GY, suggesting that PC1 captures the genotypic variation in reproductive potential and branching capacity. Genotypes that loaded higher on PC1, such as VI003379BG and VI003744AG, exhibited superior performance in NSP and PL, indicating that these genotypes possess better yield components (i.e., more seeds per pod and longer pods). This highlights the importance of PC1 in determining genotypic performance related to reproduction and overall productivity under both well-watered and drought-stressed conditions.

Principal Component Analysis (PCA) biplot and scree plot illustrating the relationships among morpho-physiological traits, the distribution of 122 mungbean genotypes under drought stress, and the proportion of variance explained by each principal component. PC1 and PC2 together capture the majority of variation and effectively differentiate genotype responses.
In contrast, PC2 (13.4%) primarily represented the variation in physiological traits, including E, LA, VPD, and GH2O, reflecting differences in canopy activity, water-use efficiency, and stomatal regulation under drought stress. Genotypes such as VI000316AG and VI001400AG, which had higher loadings on PC2, were positioned closer to the PC2 axis, suggesting these genotypes possess better vegetative architecture and enhanced physiological responses to water stress. These genotypes demonstrated efficient gas exchange and canopy development, which are critical for sustaining productivity under drought conditions by improving the plant’s ability to capture and use water efficiently. The PCA biplot (Fig. 4) provides a visual representation of how the various traits contribute to genotype performance across water regimes. Traits such as GY, NSP, PL, and NPB are strongly associated with PC1, emphasising their importance in determining yield potential. This aligns with the fact that higher PC1 loadings correspond to superior yield components. Conversely, physiological traits such as E, GH2O, and VPD are more strongly correlated with PC2, highlighting their roles in drought adaptation mechanisms, such as water-use efficiency and stomatal conductance.
To integrate all the traits into a unified framework and identify genotypes with superior drought resilience, the Multi-trait Genotype-Ideotype Distance Index (MGIDI) was applied. The MGIDI ideotype was explicitly defined to align with breeding goals: higher values were considered desirable for PH, NPB, NSB, PP, NSP, PL, TW, GY, A, E, GH2O, NDVI, LA, and Qmax, whereas lower values were preferred for Tleaf and VPD. The ideotype thus represents an ideal genotype with balanced traits that contribute to high productivity and physiological performance under drought stress. All traits were standardised and direction-adjusted, so that lower MGIDI values indicated genotypes closer to the ideotype and thus more desirable for breeding. Genotypes such as VI003685AG, VI002051BG, and VI003957AG were found to be closest to the ideotype, demonstrating a balanced combination of desirable morphological, yield, and physiological traits. These genotypes exhibited superior performance across a wide range of traits, including yield components, canopy development, and physiological performance under drought conditions, making them strong candidates for drought-tolerant breeding. MGIDI rankings further confirmed that these genotypes were among the best performers, combining productive plant architecture with enhanced drought-resilience mechanisms.
Factor analysis grouped traits into seven latent factors representing plant architecture, reproductive attributes, physiological drought-response mechanisms, and grain-yield determinants (Table 7). Branching traits (NPB and NSB) showed strong loadings within FA1, NSP, and PL defined FA2, and key physiological traits (E, gH2O, VPD) clustered within FA3, with high communalities indicating robust trait representation. The ideotype was explicitly defined to align with breeding goals: higher values were considered desirable for PH, NPB, NSB, PP, NSP, PL, TW, GY, A, E, gH2O, NDVI, LA and Qmax, whereas lower values were desirable for VPD and Tleaf. All traits were standardised and direction-adjusted so that lower MGIDI values indicated greater similarity to the ideotype (Fig. 5).

MGIDI scores and factor contributions for 122 mungbean genotypes under drought stress. Genotypes with lower MGIDI values are closer to the ideotype and considered more drought-tolerant. Factor contributions indicate which trait groups most influenced each selected genotype’s performance.
Selection differentials demonstrated substantial improvement potential (Table 8). Grain yield exhibited the highest predicted gain (+ 23.0%), followed by strong gains for NSB (+ 20.2%), gH2O (+ 19.2%) and NPB (+ 19.1%). Moderate positive gains in NDVI, LA and E further supported their contribution to drought adaptation. Based on MGIDI rankings, 18 genotypes were identified as closest to the ideotype, including VI002051BG, VI003685AG, VI001408BG, VI003957AG, VI002402BG, VI000316AG, VI001126BG, VI001435AG, VI003232AG, VI001448A-BLM, VI000852AG, VI001482BG, VI001284AG, VI004811BG, VI001419BG, VI000942AG, VI001400AG and VI004096AG. These genotypes demonstrated a balanced combination of productive plant architecture, favourable yield components and enhanced physiological performance under moisture stress. Overall, the application of the combined PCA–MGIDI approach provided a robust, multivariate framework for identifying genotypes that balance high productivity with physiological efficiency under water stress, offering valuable insights for targeted breeding programs aimed at developing drought-resilient mungbean cultivars.
Discussion
Drought stress represents a major limitation in rainfed mungbean production, particularly during the flowering and pod-filling stages, which are critical for yield determination4,6. In this study, 122 genetically diverse genotypes were evaluated under well-watered (WW) and drought-stressed (DS) conditions using a semi-controlled greenhouse setup. This approach allowed for precise stress imposition and phenotyping. However, the confined rooting volume in pots likely intensified drought stress compared with field conditions. Restricted root growth can limit water acquisition and exaggerate differences among genotypes, particularly for traits related to drought avoidance or deeper soil water extraction31.
Additionally, the study imposed only a single terminal drought, while mungbean often encounters intermittent or early-season drought in the field. Genotypes performing well under terminal stress may not exhibit the same resilience under fluctuating stress patterns. Similarly, grain yield per plant in pots does not fully reflect yield per unit area in the field, where factors like competition, plant architecture, and lodging susceptibility influence overall productivity. These factors underscore the importance of field validation of top-performing genotypes to ensure greenhouse results are representative of field performance.
Despite these limitations, substantial genotypic variation was observed for both morphological and physiological traits under WW and DS conditions. Yield-contributing traits, including plant height (PH), branching (NPB, NSB), pod number (PP), and leaf area (LA), exhibited strong positive correlations with grain yield (GY). In contrast, Tleaf and VPD were negatively associated with GY under DS, underscoring the importance of canopy development and efficient water-use regulation for drought resilience32,33. Physiological traits such as net photosynthesis (A), stomatal conductance (gH2O), and transpiration rate (E) retained moderate values in high-performing genotypes, suggesting partial maintenance of photosynthetic activity despite water limitation34,35,36,37. High heritability and genetic advance for these traits further suggest that direct selection for yield and key physiological parameters can be effective for improving drought adaptation38,39,40,41.
To integrate multivariate trait responses, Principal Component Analysis (PCA) and the Multi-trait Genotype-Ideotype Distance Index (MGIDI) were employed. PCA revealed six principal components accounting for over 87% of the total variation, with the first component representing yield-related and architectural traits and the second capturing physiological attributes. This approach allowed the identification of genotypes with complementary adaptation strategies. For example, G91 (VI003379BG) and G104 (VI003744AG) were associated with reproductive yield components, while G8 (VI000316AG) and G98 (VI003648BG) were associated with vegetative branching, indicating distinct approaches to stress tolerance. The MGIDI further refined selection by integrating trait desirability, highlighting genotypes such as VI003685AG, VI002051BG, and VI004811BG, which combined high yield, stable branching, and favourable physiological traits, with predicted selection gains of + 23% for grain yield and + 20.2% for secondary branching42. This multivariate framework emphasises stable, integrated performance rather than focusing on single-trait extremes, partially mitigating the limitations inherent in pot-based screening.
However, the physiological mechanisms underlying tolerance in the top-ranked genotypes remain largely theoretical. The absence of root system data, water relations measurements, and biochemical assessments (e.g., osmolytes, antioxidants) limits our ability to interpret these mechanisms fully. High-performing genotypes likely combine efficient stomatal regulation, sustained photosynthesis, and canopy cooling to maintain productivity under drought stress, but these processes require validation through targeted physiological and molecular analyses34,35,36,37. Therefore, while the study provides valuable insights into genotype performance and trait associations, the findings must be regarded as preliminary until confirmed under field conditions, where natural variation in drought timing and intensity is present.
Overall, this study identifies genotypes with promising drought resilience and demonstrates the utility of integrating PCA and MGIDI to capture multidimensional trait performance. The combined evaluation of morphological, physiological, and yield traits provides a robust framework for preliminary selection, laying the foundation for breeding programs targeting drought-adaptive mungbean varieties. However, multi-environment field trials, along with complementary analyses of root traits, water-use efficiency, and biochemical responses, will be crucial to confirm the durability and mechanistic basis of the observed tolerance and to ensure that high-performing genotypes identified under controlled conditions perform well in real-world cultivation scenarios.
These findings are preliminary and require multi-location field validation before being utilised for breeding purposes. Although the greenhouse-based screening provided valuable insights into genotype performance under controlled drought stress, these results may not fully capture the complexity of field conditions. Therefore, further evaluation across diverse agro-ecological zones is essential to confirm the stability and reliability of the identified drought-resilient genotypes under real-world conditions.
Conclusion
This study identified drought-tolerant mungbean genotypes through an integrated approach combining phenotypic evaluation, physiological screening, stress indices, PCA and the MGIDI. Significant genetic variability was observed among the genotypes, highlighting strong potential for preliminary selection and improvement. Stress indices, such as the STI and GMP, efficiently identified genotypes with stable yield across environments, while PCA pinpointed key traits like grain yield, stomatal conductance, and transpiration rate as critical to drought adaptation. The MGIDI index provided a refined selection tool, identifying genotypes such as VI003685AG, VI002051BG, and VI003957AG as the most preliminarily identified drought-resilient genotypes. These genotypes exhibited superior performance in both morphological and physiological traits, demonstrating the value of a multifaceted selection approach. While pot-based screening was effective for controlled drought stress, further multi-location field validation is needed to confirm the stability and reliability of these genotypes under real-field conditions. The greenhouse conditions may not fully reflect the complexities of field environments, where factors such as competition, soil conditions, and environmental stress patterns influence genotype performance. Therefore, further evaluation across diverse agro-ecological zones is essential to confirm the stability and reliability of these drought-resilient genotypes. Overall, this study provides valuable insights and promising genetic material for breeding potential drought-tolerant mungbean cultivars, contributing to food security in the face of climate change.
Data availability
The manuscript presents all the data generated in this research. Additional data is provided in the supplementary material file.
References
Yi-Shen, Z., Shuai, S. & FitzGerald, R. Mung bean proteins and peptides: Nutritional, functional, and bioactive properties. Food Nutr. Res. 62, 1290. https://doi.org/10.29219/fnr.v62.1290 (2018).
Islam, M. R. et al. Drought tolerance in mung bean is associated with the genotypic divergence, regulation of proline, photosynthetic pigment, and water relation. Phyton-Int. J. Exp. Bot. 92, 955–981. https://doi.org/10.32604/phyton.2023.025138 (2023).
Indiastat. Agriculture: Selected State, Season-Wise Area, Production, Product. Indiastat https://www.indiastat.com/table/agriculture/selected-state-season-wise-area-production-product/1457200 (2024).
Ikram, S., Bhattarai, S. & Walsh, K. B. Screening new mungbean varieties for terminal drought tolerance. Agriculture 14, 1328. https://doi.org/10.3390/agriculture14081328 (2024).
Nunes, C. et al. Cowpea physiological responses to terminal drought—Comparison between four landraces and a commercial variety. Plants 11, 593. https://doi.org/10.3390/plants11050593 (2022).
Nivethitha, T., Babu, C. & Jayamani, P. Impact of drought stress on seedling vigor and physio-biochemical attributes of diverse mungbean genotypes for drought tolerance through in vitro and field evaluation. Euphytica 220, 182. https://doi.org/10.1007/s10681-024-03438-8 (2024).
Ullah, I. et al. Projected changes in socioeconomic exposure to heatwaves in South Asia under changing climate. Earth’s Future 10, e2021EF002240 (2022).
Bonaccorsi, G. et al. Economic and social consequences of human mobility restrictions under COVID-19. Proc. Natl. Acad. Sci. USA 117, 15530–15535. https://doi.org/10.1073/pnas.2007658117 (2020).
Haeften, V. S. et al. Building a better mungbean: Breeding for reproductive resilience in a changing climate. Food Energy Secur. 12, e467. https://doi.org/10.1002/fes3.467 (2023).
Farooq, M. et al. Heat stress in grain legumes during reproductive and grain-filling phases. Crop Pasture Sci. 68, 985–1005. https://doi.org/10.1111/jac.12169 (2017).
Basavaraj, P. S. et al. Stress tolerance indices for the identification of low phosphorus tolerant introgression lines derived from Oryza rufipogon Griff. Plant Genet. Resour. 19, 328–338. https://doi.org/10.1017/S1479262121000381 (2021).
Basavaraj, P. S. et al. Index-based selection of chickpea (Cicer arietinum L.) genotypes for enhanced drought tolerance. Sci. Rep. 15, 8282. https://doi.org/10.1038/s41598-025-93273-1 (2025).
Biju, S., Fuentes, S. & Gupta, D. The use of infrared thermal imaging as a non-destructive screening tool for identifying drought-tolerant lentil genotypes. Plant Physiol. Biochem. 127, 11–24. https://doi.org/10.1016/J.PLAPHY.2018.03.005 (2018).
Wang, J. et al. Analysis of physiological indicators associated with drought tolerance in wheat under drought and re-watering conditions. Antioxidants 11, 2266. https://doi.org/10.3390/antiox11112266 (2022).
Hussain, T., Akram, Z., Shabbir, G., Manaf, A. & Ahmed, M. Identification of drought-tolerant chickpea genotypes through multi-trait stability index. Saudi J. Biol. Sci. 28, 6818–6828. https://doi.org/10.1016/j.sjbs.2021.07.056 (2021).
Reddy, S. S. et al. Spatio-temporal evaluation of drought adaptation in wheat revealed NDVI 1 and MTSI as powerful tools for selecting tolerant genotypes. Field Crops Res. 311, 109367. https://doi.org/10.1101/2023.01.29.526148 (2024).
Schafleitner, R. et al. The AVRDC–The World Vegetable Center mungbean (Vigna radiata) core and mini core collections. BMC Genomics 16, 344. https://doi.org/10.1186/s12864-015-1556-7 (2015).
Govaerts, B. & Verhulst, N. The normalized difference vegetation index (NDVI) Greenseeker handheld sensor: Toward the integrated evaluation of crop management. Part A: Concepts and case studies (2010).
Nedbal, L., Soukupová, J., Kaftan, D., Whitmarsh, J. & Trtílek, M. Kinetic imaging of chlorophyll fluorescence using modulated light. Photosynth. Res. 66, 3–12 (2000).
Diao, H. et al. Dry inside: Progressive unsaturation within leaves with increasing vapour pressure deficit affects estimation of key leaf gas exchange parameters. New Phytol. 244, 1275–1287. https://doi.org/10.1111/nph.20078 (2024).
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4, 9 (2001).
Johnson, H. W., Robinson, H. F. & Comstock, R. E. Estimates of genetic and environmental variability in soybeans. Agron. J. 47, 314–318 (1955).
R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. https://doi.org/10.59350/t79xt-tf203 (2023).
Olivoto, T., Diel, M. I., Schmidt, D. & Lúcio, A. D. MGIDI: a powerful tool to analyze plant multivariate data. Plant Methods 18, 121 (2022).
Rosielle, A. A. & Hamblin, J. Theoretical aspects of selection for yield in stress and non-stress environments. Crop Sci. 21, 943–946. https://doi.org/10.2135/cropsci1981.0011183X002100060033x (1981).
Fernandez, G. C. J. Effective selection criteria for assessing plant stress tolerance. In Adaptation of Food Crops to Temperature and Water Stress (ed. Kuo, C. G.), 257–270. AVRDC (1992).
Fischer, R. A. & Maurer, R. Drought resistance in spring wheat cultivars. I. Grain yield responses. Aust. J. Agric. Res. 29, 897–912. https://doi.org/10.1071/AR9780897 (1978).
Gavuzzi, P. et al. Evaluation of field and laboratory predictors of drought and heat tolerance in winter cereals. Can. J. Plant Sci. 77, 523–531. https://doi.org/10.4141/P96-130 (1997).
Bouslama, M. & Schapaugh, W. T. Stress tolerance in soybeans: Evaluation of three screening techniques for heat and drought tolerance. Crop Sci. 24, 933–937. https://doi.org/10.2135/cropsci1984.0011183X002400050026x (1984).
Farshadfar, E. & Elyasi, P. Screening quantitative indicators of drought tolerance in bread wheat (Triticum aestivum L.) landraces. Eur. J. Exp. Biol. 2, 577–584 (2012).
Turner, N. C. Imposing and maintaining soil water deficits in drought studies in pots. Plant Soil 439, 45–55. https://doi.org/10.1007/s11104-018-3893-1 (2019).
Wang, J. et al. Exploitation of drought tolerance-related genes for crop improvement. Int. J. Mol. Sci. 22, 10265. https://doi.org/10.3390/ijms221910265 (2021).
Amitrano, C., Arena, C., Cirillo, V., De Pascale, S. & De Micco, V. Leaf morpho-anatomical traits in Vigna radiata L. affect plant photosynthetic acclimation to changing vapor pressure deficit. Environ. Exp. Bot. 186, 104453. https://doi.org/10.1016/j.envexpbot.2021.104453 (2021).
Qiao, M., Hong, C., Jiao, Y., Hou, S. & Gao, H. Impacts of drought on photosynthesis in major food crops and the related mechanisms of plant responses to drought. Plants 13, 1808. https://doi.org/10.3390/plants13131808 (2024).
Yuan, X. et al. Impacts of global climate change on agricultural production: A comprehensive review. Agronomy 14, 1360. https://doi.org/10.3390/agronomy14071360 (2024).
Dong, X. et al. Impacts of high temperature, relative air humidity, and vapor pressure deficit on the seed set of contrasting maize genotypes during flowering. J. Integr. Agric. 23, 2955–2969. https://doi.org/10.1016/j.jia.2023.09.007 (2024).
Salehi-Soghadi, Z., Islam, M. S., Manschadi, A. M. & Kaul, H.-P. Transpiration efficiency of some potato genotypes under drought. Agronomy 13, 996; https://doi.org/10.3390/agronomy13040996 (2023).
Kumar, J. et al. Phenotyping of traits imparting drought tolerance in lentil. Crop Pasture Sci. 63, 547–554. https://doi.org/10.1071/CP12168 (2012).
Singh, C. M., Singh, P., Tiwari, C., Purwar, S., Kumar, M., Pratap, A. & Mishra, A. K. Improving drought tolerance in mungbean (Vigna radiata L. Wilczek): Morpho-physiological, biochemical and molecular perspectives. Agronomy 11, 1534. https://doi.org/10.3390/agronomy11081534 (2021).
Roy, S. C. & Shil, P. Assessment of genetic heritability in rice breeding lines based on morphological traits and caryopsis ultrastructure. Sci. Rep. 10, 7830. https://doi.org/10.1038/s41598-020-63976-8 (2020).
Mutari, B., Sibiya, J., Gasura, E., Matova, P. M., Simango, K. & Kondwakwenda, A. Genetic analysis of grain yield and yield-attributing traits in navy bean (Phaseolus vulgaris L.) under drought stress. Euphytica 218, 51. https://doi.org/10.1007/s10681-022-03001-3 (2022).
Olivoto, T. & Nardino, M. MGIDI: A novel multi-trait index for genotype selection in plant breeding. BioRxiv 2020–07, 2020. https://doi.org/10.1101/2020.07.23.217778 (2020).
Acknowledgements
The authors are thankful to ICAR-NICRA for the support through the Project “Phenotyping of Pulses for Enhanced Tolerance to Drought and Heat (XX01737) and ICAR-NIASM for the logistic support. The authors also thank the World Vegetable Centre, South & Central Asia office, Hyderabad, for sharing the seeds for experimentation.
Source of seeds
Seeds were procured from World Vegetable Center - South and Central Asia, IN, Hyderabad- 502324, India.
Funding
This research was supported by ICAR-NICRA under the project “Phenotyping of Pulses for Enhanced Tolerance to Drought and Heat (XX01737)”.
Author information
Authors and Affiliations
Contributions
BPS: Conception, writing and editing; RB, IVG, AG, SS, SG; data collection: JR, BKM, HCB, HMH, and SRK Manuscript editing; DS: editing and formal data analysis RN: Seed Supply and Editing manuscript. All the authors read the manuscript and approved it for submission.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Basavaraj, P.S., Babar, R., Gangurde, A. et al. Unveiling drought-tolerant mungbean genotypes through integrated multi-trait selection. Sci Rep 16, 6018 (2026). https://doi.org/10.1038/s41598-026-36830-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-36830-6
