
Abstract
Juvenile blacktip sharks (Carcharhinus limbatus) are highly abundant in the mangrove-fringed bays throughout the Galapagos Archipelago, utilizing these habitats as nursery grounds. Due to spatial variation in factors including temperature and larval connectivity, fish species richness is higher in the eastern portion of the archipelago. Predator diets often reflect regional prey availability, leading us to hypothesize that juvenile blacktip sharks in the eastern Galapagos would have greater prey taxa richness than those in the west. Here, we used DNA metabarcoding of fecal matter collected via non-lethal cloacal swabs to describe and quantify juvenile blacktip shark diets. We collected samples from 107 sharks across 240 km, spanning 14 bays on four islands, and released all sharks alive. The most common prey taxa identified were Thoburn’s mullet (Mugil thoburni; 20.0%, based on percent of occurrence), followed by fishes in the genus Abudefduf (sergeant-majors; 13.4%), Galapagos ringtail damselfish (Stegastes beebei; 12.7%), scalloped hammerhead shark (Sphyrna lewini; 11.7%), and yellowtail damselfish (Stegastes arcifrons; 10.7%). Prey taxa richness was highest among sharks sampled in the bays of San Cristobal, possibly due to geographic variation in prey availability. Additionally, sharks sampled during a La Niña event in 2021 had higher prey taxa richness than those sampled during an El Niño event in 2023, which could be due to El Niño-induced declines in species richness and abundance resulting from shifts in primary productivity and upwelling. A deeper understanding of shark diets can improve our ability to predict how changes in food web dynamics and prey availability affect shark populations. This knowledge is especially vital in the face of overfishing and climate change, helping to inform more effective conservation strategies and support shark survival.
Similar content being viewed by others

Introduction
Prey availability and consumption are pivotal in understanding the ecological interactions and behaviors of consumers across different environments. What and how much consumers eat, has a strong effect on their behavior, survival, and fitness, as well as the demography of the population. Moreover, consumer diets vary enormously with seasonality and among locations as a function of caloric demands, prey availability, and the presence or absence of competitors and predators. For example, the diet of pumas (Felis concolor) varies by latitude, with temperate populations typically specializing in a few larger prey species, while tropical populations consume a greater diversity of smaller prey1. Understanding biogeographic variation in consumer diets can provide insight into an animal’s ability to adapt to environmental disturbances and fluctuations in prey availability across time and space. For example, wolves (Canis lupus ligoni) in Alaska’s Alexander Archipelago responded to a decline in ungulate prey abundance and availability by broadening their overall dietary diversity rather than shifting to a single alternative prey source2.
The Galapagos Archipelago is one of the most unique biogeographical regions on Earth, shaped by an anomalous ocean climate that fosters distinct marine ecosystems in close proximity3. As a result, its biota includes a unique mix of species typically found in tropical (e.g., manta rays, reef sharks, corals), temperate (e.g., sea lions, kelp), and even subantarctic (e.g., fur seals, penguins, albatross) environments. Species richness of both reef fishes and macro-invertebrates vary throughout the Galapagos3 and the archipelago is divided into bioregions characterized by variable species composition driven by temperature and productivity3. In general, the western portion of the archipelago has cooler water temperatures with more temperate species, while the eastern side of the archipelago is relatively warmer and has more tropical species and greater overall richness. The temperate western regions have the lowest overall fish species richness3,4 and exhibit the highest levels of endemism3. These different bioregions and the distinct variation of fish and invertebrate species across the archipelago have led the diet of some predator species, such as the Galapagos sea lion (Zalophus wollebaeki), to differ depending on where they are foraging5.
While the Galapagos Archipelago’s diverse marine ecosystems support a wide array of species, they also provide critical habitats for specific life stages of certain animals, particularly in the case of shark nurseries. Numerous shark species, including the blacktip (Carcharhinus limbatus) and scalloped hammerhead (Sphyrna lewini), use nursery habitats as juveniles6,7,8. Shark nurseries are often located in coastal, protected bays and estuaries7, which can be susceptible to increased anthropogenic impacts (e.g., pollution, tourism, and fishing)9. Shark nurseries are typically in mangrove-fringed bays7,9, which also provide nurseries for other juvenile fishes, potentially increasing the abundance of possible prey for the juvenile sharks1,4. It has been hypothesized that sharks use nursery habitats for their greater prey abundance and accessibility to food sources as well as a refuge from predation10,11, although more research is required to fully understand their purpose8.
Blacktip sharks have a circumglobal distribution, inhabiting continental and insular shelves in tropical and subtropical regions12. In the Gulf of Mexico, female blacktip sharks reach maturity at 6–7 years of age (158–162 cm TL), whereas males reach maturity at 4–5 years of age (133–136 cm TL)13. The length at birth is estimated to be 50–60 cm TL14. Juvenile blacktips are also abundant throughout the Galapagos Archipelago9,15,16. Based on previous studies that have used morphological stomach contents analysis, Clupeidae and Sciaenidae fishes had the largest contribution to the blacktip shark’s diet worldwide8. However, the importance of specific prey species in their diet varies geographically, for example due to the variation in size/age of the sharks analyzed and differentiation in prey availability8. While blacktip sharks exhibit ontogenetic dietary shifts17, research on juvenile blacktip diets has often been limited due to the reliance on morphological stomach content analysis, which can be imprecise11,17,18. Additionally, most studies have focused on juvenile blacktip sharks caught in the Gulf of Mexico11,19,20,21,22,23.
Only one study has investigated the trophic ecology of juvenile blacktips in the Galapagos, in which they used stable isotope analysis (SIA) from sharks caught on two different islands (i.e., Santa Cruz and Santiago)24. While SIA is valuable for determining the relative trophic position of a predator over extended periods, it lacks the taxonomic resolution needed to identify specific prey species. Alternatively, DNA metabarcoding of fecal DNA (fDNA) provides a more precise alternative for diet analysis, enabling researchers to identify prey at higher taxonomic resolutions (e.g., to species). DNA metabarcoding is a technique used to identify many taxa within the same sample and has become a popular method for diet reconstruction studies5,25,26,27,28. The use of cloacal swabs to collect fDNA from sharks for DNA metabarcoding is also a less invasive, non-lethal, and more precise method for diet analysis compared to traditional morphological stomach contents analysis29,30,31. Cloacal swabbing for shark diet reconstruction was first applied by van Zinnicq Bergmann et al. (2021) 67, who collected samples from captive lemon sharks (Negaprion brevirostris) during a controlled feeding trial and from wild bull sharks (Carcharhinus leucas). Ryburn et al. (2026)33 found no statistical difference in overall diet composition when comparing metabarcoded stomach contents and cloacal swabs collected from the same individuals across four shark species: blacktip, bonnethead (Sphyrna tiburo), blacknose (Carcharhinus acronotus), and Atlantic sharpnose (Rhizoprionodon terraenovae).
Here we describe and quantify the diet of the juvenile blacktip shark across 14 bays on four islands in the Galapagos using DNA metabarcoding of fDNA collected with non-lethal cloacal swabs. The purpose of our study was to conduct a biogeographical analysis of juvenile blacktip shark foraging patterns. Given that fish species richness is higher on the eastern side of the archipelago, we hypothesized that the dietary richness of juvenile blacktip sharks would reflect this pattern.
Methods
Shark capture
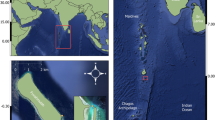
We sampled 107 juvenile blacktip sharks (694 ± 7 mm stretched total length [STL], 495–940 mm range) across four islands in the Galapagos (i.e., San Cristobal, Isabela, Fernandina, and Santiago; Fig. 1). Sampling took place in 14 bays between June 2021 and July 2023 (Table 1). These bays were chosen based on knowledge from local fishermen and from our team’s experience in observing blacktips. San Cristobal had a larger sample size due to ease of daily access because the Galapagos Science Center (GSC) is located on San Cristobal, whereas samples from the other islands were collected during two research cruises in June of 2022 and February of 2023. To capture the sharks, we used gillnets (75 of the 107 sharks), similar to those used by local artisanal fishermen, seine nets (28 of the 107 sharks), and handlines (4 of the 107 sharks). When possible, we used a SonTek CastAway-CTD to measure the water temperature for each sampling before the hooks or net were placed into the water.

Map of the Galapagos Archipelago showing the 14 locations where juvenile blacktip sharks were sampled. Each point represents a specific sampling site. Puerto Grande and Cartago Bay are the only two nursery habitats where juvenile blacktip sharks and juvenile scalloped hammerhead sharks co-occur.
A large beach seine, measuring 100 m in length and 3 m in depth with 1.27 cm eye-mesh, was deployed in Puerto Grande to encircle the shallow area along the beach. This net was used exclusively at this site, as Puerto Grande was the only bay with a sandy bottom and a sufficiently large beach to accommodate its use. In contrast, the other 13 bays had rocky bottoms or lacked suitable beaches, making the seine net impractical. As a result, gillnets and handlines were the primary sampling methods used in the other locations. Sampling with the seine net in Puerto Grande took place at sunrise. The process began with one end of the net being anchored at one side of the beach, while the rest of the net was deployed from a boat to encircle the area. Once the net looped around to the opposite end of the beach, it was anchored there and then pulled ashore, creating a small, confined area where sharks were enclosed. The entire process took approximately 10 min, during which the sharks remained in the water, swimming freely within the enclosed area, until they were individually worked up.
A 100 × 3.5 m gillnet with 6.35 cm eye-mesh was used in the shallow areas at all bays and soaked for approximately one hour per deployment. To avoid shark mortality, the net was constantly monitored and sharks were immediately removed from the net as soon as they were captured.
Handlines were also used in all bays to target deeper areas that were inaccessible to the gillnet and seine net. Each handline was free-floating and comprised of a #4/0 barbless steel octopus hook attached to 0.7 mm, 25 kg monofilament line, 200 g polypropylene rope, and a foam buoy. We baited the hooks with either squid purchased from a local grocery or fresh mullet. Eight to 14 hooks were dispersed throughout the bay at a time. The buoys were constantly monitored, allowing sharks to be immediately brought onto the boat for the work up once hooked. We checked and re-baited the hooks as needed, approximately every 20–30 min.
Fieldwork and data collection were conducted under research permits PC-13-21, PC-18-22, PC-13-23, and MAATE-DBI-CM-2021-0174 granted by the Galapagos National Park and the Ecuadorian Ministry of Environment. Animal care and use were approved and overseen by UNC-CH’s IACUC (23–210.0) and was performed in accordance with relative guidelines and regulations. All sharks were released alive immediately after a brief workup, which lasted approximately three minutes. No sharks were harmed or killed as a result of this research.
fDNA collection
We followed the protocol outlined by Ryburn (2025) 32 and Ryburn et al. (2026) 33 to collect fecal DNA (fDNA) samples. If the shark was caught using a hook, the hook was immediately removed upon capture. The shark was then placed ventral side up on a flat surface, with its pelvic fins spread apart to prevent the swab from making contact with the exterior skin. The cloacal area was sterilized and dried by wiping the exterior with a Kimwipe soaked in 98% ethanol, which was discarded after a single use. Using a sterile, individually packed cotton-tipped swab with a wooden handle (Puritan 25–806 1WC FDNA; length 15.2 cm, tip diameter 0.46 cm) we carefully inserted the swab approximately 2.5 cm into the shark’s cloaca, past the sphincter, and rotated it for about 10 s. After removal, the swab was immediately placed into a sterile vial, stored in a cooler with ice, and the shark was released. The swabs were then stored at -20 °C until further processing.
Metabarcoding laboratory protocol and bioinformatics
We used the metabarcoding laboratory protocol from Ryburn (2025) 32 and Ryburn et al. (2026)33 . The full protocol is available on protocols.io (https://doi.org/10.17504/protocols.io.261ge5dnwg47/v2)34. For DNA extraction, we used the DNeasy Blood and Tissue Kit (www.qiagen.com), adhering to the manufacturer’s protocol with two adjustments: samples were digested at 70 °C for 24–48 h, and the DNA was eluted to 100 µl. The extracted DNA was stored at -20 °C until further processing.
We amplified two target regions for all samples: the MiFish-U35 region of the mitochondrial 12S rRNA gene and the Crustacean_16S26 mitochondrial rRNA gene. These primer sets were chosen based on previous diet studies that identified teleost fishes and crustaceans as primary components of the blacktip shark’s diet11,17. For both primer sets, Illumina Nextera-style adapters were appended to the 5’ ends of both the forward and reverse primer binding regions. PCR reactions were performed in triplicate for each sample and primer combination using the Takara High Fidelity PCR EcoDry™ Premix. DNA extraction and PCR were completed at the GSC for samples collected in 2021, and the resulting PCR products were exported to the United States at room temperature. However, for the 2022 and 2023 samples, DNA extraction took place at the GSC, and the extracted DNA was exported to the United States in a cooler on ice. PCR amplification and all subsequent laboratory procedures for these samples were carried out in the Department of Biology at the University of North Carolina at Chapel Hill (UNC).
We combined 10 µl of PCR product from each reaction (i.e., both MiFish and Crustacean_16S), resulting in a total of 60 µl per sample. Negative controls were pooled separately by primer set, with 20 µl from each PCR reaction combined to create 60 µl of MiFish PCR control and 60 µl of Crustacean_16S PCR control. All pooled PCR products were cleaned using a 1.5x ratio of Ampure XP beads and eluted in 30 µl of Qiagen Buffer EB. In a second PCR step, we applied a unique dual-indexing strategy by adding IDT for Illumina UDI-UMI indexes to 2.5 µl of the combined and cleaned amplicons.
We then combined 10 µl of each indexed sample and cleaned the combined amplicons using a 0.9x ratio of Ampure XP beads. The 10 nM pooled library was sent to the High-Throughput Sequencing Facility (HTSF) at UNC for paired-end sequencing on a single MiSeq lane using 250 cycles. Throughout all laboratory procedures, we used single-use disposable gloves and all equipment and surfaces were sterilized with a 20% bleach solution before and after use.
We used the same bioinformatics protocol as Ryburn (2025) 32 and Ryburn et al. (2026)33 . In January of 2024, we downloaded all vertebrate sequences (for fish) and all invertebrate sequences (for crustaceans) from the European Molecular Biology Laboratory (EMBL). Based on the distribution of percent sequence similarity scores, we filtered the MiFish sequences to an increased stringency of 98% similarity and Crustacean_16S to 97% similarity in the final dataset.
Statistical analysis
We conducted all statistical analyses in R (version 4.2.2; www.R-project.org) with RStudio (version 2022.12.0 + 353). The map of the sampling locations (Fig. 1) was also created in R using the Galapagos Archipelago shapefiles obtained from the Instituto Oceanográfico y Antártico de la Armada (INOCAR; https://www.inocar.mil.ec/web/). Spatial data were imported and handled using the sf package (version 1.0–16). Map visualization and figure layout were produced with ggplot2 (version 3.5.1), including coastline rendering and spatial layering of geographic features. Sample-based rarefaction curves, which included all identified prey taxa, were created with the vegan package (version 2.6-4; 36) for all blacktip sharks (Fig. 2), male and female sharks (Fig. 3), and sharks sampled during El Niño and La Niña (Fig. 4). We calculated the Levin’s niche width at each shark nursery bay using the spaa package (version 0.2.2; 37; Table 3). To assess dietary differentiation among sharks across the nursery bays, we used a permutational multivariate analysis of variance (perMANOVA) with the adonis2 function in the vegan package36. This analysis used the Jensen-Shannon distance of relative prey read abundance as the metric. Additionally, species richness for each shark was calculated using the phyloseq package (version 1.42.0; 38; Table 3). Statistical analysis R Markdown code is available at https://github.com/sryburn95/Galapagos_Blacktip_Diet.

Rarefaction curve illustrating cumulative prey richness as a function of blacktip sharks sampled. Prey richness includes all 25 classified prey taxa. Figure includes all 107 juvenile blacktips, with the shaded area representing the 95% confidence interval after 100 permutations.

Rarefaction curves illustrating cumulative prey richness as a function of female (A) and male (B) blacktip sharks sampled. Prey richness includes all 25 classified prey taxa. Figure includes all 107 juvenile blacktips, 53 females and 54 males. The shaded area representing the 95% confidence interval after 100 permutations.

Rarefaction curves illustrating cumulative prey richness as a function of blacktip sharks sampled from Rosa Blanca 2 during La Niña (A) and El Niño (B). Prey richness includes 14 classified prey taxa. Figure includes all 13 juvenile blacktips sampled from Rosa Blanca 2, eight during La Niña and five during El Niño. The shaded area representing the 95% confidence interval after 100 permutations.
Results
From the 107 cloacal swabs collected, we identified 989,789 reads of amplicon sequence variants (ASVs), with an average count of 9,029 ± 1,593 (mean ± standard error) per individual shark. A total of 25 prey taxa were identified, including 19 to species, five to genus, and one to class. On average, each juvenile blacktip shark had 3.8 ± 0.2 prey taxa, with the number of taxa per individual ranging from one to eight. The diet of the juvenile blacktip was predominantly composed of fishes (98.8%, based on percent of occurrence), with smaller contributions from crustaceans (0.9%) and a single species of brittle star, Amphipholis squamata (dwarf brittle star; 0.2%). The Thoburn’s mullet (Mugil thoburni) was the most common prey taxon (20.0%) followed by fishes in the genus Abudefduf (sergeant-majors; 13.4%), the Galapagos ringtail damselfish (Stegastes beebei; 12.7%), the scalloped hammerhead shark (Sphyrna lewini; 11.7%), and the yellowtail damselfish (Stegastes arcifrons; 10.7%; Table 2). Among all identified prey, the Thoburn’s mullet was the only prey taxon endemic to the Galapagos. Scalloped hammerhead shark was identified in the diet of the juvenile blacktip shark in all six nursery bays located on San Cristobal, despite Puerto Grande being the only bay on the island classified as a scalloped hammerhead nursery. However, scalloped hammerhead was absent from the diets of the blacktip sharks from all nursery bays on the other three islands (i.e., Isabela, Santiago, and Fernandina), even though juvenile scalloped hammerheads are present in Cartago Bay. The rarefaction curve (Fig. 2) began to reach a plateau, suggesting that the 107 cloacal swabs provided a comprehensive representation of the dietary richness of juvenile blacktip sharks in the Galapagos. Of the 107 sharks, 54 were male and 53 were female. Sex-specific rarefaction curves (Fig. 3) for both males and females also began to plateau, suggesting that sampling effort was sufficient to capture dietary richness for each sex. In addition, we found a statistical difference between male and female juvenile blacktips when comparing the prey composition between sexes (perMANOVA: pseudo F = 2.424, R2 = 0.023, df = 1, P = 0.035).
A statistically significant difference in prey composition was also found when comparing the diets of juvenile blacktip sharks across the different nursery bays (perMANOVA: pseudo F = 2.40, R2 = 0.251, df = 13, P = 0.001). Prey taxa richness was higher in the nursery bays located on San Cristobal compared to all other nursery bay locations (Figs. 5and 6; Table 3). Crustaceans were only found in the diets of the sharks sampled from San Cristobal. The Thoburn’s mullet was present in the diet of the sharks at every nursery bay except Albemarle (Fig. 5).
Rosa Blanca 2 was the only nursery bay where samples were collected across multiple years (i.e., 2021 and 2023). In 2021, the Galapagos experienced a La Niña event and in 2023 an El Niño event. In total, 13 juvenile blacktips were sampled from Rosa Blanca 2, eight during La Niña and five during El Niño. As shown in Fig. 7, the average prey taxa richness for juvenile blacktips caught in Rosa Blanca 2 was higher in 2021, during La Niña conditions, than in 2023, when the archipelago was experiencing an El Niño event. However, neither of the rarefaction curves (Fig. 4) from the sharks sampled from Rosa Blanca 2 during La Niña and El Niño reached an asymptote, suggesting that shark diet richness may be underestimated.

Relative percent of occurrence (POO) of all prey taxa identified in the diets of the 107 juvenile blacktip sharks sampled across the 14 nursery bays. The x-axis represents the individual nursery bays: Puerto Grande, La Seca, Cerro Brujo, Rosa Blanca 1, Rosa Blanca 2, and La Tortuga are located on San Cristobal; Cartago Bay, Albemarle, Grimanesa, Puerto Chino, Baleado, and El Muelle de Balleno are located on Isabela; La Bomba is located on Santiago; and Punta Espinoza is located on Fernandina.

Prey taxa richness across juvenile blacktip shark nursery bays in the Galapagos Archipelago. Colors indicate the island where each bay is located: San Cristobal, Isabela, Santiago, and Fernandina. Taxa richness encompasses all 25 identified prey taxa, and the figure includes all 107 sampled sharks. Grey error bars indicate a 95% confidence interval.

Relationship between capture date and prey for juvenile blacktip sharks sampled at Rosa Blanca 2 on San Cristobal. Data includes all 13 sharks collected from this location. In 2021 there was a La Niña event and in 2023 there was an El Niño event. A) Boxplot comparing capture date, prey taxa richness, and water temperature. Grey error bars indicate a 95% CI. B) Relative percent of occurrence (POO) of all prey taxa identified in the diets of juvenile blacktip sharks across the three different sampling dates.
Discussion
In the Galapagos, juvenile blacktip sharks primarily consumed fishes, particularly Thoburn’s mullet. We found that the diet of juvenile blacktip sharks varied by location; prey taxa richness at the population and individual level was significantly higher on San Cristobal than the other three islands (Table 3). Additionally, diet varied between samples collected at the same location during a La Niña event and those collected during an El Niño event (Fig. 7).
Previous studies focused on the diet of the juvenile blacktip shark relied on morphological stomach content analysis, which frequently resulted in high proportions of unidentified prey (e.g., 99.3% of the Index of Relative Importance [IRI] consisted of unidentified teleosts)11,17,18. In contrast, using DNA metabarcoding of fecal matter, we were able to identify 19 of the 25 prey taxa to the species level (Table 2). The diet of blacktip sharks is known to vary geographically17, likely reflecting differences in prey availability. However, Clupeids and Sciaenids are recorded as being the most common prey consumed by blacktip sharks worldwide17. In our study, we did not detect any Sciaenids in the diet of juvenile blacktips from the Galapagos and observed only a small contribution of Clupeids to their overall diet (1.2% based on percent of occurrence; Table 2). Llerena-Martillo et al. (2018) 65 reported that blacktip shark and Thoburn’s mullet were the two most abundant fish species inhabiting the mangrove bays of Santa Cruz Island in the Galapagos. The ample abundance of Thoburn’s mullet could explain why it had the largest contribution to the diet of the juvenile blacktips in our study. Additionally, Puerto Grande exhibited the largest Levin’s niche width among the 14 sampled bays, followed by Cerro Brujo and La Seca (Table 3). Geographically, La Seca and Cerro Brujo are the closest bays to Puerto Grande among the other 13 sampled locations. Further research on prey species abundance and richness is needed to better understand the ecological significance of Puerto Grande and how its prey composition effects the diet of juvenile blacktip sharks.
Overall, scalloped hammerhead shark was the fourth most common prey species (11.7%; Table 2). Our methodology focused on identifying the presence of scalloped hammerheads in the blacktips’ diet but did not differentiate the age or size of the individuals consumed, nor whether they were actively hunted or scavenged. While it is implausible that juvenile blacktips were hunting adult scalloped hammerheads, it is possible that they targeted juveniles. Juvenile blacktip sharks are known to be highly mobile39 and are thought to utilize areas much larger than the boundaries of individual nursery bays. For instance, juvenile blacktips originally caught in Puerto Grande were documented traveling up to 46.8 km within 35 h9. Puerto Grande was the only bay on San Cristobal where juvenile scalloped hammerheads were reliably observed. However, juvenile blacktips from other nursery bays on San Cristobal could have ventured into Puerto Grande, providing opportunities for them to consume scalloped hammerheads. Further research is needed to better understand the home range and movement patterns of juvenile scalloped hammerheads in the Galapagos40. It remains unclear how far and frequently these juvenile scalloped hammerheads travel from their specific nursery bays, but they likely venture into other bays on the island, increasing their risk of predation. Additionally, neonate scalloped hammerheads are known to have high mortality rates due to malnutrition and starvation41, so it may be more likely that juvenile blacktips scavenged on deceased neonate scalloped hammerheads in or near Puerto Grande.
The role of ENSO
The El Niño Southern Oscillation (ENSO) cycle influences the Galapagos Archipelago in various ways, including shifts in primary productivity and upwelling42. During El Niño events, primary productivity declines dramatically due to increased stratification and reduced upwelling43,44 which has cascading effects on the food web42,45. For example, during the 1997–1998 El Niño, the Galapagos Archipelago had elevated temperatures for over 12 months, resulting in clearer oceanic waters, declines in dissolved nutrients, and reduced phytoplankton productivity46. This prolonged disruption at the base of the marine food chain ultimately led to a decrease in biomass. Most algal species became scarce or declined, leading to mortality and reduced densities of herbivores such as marine iguanas (Amblyrhynchus cristatus) and crabs45. The impacts extended throughout the food web, leading to population declines in higher trophic-level species, including the Galapagos penguins (Spheniscus mendiculus), flightless cormorants (Phalacrocorax harrisi), Galapagos fur seals (Arctocephalus galapagoensis), and Galapagos sea lions (Zalophus wollebaeki)46,47. As food availability declines, many species, such as marine iguanas and Galapagos penguins, face an increased risk of starvation45,48. Given that El Niño events have intensified in the eastern tropical Pacific over the past millennium, with the last 50 years marking an unprecedented period of warming49, such disruptions to marine food webs may become more frequent and severe.
The majority of cloacal swabs (98 out of 107) were collected in 2021 and 2022 during La Niña conditions, however, samples were collected from Rosa Blanca 2 in both 2021 (La Niña) and 2023 (El Niño). We found that prey richness was lower in the samples collected from Rosa Blanca 2 during the El Niño event in 2023 compared to those collected in 2021 during a La Niña event (Fig. 7A). However, this result is derived from a very small sample size and therefore should be interpreted with caution. Previous studies have reported that fish biomass and species richness are greater during La Niña, when upwelling and productivity are much greater50,51. For example, the abundance of Thoburn’s mullet in the Galapagos artisanal fisheries landings decreased during the 1997–1998 El Niño but increased during the 1998–1999 La Niña52. The reduction in prey availability during El Niño events could explain the observed decline in dietary richness among juvenile blacktips in Rosa Blanca 2. Notably, two damselfish species (Galapagos ringtail damselfish and yellowtail damselfish) and scalloped hammerhead sharks were present in the diet of the juvenile blacktips in Rosa Blanca 2 in 2021 but were absent in 2023 (Fig. 7B). Damselfish primarily consume algae53 and since El Niño events reduce algal production and biomass45, damselfish populations could have declined during this period. For example, the Galapagos damselfish (Azurina eupalama) has not been observed in the Galapagos since the 1982 El Niño event and is currently classified as possibly extinct54. Additionally, juvenile scalloped hammerheads are particularly vulnerable to starvation in general55, and their abundance could have been similarly affected by the El Niño event.
Prey richness across the archipelago
Due to variation in temperature, nutrient availability, and primary productivity, fish species richness is lower on the western side of the Galapagos Archipelago3. The region’s heightened productivity and strong upwelling create a unique environment that supports a smaller number of species, favoring those that are well-adapted to these conditions. Additionally, cooler water temperatures in the west make it difficult for many tropical species to survive. Juvenile blacktip sharks could be consuming few prey species in this region simply because there are fewer prey options compared to the more species-rich eastern region (Fig. 6). Water temperature in the Galapagos varies considerably due to multiple factors, including ENSO events, upwelling, ocean currents, and seasonal fluctuations56,57,58. However, the western region is characterized by a temperate climate with colder waters that support more Peruvian fish species, while the eastern region has a more tropical climate, favoring Panamic and Indo-Pacific species3. The strong regional divisions in marine fauna are a reflection of local environment as well as connectivity of larval propagules with external source regions3. Fish species with Indo-Pacific distributions likely maintain gene flow across the East Pacific Barrier, contributing to higher species richness in the region3. In contrast, populations of Panamic and Peruvian species in the Galapagos are likely self-sustaining, with Panamic species experiencing intermittent recruitment from the mainland, while Peruvian species persist without reliance on larval input from the continent3. The environmental conditions in the western Galapagos resemble those of central South America, with cooler temperatures and high productivity59. As a result, Peruvian fish species may have a stronger affinity for this distinct region of the archipelago3. Conversely, tropical species could be intolerant of the cooler waters of the western Galapagos, thereby reducing the number of potential prey species available to juvenile blacktips residing in that area.
Conservation and conclusions
Mangrove-fringed bays are not only critical habitats for juvenile blacktip sharks but also serve as vital fishing grounds for artisanal fishermen in the Galapagos, where they catch mullet and bait for offshore fishing52. Mullet, specifically Thoburn’s mullet and Flathead grey mullet (Mugil cephalus), are the most important target species of the artisanal gillnet fisheries operating within the Galapagos Marine Reserve (GMR)60,61. These two species accounted for 30% of the total fish biomass landed in the GMR from 1998 to 200662. Thoburn’s mullet, an endemic species to the Galapagos Archipelago, also accounted for the largest percentage of prey found in the blacktip’s diet (Table 2). Sustained fishing pressure can lead to declines in species density63 and diversity64, making it imperative to prevent overfishing of these mangrove bays and, in particular, Thoburn’s mullet. Protecting these ecosystems is crucial to maintaining both the ecological balance and the livelihoods of local fishers.
In conclusion, our findings indicate that location and the ENSO cycle had a considerable effect on the diet of the juvenile blacktip in the Galapagos, likely due to their effects on prey availability. These results are in concordance with previous research identifying water temperature as the most important environmental variable driving variations in fish abundance within the mangrove bays of the Galapagos65. For example, Thoburn’s mullet, a critical prey species for juvenile blacktips, has been found to be more abundant in the cold season than the warm. With rising sea surface temperatures due to climate change, this could negatively affect the abundance of one of the juvenile blacktip’s primary prey species. Many shark species are often categorized as generalist predators66. However, using metabarcoding techniques on non-lethal fecal samples to expand our understanding of shark diets can improve our ability to predict how disruptions in food web structure and prey availability, particularly due to overfishing and climate change, may impact shark survival and conservation efforts.
Data availability
The raw data is available at Figshare: 10.6084/m9.figshare.30359203.
References
Iriarte, J. A., Franklin, W. L., Johnson, W. E. & Redford, K. H. Biogeographic variation of food habits and body size of the America Puma. Oecologia 85, 185–190 (1990).
Roffler, G. H., Allen, J. M., Massey, A. & Levi, T. Metabarcoding of fecal DNA shows dietary diversification in wolves substitutes for ungulates in an Island Archipelago. Ecosphere 12, e03297 (2021).
Edgar, G. J., Banks, S., Fariña, J. M., Calvopiña, M. & Martínez, C. Regional biogeography of shallow reef fish and macro-invertebrate communities in the Galapagos Archipelago. J. Biogeogr. 31, 1107–1124 (2004).
Fierro-Arcos, D. et al. Mangrove fish assemblages reflect the environmental diversity of the Galapagos Islands. Mar. Ecol. Prog Ser. 664, 183–205 (2021).
Urquía, D. O. et al. DNA -metabarcoding supports trophic flexibility and reveals new prey species for the Galapagos sea Lion. Ecol. Evol. 14, e10921 (2024).
Simpfendorfer, C. A. & Milward, N. E. Utilisation of a tropical Bay as a nursery area by sharks of the families Carcharhinidae and Sphyrnidae. Environ. Biol. Fish. 37, 337–345 (1993).
Heupel, M., Carlson, J. & Simpfendorfer, C. Shark nursery areas: concepts, definition, characterization and assumptions. Mar. Ecol. Prog Ser. 337, 287–297 (2007).
Matich, P. et al. Delineation of Blacktip shark (Carcharhinus limbatus) nursery habitats in the north-western Gulf of Mexico. J. Fish Biol. 101, 236–248 (2022).
Llerena, Y. et al. Nursery grounds of Blacktip sharks (Carcharhinus limbatus) in mangrove-fringed Bays in the central part of the Galapagos Archipelago. (2015).
Springer, S. Social organization of shark population. Sharks Skate Rays 149–174 (1967).
Heupel, M. R. & Hueter, R. E. Importance of prey density in relation to the movement patterns of juvenile Blacktip sharks (Carcharhinus limbatus) within a coastal nursery area. Mar. Freshw. Res. 53, 543 (2002).
IUCN et al. The IUCN Red List of Threatened Species 2021: e.T3851A2870736. (2020). https://doi.org/10.2305/IUCN.UK.2021-2.RLTS.T3851A2870736.en
Killam, K. A. & Parsons, G. R. Age and Growth of the Blacktip Shark, Carcharhinus Limbatus, Near Tampa Bay, Florida 87 (Fishery Bulletin, 1989).
Branstetter, S. Age and growth estimates for Blacktip, carcharhinus limbatus, and Spinner, C. brevipinna, sharks from the Northwestern Gulf of Mexico. Copeia 964, 1987 (1987).
The Galapagos Marine Reserve: A Dynamic Social-Ecological System. (Springer International Publishing, Cham, (2014). https://doi.org/10.1007/978-3-319-02769-2
Goodman, L. S. Feasability of New Methods for Nursery Ground Identification for the Juvenile Blacktip Shark (Carcharhinus limbatus) Using Drone Technology – San Cristobal Island, Galapagos. ProQuest Dissertations and Theses (The University of North Carolina at Chapel Hill, United States -- North Carolina, (2020).
Matich, P., Plumlee, J. D., Weideli, O. C. & Fisher, M. New insights into the trophic ecology of Blacktip sharks (Carcharhinus limbatus) from a subtropical estuary in the Western Gulf of Mexico. J. Fish Biol. 98, 470–484 (2021).
Tavares, R. & Occurrence Diet and growth of juvenile Blacktip Sharks, Carcharhinus limbatus, from Los Roques Archipelago National Park, Venezuela. Caribb. J. Sci. 44, 291–302 (2008).
Barry, K. & College Feeding habits of blacktip sharks, Carcharhinus limbatus, and Atlantic sharpnose sharks, Rhizoprionodon terraenovae, in Louisiana coastal waters. (Louisiana State University and Agricultural and Mechanical doi:https://doi.org/10.31390/gradschool_theses.66. (2002).
Hoffmayer, E. R., Parsons, G. R., FOOD HABITS OF THREE & SHARK SPECIES FROM THE MISSISSIPPI SOUND IN THE NORTHERN GULF OF MEXICO. Southeast. Nat. 2, 271–280 (2003).
Bethea, D., Buckel, J. & Carlson, J. Foraging ecology of the early life stages of four sympatric shark species. Mar. Ecol. Prog Ser. 268, 245–264 (2004).
Barry, K. P., Condrey, R. E., Driggers, W. B. & Jones, C. M. Feeding ecology and growth of neonate and juvenile Blacktip sharks Carcharhinus limbatus in the Timbalier–Terrebone Bay complex, LA, U.S.A. J. Fish Biol. 73, 650–662 (2008).
Plumlee, J. & Wells, R. Feeding ecology of three coastal shark species in the Northwest Gulf of Mexico. Mar. Ecol. Prog Ser. 550, 163–174 (2016).
Páez-Rosas, D. et al. Trophic variation during the early stages of Blacktip sharks (Carcharhinus limbatus) within coastal nurseries of the Galapagos marine reserve. J. Sea Res. 170, 102023 (2021).
Ford, M. J. et al. Estimation of a killer Whale (Orcinus orca) population’s diet using sequencing analysis of DNA from feces. PLoS ONE. 11, e0144956 (2016).
Berry, T. E. et al. DNA metabarcoding for diet analysis and biodiversity: A case study using the endangered Australian sea Lion (Neophoca cinerea). Ecol. Evol. 7, 5435–5453 (2017).
Anslan, S. et al. DNA metabarcoding reveals fine scale geographical differences of consumed algae in the Galápagos marine iguanas (Amblyrhynchus cristatus). Amphib -Reptilia. 42, 471–480 (2021).
Snider, A. M., Bonisoli-Alquati, A., Pérez-Umphrey, A. A., Stouffer, P. C. & Taylor, S. S. Metabarcoding of stomach contents and fecal samples provide similar insights about seaside sparrow diet. Ornithological Appl. 124, duab060 (2022).
van Zinnicq Bergmann, M. P. M. et al. Elucidating shark diets with DNA metabarcoding from cloacal swabs. Mol. Ecol. Resour. 21, 1056–1067 (2021).
Clark, Z. S. R. et al. Insights into the diet and trophic ecology of white sharks (Carcharodon carcharias) gained through DNA metabarcoding analyses of cloacal swabs. Environ. DNA. https://doi.org/10.1002/edn3.454 (2023). edn3.454.
Olin, J. A. et al. DNA metabarcoding of cloacal swabs provides insight into diets of highly migratory sharks in the Mid-Atlantic bight. Journal Fish. Biology Jfb. 15543 https://doi.org/10.1111/jfb.15543 (2023).
Ryburn, S. J. Elucidating the Diet of Sharks Through DNA Analysis of Fecal Matter. Doctoral Dissertation, The University of North Carolina at Chapel Hill. (2025)
Ryburn, S. J. et al. A comparison of DNA metabarcoding cloacal swabs and stomach contents for shark diet reconstruction. Environmental DNA. https://doi.org/10.1002/edn3.70247 (2026).
Wisely, E. Metabarcoding for fish and crustaceans in diet samples using 2-PCR protocol with unique dual indexing v2. Preprint at (2025). https://doi.org/10.17504/protocols.io.261ge5dnwg47/v2
Miya, M. et al. MiFish, a set of universal PCR primers for metabarcoding environmental DNA from fishes: detection of more than 230 subtropical marine species. R Soc. open. sci. 2, 150088 (2015).
Oksanen, J. et al. vegan: Community Ecology Package. 2.6–6.1 (2001). https://doi.org/10.32614/CRAN.package.vegan
Zhang, J. SPecies Association Analysis. (2022).
McMurdie, P. J. & Holmes, S. Phyloseq: an R package for reproducible interactive analysis and graphics of Microbiome census data. PLoS ONE. 8, e61217 (2013).
Heupel, M. R., Simpfendorfer, C. A. & Hueter, R. E. Running before the storm: Blacktip sharks respond to falling barometric pressure associated with tropical storm gabrielle: response of sharks to a tropical storm. J. Fish Biol. 63, 1357–1363 (2003).
Chiriboga-Paredes, Y. et al. Discovery of a putative scalloped hammerhead shark Sphyrna Lewini (Carcharhiniformes: Sphyrnidae) nursery site at the Galapagos Islands, Eastern tropical Pacific. Environ. Biol. Fish. https://doi.org/10.1007/s10641-021-01207-3 (2022).
Duncan, K. & Holland, K. Habitat use, growth rates and dispersal patterns of juvenile scalloped hammerhead sharks Sphyrna Lewini in a nursery habitat. Mar. Ecol. Prog Ser. 312, 211–221 (2006).
Eddy, T. D. & Friedlander, A. M. Salinas de León, P. Ecosystem effects of fishing & El Niño at the Galápagos marine reserve. PeerJ 7, e6878 (2019).
Gluschankoff, N., Santoro, A. E., Buchwald, C. & Casciotti, K. L. Shifts in the Isotopic Composition of Nitrous Oxide Between El Niño and La Niña in the Eastern Tropical South Pacific. Global Biogeochemical Cycles 37, e2023GB007959 (2023).
Hernández-Moreno, Y. et al. Phytoplankton biomass in a Ramsar-listed coastal lagoon of the Southern Gulf of Mexico during La Niña 2022 and El Niño 2023 events. Lat Am. J. Aquat. Res. 52, 845–856 (2024).
Vinueza, L. R., Branch, G. M., Branch, M. L. & Bustamante, R. H. Top-down herbivory and bottom-up el niño effects on galápagos rocky-shore communities. Ecol. Monogr. 76, 111–131 (2006).
Robinson, G. & Del Pino. El Niño in the Galápagos Islands: the 1982– 1983 Event. (1985).
Trillmich, F. & Limberger, D. Drastic effects of El Nifio on Galapagos pinnipeds. (1985).
Boersma, P. D. Population trends of the Galápagos penguin: impacts of El Niño and La Niña. Condor 100, 245–253 (1998).
Conroy, J. L. et al. Unprecedented recent warming of surface temperatures in the Eastern tropical Pacific ocean. Nat. Geosci. 2, 46–50 (2009).
Godı́nez-Domı́nguez, E., Rojo-Vázquez, J., Galván-Piña, V. & Aguilar-Palomino, B. Changes in the structure of a coastal fish assemblage exploited by a small scale Gillnet fishery during an El Niño–La Niña event. Estuar. Coast. Shelf Sci. 51, 773–787 (2000).
Rastoin-Laplane, E. et al. Fluctuations of Galapagos mid-water and benthic reef fish populations during the 2015–16 ENSO. Estuar. Coast. Shelf Sci. 294, 108523 (2023).
Murillo, J. C. & Bautil, B. Evaluación de Las pesquerías En La reserva Marina de Galápagos. Informe Compendio. 2002, 124 (2002).
DeVaan, I., Koch, B., Goodman, P. & Madden, C. Damselfish and their Effect on the Biodiversity of Algal Species: A Continuation of Research in the Galápagos Islands. (2016).
Grove, J. S. & Victor, B. C. Has climate change driven the Galapagos Damselfish, Azurina eupalama, to extinction? Preprint at (2025). https://doi.org/10.5281/ZENODO.14846312
Lowe, C. G. Bioenergetics of free-ranging juvenile scalloped hammerhead sharks (Sphyrna lewini) in Kāne’ohe Bay, Ō’ahu, HI. J. Exp. Mar. Biol. Ecol. 278, 141–156 (2002).
Houvenaghel, G. T. Equatorial undercurrent and climate in the Galapagos Islands. Nature 250, 565–566 (1974).
Houvenaghel, G. Oceanographic setting of the Galapagos Islands. Key environments: Galapagos. Pergamon Press. Oxford 43–54 (1984).
Wellington, G. M., Strong, A. E., Merlen, G., Sea surface temperature variation in the galápagos archipelago: a comparison between avhrr nighttime satellite data and in situ instrumentation (1982–1998). Bulletin of marine science 69, 27–42 (2001).
Banks, S. J. Ambiente Fısico. Reserva Marina de Galápagos, Línea base de La Biodiversidad. Charles Darwin Foundation Galapagos Natl. Park Service Galapagos Ecuador 18–33 (2002).
IATTC (Inter American Tropical Tuna Comission). Inter American Tropical Tuna Comission: La Jolla, California Annual Report of the year 1955. (1956).
Reck, G. K. The coastal fisheries in the Galápagos Islands, Ecuador. Description and consequences for management in the context of marine environmental protection and regional development. (Doctoral Thesis) Kiel University, Kiel, Germany, (1983).
Peñaherrera-Palma, C. Variaciones espacio-temporales de los ensambles de peces de la Reserva Marina de Galápagos basados en registros pesqueros. (Bachelor Thesis) Pontificia Universidad Católica del Ecuador, Ecuador, (2007).
Mangi, S. C. & Roberts, C. M. Quantifying the environmental impacts of artisanal fishing gear on kenya’s coral reef ecosystems. Mar. Pollut. Bull. 52, 1646–1660 (2006).
Hall, S. The Effects of Fishing on Marine Ecosystems and Communities (Blackwell Science Ltd Oxford, U.K., 1999).
Llerena-Martillo, Y., Peñaherrera-Palma, C. & Espinoza, E. R. Fish assemblages in three fringed mangrove bays of Santa Cruz Island, Galapagos Marine Reserve. RBT 66, 674 (2018).
Munroe, S. E. M., Simpfendorfer, C. A. & Heupel, M. R. Defining shark ecological specialisation: concepts, context, and examples. Rev. Fish. Biol. Fisheries. 24, 317–331 (2014).
van Zinnicq Bergmann P. M., Bautisse D., Postaire Kirk, Gastrich Michael R., Heithaus Lisa A., Hoopes Kady, Lyons Yannis P., Papastamatiou Eric V. C., Schneider Bradley A., Strickland Brendan S., Talwar Demian D., Chapman Judith, Bakker (2021) Elucidating shark diets with DNA metabarcoding from cloacal swabs Abstract Molecular Ecology Resources 21(4) 1056-1067 10.1111/men.v21.4 10.1111/1755-0998.13315
Acknowledgements
We thank Isabel Silva, Esteban Agudo, Salomé Jaramillo Gil, Mazarine Bruno, Kelly Ong, Emma Rudy Srebnik, Caroline Parker, Sydney Mantell, and Alex Reulbach for their help with the fieldwork and sampling. We also thank our boat captains, Yuri Revelo and Manuel Yépez. Animal care and use were overseen by UNC-CH’s IACUC (23-210.0). Fieldwork and data collection were performed under research permits PC-13-21, PC-18-22, PC-13-23, and MAATE-DBI-CM-2021-0174 granted by the Galapagos National Park and the Ecuadorian Ministry of Environment.
Funding
This study was funded in part by the National Science Foundation (OCE #2128592 to JFB), UNC E3P Graduate Student Research Grant (SJR), the Galapagos Conservancy (EE), and the Save Our Seas Foundation (EE).
Author information
Authors and Affiliations
Contributions
*SJR* and *EW* designed the study. *SJR* , *EW* , *EE* , *DA* , *HEC* , and *JFB* collected the samples. *DAP* and *AH* provided the permits. *SJR* and *EW* completed the DNA analysis. *SJR* analyzed the data. The manuscript was written by *SJR* with help from *JFB* . All authors revised the manuscript and gave final approval for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ryburn, S.J., Wisely, E., Espinoza, E. et al. Biogeographic variation in the diet of juvenile blacktip sharks across the Galapagos Archipelago. Sci Rep 16, 9110 (2026). https://doi.org/10.1038/s41598-026-37386-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-37386-1
