
Abstract
Tomato (Solanum lycopersicum L.) is one of the most widely cultivated vegetable crops globally. Still, its productivity is significantly constrained by tomato leaf curl virus (ToLCV), a devastating begomovirus transmitted by whiteflies. This study examined the diversity and plant growth-promoting potential of culturable endophytes associated with tomato cultivars differing in resistance to tomato leaf curl virus (ToLCV). A total of 59 fungal and bacterial endophytes were isolated. Resistant cultivars (Nandi, Sankranthi, and Vybhav) harboured more diverse and compositionally distinct communities than the susceptible cultivar Arka Vikas, as indicated by Shannon, Simpson, and Chao-1 indices and multivariate analyses. Several isolates, particularly from the genera Xylaria, Fusarium, Arcopilus, Epicoccum, Bacillus, Pseudomonas, Stutzerimonas, and Paenibacillus, displayed strong nutrient-solubilizing traits in vitro, highlighting their potential as plant growth-promoting candidates. Eleven promising isolates were further evaluated on the susceptible cultivar Arka Vikas. At 30 days after sowing, Epicoccum nigrum and Bacillus subtilis significantly increased seedling height, biomass, and leaf number relative to the control. Overall, the study reveals that resistant cultivars are associated with greater culturable endophyte diversity and identifies several isolates with strong potential for promoting plant growth. Future research should assess the antiviral potential of these endophytes under ToLCV challenge and employ metagenomic studies to elucidate their functional roles in enhancing plant health.
Similar content being viewed by others

Introduction
Tomato (Solanum lycopersicum L.) is one of the most important vegetable crops worldwide, valued for its nutritional, economic, and industrial significance. In India, its cultivation faces major challenges due to various biotic stresses, among which the tomato leaf curl virus (ToLCV) is particularly devastating. Tomato leaf curl virus (ToLCV), a monopartite or bipartite begomovirus transmitted persistently by the whitefly Bemisia tabaci1,2,3 causes tomato leaf curl disease (ToLCD), which manifests as leaf curling, vein thickening, stunted growth, and flower drop, often resulting in yield losses ranging from 30 to 100%4,5,6 especially when infection occurs at early stages7,8.
Traditionally, the management of ToLCV has primarily relied on resistant cultivars, vector control, and cultural practices. The development and deployment of ToLCV-resistant tomato cultivars have been a major strategy to manage the disease. Earlier screening of Lycopersicon cultivars and wild accessions also revealed natural sources of resistance that have since been exploited in breeding programs1,9. Breeding efforts have led to the identification of resistant lines and hybrids that exhibit significant resistance to ToLCV under field conditions10. However, the frequent emergence of new viral strains and vector biotypes complicates long-term resistance3,10,11. This highlights the need to explore complementary strategies for sustainable disease management. Beyond host genetics, there is increasing recognition of the plant microbiome, especially endophytic microbial communities, in modulating plant health and resistance.
Endophytes (fungi and bacteria) that reside within plant tissues without causing disease play a crucial role in enhancing plant tolerance to biotic and abiotic stresses12. They are known to produce antimicrobial compounds, stimulate induced systemic resistance (ISR), and improve nutrient uptake and growth via plant growth-promoting traits, phosphate solubilization, and siderophore secretion13,14,15. Recent studies suggest that the composition and functional diversity of endophytes can vary significantly between resistant and susceptible plant genotypes, likely due to differences in host metabolites and immune signaling pathways16 and recent advances in plant microbiome research propose that plants and their associated microbes function as an integrated unit, or holobiont. Within this framework, host genotype influences both the composition and activity of microbial communities, leading resistant and susceptible cultivars to assemble distinct microbiomes. Such selective assembly can affect nutrient acquisition, growth, stress tolerance, and interactions with pathogens. Consequently, breeding for particular traits may inadvertently shape the associated microbiome, linking plant genetics with microbial community structure and function. This emerging concept emphasizes that future strategies could deliberately integrate host–microbiome interactions, enabling the development of crops that harness both genetic resistance and microbiome-mediated resilience17. For instance, tomato cultivars resistant to Ralstonia solanacearum were found to harbour more antagonistic and functionally diverse bacterial endophytes than susceptible ones14. Similarly, genotypic differences in microbial recruitment were observed in rice and maize under pathogen pressure, influencing disease outcomes18. Despite progress in understanding plant–microbe interactions, information on the diversity and functional potential of endophytes in ToLCV-resistant tomato cultivars remains limited. In this study, we isolated and characterized fungal and bacterial culturable endophytes from three resistant cultivars (Nandi, Sankranthi, and Vybhav) and compared them with a susceptible cultivar (Arka Vikas)8,10 to assess their diversity. Furthermore, the plant growth-promoting traits of the isolated endophytes were evaluated for their ability to enhance growth and development, exploring their role in plant growth promotion and potential biocontrol applications.
Materials and methods
Collection and Raising of tomato cultivars
ToLCV-resistant tomato cultivars Nandi (TLB-130), Sankranthi (TLB-111), and Vybhav (TLB-182) and one susceptible cultivar (Arka Vikas) were selected for endophyte isolation. Certified seeds of Nandi, Sankrathi and Vybhav were obtained from the Department of Plant Pathology, University of Agricultural Sciences (UAS), Bangalore, and Arka Vikas from the Indian Institute of Horticultural Research (IIHR), Bengaluru. Prior to sowing, seeds were surface sterilized using 70% ethanol for 1 min, followed by treatment with 1% sodium hypochlorite for 1 min, and rinsed thoroughly five times with sterile distilled water. Sterilized seeds were then sown in autoclaved soil and maintained under greenhouse conditions at a temperature of 35 ± 2 °C, 12 h photoperiod, light intensity of approximately 250–300 µmol m⁻² s⁻¹. Plants were grown for 30–40 days until sampling.
Sample collection and surface sterilization
Healthy, asymptomatic tissues (roots, stems and leaves) were collected from 40-day-old tomato plants. Three plants per genotype were sampled, the plants were collected. Later, the plants were carried in individual polybags to the laboratory and washed under running tap water to remove adhering soil and debris. Under aseptic conditions inside a laminar air flow cabinet, surface sterilization was performed to eliminate epiphytic microbes. Plant tissues were treated with 70% ethanol for 1 min, followed by 1% sodium hypochlorite for 30 s and then treated with 70% ethanol for 30 s. Further, tissues were rinsed thoroughly three times with sterile distilled water. To verify the efficacy of surface sterilization, 100 µl from the final rinse water was plated and some tissue segments were imprinted on potato dextrose agar (PDA) and nutrient agar (NA) and incubated at 24–28 °C for 3–7 days for fungal growth and 30–34 °C for 24–48 h for bacterial growth. Plates showing no microbial growth after 3–5 days were considered sterile, confirming the effectiveness of the surface sterilization procedure19.
Isolation of endophytes
For isolation of fungal endophytes, sterile plant tissues (leaf, stem and roots) were aseptically made into segments (1 cm × 1 cm) and placed on potato dextrose agar (PDA), rose bengal agar (RBA) and Richard’s synthetic agar (RSA). For bacterial isolation, plant tissues were macerated in 1 mL sterile potassium phosphate buffer (pH 7.2) using a sterilized pestle and mortar. The resulting suspensions were serially diluted up to 10− 5 and aliquots from appropriate dilutions were plated on nutrient agar (NA), Luria-Bertani agar (LBA) and King’s B medium (KBM). Plates were incubated at 24–28 °C for 3–7 days for fungal growth and 30–34 °C for 24–48 h for bacterial growth. Morphologically distinct colonies were subcultured within 2 days of fungus and bacterial growth to obtain a pure culture for further use20. A lab-wide culture tracking system with unique alphanumeric IDs for every isolate and subculture was implemented. IDs encode: tomato genotype code, part of plant, endophyte code and isolate sequence number (format: tomato genotype code (G-Nandi, H-Sankrathi, I-Vybhav, and V-Arka Vikas); Part of plant (L-Leaf, S-Stem, and R-Root); endophyte code (F-fungus, B-Bacteria); Numerical isolate sequence (1,2,3,….). Three individual plants per cultivar were sampled, and each plant was treated as an independent biological replicate. Leaf, stem, and root tissues from each plant were processed separately, all isolations were performed with three biological replicates per cultivar.
Morphological and molecular characterization
All endophytic isolates were preliminarily characterised based on colony morphology. Fungal isolates were identified based on colony characteristics like colony colour, pigmentation, form, elevation, margin, surface texture and growth rate and microscopic examination, while bacterial isolates were assessed for colony characteristics like colony color, form, elevation, margin, surface and Gram reaction. For molecular identification, genomic DNA was extracted from fungal and bacterial isolates using the CTAB protocol21,22. DNA quality and concentration were confirmed using agarose gel electrophoresis and a Nanodrop. PCR amplification of the internal transcribed spacer (ITS) region was performed for fungal DNA using ITS-1 (5’-TCCGTAGGTGAACCTGCGG-3’) and ITS-4 (5’-TCCTCCGCTTATTGATATGC-3’) primers23, while the 16 S rDNA gene was amplified for bacterial isolates using universal primers 27 F 5’-AGAGTTTGATCCTGGCTCAG-3’ and 1492R 5’-TACGGYTACCTTGTTACGACTT-3’)24. Amplification was performed in 20 µl reaction mixtures containing 2 µl of DNA template, 1 µl of each primer, 6 µl of double-distilled water (ddH2O), and 10 µl of master mix (TAKARA®, India). The thermal cycling conditions were as follows for ITS primer : initial denaturation at 95 °C for 5 min, followed by 35 cycles consisting of denaturation at 95 °C for 60 s, annealing at 60 °C for 90 s, extension at 72 °C for 60 s, and a final extension at 72 °C for 7 min22, for 16 S rDNA Primer: initial denaturation at 95 °C for 5 min, followed by 35 cycles consisting of denaturation at 95 °C for 60 s, annealing at 57 °C for 90 s, extension at 72 °C for 60 s, and a final extension at 72 °C for 10 min24. PCR products were visualized on a 1% agarose gel, purified, and sequenced commercially at Barcode Bioscience Limited, Bengaluru. BLASTn searches were conducted against the NCBI GenBank database to identify isolates based on sequence similarity. The sequences were submitted to the NCBI GenBank and obtained accession numbers.
Analysis of diversity indices
The diversity of endophytic communities in the resistant and susceptible tomato cultivars was analyzed using alpha diversity indices, such as the Dominance (D), Simpson (1 − D), Shannon (H’), Evenness (eH/S), Chao-1, and Bray–Curtis dissimilarity index. These indices were computed using PAST software25,26. Venn diagrams were created to determine the shared and unique taxa among the four cultivars27,28. Additionally, tornado graph, heatmaps and hierarchical cluster analysis, as well as principal component analysis (PCA), were constructed using R software (Packages: VennDiagram, stats, pheatmap, vegan and ggplot) to visualise and compare community structures based on presence/absence and relative abundance data29. Diversity analyses were performed at the plant (replicate) level rather than at the colony level, thereby avoiding pseudoreplication.
In vitro plant growth-promoting activity assays
Representative endophytes from each genotype were screened for plant growth-promoting traits for their ability to solubilize phosphorus (P), potassium (K), and zinc (Zn), using standard in vitro methods. Phosphorus solubilization was assessed on Pikovskaya’s agar medium30, potassium solubilization on Aleksandrow medium31, and zinc solubilization using a modified minimal salt medium supplemented with zinc oxide (ZnO) as the insoluble Zn source32. For fungal isolates, 5 mm diameter mycelial plugs from actively growing cultures were aseptically placed onto the respective media plates. All plates were incubated at 28 ± 2 °C for 7 days in dark conditions15. Bacterial isolates were pre-cultured in nutrient broth at 30 °C and 120 rpm for 24 h33. A 5 µl aliquot of each culture was then spot-inoculated onto the respective media and incubated at 30 °C for three days. The appearance of solubilization zones around the colonies or mycelial plugs was indicative of nutrient solubilization (P, K, Zn).
In-planta evaluation of plant growth-promoting (PGP) traits
To evaluate the growth-promoting potential of the endophytes, tomato seeds of Arka Vikas were surface sterilized with 70% ethanol, then rinsed with sterile double-distilled water thrice and treated with respective fungal spores (1 × 10⁸ spores/mL) or bacterial suspensions (1 × 10⁸ CFU/mL) by soaking for 24 h15,33. Treated seeds were sown in sterile soil under greenhouse conditions. Treatment details were mentioned in Supplementary Table 1. Each treatment was carried out with three replicates and each replicate consisted of ten plants. Pots were arranged in a completely randomized design (CRD) inside the greenhouse to minimize positional bias. Plants were irrigated regularly with sterile distilled water to maintain adequate soil moisture, and no fertilizers were applied during the experimental period to ensure that the observed effects were solely due to the endophyte treatments. Growth parameters such as plant height and fresh weight of root and shoot (g/plant), the number of branches and leaves were recorded 30 and 60 days after sowing (DAS). Untreated seeds served as the control. Furthermore, In-Planta data were analyzed using ANOVA models in which treatment was specified as a fixed factor and replicate as a random factor, followed by Tukey’s range test at a significance level of p ≤ 0.05, using OPISTAT software.
Results
Isolation of endophytes
Endophytic microorganisms were successfully isolated from surface-sterilized root, stem, and leaf tissues of all four tomato genotypes, three ToLCV-resistant tomato cultivars (Nandi, Sankranthi and Vybhav) and the susceptible cultivar Arka Vikas (Fig. 1). Among the ToLCV-resistant cultivars, both Sankranthi (9 fungal and 8 bacterial isolates) and Vybhav (8 fungal and 9 bacterial isolates) cultivars harbored the highest number of endophytes (n = 17 each) and Nandi (n = 15, comprising 8 fungal and 7 bacterial isolates). The ToLCV-susceptible cultivar, Arka Vikas, yielded 10 endophytes (6 fungal and 4 bacterial isolates), resulting in a total of 59 isolates across all four cultivars (Tables 1 and 2). The epiphytic confirmation test showed no microbial growth on the imprints and rinse water, indicating the absence of surface contamination and confirming that the surface sterilization procedure was effective.

ToLCV-resistant tomato cultivars. (a) Nandi (TLB-130), (b) Sankranthi (TLB-111) (c) Vybhav (TLB-182) and ToLCV-susceptible tomato cultivar (d) Arka Vikas.
The diversity of culturable endophytic communities isolated from different plant parts of three ToLCV-resistant tomato cultivars (Nandi, Sankranthi and Vybhav) and the susceptible cultivar Arka Vikas revealed distinct differences in both fungal and bacterial endophyte composition and tissue-specific colonization. Across all cultivars, leaf tissues contributed the highest number of fungal isolates (41.9%), followed by stems (32.3%) and roots (25.8%), while for bacterial endophytes, stems harbored the highest number of isolates (50.0%), followed by roots (42.9%) and leaves (7.1%) indicating tissue-specific preferences of endophyte colonization (Fig. 2).

Tissue-specific colonization of fungal and bacterial endophytes in tomato genotypes. A tornado chart illustrating the percentage distribution of fungal (blue, left) and bacterial (red, right) endophytes isolated from different plant parts of tomato cultivars.
Morphological and molecular identification
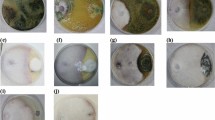
Morphological and molecular identification based on sequencing of the internal transcribed spacer (ITS) region for fungi and 16 S rDNA for bacteria confirmed morphological identifications (Tables 3 and 4). Morphological characterization of culturable endophytes from tomato cultivars (Nandi, Sankranthi, Vybhav, and Arka Vikas) revealed high diversity among 31 fungal and 28 bacterial isolates (Figs. 3, 4 and 5). Fungal colonies displayed varied colours (blackish grey to whitish purple), pigmentation (none to red, black, yellow-brown), and textures (fluffy, woolly, velvety), with forms ranging from irregular to circular and growth rates from medium to very fast (Table 1). Dominant fungal genera included Chaetomium, Fusarium, Xylaria, Ovatospora, and Arcopilus. Bacterial isolates varied in colour (white to yellow), shape, elevation, and margin, with both Gram-positive (e.g., Bacillus, Paenibacillus) and Gram-negative (e.g., Pseudomonas, Enterobacter) genera as dominant genera (Table 2). The observed morphological traits suggest a diverse and rich endophytic community in tomato plants.

Fungal endophytes isolated from ToLCV-resistant tomato cultivars (Nandi, Snakranthi and Vybhav) (a) GLF1, (b) GLF2, (c) GLF3, (d) GLF4, (e) GSF1, (f) GSF2, (g) GRF1, (h) GRF2, (i) HLF1, (j) HLF2, (k) HLF3, (l) HLF4, (m) HSF1, (n) HSF2, (o) HSF3, (p) HSF4, (q) HRF1, (r) ILF1, (s) ILF2, (t) ILF3, (u) ILF4, (v) ISF1, (w) ISF2, (x) IRF1, (y) IRF2. G - Nandi, H - Sankranthi, I - Vybhav, L - Leaf, S - Stem, R - Root, F - Fungal endophyte.

Bacterial endophytes isolated from ToLCV-resistant tomato cultivars (Nandi, Sankranthi and Vybhav) (a) GSB1, (b) GSB2, (c) GSB3, (d) GSB4, (e) GRB1, (f) GRB2, (g) GRB3, (h) HLB1, (i) HSB1, (j) HSB2, (k) HSB3, (l) HSB4, (m) HRB1, (n) HRB2, (o) HRB3, (p) ILB1, (q) ISB1, (r) ISB2, (s) ISB3, (t) ISB4, (u) IRB1, (v) IRB2, (w) IRB3, (x) IRB4. G - Nandi, H - Sankranthi, I - Vybhav, L - Leaf, S - Stem, R - Root, B- Bacterial endophyte.

Endophytes isolated from ToLCV-susceptible tomato cultivars. Fungal Endophytes: (a) VLF1, (b) VSF1, (c) VSF2, (d) VRF1, (e) VRF2, (f) VRF3. Bacterial endophytes (g) VSB1, (h) VSB2, (i) VRB1, (j) VRB2. V- Arka Vikas, L - Leaf, S - Stem, R - Root, F- Fungal Endophyte, B- Bacterial endophyte.
For Molecular identification fungal isolates showing ≥ 97% sequence similarity in the ITS region and bacterial isolates showing ≥ 98% sequence similarity in the 16 S rDNA region with reference sequences in the NCBI GenBank database were considered. Further molecular identification revealed that those fungal isolates were distributed among Chaetomium, Fusarium, Xylaria, Nigrospora, Arcopilus, Ovatospora, Pestalotiopsis, Peroneutypa, Epicoccum, and Alternaria, with Chaetomium globosum being the most frequently isolated species. Bacterial endophytes were identified as members of Bacillus, Paenibacillus, Pseudomonas, Enterobacter, Neobacillus, Klebsiella, Kosakonia, Stutzerimonas, Peribacillus, and Cytobacillus. Sequencing Data were submitted to NCBI GenBank, and accession numbers are provided in Tables 3 and 4.
Diversity analysis of fungal and bacterial endophytic communities
Diversity analysis using alpha diversity indices revealed clear differences in species richness, evenness, and dominance across the four tomato cultivars ToLCV-resistant tomato cultivars (Nandi, Sankranthi, and Vybhav) and the susceptible cultivar Arka Vikas. Among the fungal endophytes, all three ToLCV-resistant cultivars harboured a greater number of taxa compared to the susceptible cultivar Arka Vikas, indicating higher species richness in the resistant genotypes.
Resistant cultivars (Nandi, Sankranthi, and Vybhav) showed higher diversity of fungal and bacterial endophytes than the susceptible Arka Vikas. Each resistant cultivar had 6 fungal taxa (8–9 isolates), while Arka Vikas had only 3 taxa (6 isolates) (Table 5). Fungal diversity was highest in Nandi and Vybhav (Simpson’s 1-D = 0.9286; Shannon’s H = 2.045), followed by Sankranthi (0.8889; 1.955), with Arka Vikas showing the lowest diversity (0.7333; 1.178) and highest dominance (D = 0.2667). Evenness was greatest in Nandi and Vybhav (1.289). Chao-1 estimates also reflected higher richness in Sankranthi (8.667) and Nandi/Vybhav (7.75) compared to Arka Vikas (3.417). For bacterial endophytes, Vybhav showed the highest diversity (8 taxa, 9 isolates; Simpson = 0.9722; Shannon = 2.432; evenness = 1.423), followed by Sankranthi (7 taxa; 0.9643; 2.281) and Nandi (5 taxa; 0.9048; 1.836). Arka Vikas again had the lowest bacterial diversity (3 taxa, 4 isolates; 0.8333; 1.29) and highest dominance (D = 0.1667). Chao-1 estimates predicted higher unseen richness in Sankranthi (13.56) and Vybhav (17.33) than in Arka Vikas (3.375).
Venn analysis revealed distinct, cultivar-specific communities of fungal and bacterial endophytes among the four tomato genotypes (Fig. 6). Among fungal endophytes, Nandi had four unique taxa, including Xylaria adscendens, Nigrospora sphaerica N. hainanensis and Fusarium delphinoides; Sankranthi (Eutypella scoparia and Epicoccum nigrum) and Vybhav (Pestalotiopsis kenyana and Alternaria burnsii) each had two unique species, while Arka Vikas hosted only Alternaria alternata. Overlap was minimal, with no fungal taxa shared across all cultivars. In bacterial communities, Vybhav exhibited the highest number of unique taxa (Stutzerimonas stutzeri, Neobacillus niacini, Peribacillus asahii, Cytobacillus firmus, Pseudomonas aeruginosa, Pseudomonas putida and Pseudomonas plecoglossicida), followed by Sankranthi (Bacillus pumilus, Bacillus safensis, Neobacillus bataviensis, Klebsiella quasipneumoniae, Klebsiella variicola and Kosakonia sacchari) and Nandi (Paenibacillus lautus, Paenibacillus cineris, Pseudomonas guariconensis and Enterobacter roggenkampii), while Arka Vikas had only two unique species (Bacillus siamensis and Enterobacter cloacae). Bacillus cereus was the only bacterial taxon shared between Vybhav and Arka Vikas, with no species common to all cultivars.

Venn diagrams showing shared and unique culturable fungal (a) and bacterial (b) endophyte taxa among tomato cultivars. Each circle represents one cultivar (Nandi, Sankranthi, Vybhav, and Arka Vikas). Overlaps indicate shared taxa, while non-overlapping regions represent unique taxa.
Cluster analysis and heatmap visualization (Fig. 7), based on the Bray–Curtis dissimilarity index, demonstrated clear separation of endophytic communities between resistant and susceptible tomato cultivars. Fungal endophyte communities exhibited distinct clustering, with Nandi and Arka Vikas showing the highest dissimilarity (1.0), suggesting no shared taxa. Vybhav and Sankranthi had the lowest dissimilarity (0.50), reflecting a more similar fungal assemblage, whereas moderate similarity was observed between Arka Vikas and Vybhav (0.55) indicating relatively greater similarity in their fungal profiles. Similarly, bacterial endophyte communities showed strong cultivar specificity, with the highest dissimilarity (1.0) observed between Nandi and both Vybhav and Arka Vikas and between Sankranthi and the same cultivars, indicating no shared bacterial taxa. Moderate dissimilarity (0.83) between Nandi and Sankranthi suggested minimal overlap, while the lowest dissimilarity (0.81) was between Vybhav and Arka Vikas. These patterns underscore the presence of genotype-specific microbial assemblages, with resistant cultivars harboring more distinct and diverse endophytic communities than the susceptible Arka Vikas. Detailed Bray–Curtis dissimilarity values are presented in Table 6.

Cluster analysis and heatmaps showing similarity of culturable fungal (a) and bacterial (b) endophytic communities among tomato cultivars based on the Bray–Curtis index. Colour intensity reflects pairwise dissimilarity (0 = more similar; 1 = more dissimilar).
Principal Component Analysis (PCA) of bacterial and fungal endophytes showed clear clustering based on tomato cultivars, indicating that each cultivar had a distinct endophytic community. For bacterial endophytes, the first two components explained 79% of the variation, and for fungal endophytes, 78.6%, showing that most of the differences in microbial diversity were well captured in the analysis. In both biplots, the resistant cultivars Nandi, Sankranthi, and Vybhav occupied separate positions, reflecting unique bacterial and fungal community assemblages associated with each genotype (Fig. 8). In contrast, the susceptible cultivar Arka Vikas was positioned near the center of the bacterial PCA and partially overlapped with resistant cultivars in the fungal PCA, suggesting a more mixed or less specialized endophytic profile.

Principal Component Analysis (PCA) biplots showing the distribution of culturable fungal (a) and bacterial (b) endophyte communities among tomato cultivars. Each point represents the community associated with a cultivar (Nandi, Sankranthi, Vybhav, and Arka Vikas). Closer points indicate more similar communities, while greater distances reflect distinct community structures.
Plant growth-promoting traits of endophytes isolated from resistant cultivars
In vitro screening of diverse endophytic isolates from resistant cultivars for key plant growth-promoting traits, namely phosphate (P), potassium (K), and zinc (Zn) solubilization, revealed that while many strains exhibited one or two of these activities, a select group demonstrated robust functionality across all three. Notably, Xylaria adscendens, Fusarium delphinoides, Arcopilus aureus, Epicoccum nigrum, Xylaria curta, Bacillus subtilis, Paenibacillus lautus, Bacillus pumilus, Stutzerimonas stutzeri (formerly Pseudomonas stutzeri), Pseudomonas putida, and Pseudomonas plecoglossicida showed consistently strong solubilization halos in vitro for P, K and Zn, indicating their multifunctionality (Figs. 9 and 10). The multifunctional strains were therefore prioritized for subsequent in planta bioassays aimed at assessing their effects on growth promotion under greenhouse conditions.

In vitro evaluation of plant growth-promoting (PGP) traits of fungal endophytes isolated from ToLCV-resistant tomato cultivars. Bubble plot shows solubilization activity for phosphate (P), potassium (K), and zinc (Zn) by each fungal isolate.

In vitro evaluation of plant growth-promoting (PGP) traits of bacterial endophytes isolated from ToLCV-resistant tomato cultivars. Bubble plot shows solubilization activity for phosphate (P), potassium (K), and zinc (Zn) by each bacterial isolate.
In-planta PGP activity on susceptible cultivar (Arka Vikas)
Eleven high-performing endophytic isolates (five fungal and six bacterial isolates) were evaluated on Arka Vikas seedlings to assess their plant growth-promoting effects compared to an untreated control. Growth parameters were recorded at 30 and 60 days after sowing (DAS) (Fig. 11). At 30 DAS, treatments with Epicoccum nigrum (T4) and Bacillus subtilis (T10) showed significantly greater plant height (T4-24.83 cm and T10-22.7 cm), root weight (T4-1.45 g and T10-1.41 g), shoot weight (T4-5.8 g and T10-5.5 g), and number of leaves (T4-10.6 and T10-9.6) compared to the control (Suplementary Table 2). Most other treatments also performed significantly better than the control at this stage. However, by 60 DAS, differences in plant height and root weight were no longer significant. Shoot weight remained significantly higher in T4, T10, Arcopilus aureus (T3), and Paenibacillus lautus (T9). Although branching showed no significant difference at 30 DAS, T4 showed the highest number of branches at 60 DAS, followed by T10, T3, T7 and T11. Leaf number was consistently higher in T4 and T10 at both time points. Overall, Epicoccum nigrum and Bacillus subtilis stood out as the most effective in enhancing early vegetative growth in tomato, with some sustained effects at 60 DAS.

Bar graphs showing in-planta evaluation of plant growth-promoting effects of endophytes from ToLCV-resistant cultivars on the susceptible cultivar Arka Vikas: (a) plant height, (b) fresh root weight, (c) fresh shoot weight, (d) number of branches, and (e) number of leaves. Treatments T1–T11 correspond to eleven selected endophytes, and T12 is the untreated control. Different letters above bars indicate significant differences at p < 0.05.
Discussion
This study examined the diversity and functional potential of culturable endophytic communities associated with tomato leaf curl virus (ToLCV)-resistant tomato cultivars (Nandi, Sankranthi, and Vybhav) in comparison with a susceptible cultivar (Arka Vikas), and evaluated the plant growth-promoting traits of these endophytes. The resistant cultivars consistently harboured a more diverse and distinct assemblage of endophytes, as evidenced by alpha and beta diversity indices, PCA clustering, and Venn diagram analysis. These findings indicate that host genotype is likely to influences microbial recruitment and structuring within plant tissues16,34.
The results indicate that ToLCV-resistant cultivars (Nandi, Sankranthi, and Vybhav) harbour richer and more functionally diverse endophytic communities than the susceptible cultivar Arka Vikas. The greater recovery of fungal and bacterial isolates from resistant cultivars suggests enhanced microbial recruitment or compatibility, possibly influenced by host metabolites and immune cues, as reported in other crops18. Conversely, the lower diversity in Arka Vikas is consistent with reports that susceptible genotypes often maintain fewer or less beneficial endophytes14. Diversity indices confirmed that resistant cultivars possessed more balanced communities, with higher fungal richness and greater bacterial diversity, particularly in Vybhav and Sankranthi. Tissue-specific patterns were evident, with fungi predominating in leaves and bacteria more common in roots and stems, reflecting nutrient availability and entry routes34,35. Morphological and molecular analyses revealed several beneficial species, including Ovatospora brasiliensis, Arcopilus aureus, Xylaria curta, Pestalotiopsis microspora, Paenibacillus lautus, Kosakonia sacchari, and Stutzerimonas stutzeri, which are with reported plant-beneficial traits like promote growth, enhance nutrient uptake, and activate systemic resistance13,14,35. The consistent detection of Chaetomium globosum across all cultivars highlights its broad adaptability and antagonistic potential36. Overall, these findings indicate that resistant genotypes are associated with a more diverse and spatially structured endophytic community. While such communities may have implications for plant health and stress tolerance, our results are correlative, and functional validation under ToLCV infection will be required to determine whether specific endophytes contribute to resistance37,38,39,40,41.
The Venn diagram, Bray–Curtis clustering, and PCA revealed clear genotype-associated structuring of endophytic communities in tomato. Resistant cultivars (Nandi, Sankranthi, and Vybhav) contained more unique fungal and bacterial taxa than the susceptible cultivar Arka Vikas. Vybhav showed the highest bacterial diversity, while Nandi was enriched in distinct fungal taxa, indicating host-related differences in microbial assembly. Arka Vikas exhibited fewer unique taxa and limited overlap with resistant cultivars, supporting the role of host genotype in shaping communities34,39,41. Bray–Curtis clustering and PCA further separated Arka Vikas from the resistant cultivars, consistent with genotype-mediated ecological filtering16,42,43. These trends align with earlier reports that resistant genotypes tend to enrich endophytes with putative beneficial traits17,44,45. However, the link between these communities and ToLCV resistance remains correlative. Functionally, the higher endophytic diversity observed in resistant cultivars could, in theory, contribute to improved plant performance through mechanisms such as niche competition, resource exclusion, or complementary functional roles. Several of the isolates identified here have previously been reported to synthesize antimicrobial metabolites, promote growth, or induce systemic resistance (e.g., Chaetomium globosum, Epicoccum spp., Pestalotiopsis spp., Bacillus subtilis, and Paenibacillus lautus)46,47,48,49,50,51,52,53,54,55. Some species within these genera have also shown antiviral activity in other pathosystems.
A diverse microbiome can strengthen plant resistance through multiple, complementary mechanisms. Different microbial species can activate distinct defense pathways, enhance the plant’s immune system, and enable it to respond more effectively to a broad range of pathogens. Beneficial microbes also compete with pathogens for resources and colonization sites, which helps reduce pathogen establishment and disease incidence. In addition, many endophytes synthesize antimicrobial metabolites that directly inhibit pathogen growth and proliferation. Taken together, these results suggest that resistant cultivars are associated with a more diverse and functionally redundant endophytic community, which may have implications for plant health and resilience. We emphasize that these findings indicate association, and future work involving virus-challenge assays and inoculation experiments will be essential to test the hypothesis that specific endophytes contribute to ToLCV resistance.
In vitro screening revealed that several endophytes from ToLCV-resistant tomato cultivars expressed multiple plant growth-promoting (PGP) traits, notably the solubilization of phosphate, potassium, and zinc, which are critical for nutrient availability and plant vigor13,56,57,58. Both fungal isolates such as Xylaria adscendens, Fusarium delphinoides, Arcopilus aureus, Epicoccum nigrum, and Xylaria curta and bacterial strains including Bacillus pumilus, Pseudomonas plecoglossicida, Stutzerimonas stutzeri, Paenibacillus lautus, Bacillus subtilis, and Pseudomonas putida displayed strong nutrient-solubilizing capacity, suggesting their potential contribution to ToLCV tolerance through enhanced nutrition. Such multifunctional microbes are well-documented for roles in nutrient cycling and disease suppression45,59,60. In planta experiments with eleven promising isolates further confirmed their positive impact on the susceptible cultivar Arka Vikas, with several treatments significantly improving seedling growth compared to controls, especially during the first 30 days after sowing (DAS). Among these, Epicoccum nigrum and Bacillus subtilis consistently promoted superior root and shoot development, highlighting their robust PGP potential61. Recent discoveries in peptide-mediated signaling, such as the REF1–PORK1 pathway, illustrate additional host regulatory layers that may interact with microbiome dynamics to influence plant growth and stress responses62. The early effects were probably caused by improved solubilization of mineral nutrients, a crucial factor in rapid seedling establishment, although differences lessened by 60 DAS, potentially due to plant growth convergence or changed rhizospheric conditions that reduced endophyte activity63. This temporal pattern may reflect transient colonization, reduced expression of PGP traits as plants matured, or nutrient competition as soil resources became depleted64,65,66. It is also possible that single-time inoculation was insufficient to maintain active populations later in the growth cycle. Earlier reports show that early improvements in nutrient uptake and seedling vigor after inoculation are common, but sustained benefits over the full growth cycle often require continued presence or reinoculation of beneficial strains [67]. Further work is needed to determine persistence, colonization dynamics, and whether repeated inoculation would produce longer-term benefits.
The findings from this study suggest that consideration of the plant-associated microbiota, alongside genetic traits, may be important for future microbiome-informed crop improvement strategies. The distinct clustering of endophytic communities in resistant cultivars points to the possibility of developing microbial “fingerprints” associated with resistance phenotypes, although these relationships remain associative at present. A key limitation of this work is that it focuses exclusively on the culturable fraction of the endophytic microbiome. Culture-dependent techniques tend to favour fast-growing and metabolically versatile taxa and may underrepresent slow-growing, obligate symbionts, or rare community members. Future studies integrating amplicon-based, metagenomic, and transcriptomic approaches will be essential to capture unculturable taxa, resolve community structure at higher resolution, and provide deeper insight into the functional interactions between the host and microbiome during ToLCV infection. In addition, studies should experimentally evaluate endophytes under controlled ToLCV challenge using either mechanical or vector-mediated inoculation. Their effects could be assessed through plant symptom scoring, quantification of viral load (qPCR), and monitoring of defense-related marker genes to determine whether any strains contribute to antiviral protection. Identifying microbial taxa that consistently associate with resistant cultivars could ultimately inform the rational development of biocontrol consortia or probiotic formulations, pending functional validation under controlled infection conditions.
Conclusion
This study shows that ToLCV-resistant tomato cultivars (Nandi, Sankranthi, and Vybhav) are associated with richer and more functionally diverse communities of culturable endophytes than the susceptible cultivar Arka Vikas. The composition of these microbial communities appeared to be influenced by host genotype, as indicated by diversity indices, Venn analysis, Bray–Curtis clustering, and PCA. Several endophytes, notably Epicoccum nigrum and Bacillus subtilis, exhibited consistent plant growth-promoting traits and enhanced seedling vigour under controlled conditions, highlighting their potential as candidate bioinoculants. The recovery of unique taxa from resistant cultivars suggests possible roles in defense priming or stress tolerance; however, this relationship remains correlative. Clustering and PCA analyses indicated genotype-associated microbiome assembly, pointing to microbial profiles that may be linked with resistance phenotypes. Overall, these findings support the idea that microbiome considerations could complement conventional breeding and disease-management strategies. Future research should prioritize in planta functional validation under ToLCV challenge, long-term field trials and omics-based approaches to clarify host–microbe–virus interactions and to identify robust, reproducible microbial taxa that could eventually inform probiotic development in virus-affected cropping systems.
Data availability
Data will be made available on request.
References
Muniyappa, V. et al. Reaction of Lycopersicon cultivars and wild accessions to tomato leaf curl virus. Euphytica 56, 37–41 (1991).
Banks, G. K. et al. First report of the Bemisia tabaci B biotype in India and an associated tomato leaf curl virus disease epidemic. Plant. Dis. 85, 231–231. https://doi.org/10.1094/PDIS.2001.85.2.231C (2001).
Ponselvakumari, M. K. et al. Assessment of tomato genotypes against the whitefly, Bemisia tabaci gennadius (Hemiptera: Aleyrodidae) and tomato leaf curl virus under controlled condition. Int. J. Trop Insect Sci. 44, 581–593. https://doi.org/10.1007/s42690-024-01167-y (2024).
Singh, R. K. et al. Genetic and molecular characterisations of tomato leaf curl virus resistance in tomato (Solanum lycopersicum L). J. Hortic. Sci. Biotechnol. 90, 503–510. https://doi.org/10.1080/14620316.2015.11668706 (2015).
Prasad, A., Sharma, N., Hari-Gowthem, G., Muthamilarasan, M. & Prasad, M. Tomato yellow leaf curl virus: impact, challenges, and management. Trends Plant. Sci. 25, 897–911. https://doi.org/10.1016/j.tplants.2020.03.015 (2020).
Chowdhury, S. et al. Disruption of tomato TGS machinery by ToLCNDV causes reprogramming of vascular tissue-specific TORNADO1 gene expression. Planta 256, 78. https://doi.org/10.1007/s00425-022-03985-1 (2022).
Nateshan, H. M., Muniyappa, S. H., Jalikop, S. H. & Ramappa, H. K. Resistance of Lycopersicon species and hybrids to tomato leaf curl geminivirus. In Bemisia: 1995 taxonomy, biology, damage, control and management (1996).
Shankarappa, K. S. et al. Development of tomato hybrids resistant to tomato leaf curl virus disease in South India. Euphytica 164, 531–539. https://doi.org/10.1007/s10681-008-9750-1 (2008).
Marchant, W. G., Gautam, S., Hutton, S. F. & Srinivasan, R. Tomato yellow leaf curl virus-resistant and -susceptible tomato genotypes similarly impact the virus population genetics. Front. Plant. Sci. 11, 599697. https://doi.org/10.3389/fpls.2020.599697 (2020).
Muniyappa, V. et al. Tomato leaf curl virus-resistant tomato lines TLB111, TLB130, and TLB182. HortSci. ( (2002).
Muniyappa, V. et al. Tomato leaf curl virus from Bangalore (ToLCV-Ban4): sequence comparison with Indian ToLCV isolates, detection in plants and insects, and vector relationships. Arch. Virol. 145, 1583–1598 (2000).
Gowtham, H. G. et al. N. Fungal endophytes as mitigators against biotic and abiotic stresses in crop plants. J. Fungi. 10, 116. https://doi.org/10.3390/jof10020116 (2024).
Glick, B. R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 169, 30–39. https://doi.org/10.1016/j.micres.2013.09.009 (2014).
Liu, H. et al. Inner plant values: diversity, colonization and benefits from endophytic bacteria. Front. Microbiol. 8, 2552. https://doi.org/10.3389/fmicb.2017.02552 (2017).
Kavya, B. S., Arati, Chethan, D. & Nagaraju, N. Exploring the potential of endophytic fungi in enhancing the growth of cultivated Pigeonpea. Mysore J. Agric. Sci. 59, 110–144 (2025).
Berg, G. & Raaijmakers, J. M. Saving seed microbiomes. ISME J. 12, 1167–1170 (2018).
Trivedi, P., Leach, J. E., Tringe, S. G., Sa, T. & Singh, B. K. Plant–microbiome interactions: from community assembly to plant health. Nat. Rev. Microbiol. 18, 607–621. https://doi.org/10.1038/s41579-020-0412-1 (2020).
Chen, T. et al. A plant genetic network for preventing dysbiosis in the phyllosphere. Nature 580, 653–657. https://doi.org/10.1038/s41586-020-2185-0 (2020).
Sahu, P. K. et al. Surface sterilization for isolation of endophytes: ensuring what (not) to grow. J. Basic. Microbiol. 62, 647–668. https://doi.org/10.1002/jobm.202100462 (2022).
Manias, D., Verma, A. & Soni, D. K. Isolation and characterization of endophytes: biochemical and molecular approach. In Microbial Endophytes (1–14). Woodhead Publishing (2020).
Patel, R., Vinchurkar, M., Patkar, R., Ramesh, R. & BaghiniM. S. Identification, Characterization, Isolation, and pathogenicity study of ralstonia solanacearum: foundation for development of a highly selective DNA-Biosensing platform. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 93, 595–601. https://doi.org/10.1007/s40011-023-01464-5 (2023).
Chethan, D. et al. R. A novel report of Rhizoctonia solani AG-4 HG-I associated with damping-off and wilt of quinoa (Chenopodium quinoa) in India. Physiol. Mol. Plant Pathol. 102725; (2025). https://doi.org/10.1016/j.pmpp.2025.102725
White, T. J., Bruns, T., Lee, S. J. W. T. & Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protocols: Guide Methods Appl. 18, 315–322 (1990).
Zaoti, Z. F. et al. Biochemical and molecular characterization of bacterial wilt disease of banana and evaluation of their antibiotic sensitivity. Microbiol. Res. J. Int. 22, 1–10 (2018).
Hammer, O. P. A. S. T. Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4, 9 (2001).
Kavitha, T. R. et al. Alpha diversity analysis using Shannon index for each species in the rhizosphere soil. In Plant Microbiome Engineering (pp. 293–298). Springer US; (2024). https://doi.org/10.1007/978-1-0716-4180-4_36
Deutsch, Y., Gur, L., Berman Frank, I. & Ezra, D. Endophytes from algae, a potential source for new biologically active metabolites for disease management in aquaculture. Front. Mar. Sci. 8, 636636. https://doi.org/10.3389/fmars.2021.636636 (2021).
Barta, M., Pastirčáková, K., Ostrovský, R. & Kobza, M. Kádasi Horáková, M. Culturable endophytic fungi in Fraxinus excelsior and their interactions with Hymenoscyphus Fraxineus. Forests 13, 1098. https://doi.org/10.3390/f13071098 (2022).
Harrison, J. G. & Griffin, E. A. The diversity and distribution of endophytes across biomes, plant phylogeny and host tissues: how Far have we come and where do we go from here? Environ. Microbiol. 22, 2107–2123. https://doi.org/10.1111/1462-2920.14968 (2020).
Jasim, B., Jimtha, J., Jyothis, C., Radhakrishnan, E. K. & M. & Plant growth promoting potential of endophytic bacteria isolated from Piper nigrum. Plant. Growth Regul. 71, 1–11. https://doi.org/10.1007/s10725-013-9802-y (2013).
Hu, X., Chen, J. & Guo, J. Two phosphate-and potassium-solubilizing bacteria isolated from Tianmu Mountain, Zhejiang, China. World J. Microbiol. Biotechnol. 22, 983–990. https://doi.org/10.1007/s11274-006-9144-2 (2006).
Fomina, M. A., Alexander, I. J., Colpaert, J. V. & Gadd, G. M. Solubilization of toxic metal minerals and metal tolerance of mycorrhizal fungi. Soil. Biol. Biochem. 37, 851–866. https://doi.org/10.1016/j.soilbio.2004.10.013 (2005).
Kavya, B. S., Chethan, D., Prajwal, B. & Nagaraju, N. Biofertilizer and biostimulant potential of Pigeonpea (Cajanus scarabaeoides L.) bacterial endophytes in vitro and in planta study. Int. J. Plant. Soil. Sci. 37, 104–117. https://doi.org/10.9734/ijpss/2025/v37i45392 (2025).
Hardoim, P. R., van Overbeek, L. S. & van Elsas, J. D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 16, 463–471. https://doi.org/10.1016/j.tim.2008.07.008 (2008).
Compant, S., Clément, C. & Sessitsch, A. Plant growth-promoting bacteria in the rhizo-and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil. Biol. Biochem. 42, 669–678. https://doi.org/10.1016/j.soilbio.2009.11.024 (2010).
Goda, M. S. et al. Antimicrobial potential of different isolates of Chaetomium globosum combined with liquid chromatography tandem mass spectrometry chemical profiling. Biomolecules 13, 1683; (2023). https://doi.org/10.3390/biom13121683
Compant, S., Duffy, B., Nowak, J., Clément, C. & Barka, E. A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 71, 4951–4959. https://doi.org/10.1128/AEM.71.9.4951-4959.2005 (2005).
Lugtenberg, B. & Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 63, 541–556 (2009).
Berg, G. & Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 68, 1–13. https://doi.org/10.1111/j.1574-6941.2009.00654.x (2009).
Marquez-Santacruz, H. A., Hernandez-Leon, R., Orozco-Mosqueda, M. D. C., Velazquez-Sepulveda, I. & Santoyo, G. Diversity of bacterial endophytes in roots of Mexican husk tomato plants (Physalis ixocarpa) and their detection in the rhizosphere. Genet. Mol. Res. 9, 2372–2380 (2010).
Longoria-Espinoza, R. M., Félix-Gastélum, R. & Cordero-Ramírez, J. D. Diversity of endophytic bacteria associated with tomato plants (Solanum lycopersicum). Rev. Mex Fitopatol. 38, 307–319 (2020).
Edwards, J. et al. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl Acad. Sci. U.S.A. 112, E911–E920; (2015). https://doi.org/10.1073/pnas.1414592112
Walters, W. A. et al. Large-scale replicated field study of maize rhizosphere identifies heritable microbes. Proc. Natl Acad. Sci. U.S.A. 115, 7368–7373; (2018). https://doi.org/10.1073/pnas.1800918115
Upreti, R. & Thomas, P. Root-associated bacterial endophytes from Ralstonia solanacearum resistant and susceptible tomato cultivars and their pathogen antagonistic effects. Front. Microbiol. 6, 255. https://doi.org/10.3389/fmicb.2015.00255 (2015).
Liu, H., Brettell, L. E., Qiu, Z. & Singh, B. K. Microbiome-mediated stress resistance in plants. Trends Plant. Sci. 25, 733–743. https://doi.org/10.1016/j.tplants.2020.03.014 (2020).
Sharma, S., Pandey, S., Kulshreshtha, S. & Dubey, M. Biology and application of Chaetomium globosum as a biocontrol agent: current status and future prospects. Microorganisms 13, 1646. https://doi.org/10.3390/microorganisms13071646 (2025).
Taguiam, J. D., Evallo, E. & Balendres, M. A. Epicoccum species: ubiquitous plant pathogens and effective biological control agents. Eur. J. Plant. Pathol. 159, 713–725. https://doi.org/10.1007/s10658-021-02207-w (2021).
An, Q. et al. Culture filtrates derived from endophytes Epicoccum spp. Confer resistance against tobacco mosaic virus in Nicotiana benthamiana. Physiol. Mol. Plant. Pathol. 138, 102732. https://doi.org/10.1016/j.pmpp.2025.102732 (2025).
Bernardino, M. C., Couto, M. L. C., Vaslin, M. F. & Barreto-Bergter, E. Antiviral activity of glucosylceramides isolated from Fusarium oxysporum against tobacco mosaic virus infection. PLoS One. 15, e0242887. https://doi.org/10.1371/journal.pone.0242887 (2020).
Deshmukh, S. K., Prakash, V. & Ranjan, N. Recent advances in the discovery of bioactive metabolites from Pestalotiopsis. Phytochem Rev. 16, 883–920. https://doi.org/10.1007/s11101-017-9495-3 (2017).
Wu, C., Wang, Y. & Yang, Y. Pestalotiopsis diversity: Species, dispositions, secondary metabolites, and bioactivities. Molecules 27, 8088. https://doi.org/10.3390/molecules27228088 (2022).
Etesami, H., Jeong, B. R. & Glick, B. R. Potential use of Bacillus spp. As an effective biostimulant against abiotic stresses in crops — A review. Curr. Res. Biotechnol. 5, 100128. https://doi.org/10.1016/j.crbiot.2023.100128 (2023).
Grady, E. N., MacDonald, J., Liu, L., Richman, A. & Yuan, Z. C. Current knowledge and perspectives of Paenibacillus: a review. Microb. Cell. Fact. 15, 203. https://doi.org/10.1186/s12934-016-063-7 (2016).
Kavino, M. et al. Rhizosphere and endophytic bacteria for induction of systemic resistance of banana plantlets against bunchy top virus. Soil. Biol. Biochem. 39, 1087–1098. https://doi.org/10.1016/j.soilbio.2006.11.020 (2007).
Pieterse, C. M. et al. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 52, 347–375 (2014).
Sharma, S. B., Sayyed, R. Z., Trivedi, M. H. & Gobi, T. A. Phosphate solubilizing microbes: sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2, 587. https://doi.org/10.1186/2193-1801-2-587 (2013).
Sinno, M., Ranesi, M., Gioia, L., d’Errico, G. & Woo, S. L. Endophytic fungi of tomato and their potential applications for crop improvement. Agriculture 10, 587. https://doi.org/10.3390/agriculture10120587 (2020).
Yadav, R. C. et al. Zinc-solubilizing Bacillus spp. In conjunction with chemical fertilizers enhance growth, yield, nutrient content, and zinc biofortification In wheat crop. Front. Microbiol. 14, 1210938. https://doi.org/10.3389/fmicb.2023.1210938 (2023).
Devi, R. et al. Minerals solubilizing and mobilizing microbiomes: A sustainable approach for managing minerals’ deficiency in agricultural soil. J. Appl. Microbiol. 133, 1245–1272. https://doi.org/10.1111/jam.15627 (2022).
Pradhan, N. et al. A review on Microbe-Mineral transformations and their impact on plant growth. Front. Microbiol. 16, 1549022. https://doi.org/10.3389/fmicb.2025.1549022 (2025).
Shaffique, S. et al. J. A review on the role of endophytes and plant growth promoting rhizobacteria in mitigating heat stress in plants. Microorganisms 10, 1286. https://doi.org/10.3390/microorganisms10071286 (2022).
Harman, G. E., Howell, C. R., Viterbo, A., Chet, I. & Lorito, M. Trichoderma species — opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2, 43–56. https://doi.org/10.1038/nrmicro797 (2004).
Hashem, A., Tabassum, B. & Abd Allah, E. F. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 26, 1291–1297. https://doi.org/10.1016/j.sjbs.2019.05.004 (2019).
Blake, C., Christensen, M. N. & Kovács, Á. T. Molecular aspects of plant growth promotion and protection by Bacillus subtilis. Mol. Plant. Microbe Interact. 34, 15–25. https://doi.org/10.1094/MPMI-08-20-0225-CR (2021).
El-Sharkawy, H. H., Rashad, Y. M. & Elazab, N. T. Biocontrol potential of the endophytic Epicoccum nigrum HE20 against Stripe rust of wheat. Pestic Biochem. Physiol. 194, 105517. https://doi.org/10.1016/j.pestbp.2023.105517 (2023).
Ali, S. A. B., Abdelmoaty, H., Ramadan, H. & Salman, Y. The endophytic fungus Epicoccum nigrum: isolation, molecular identification and study its antifungal activity against phytopathogenic fungus Fusarium Solani. J Microbiol. Biotechnol Food Sci. 13, e10093. https://doi.org/10.55251/jmbfs.10093 (2024).
Acknowledgements
The authors thank the Department of Plant Pathology, UAS, GKVK, Bengaluru, for providing the inputs and laboratory facilities to carry out this investigation.
Funding
No funding was received for conducting this study.
Author information
Authors and Affiliations
Contributions
**DC: ** Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing-original draft, Writing-review and editing; **KBS: ** Conceptualization, Formal analysis, Methodology, Supervision, Visualization, Writing: original draft, Writing-review and editing; **A: ** Formal analysis, Supervision, Visualization, Writing-review and editing; **RC: ** Writing-review and editing; **AS: ** Formal analysis; **HPG: ** Writing-review and editing; **SC: ** Formal analysis; **NN: ** Conceptualization, Project administration, Software, Resources, Supervision and Visualization, **CNLR: ** Conceptualization, Methodology, Supervision and Visualization; **SGK: ** Conceptualization and Supervision; **TMN: ** Conceptualization and Supervision.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Chethan, D., Kavya, B.S., Arati et al. Endophyte profiling of tomato leaf curl virus (ToLCV) resistant and susceptible tomato genotypes: Insights into microbial diversity and growth promotion. Sci Rep 16, 5348 (2026). https://doi.org/10.1038/s41598-026-37677-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-37677-7
