
Abstract
Ginsenoside Rg1 (Rg1), a major active component of ginseng, exerts antidepressant-like effects, but the mechanisms underlying its anti-inflammatory activity and association with hippocampal neuronal remain unclear. A depression model was established in ICR mice via lipopolysaccharide (LPS) injection to investigate the effects of Rg1 on neuroinflammation and neuronal injury, as well as its antidepressant mechanism. Cognitive dysfunctions were evaluated via forced swim test (FST), sucrose preference test (SPT) and elevated plus maze (EPM). The effects of LPS-induced neuronal damage and Rg1 on anti-inflammatory pathways were subsequently evaluated through biochemical assays, and histological analysis, including Hematoxylin and Eosin staining, Nissl staining, Immunohistochemistry, ELISA and transmission electron microscopy (TEM). After confirming Rg1’s antidepressant activity, its possible mechanisms were analyzed through Proteomic. Finally, Western blot was used to detect the key targets for predictive mechanisms. Results showed Rg1 significantly ameliorated LPS-induced depressive-like behaviors, reduced serum pro-inflammatory cytokines, attenuated neuronal loss, and increased hippocampal NeuN-positive cell density. The p-MST1/MST1 and p-YAP/YAP ratios were elevated in the model group but reversed by Rg1 and fluoxetine treatment. This study indicated Rg1 exerts antidepressant effect in mice, possibly by inhibiting hyperphosphorylation of key Hippo-YAP signaling pathway proteins.
Similar content being viewed by others
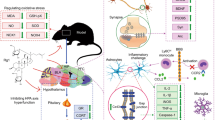

Introduction
Depression, a mood disorder causing persistent sadness and loss of interest, affects approximately 350 million people globally1,2. The World Health Organization has ranked it as the leading contributor to global disability3. Its etiology is complex, characterized by altered neuronal structure, functional impairments, and changes in interneuronal interaction4,5,6,7. Commonly prescribed antidepressants, such as fluoxetine, have limitations including cardiotoxicity, suicidal tendencies, sexual dysfunction, and sleep disorders8,9,10,11. Therefore, there is an urgent need to explore effective alternative treatments for depression.
Ginsenoside Rg1, a protopanaxadiol isolated from Panax ginseng root, exhibits neuroprotective and anti-inflammatory activities5,12. Rg1 exerts diverse pharmacological effects primarily by inhibiting the secretion of inflammatory factors, downregulating the expression of pro-apoptotic genes, and modulating redox homeostasis in the organism13,14.Importantly, Rg1 is involved in signaling pathways including BDNF/TrkB/ERK, NF-κB/NLRP3, and Hippo-YAP, and it exhibits neuroprotective properties, which have been thoroughly investigated in other neurological conditions15,16,17. However, the molecular mechanisms underlying its antidepressant effects remain to be fully elucidated.
To investigate the role of Rg1 in depression, the depressive-like mouse model induced by LPS injection was utilized in this study to elicit cerebral neuroinflammation. After four weeks of continuous Rg1 administration, behavioral tests were conducted. Furthermore, to continue exploring and explaining the mechanism of Rg1 in depression, this study used gene ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG), and protein–protein interaction (PPI) network analyses to predict underlying molecular mechanisms of the Rg1 in depression treatment.
Methods
Animals
Based on our previous experience in the laboratory, we assessed the efficacy and mortality outcomes of depression modeling and ultimately selected 60 ICR mice (7–8 weeks old) for our current study. These mice, weighing between 20 and 25 g, were procured from Beijing Vital River Laboratory Animal Technology Co., Ltd. The animal production certificate number was SCXK(Ji)20190008. The mice were housed in a well-lit environment with alternating 12-h light and dark cycles, a temperature range of 22–24 °C, and a humidity level of 50–60%. Our study examined the anxiety and depressive-like behaviors of depression mice with depression. To prevent potential communication between groups of mice that could compromise the validity of our findings, we housed six mice from each group in separate cages, with opaque cardboard barriers separating them. They were accommodated in the animal room of Hebei Academy of Chinese Medical Sciences. This experiment has secured review and approval from the Animal Experiment Ethics Committee of Hebei University of Chinese Medicine (DWLL202302019). All experiments were performed in accordance with relevant named guidelines and regulations. The authors complied with the ARRIVE guidelines.
LPS injection
All mice except those in the normal group were subjected to intracerebroventricular injection of lipopolysaccharide (LPS, Beijing Solarbio Life Sciences, China, L8880) for neuroinflammation induction. Injection coordinates were defined with the bregma as the origin: 0.5 mm anteroposterior, 1.0 mm mediolateral, and 2.5 mm dorsoventral. LPS was prepared by dissolving it in distilled water, and 3 µL (4 µg/µL) of LPS was injected into the ICV region of mouse brains once, according to the previously documented approach18. As a sham control, mice in the normal group were administered an equivalent volume of sterile normal saline via the same ICV injection protocol. The experiment was performed as illustrated in Fig. 1.

Flow chart of experiment.
Groups
The mice were randomly divided into five groups: (1)control mice distilled water through gavage (Control group, n = 12), (2) depression mice treated with normal saline (NS) (Model group, n = 12), (3) depression mice treated with fluoxetine (10 mg/kg/day, Fluoxetine group, n = 12, Lilly Suzhou Pharmaceutical Co., Ltd. HJ20181215). (4) depression mice treated with Rg1 low-dose (20 mg/kg/day, LRg group, n = 12, purity ≥ 98, confirmed by high-performance liquid chromatography analysis, Shanghai yuanye Bio-Technology Co., Ltd. B21057), and (5) depression mice treated with Rg1 high-dose (40 mg/kg/day, HRg group, n = 12). The dose of Rg depends on the previous report. The treatments started from day 7 till 35 and were given between 9:00 and 11:00 h once daily via gastric gavage (Fig. 2).

Representative diagram for the experimental process. All mice except the normal group received intracerebroventricular injection of LPS on day 7 to induce neuroinflammation. The drug administration was began on day 8 and continued for 28 days to assess the effect of Rg1 on depression.
Forced swimming test, FST
The mice were positioned in a transparent cylindrical container filled with clean water (height × diameter: 25 cm × 10 cm). The container was filled with warm water maintained at 24 ± 1 °C. The mice’s behavior during the 6-min swim was recorded by a video camera positioned above the glass cylinder. Each test was performed using fresh water. The immobility time was recorded during the final 4 min of the test.
Sucrose preference test, SPT
Before the test, mice were individually caged and acclimated to a sucrose solution and water in two bottles for 24 h. The positions of the bottles were alternated every 12 h to minimize side bias. After 24 h, a bottle filled with 2% sucrose and a bottle of regular water were placed in each cage. Twelve hours later, the water and sucrose bottles were removed and weighed to calculate the amounts of water and sucrose consumed. Sucrose preference was calculated using the following formula: (sucrose solution consumed/total liquid consumed) × 100%.
Elevated plus-maze test, EPM
The elevated plus-maze test apparatus consisted of black plexiglass. It had two open and two enclosed arms of equal length (50 cm), symmetrically installed in a crisscross pattern (10 × 10 cm) and positioned 50 cm above the floor. A square central area was formed at the intersection of the four arms. During the experiment, each mouse was placed in the central region, facing an open arm, and allowed to explore freely for 10 min. The number of entries into the open arms and the the time spent in the open arms was recorded.
Hematoxylin and eosin staining, HE
Following treatment, mice were deeply anesthetized with 2% isoflurane to ensure complete loss of consciousness, after which euthanasia was performed via cervical dislocation. Immediately following humane euthanasia, hippocampal tissues were rapidly dissected and fixed in 4% paraformaldehyde, and subsequently subjected to gradient dehydration using ethanol solutions of ascending concentrations. Following paraffin embedding, the tissues were sectioned to a thickness of 4–6 μm and then subjected to sequential dewaxing and rehydration. The sections were stained with hematoxylin for 5 min, followed by eosin counterstaining for 3 min. After staining, the sections were processed sequentially through 95% ethanol, absolute ethanol, xylene, and then mounted in a resinous medium. Histological alterations in the mouse hippocampus were subsequently assessed by light microscopy.
Nissl staining
The mouse brain tissue underwent dewaxing of the wax mass. The tissue slices were rinsed in distilled water and then stained with Toluidine blue. The staining vat was maintained at 50–60 °C for 25–50 min. After staining, the slices were gently rinsed in distilled water and then in 70% ethanol. Rapid fractionation was performed with 95% ethanol. Subsequently, the sections were rapidly dehydrated in anhydrous ethanol, then cleared in xylene. Finally, the sections were mounted with neutral gum. The mouse hippocampus was observed under a microscope, and photographs were taken for analysis.
Immunohistochemistry
The mouse brain tissues were embedded in paraffin and cut into 5-μm coronal sections using an automatic microtome. After the tissues were placed on slides, deparaffinized, and rehydrated, they were immersed in sodium citrate for antigen retrieval. The sections were blocked with standard goat serum solution and then incubated overnight at 4 °C with NeuN Rabbit Monoclonal Antibody (Beyotime AF1072). After three washes with PBS, these sections were incubated with a biotinylated rabbit anti-goat IgG secondary antibody for 30 min at room temperature, then washed three times with PBS. A DAB kit was employed for chromogenic detection, and the sections were stained with hematoxylin and mounted. Finally, Optical density were quantified using ImageJ software.
ELISA
Following the experiments, the tissues were promptly collected. For each mouse, blood serum was prepared by allowing the blood to clot for at least 30 min. Subsequently, the blood was centrifuged at 3000 revolutions per minute (r/min) at 4 °C for 10 min. The supernatant (serum) was then collected, stored at -80 °C, and thawed immediately before testing. Levels of IL-1β (Elab science Biotechnology, E-EL-M0037) and TNF-α (Elab science Biotechnology, E-EL-M3063) in serum were measured using an ELISA kit in 96-well plates, according to the manufacturer’s instructions. The absorbance of each well of the 96-well plate was measured using a microplate reader, and the IL-1β and TNF-α levels of each sample were calculated based on a standard curve.
Transmission electron microscopy
Mice were anesthetized. Subsequently, their brains were removed and placed in an electron microscopy fixative solution composed of 2% paraformaldehyde and 2% glutaraldehyde. After being rinsed three times with 0.1 M PBS (pH 7.4), the samples were thoroughly fixed in 1% osmium tetroxide at 4 °C for 2 h. Then, they were dehydrated using different gradients of ethanol concentrations (30%, 50%, 70%, 80%, 95%, 100%, and 100%) and 100% acetone. Next, the hippocampal sections were embedded in an embedding solution at room temperature for 2 h, followed by overnight incubation at 37 °C, 12 h at 45 °C, and 24 h at 60 °C. Ultra-thin brain slices (90 nm) were obtained using an ultra-thin microtome and stained with 3% uranium acetate-lead citrate (UA-LC). Finally, the ultrastructure of the hippocampus was observed using transmission electron microscopy (TEM).
Proteomic analysis
The sample was ground into a cell powder using liquid nitrogen and then transferred to a 5-mL centrifuge tube. After that, four volumes of lysis buffer (8 M urea, 1% protease inhibitor cocktail) were added to the cell powder, followed by three rounds of sonication on ice using a high-intensity ultrasonic processor. The remaining debris was removed by centrifugation at 12,000 g for 10 min at 4 °C. Finally, the supernatant was collected, and the protein concentration was determined using a BCA kit according to the manufacturer’s instructions. For digestion, the protein solution was reduced with 5 mM dithiothreitol for 30 min at 56 °C, then alkylated with 11 mM iodoacetamide for 15 min at room temperature in the dark. The protein sample was diluted with 100 mM TEAB to reduce the urea concentration to less than 2 M. Trypsin was added at a 1:50 trypsin-to-protein mass ratio for the first overnight digestion and at a 1:100 trypsin-to-protein mass ratio for the second 4-h digestion. After digestion, the peptides were desalted by a C18 SPE column.
The tryptic peptides were dissolved in solvent A (1% formic acid, 2% acetonitrile in water) and directly loaded onto a reversed-phase analytical column (25 cm in length, 75 μm in diameter). Peptides were separated with a gradient from 5 to 25% solvent B (0.1% formic acid in 90% acetonitrile) over 60 min, 25% to 35% in 22 min and climbing to 80% in 4 min then holding at 80% for the last 4 min, all at a constant flowrate of 450nL/min on an EASY-nLC 1200 UPLC system (ThermoFisher Scientific). The separated peptides were analyzed in Q Exactive HF-X with a nano-electrospray ion source. The electrospray voltage applied was 2.0 kV. The full MS scan resolution was set to 60,000 for a scan range of 350–1600 m/z. Up to 20 of the most abundant precursors were then selected for further MS/MS analyses with 30-s dynamic exclusion.
GO and KEGG pathway analyses
The functional annotations of the UniProt database was used for GO functional enrichment analysis. GO enrichment analysis was exploited to explain and annotate genes by three aspects, including cellular component (CC), molecular function (MF), and biological process (BP) analyses. The KEGG database was utilized for pathway analysis19,20,21. Apart from this, enriched GO terms and relevant pathways with P-values < 0.05 were selected for better prediction and verification of the biological process and mechanism.
Western blotting
Western blotting analysis was performed using proteins extracted from brain tissues (n = 3 mice per group). Hippocampus tissues were dissected on a cold surface and immediately frozen in liquid nitrogen. Hippocampus tissues were homogenized in RIPA buffer, containing protease and phosphatase inhibitors (Sevenbio, Beijing, China; SW103-02), incubated on ice for 30 min, and then centrifuged at 12,000×g for 20 min at 4 °C; the resulting supernatant was collected as the total protein from hippocampal samples. The protein concentration of the final supernatant was determined using a BCA protein quantification kit (Sevenbio, Beijing, China; SW101-02). According to the quantified protein concentration, 5× loading buffer was added to the total protein samples, which were then denatured at 95 °C for 10 min and stored on ice. Equal amounts of protein were separated via sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene difluoride (PVDF) membranes (Sevenbio, Beijing, China; SW120-01). The membranes were incubated overnight at 4 °C with the appropriate primary antibodies: anti-p-MST1 (Proteintech, Wuhan, China; 80093-1-RR, 1:5,000), anti-MST1 (Proteintech, Wuhan, China;22245-1-AP,1:2000), anti-p-YAP (Proteintech, Wuhan, China;22245-1-AP,1:5000), anti-YAP (Immunoway, Suzhou, China; 1:800), and anti-β-catenin (Proteintech, Wuhan, China; 66009-1-Ig, 1:20000). Subsequently, the PVDF membranes were incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies. Finally, protein bands were visualized using enhanced chemiluminescence reagent, and band intensities were quantified using ImageJ software.
Statistical analysis
The statistical analysis of the data was performed using GraphPad Prism 8 software (La Jolla, USA). Data normality was determined using the Shapiro–Wilk test. The Levene test was used to assess the assumption of homogeneity of variance. For multiple group comparisons, a one-way ANOVA and subsequent Tukey post hoc tests were employed. Furthermore, to compare several related samples, a two-way ANOVA was utilized. Values that were not normally distributed were analyzed by nonparametric methods. Differences were considered statistically significant at P < 0.05. The data were represented as mean ± SD (standard deviation).
Results
Effects of Rg1 on depression-like behavior following LPS-induced in depression mice
The FST, SPT, and EPM were the recognized behavioral tests to evaluate depression activities in mice. Compared to the Control group, the immobility time in the FST was significantly increased (P < 0.05.Fig. 3D). However, following treatment, a statistically significant decrease was observed in both the Fluoxetine and Rg1 groups (P < 0.05). In addition, Rg1 administration enhanced the sucrose preference of LPS-induced mice in the SPT (P < 0.05. Fig. 3E), manifesting that Rg1 alleviated anhedonia in depression mice. In the EPM, the Model group displayed significantly fewer open-arm entries and less time spent in the open arms compared with the Control group, while these parameters were notably reversed by treatment with Rg1(P < 0.05, Fig. 3F,G).The behavioral findings sustained that Rg1 treatment could efficiently alleviate the depression-like performance.

Effects of Rg1 on depression-like behavior. (A) Schematic diagram of the forced swim test. (B) Schematic diagram of the sucrose preference test. (C) Schematic diagram of the Elevated plus-maze. (D) Time of immobility. (E) Sucrose preference values. (F) Number of entries in open arm. (G) Time spend in open arm.The values were presented as the mean ± SD. (n = 12. *P < 0.05 vs. Control group; #P < 0.05 vs. Model group).
Effects of Rg1 on hippocampal neurons following LPS-induced in depression mice
HE staining was utilized to evaluate the structural organization of the CA1, CA3, and DG regions in the hippocampus among different experimental groups. In the Control group, the neurons in hippocampal regions showed a well-organized arrangement, with intact structures, uniform chromatin distribution, and distinct nucleoli, and no significant neuronal loss was observed. In contrast, the hippocampus of the Model group exhibited disrupted neuronal organization, particularly in the CA1 and DG regions, with a reduction in neuronal number or even neuronal loss. These neurons showed abnormal morphologies, including darker staining, smaller nuclei, loss of distinct nucleoli, and the presence of neuronal vacuolation. Following treatment with fluoxetine and Rg1, these pathological changes were partially improved (Fig. 4A).

Effects of Rg1 on neuronal damage. (A) HE staining. (B) Nissl staining. Scale bar = 100 μm (200×). The values were presented as the mean ± SD. The red arrow refered to the pathological injury site. (n = 3.).
Similarly, Nissl staining demonstrated that neurons in the Control group were arranged in an orderly manner, with intact cellular structures, intact nucleus, and distinct Nissl bodies (Fig. 4B). In the Model group, nucleus were pyknotic, Nissl bodies were barely distinguishable, and some cells in the DG exhibited vacuolation. Additionally, the cell membrane boundaries in the CA1 and CA3 regions were indistinct. Following fluoxetine and Rg1 treatment, vacuolation in the CA1 and CA3 regions was ameliorated, nuclei and cytoplasm were clearly defined, and cellular arrangement and morphology were more regular. Collectively, these findings indicated that Rg1 effectively suppresses hippocampal neuronal loss in LPS-induced depressive mice.
Effects of Rg1 on NeuN following LPS-induced in depression mice
NeuN, a neuron-specific nuclear protein, is a well-established marker for mature neurons in the vertebrate nervous system22. Compared to the Control group, NeuN expression was significantly reduced in the CA3 and DG regions of the Model group (P < 0.05, Fig. 5C,D). Nevertheless, Rg1 significantly upregulated NeuN expression (P < 0.05, Fig. 5C,D). Notably, in the CA1 region, NeuN expression was increased in the Rg group compared to the Model group, this elevation did not reach statistical significance (P > 0.05, Fig. 5A). These findings indicated that NeuN may exert distinct functions in different hippocampal subregions.

Effects of Rg1 on the expression of NeuN. (A) Expression of NeuN was determined by immunohistochemistry. Scale bar = 100 μm (200×). (B) The change in the number of NeuN positive cells in CA1 of each group. (C) The change in the number of NeuN positive cells in CA3 of each group. (D) The change in the number of NeuN positive cells in DG of each group.The values were presented as the mean ± SD. Red arrows in photomicrographs indicated the positive expression. (n = 3. *P < 0.05 vs. Control group; #P < 0.05 vs. Model group).
Effects of Rg1 on the expression of inflammatory cytokines in the hippocampus
Figure 6 illustrated the effect of Rg1 on the levels of inflammatory cytokines using ELISA in the hippocampus. Hippocampal levels of TNF-α and IL-1β were significantly increased in the Model group compared to the Control group. Following Rg1 treatment, the levels of TNF-α and IL-1β were significantly reduced (P < 0.01, Fig. 6A,B).

Effects of Rg1 on the expressions of TNF-α and IL-1β. (A) Expression of TNF-α in the hippocampus was determined by ELISA. (B) Expression of IL-1β in the hippocampus was determined by ELISA. The values were presented as the mean ± SD. (n = 6. *P < 0.05 vs. Control group; #P < 0.05 vs. Model group).
Effect of Rg1 on hippocampal ultrastructure
Electron microscopy results showed that the neurons in the hippocampus of the Control group were intact at high magnification, with intact nuclear membranes. The mitochondria exhibited a regular shape, intact double membranes, clear internal cristae, and uniform matrix density, alongside structurally normal endoplasmic reticulum. In contrast, the Model group showed distinct pathological changes: indistinct cell membrane structure, widened inter-cellular gaps, and prominent mitochondrial damage characterized by blurred membrane edges, and reduced or fragmented cristae. Following treatments of fluoxetine and HRg1, the cell membranes remained distinct. The mitochondria showed relatively regular morphology, and clearer internal cristae, with no obvious swelling or deformation. In the LRg1 group, the cell membrane was also clearly visible, the cytoplasm was exhibited edema, partial mitochondria exhibited structural impairment, such as incomplete membranes or slightly diminished cristae (Fig. 7).

Effects of Rg1 on morphological changes of neuronal cells of hippocampus. Scale bar = 500 nm (20,000×). The red arrow points to mitochondrial structures. (n = 3.).
Proteomics analysis of depression mice hippocampus
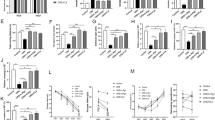
Quantitative proteomics was performed to identify differentially expressed proteins (DEPs) in Control, Model, and HRg1 group (Fig. 8). 6225 proteins were screened. Heatmap of DEPs is shown in Fig. 8A.

Proteomics analysis of depression mice hippocampus. (A) Heatmap of DEPs. (B) PCA of DEPs. (C) Volcano plots of DEPs in Control group and Model group. (D) Volcano plots of DEPs in Model group and Rg1 high-dose group. (E) Volcano plots of DEPs in Control group and Rg1 high-dose group. (F) GO analysis. (G) KEGG pathway enrichment analysis. (n = 3).
Principal Component Analysis (PCA), an unsupervised multivariate statistical method, reveals the internal structure of a dataset and visually illustrates the separation trends between different groups through their spatial distribution on the principal component coordinate plot. PCA was performed on hippocampal samples from each group to identify differential protein expression (Fig. 8B). The results demonstrated a clear separation among the Model, HRg, and Control groups, indicating significant differences in their protein profiles.
Our results revealed that 115 DEPs were identified between the Model and Control groups, of which 20 were downregulated and 95 were upregulated in the Model group (Fig. 8C). Similarly, 33 DEPs were identified in the HRg1-treated group compared with the Model group, with 19 upregulated and 14 downregulated proteins (Fig. 8D).
To study the changes in DEPs of different groups of mice, it is important to understand the function, cellular localization, and biological processes of each protein. For each comparison group, we selected the top 10 significantly enriched GO terms associated with differentially expressed proteins, ranked by ascending p-value, all significant GO terms were included if the number of such terms was fewer than 10 (Fig. 8F). GO enrichment analysis results revealed notable alterations in the Model group and HRg group. In BP, basement membrane organization and regulation of protein dephosphorylation have changed significantly. In CC, chromosome and chromatin showed a significant difference. In MF, phosphatase regulator activity was the main functional category. The results of the GO analysis suggested that basement membrane organization and phosphatase regulator activity may be the main biological processes of the effect of Rg1 in alleviating the depression.
Next, we analyzed the pathways of these expressed proteins by KEGG enrichment analysis (Fig. 8G). Notably, Hippo signaling pathway was the most enriched pathways in the model group and HRg group. This suggested that the role of Rg1 may be related to Hippo signaling pathway.
Effects of Rg1 on the expression of the Hippo/YAP signaling pathway- related proteins in the hippocampus
The Hippo/YAP-related proteins expression level in the hippocampus was determined to explore the mechanism of Rg1 on hippocampal neurogenesis. Compared with the control group, the ratio of p-MST1 to MST1 and p-YAP to YAP were significantly increased in the Model group (Fig. 9B,C). Compared with the Model group, the ratio of p-MST1 to MST1 and p-YAP to YAP were in the Fluoxetine group and HRg1 group were significantly decreased (P < 0.05), as treatments of Rg1 normalized the changes with the consistent effect of fluoxetine. Taken together, this suggests Rg1 might regulate the Hippo/YAP pathway-related proteins to affect neurogenesis.

Effect of Rg1 on the expression of the Hippo/YAP signaling pathway-related proteins in the hippocampus. (A) Representative western blots of Hippo/YAP signaling pathway-related proteins. (B) Quantitative analysis of p-MST1/MST1 in the hippocampus. (C) Quantitative analysis of p-YAP/YAP in the hippocampus. The values were presented as the mean ± SD. (n = 3. *P < 0.05 vs. Control group; #P < 0.05 vs. Model group).
Discussion
The hippocampus, a key regulator of depressive-like behaviors, displays high susceptibility to stress and marked vulnerability to LPS23,24.
Cytokine inducer LPS is widely utilized in animal models of neuropsychiatric disorders to investigate the link between neuroinflammation and memory deficits, anxiety, and depression25,26,27. Systemic administration of LPS at doses below the septic threshold induces time-dependent behavioral alterations28. The immune reaction elicited by this intervention unfolds via the canonical stages of inflammation, typified by an initial transient upregulation of pro-inflammatory cytokines and a progressive increment in the population of phagocytic microglia displaying distinct activated morphological alterations29. This inflammatory response exhibits considerable phenomenological similarities to clinical depression, with shared manifestations including behavioral despair, anhedonia, reduced social interaction, fatigue, and anorexia, strongly validating the suitability of this model for studying neuroinflammation-related depression30,31. A single intracerebroventricular administration of LPS in mice is sufficient to induce stable depressive-like behaviors, which are temporally segregated from transient acute sickness responses, thus validating the sustainability of depression-related behavioral phenotypes32. Although LPS-induced depressive models have been widely used to study inflammation-related mood disorders, most studies have focused on acute behavioral changes within 24 h after injection. Research investigating whether depressive-like behaviors induced by a single LPS injection can persist for up to 28 days post-administration remains relatively limited. Notably, the depression-like behaviors induced by LPS in our study were consistent with previous reports, further supporting the reliability of our experimental protocol and the reproducibility of this model33,34,35,36. Our findings not only fill the research gap regarding the long-term effects of the LPS-induced depression model, but also provide new experimental evidence for further understanding the pathological relationship between inflammation and depression.
Accumulating evidence highlights neuronal loss, synaptic damage, the reduced neuronal density, and hippocampal volume shrinkage as core pathological features of depression37,38,39. The hippocampus has three major excitatory synaptic systems formed by its afferent fibers and intrinsic circuitry40,41,42,43. The integrity of these neuronal and synaptic structures is indispensable for normal function. In line with these pathological characteristics, our TEM analysis confirmed that LPS administration severely disrupted hippocampal synaptic architecture, as reflected by decreased synapse number, reduced postsynaptic density thickness, and signs of neuronal death. Consistently, Nissl staining and immunohistochemistry also demonstrated significant neuronal loss in the hippocampus of LPS-induced mice. Notably, our findings revealed that Rg1 treatment ameliorated LPS-induced structural deficits and enhanced neuronal integrity. These results are meaningful given the critical role of hippocampal neuronal and synaptic integrity in depression.
The nervous and immune systems interact tightly to maintain physiological homeostasis, and the release of pro-inflammatory cytokines is a key disruptor of this balance. These cytokines are widely recognized as central mediators of depression pathophysiology44,45,46. Among these cytokines, TNF-α is linked to depressive phenotypes, including locomotor suppression, anhedonia, fatigue, and cognitive impairment, while IL-1β acts as a critical mediator of neuropathology; notably, inhibition of IL-1β had been shown to exert antidepressant effects47,48,49. Furthermore, numerous studies have indicated a relationship between depression and inflammatory cytokines. Pro-inflammatory cytokines are expressed at low, homeostatic levels in the brain, but their expression is markedly up-regulated in response to stressors or immune challenges, leading to neuroinflammation50,51,52,53,54,55. This elevation in pro-inflammatory cytokines is not only a characteristic feature of depression in patients but also recapitulated in depression models56.In line with these characteristics, LPS significantly up-regulated serum levels of both IL-1β and TNF-α, and Rg1 treatment effectively reduced their expression. This indicated that Rg1 exerts potent anti-inflammatory effects by targeting key pro-inflammatory cytokines to alleviate depression.
To obtain a more in-depth understanding of the effects of Rg1 on depression, we performed proteomic analysis, through which the Hippo/YAP signaling pathway was identified via screening (Fig. 10). The Hippo signaling pathway was recognized as the most significantly enriched pathway among HRg1-modulated differentially expressed proteins (DEPs). This signaling cascade is well-documented to play a pivotal role in regulating cellular processes critical to neuronal homeostasis—including cell survival, proliferation, and apoptosis57. Notably, the hippocampus has emerged as a key site of Hippo signaling pathway activity, implicating its involvement in stress-related psychiatric disorders58,59,60,61. The core transcriptional output of the Hippo signaling pathway is mediated by the co-activators Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ), whose activity is tightly regulated by a kinase cascade involving MST1/2 and LATS1/262. These kinases phosphorylate YAP/TAZ to inhibit their nuclear translocation. Upon activation, unphosphorylated YAP translocates to the nucleus, where it interacts with TEA domain family (TEAD) transcription factors to drive the expression of genes associated with cellular homeostasis, neuronal development, and tissue regeneration. Intriguingly, this regulatory axis is consistent with the results of our GO enrichment analysis. Specifically, the enrichment of phosphatase regulator activity reflects HRg1-mediated modulation of MST1/2 and LATS1/2 kinase activity, which are critical regulators governing the phosphorylation status of YAP and TAZ. Importantly, accumulating evidence supports the relevance of the Hippo-YAP signaling pathway to the pathogenesis of depression. For instance, YAP knockdown in HT22 hippocampal neurons induces mitochondrial dysfunction. Moreover, studies utilizing YAP conditional knockout and overexpression mouse models have demonstrated that YAP signaling is essential for the modulation of depressive-like behaviors63. Biological validation represents a critical step to confirm that the predicted key targets are functionally relevant and to exclude non-specific signaling pathways. For this reason, based on the aforementioned proteomic analysis results and literature reviews, the Hippo/YAP signaling pathway was further validated to explore the anti-depression mechanism of Rg1. Western blot assay results demonstrated that the levels of p-MST1 and p-YAP were markedly elevated in the hippocampus of LPS-induced mice, indicating that the Hippo-YAP signaling pathway was dysregulated following LPS exposure. Notably, Rg1 treatment reversed this pathological dysregulation, as evidenced by significant reductions in the p-MST1/MST1 and p-YAP/YAP ratios in the hippocampus. Collectively, these findings demonstrate that LPS enhances the phosphorylation of key proteins associated with the Hippo-YAP signaling pathway, whereas Rg1 administration effectively inhibited this hyperphosphorylation event and restored hippocampal neurogenesis. These observations further highlight that the Hippo signaling pathway is not only a critical mediator of depression-associated molecular and cellular dysfunction but also a key target through which Rg1 exerts its antidepressant effects by restoring hippocampal protein homeostasis.

Hippo signaling pathway mechanism diagram.
Conclusion
Collectively, the results demonstrated that Rg1 alleviated depressive-like behaviors. These behavioral improvements were accompanied by attenuated hippocampal pathological damage and reduced pro-inflammatory cytokine levels, mediated by Hippo/YAP pathway.
Data availability
All data generated or analysed during this study are included in this published article.
Abbreviations
- Rg1:
-
Ginsenoside Rg1
- LPS:
-
Lipopolysaccharide
- TEM:
-
Transmission electron microscopy
- FST:
-
Forced swim test
- SPT:
-
Sucrose preference test
- EPM:
-
Elevated plus maze
- HE:
-
Hematoxylin–eosin staining
- IL-1β:
-
Interleukin-1 beta
- TNF-α:
-
Tumor necrosis factor alpha
- GO:
-
Gene ontology
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- PPI:
-
Protein–protein interaction
- ICV:
-
Intracerebroventricular
- PBS:
-
Phosphate buffered saline
- ELISA:
-
Enzyme-linked immunosorbent assay
- PCA:
-
Principal components analysis
- YAP:
-
Yes-associated protein
- TAZ:
-
Transcriptional coactivator with a PDZ-binding domain
References
Li, S. et al. Sex difference in incidence of major depressive disorder: an analysis from the global burden of disease study 2019. Ann. Gen. Psychiatry. 22(1), 53 (2023).
Malhi, G. S. & Mann, J. J. Depression. Lancet 392(10161), 2299–2312 (2018).
Moreno-Agostino, D. et al. Global trends in the prevalence and incidence of depression:a systematic review and meta-analysis. J. Affect. Disord. 15(281), 235–243 (2021).
Marx, W. et al. Major depressive disorder. Nat. Rev. Dis. Primers. 9(1), 44 (2023).
Li, Y. et al. Ginsenoside Rb1 protects hippocampal neurons in depressed rats based on mitophagy-regulated astrocytic pyroptosis. Phytomedicine 1(121), 155083 (2023).
Sun, N. et al. A new perspective on hippocampal synaptic plasticity and post-stroke depression. Eur. J. Neurosci. 58(4), 2961–2984 (2023).
Duman, R. S., Aghajanian, G. K., Sanacora, G. & Krystal, J. H. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants. Nat. Med. 22(3), 238–249 (2016).
Xie, S. et al. Lacticaseibacillus rhamnosus KY16 improves depression by promoting intestinal secretion of 5-HTP and altering the gut microbiota. J. Agric. Food Chem. 72(39), 21560–21573 (2024).
Amiri, H., Zamani, N., Hassanian-Moghaddam, H. & Shadnia, S. Cardiotoxicity of tricyclic antidepressant treated by 2650 mEq sodium bicarbonate: A case report. JRSM Cardiovasc. Dis. 5, 2048004016682178 (2016).
Polychroniou, P. E. et al. Temporal profiles and dose-responsiveness of side effects with Escitalopram and Duloxetine in treatment-naïve depressed adults. Behav. Sci. 8(7), 64 (2018).
Clayton, A. H., Croft, H. A., Yuan, J., Brown, L. & Kissling, R. Safety of flibanserin in women treated with antidepressants: A randomized, placebo-controlled study. J. Sex Med. 15(1), 43–51 (2018).
Zhao, B. S. et al. Effects of ginsenoside Rg1 on the expression of toll-like receptor 3, 4 and their signalling transduction factors in the NG108-15 murine neuroglial cell line. Molecules 19(10), 16925–16936 (2014).
Lin, Z. et al. Recent progress (2015–2020) in the investigation of the pharmacological effects and mechanisms of ginsenoside Rb1, a main active ingredient in panax ginseng meyer. J. Ginseng Res. 46(1), 39–53 (2022).
Ling, G. et al. Progress of ginsenoside Rb1 in neurological disorders. Front. Pharmacol. 15, 1280792 (2024).
Kezhu W et al. Effects of ginsenoside Rg1 on learning and memory in a reward-directed instrumental conditioning task in chronic restraint stressed rats. [cited 2026 Jan 29].
Zhang, Y. Q., Wang, X. B., Xue, R. R., Gao, X. X. & Li, W. Ginsenoside Rg1 attenuates chronic unpredictable mild stress-induced depressive-like effect via regulating NF-κB/NLRP3 pathway in rats. NeuroReport 30(13), 893 (2019).
Gao, L. et al. Rinsenoside Rg1 and its involvement in hippo–YAP signaling pathway alleviating symptoms of depressive-like behavior. Sci. Rep. 15(1), 14441 (2025).
Kwon, O. Y. & Lee, S. H. Ishige okamurae attenuates neuroinflammation and cognitive deficits in mice intracerebroventricularly injected with LPS via regulating TLR-4/MyD88-dependent pathways. Antioxidants 12(1), 78 (2023).
Kanehisa, M., Furumichi, M., Sato, Y., Matsuura, Y. & Ishiguro-Watanabe, M. KEGG: Biological systems database as a model of the real world. Nucleic Acids Res. 53(D1), D672–D677 (2024).
Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 28(11), 1947–1951 (2019).
Kanehisa, M. & Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. https://doi.org/10.1093/nar/28.1.27 (2000).
Effects of involuntary treadmill running in combination with swimming on adult neurogenesis in an alzheimer’s mouse model. Neurochem Int. 155, 105309 (2022).
Planchez, B., Surget, A. & Belzung, C. Adult hippocampal neurogenesis and antidepressants effects. Curr. Opin. Pharmacol. 1(50), 88–95 (2020).
Schmuckermair, C. et al. Behavioral and neurobiological effects of deep brain stimulation in a mouse model of high anxiety- and depression-like behavior. Neuropsychopharmacology 38(7), 1234–1244 (2013).
Li, M. et al. Lentivirus-mediated interleukin-1β (IL-1β) knock-down in the hippocampus alleviates lipopolysaccharide (LPS)-induced memory deficits and anxiety- and depression-like behaviors in mice. J. Neuroinflamm. 14(1), 190 (2017).
Badshah, H., Ali, T. & Kim, M. O. Osmotin attenuates LPS-induced neuroinflammation and memory impairments via the TLR4/NFκB signaling pathway. Sci. Rep. 6(1), 24493 (2016).
Deyama, S. et al. Resolvin D1 and D2 reverse lipopolysaccharide-induced depression-like behaviors through the mTORC1 signaling pathway. Int. J. Neuropsychopharmacol. 20(7), 575–584 (2017).
Zhao, X. et al. Behavioral, inflammatory and neurochemical disturbances in LPS and UCMS-induced mouse models of depression. Behav. Brain Res. 17(364), 494–502 (2019).
Chugh, D., Nilsson, P., Afjei, S. A., Bakochi, A. & Ekdahl, C. T. Brain inflammation induces post-synaptic changes during early synapse formation in adult-born hippocampal neurons. Exp. Neurol. 1(250), 176–188 (2013).
Time course of the effects of lipopolysaccharide on prepulse inhibition and brain nitrite content in mice. Eur. J. Pharmacol. 713(1–3), 31–38 (2013).
Raetz, C. R. H. & Whitfield, C. Lipopolysaccharide Endotoxins. Annu. Rev. Biochem. 71, 635–700 (2002).
Lawson, M. A. et al. Intracerebroventricular administration of lipopolysaccharide induces indoleamine-2,3-dioxygenase-dependent depression-like behaviors. J. Neuroinflamm. 10(1), 875 (2013).
Wu, Z. et al. Resilience in the LPS-induced acute depressive-like behaviors: Increase of CRMP2 neuroprotection and microtubule dynamics in hippocampus. Brain Res. Bull. 162, 261–270 (2020).
Zhang, B., Xiang, Z., Pan, Z., Chen, B. Formation and characteristics of multi-dimensional tibetan traditional motion from the perspective of data mining. In 2019 Third International Conference on I-SMAC (iot in Social, Mobile, Analytics and Cloud) (I-SMAC) [Internet]. 24–28 2019 [cited 2025 Nov 1].
Guo, L. T. et al. Baicalin ameliorates neuroinflammation-induced depressive-like behavior through inhibition of toll-like receptor 4 expression via the PI3K/AKT/FoxO1 pathway. J. Neuroinflamm. 16(1), 95 (2019).
Couch, Y. et al. Low-dose lipopolysaccharide (LPS) inhibits aggressive and augments depressive behaviours in a chronic mild stress model in mice. J. Neuroinflamm. 13(1), 108 (2016).
Peng, L., Bestard-Lorigados, I. & Song, W. The synapse as a treatment avenue for alzheimer’s disease. Mol. Psychiatry 27(7), 2940–2949 (2022).
Zhang, H. et al. Role of aβ in alzheimer’s-related synaptic dysfunction. Front. Cell. Dev. Biol. 10, 964075 (2022).
Long, Q., Li, T., Zhu, Q., He, L. & Zhao, B. SuanZaoRen decoction alleviates neuronal loss, synaptic damage and ferroptosis of AD via activating DJ-1/Nrf2 signaling pathway. J. Ethnopharmacol. 6(323), 117679 (2024).
Wang, X. et al. Leucine mediates cognitive dysfunction in early life stress-induced mental disorders by activating autophagy. Front. Cell. Neurosci. 16, 1060712 (2023).
Lu, Y. et al. Baicalin regulates depression behavior in mice exposed to chronic mild stress via the rac/LIMK/cofilin pathway. Biomed. Pharmacother. 1(116), 109054 (2019).
Frodl, T. et al. Hippocampal changes in patients with a first episode of major depression. Am. J. Psychiatry 159(7), 1112–1118 (2002).
Schmidt, H. D. & Duman, R. S. The role of neurotrophic factors in adult hippocampal neurogenesis, antidepressant treatments and animal models of depressive-like behavior. Behav. Pharmacol. 18(5–6), 391 (2007).
Herbet, M. et al. α-tocopherol ameliorates redox equilibrium and reduces inflammatory response caused by chronic variable stress. Biomed. Res. Int. 2018(1), 7210783 (2018).
Gao H et al. Anti-depressant-like effect of atractylenolide I in a mouse model of depression induced by chronic unpredictable mild stress. Exp. Ther. Med. 2017 Nov 16 [cited 2025 Oct 31].
Anisman, H. & Merali, Z. Cytokines, stress and depressive illness: brain-immune interactions. Ann. Med. 35(1), 2–11 (2003).
Wang, Y., Xu, J., Liu, Y., Li, Z. & Li, X. TLR4-NF-κB signal involved in depressive-like behaviors and cytokine expression of frontal cortex and hippocampus in stressed C57BL/6 and ob/ob mice. Neural Plast. 2018(1), 7254016 (2018).
Ma, K., Zhang, H. & Baloch, Z. Pathogenetic and therapeutic applications of tumor necrosis factor-α (TNF-α) in major depressive disorder: A systematic review. Int. J. Mol. Sci. 17(5), 733 (2016).
Yamanashi, T. et al. Beta-hydroxybutyrate, an endogenic NLRP3 inflammasome inhibitor, attenuates stress-induced behavioral and inflammatory responses. Sci. Rep. 7(1), 7677 (2017).
Tonelli, L. H. & Postolache, T. T. Tumor necrosis factor alpha, interleukin-1 beta, interleukin-6 and major histocompatibility complex molecules in the normal brain and after peripheral immune challenge. Neurol. Res. 27(7), 679–684 (2005).
Howren, M. B., Lamkin, D. M. & Suls, J. Associations of depression with C-reactive protein, IL-1, and IL-6: A meta-analysis. Psychosom. Med. 71(2), 171 (2009).
Bay-Richter, C., Janelidze, S., Hallberg, L. & Brundin, L. Changes in behaviour and cytokine expression upon a peripheral immune challenge. Behav. Brain Res. 222(1), 193–199 (2011).
Yang, P. et al. Changes in proinflammatory cytokines and white matter in chronically stressed rats. Neuropsychiatr. Dis. Treat. 6(11), 597–607 (2015).
You, Z. et al. Pro- and anti-inflammatory cytokines expression in rat’s brain and spleen exposed to chronic mild stress: Involvement in depression. Behav. Brain Res. 225(1), 135–141 (2011).
Miller, A. H., Maletic, V. & Raison, C. L. Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression. Biol. Psychiatry. 65(9), 732–741 (2009).
Mahdirejei, H. A., Peeri, M., Azarbayjani, M. A. & Fattahi, M. F. Fluoxetine combined with swimming exercise synergistically reduces lipopolysaccharide-induced depressive-like behavior by normalizing the HPA axis and brain inflammation in mice. Pharmacol. Biochem. Behav. 1(232), 173640 (2023).
Zhao, Y. et al. Chronic stress dysregulates the hippo/YAP/14-3-3η pathway and induces mitochondrial damage in basolateral amygdala in a mouse model of depression. Theranostics. 14(9), 3653–3673 (2024).
Houtenbos, S. P. et al. The underlying mechanisms of the association of bone health with depression – an experimental study. Mol Biol Rep. 52(1), 163 (2025).
Stepan, J., Anderzhanova, E. & Gassen, N. C. Hippo signaling: emerging pathway in stress-related psychiatric disorders?. Front. Psychiatry. 9, 715 (2018).
Zarka, M., Haÿ, E. & Cohen-Solal, M. YAP/TAZ in bone and cartilage biology. Front. Cell. Dev. Biol. 9, 788773 (2022).
Bang, L. et al. Genome-wide analysis of anorexia nervosa and major psychiatric disorders and related traits reveals genetic overlap and identifies novel risk loci for anorexia nervosa. Transl. Psychiatry. 13(1), 291 (2023).
Yu, Y. et al. Yes-associated protein and transcriptional coactivator with PDZ-binding motif as new targets in cardiovascular diseases. Pharmacol Res. 159, 105009 (2020).
Chen, S. et al. Electroacupuncture alleviates the symptom of depression in mice by regulating the cGAS-STING-NLRP3 signaling. Aging (Milano). 16(8), 6731–6744 (2024).
Acknowledgements
We sincerely thank draw.io (https://www.drawio.com/) and Scidraw (https://scidraw.io/) for the graphic materials in this article free of charge. We also appreciate the support of the UniProt database.
Funding
The experiment was supported by the Research Foundation of Hebei Provincial Key Research and Development Program (grant number: 223777154D), China University Industry-University-Research Innovation Fund (grant number: 2024GY036), Government-Funded Clinical Medical Talent Project (grant number: ZF2023166 and grant number: ZF2024157), and Scientific Research Plan of Hebei Provincial Administration of Traditional Chinese Medicine (grant number: Z2026004).
Author information
Authors and Affiliations
Contributions
Lan Zhang: conceptualization, methodology, experiments, writing—review & editing, and formal analysis. Na Wang and Hu Chen: writing—review & editing, methodology, experiments, software and experiments; Jiachen Tang: methodology, experiments, validation and formal analysis; Zixin Fang and Gang Wang: software and resources; Lin Pei: visualization, project administration, and funding acquisition. All the authors read, discussed, and agreed to the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
All experimental protocols were approved by the the Animal Experiment Ethics Committee of Hebei University of Chinese Medicine (DWLL202302019).
Consent for publication
All authors agree to publish in this journal.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, L., Tang, J., Fang, Z. et al. Ginsenoside Rg1 ameliorates lipopolysaccharide-induced depressive-like behaviors in mice by attenuating neuroinflammation and neuronal damage. Sci Rep 16, 11948 (2026). https://doi.org/10.1038/s41598-026-41622-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-41622-z
