
Abstract
Female ear development during the pre- and post-silking period is critical for maize yield. Although plant growth regulators can enhance yield, their specific effects on ear morphogenesis and underlying mechanisms are not fully understood. In this study, two maize hybrids were foliar-treated with uniconazole (UCZ, 25 mg L⁻¹) or 5-aminolevulinic acid (ALA, 40 mg L⁻¹) at the 12-leaf stage. UCZ and ALA increased grain yield by 6.8% to 12.0% and 8.3% to 13.9%, respectively. This yield improvement resulted not only from increased 100-grain weight but, more importantly, from a greater kernel number per ear achieved through enhanced row number (by UCZ) and grains per row (by ALA). Morphological analysis revealed that UCZ significantly increased ear diameter, whereas ALA markedly promoted ear elongation. Physiologically, UCZ lowered IAA and GA levels but raised ABA, while ALA exerted opposite effects on these hormones. Both regulators enhanced ZR and carbohydrate contents. In conclusion, UCZ and ALA enhance maize yield through distinct pathways: UCZ favors radial growth and yield components related to ear thickness, whereas ALA promotes longitudinal growth and components related to ear length.
Similar content being viewed by others

Introduction
The female ear is a key reproductive organ whose morphology directly affects the maize yield1,2. Silking stage is the transitional period from vegetative growth to reproductive growth and is the key period for ear morphogenesis and the formation of yield in maize3,4,5. Cob accounts for more than half of ear tissue of maize and connects nutrition source tissues to developing the grain of ear store which such development during silking strongly influences the morphological formation of ear6,7.
The architecture of the maize ear is governed by a sophisticated phytohormonal network8. This network involves growth-promoting hormones like IAA and GA9, growth-inhibiting ABA10, and differentiation-regulating CTK11. Ultimately, the final morphology and yield potential are shaped by the dynamic balance of antagonistic and synergistic interactions among these hormones.
Structural carbohydrates are mostly used for morphogenesis of plants, while non-structural carbohydrates are mostly involved in plant metabolic activity. Maize cobs are rich in structural carbohydrates, including approximately 45% cellulose, 35% hemicellulose and 15% lignin4. Non-structural Carbohydrates include soluble sugars (i.e., sucrose and fructose) and insoluble substance (i.e., starch)12. Soluble carbohydrates supply energy and metabolic intermediates for plant growth and development13,14. Starch hydrolysis could yields soluble carbohydrates to provide energy for maize ear development15,16.
Growth retardants temporarily slow meristematic cell division and elongation, and resume normal growth after a period of time. UCZ is a typical plant growth retardant that inhibits GA bioisynthesis17. By suppressing the biosynthesis of GA, UCZ alters the endogenous hormonal balance and leading to morphological changes via inhibiting the vertical growth and promoting the horizontal growth of cells18. Agronomically, UCZ is mainly applied to delay plant growth and dwarfing to enhance reproductive growth, storage organ development, and ultimately increase yield. Foliar-treated with UCZ was found to reduces plant and ear height while thickening the stem and lignin biosynthesis, thereby reducing lodging risk and increasing yield19. Moreover, research also found that seed dressing with UCZ enlarges ear of maize diameter and increases yield under dense planting conditions20.
On the other hand, plant growth promoter stimulate cell division, differentiation, elongation and overall vegetative and reproductive organs development. ALA is a biodegradable, non-toxic growth promoter and precursor of porphyrin compounds biosynthetic21,22. ALA is a natural substance in plants and can be synthesized biologically or chemically23. Agriculturally, ALA regulates growth, development, yield and improve stress resistance24,25. In vitro, ALA-supplemented MS media promote the differentiation of adventitious roots and adventitious shoots of callus, showing the dual regulatory properties of IAA and CTK26. ALA can also improve wheat ear morphology, and increase the ear length, ear diameter, ear weight, ear surface area, 100-grain weight and yield25. ALA likewise enhances growth and yield in economic crops such as tomato27, potato24, garlic24, cucumbers28, and rape29.
Although UCZ and ALA are widely applied and known to improve the morphology and yield of crops, their specific roles in maize ear morphogenesis remain under-reported. In this field-based study, two regulators were sprayed at 12-leaf stage, and their regulation of ear morphology was investigated from the aspects of yield, morphology and physiology. This study aimed to investigate the effects and underlying mechanisms of UCZ and ALA on female ear morphogenesis in maize, providing a theoretical basis for their targeted application to improve yield.
Materials and methods
Experimental site and materials
The field trials were conducted in 2020–2021 at Daqing Branch of Heilongjiang Academy of Agricultural Sciences, Northeast China (46°40’N, 125°14’E) with temperate continental monsoon climate. The annual sunshine hours are 2600–2900, with 1300–1350 occurring during the growing period (May to October). The annual precipitation is ~ 400 mm, 85% of which falls in the growing season. Mean monthly temperature and precipitation profiles are presented in Fig. 1. The experimental soil type is weakly alkaline chernozem. Topsoil (0–20 cm) were: organic matter 26.2 g kg–1; total nitrogen (N) 2.9 g kg–1; available phosphorus (P) 0.0177 g kg–1; available potassium (K) 0.119 g kg–1; PH 8.3; humus 0.8%; water content 14.1%; saturated moisture 37.7%; bulk density 1200 g L–1.
Two widely cultivated hybrids, XY335 and ND18, were supplied by the Heilongjiang Academy of Agricultural Sciences. XY335: 3500–4000 plants per 667 square meter, 127-day growth cycle, 2700 °C growing degree days (GDD, base 10 °C), 19 leaves. ND18: 4500–5000 plants per 667 square meter, 125-day growth cycle, 2600 °C GDD, 17 leaves. UCZ and ALA were provided by the Chemical Control Laboratory of Heilongjiang Bayi Agricultural University.

Mean temperature (℃) and rainfall (mm) during the two cultivation seasons.
Experiment design
The maize varieties XY335 and ND18 were sown in early May and harvested in late September or early October. XY335 was cultivated with 60,000 plants per hectare, while ND18 was cultivated with 75,000 plants per hectare, according to the planting density of the production field. Field management measures (i.e., fertilization and weeding) were consistent with local conventional practice.
At the 12-leaf stage, UCZ and ALA were applied at 225 L ha⁻¹. Based on prior optimization30, spraying concentrations were 25 mg L–1 UCZ and 40 mg L–1 ALA; the control received tap water. Each area covered 62 m2 (8 rows × 12 m length, and 0.65 m row spacing). Treatments were arranged in a randomised complete block design with three replicates. Sampling began around the silking stage of maize, and the sampling time was controlled between 8:00 and 10:00 in the morning.
Sample collection and measurement
Morphology of female ear
Female ears were sampled at heading stage (VT), silking stage (R1), one week after silking stage (R2) and two weeks after silking stage (R3), according to their respective growth process of the two varieties. 10 representative plants per plot were selected. Ear length, diameter, and cob diameter were measured with a vernier caliper; fresh ear and cob masses were recorded with an balance. Samples were dried to constant weight at 80 ◦C in a force-draft oven (101–2SB, Lvbo Instrument Co.Ltd, Hangzhou, China) to obtain dry weight.
Biochemical analyses
For biochemical analyses, cob tissues were excised from the sampled ears, immediately frozen in liquid nitrogen, and stored at − 80 °C until analysis. For hormone quantification, frozen cob tissue (0.5 g) in two varieties were grinded with 2 ml 80% methanol. The supernatant (i.e., methanol extract), was dried under nitrogen to 3 ml after centrifugating. Indole-3-acetic acid (IAA), gibberellin GA₃ (hereafter referred to as GA for brevity), abscisic acid (ABA), and zeatin riboside (ZR) were quantified by ELISA31. The ELISA kit was provided by the Chemical Control Center of China Agricultural University. Structural carbohydrates (cellulose, hemicellulose, and lignin) were also analyzed from the frozen tissue. Cellulose and hemicellulose contents were determined according to Hussain et al.32, and lignin content was determined according to the method of Hussain et al.32 and Ahmad et al.19. Non-structural carbohydrates (starch, sucrose, fructose, and soluble sugars) in the cob tissue were analyzed. Starch content was determined by enthrone colorimetry according to the method of Yemm and Willi33; Sucrose and fructose content was determined according to the method of Wilson et al.34; and soluble sugar content was determined by enthrone colorimetry according to the method of Hu et al.35.
Grain yield
The grain yield and yield components (i.e., row number per ear, grain number per row, 100–grain mass) were obtained through measuring the 10 representative ears of maize in each plot at maturity stage. For the determination of 100-grain mass, grains were shelled from the middle region of each ear, pooled by plot, and a random sample of 100 grains from this pooled lot was weighed. Meanwhile, yield were corrected to 14.0% moisture.
Statistical analysis
Experimental data were organised in Microsoft Excel 2010, graphed with Origin 20.0 (Origin Lab Corporation, USA) and processed with Statistical Product and Service Solutions (SPSS20.0, IBM Corporation, USA). Analysis of variance (ANOVA) was performed, and where significant effects were found, means were compared using Duncan’s multiple range test at p < 0.05. The letter a and b represent the significant differences at the level of p < 0.05. Asterisks (*, **) denote significance at p < 0.05 and 0.01, respectively. Results were presented as mean ± standard deviation (SD) with measurement at least three times. Pearson’s correlations were calculated to assess relationship among hormone, carbohydrate and ear morphology. Pearson’s correlations (with n representing the total number of observations across replicates and sampling stages) were calculated to assess relationships among hormones, carbohydrates, and ear morphology.
Results
Grain yield
Across two–year experiment, UCZ and ALA treatments significantly increased the grain yield in XY335 and ND18 relative to the control (Table 1). No significant year × varieties × treatment interaction effects on grain yield (Table 1) was detected. Grain yield increased from grain number improvement per ear and grain weight per ear by both regulators. Interestingly, UCZ achieved higher grain number per ear by elevating row number per ear, while ALA did so by increasing grain number per row (Table 1).
Size of ear and cob
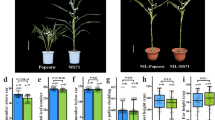
Ear elongation and radial expansion were the most dynamic from heading stage to two weeks after the silking stage. As shown in Fig. 2, the two regulators had significant regulatory effects on the size of ear of the two varieties, but in contrasting ways. No significant year × varieties × treatment interaction effects on ear length, ear diameter and cob diameter was observed (Table 2). UCZ reduced ear length but increased ear and cob diameters, whereas ALA increased ear length without significantly affecting diameters (Fig. 2).

Effect of UCZ and ALA treatments on the size of ear and cob in maize.
Weight of ear and cob
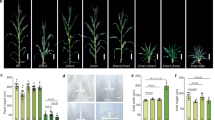
The fresh and dry weight of ears and cobs increased rapidly from VT to R3 in both hybrids by both regulators (Fig. 3). The year × varieties × treatment interaction effects on fresh weight of ear and cob, dry weight of cob were not significant (Table 2). Both regulators significantly increased the fresh and dry weights of ear and cob from VT to R3, with ALA generally showing greater increases than UCZ (Fig. 3).

Effect of UCZ and ALA treatments on the weight of ear and cob in maize.
Hormone
UCZ and ALA could effectively regulate the level of endogenous hormones from VT to R3, but their regulatory effects were different (Fig. 4). IAA levels increased from VT to R2 in both hybrids, with XY335 showing a continued steady increase to R3 and ND18 a decline at R3. In both hybrids, UCZ significantly reduced IAA levels during key developmental stages (R1–R3 in XY335; VT, R2, R3 in ND18), whereas ALA significantly increased IAA levels, particularly at VT and R2 (Fig. 4). IAA showed a positive correlation with ear length (p<0.05; Table 3). GA exhibits pattern similar to IAA in XY335 and ND18. In contrast to UCZ, which significantly suppressed GA levels (at VT and R2 in XY335; from VT to R3 in ND18), ALA markedly elevated them from VT to R1 in both hybrids (Fig. 4). The GA content also showed a positive, though not statistically significant, correlation with ear length (Table 3).ABA increase from VT to R1 stages and declined from R1 to R3 stages thereafter in both hybrids (Fig. 4). Conversely, UCZ significantly elevated ABA levels (at R1 and R3 in XY335; at VT and R1 in ND18), while ALA markedly reduced them (at R1 and R2 in XY335; from VT to R3 in ND18). ZR levels exhibited a common pattern of initial increase followed by a decline in both hybrids, with the peak occurring at R1 in XY335 and at R2 in ND18 (Fig. 4). Both UCZ and ALA treatments significantly increased ZR levels in the two hybrids, with UCZ elevating ZR from R1 to R3 in XY335 and from VT to R3 in ND18, and ALA showing a broader promotive effect from VT to R3 in both hybrids. ZR showed a positive correlation with ear diameter (p<0.05; Table 3).

Effects of UCZ and ALA treatments on hormones of cob in maize.

Effects of UCZ and ALA treatments on structural carbohydrate of cob in maize.
Structural carbohydrate
Structural carbohydrates are important components of ear cob. Cellulose, hemicellulose, and lignin correlated positively with ear morphology (Table 4). As shown in the Fig. 5, both regulators altered structural carbohydrates content in ear from VT to R3, but with different regulatory effects. Cellulose of ear of two varieties peaked at R2 stages, and then declined from R2 to R3 stages (Fig. 5). UCZ increased cellulose content in XY335 and ND18, while ALA lowered cellulose content at the R2 stage.
The hemicellulose of ear of two varieties in all treatments increased gradually from VT to R1 stages, and then decreased from R1 to R3 stages (Fig. 5). In XY335, UCZ raised hemicellulose content at R1 whereas ALA reduced it at R2; in ND18, UCZ increased hemicellulose at VT and R1 while ALA decreased it.
Lignin of ear increase to R2 in XY335 in all treatments and R3 in ND18.In XY335, UCZ increased lignin content at R1 and R2, while ALA decreased it at R3; In ND18, UCZ consistently raised lignin from VT to R3, whereas ALA had no significant effect (Fig. 5).
Non-structural carbohydrate
Non-structural carbohydrates are the substrates for ear morphogenesis. Both regulators increase non-structural carbohydrates in ear from VT to R3 (Fig. 6). Starch content of ear of XY335 and ND18 increased gradually from VT to R1 stages, and then decreased from R1 to R3 stages (Fig. 6). In both hybrids, starch content was increased by UCZ and ALA treatments, with XY335 showing a rise from R1 to R2 followed by a slight decline at R3 under ALA, while ND18 exhibited elevated starch levels at VT, R1 and R3 stages. Moreover, correlation analysis revealed that starch was extremely significantly negatively (p<0.01) correlated with ear morphology (Table 4).
Sucrose content of ear increases gradually from VT to R1 stages of XY335 and ND18 in all treatments. Sucrose content in XY335 was significantly raised by UCZ and ALA from VT to R2 stages. Similarly, sucrose content in ND18 was significantly increased by UCZ from VT to R2 and by ALA at R2. Sucrose correlated significantly positively with ear morphology (p < 0.01; Table 4).
As shown in Fig. 6, fructose responded strongly at R1. In XY335, UCZ and ALA markedly increased fructose content at R1, while UCZ also raised it at R3. In ND18, fructose content rose from VT to R2 under both regulators, then declined from R2 to R3. (Fig. 6). Fructose correlated positively with ear morphology (Table 4).
Soluble sugars content of ear of XY335 and ND18 in all treatments increased slowly from VT to R3 stages. Both UCZ and ALA elevated soluble sugars in the two hybrids, with increases observed in XY335 and similar increases from R1 to R3 in ND18 (Fig. 6). Soluble sugars correlated positively with ear traits (p < 0.01; Table 4).

Effects of UCZ and ALA treatments on non-structural carbohydrate of cob in maize.
Discussion
Grain yield and female ear morphology
Enhancing yield components including row number per ear, grain number per row and grain weight can directly elevates maize grain yield36,37. Both the growth regulators (i.e., 6-benzyladenine, diethyl aminoethyl hexanoate and kinetin) and growth retardants (i.g., UCZ, paclobutrazol) are widely applied to raise grain number per ear and grain weight of maize19,36,38,39,40. Recently, Wang et al. demonstrated that the plant growth regulator EDAH could increase the yield of summer maize under high planting density primarily by improving ear differentiation and fertilization rate, which aligns that morphological modification of the ear is a key pathway for yield improvement41. In this study, both regulators (UCZ and ALA) increased maize yield by improving the grain number per ear and grain weight. Mechanistically, UCZ increased the grain number per ear via additional rows per ear, whereas ALA did so by elevating grain number per row (Table 1), underscoring their distinct modes of action.
Female ear morphology is tightly coupled to yield components, and thus serves as a direct proxy for final yield2. Ear length correlates positively with grain number per row, grain number per ear and yield42,43, whereas ear diameter positively correlated with row number per ear, grain number per ear and yield44. Both promotive regulators and delayed regulators can modify ear morphology. For instance, exogenous application of ethylene (a retardant) inhibits ear elongation yet significantly increases ear diameter, grain weight and yield45. Conversely, exogenous foliar spraying of 6-benzyladenine (a promoter) enlarges ear length, diameter, volume, weight and yield of maize36,46. Consistent with these reports, UCZ (delayed regulator) increased the diameter and mass of ear and cob, whereas ALA (promotive regulator) lengthened ears and boosted biomass at silking. In summary, both UCZ and ALA reshape ear architecture through distinct pathways, ultimately enhancing maize yield.
Hormones
Endogenous hormones play an important role in plant growth, development and morphogenesis, which were often manipulated by plant-growth regulators in agriculture. IAA, the first-identified phytohormone, promotes ear morphological development by increasing length, diameter, row number, grain number and grain weight47. Recent molecular evidence has further solidified this view, as demonstrated by Liu et al. who identified that the YIGE2 gene regulates maize ear length and yield by modulating the auxin signaling pathway48. Consistently, IAA showed positive correlation with ear length in the present study (Table 3). UCZ and ALA altered IAA in opposite directions. Increasing exogenous UCZ concentration lowered endogenous IAA content in rapeseed, thereby suppressing shoot elongation49. By contrast, exogenous ALA increased endogenous hormone IAA in cucumber and promoted shoot and root elongation29. In this study, exogenous application of UCZ down-regulated endogenous IAA and shortened ear length. In contrast, exogenous application of ALA significantly raised endogenous IAA lengthened ear from 5.3% to 8.4%. Crucially, this promotive effect on elongation was supported by a concomitant increase in soluble sugars and sucrose (Fig. 6), which are essential substrates for cell expansion and biosynthesis. This suggests that ALA coordinates hormonal signals with enhanced carbon availability to drive ear elongation.
GA promotes cell elongation, and works synergistically with IAA to drive vertical growth50. GA biosynthesis predominates in young and vigorous meristematic tissues such as root and shoot tips51. Studies have found disruption of GA homeostasis produces rice dwarfism and decreases ears length52. Similarly, our results indicated a positive trend between GA levels and ear length in maize (Table 3). UCZ, a GA inhibitor, suppresses longitudinal growth53. Consistently, exogenous UCZ down-regulated GA in ear and restricted ear elongation. On the contrary, plant growth regulators including salicylic acid, 6-benzyladenine, ALA can increase endogenous GA, and stimulate maize growth and development54,55. Here, ALA up-regulated GA and promoted longitudinal elongation of ear, in line with prior reports.
ABA is a antagonizes growth-promoting hormones10. ABA is synthesized mainly in root caps, senescent leaves, seeds and fruits56,57,58. UCZ raises ABA in rapeseed, soybean and maize49,53,59, whereas ALA lowers endogenous ABA in rapeseed and maize29,54, leading to divergent morphogenic outcomes. Our findings mirrored those patterns which UCZ elevated endogenous ABA in the ear, whereas ALA reduced it.
CTK stimulate cell division, expansion and differentiation60. Both UCZ and ALA enhance endogenous CTK synthesis, thereby altering plant morphogenesis. UCZ treatment significantly increased CTK levels in maize and rapeseed stems, promoting radial cell expansion and stems diameter49,53. In this study, UCZ increased endogenous ZR in the ear, correlating with increased cob and ear diameter. The up-regulation of ZR, a cytokinin known to stimulate cell division and sink strength, likely established a stronger metabolic ‘sink’ in the developing ear. This enhanced sink demand is coherent with our observation of increased deposition of structural carbohydrates like cellulose and lignin (Fig. 5). Consequently, the synergistic action of elevated ZR and reinforced carbon partitioning into structural compounds underpinned the radial expansion and thickening of the cob and ear diameter, a phenomenon also observed in the regulation of ear development by other hormonal signals61. Exogenous ALA mimics CTK action by inducing adventitious root and bud differentiation in callus26. Here, exogenous ALA likewise boosted endogenous ZR in ear, corroborating with the results of Naeem et al.29 and Xu54.
Carbohydrates
More than half of the tissue of maize ear is cob around silking stage7. Structural carbohydrate are the most indispensable components of cob, including cellulose, hemicellulose and lignin, and which contents significantly affect the morphology of ear of maize62,63. In this study, it was found that cellulose, hemicellulose and lignin were positively correlated with the length, diameter and weight of ear and cob of maize (Table 4). Plant growth regulators can modify morphology by altering structural-carbohydrate deposition. UCZ, a retardant, increased the diameter and strength of maize and rice stalks by elevating structural carbohydrates40,64. Conversely, jasmonic acid (a promoter) enhances hemp hypocotyls secondary growth (diameter and mass increment) by boosting cellulose and lignin65. In this study, UCZ increased cob diameter and weight, likely via structural-carbohydrate up-regulation. ALA lengthen and thickened cobs yet reduced the content of structural carbohydrates in ear at certain stages. In the results of this experiment, although ALA treatment promoted the accumulation of non-structural carbohydrate content and provided more substrates for the synthesis of structural carbohydrates (Fig. 5), it accelerated the elongation rate of cob, resulting in the dilution of structural carbohydrate content per unit length of cob at certain stages, which is consistent with Wu18.
Non-structural carbohydrates are necessary substrates for the morphogenesis of maize ear. Sucrose, the principal soluble carbohydrate, supplies energy and acts as a signaling molecule14,66. Fructose is also a soluble carbohydrate and is the main temporary storage form of plant nutrient tissue, which can store organic matter for a short time67,68. In this study, it was found that soluble sugar, sucrose and fructose were positively correlated with morphology of maize ear (Table 4). Notably, the fructose content was substantially lower than the total soluble sugar content (Fig. 6), which aligns with the metabolic characteristics of maize ear cobs as an active sink organ. In such tissues, sucrose serves as the primary transported and metabolized sugar form, consistent with our data showing that sucrose was the predominant single sugar component (Fig. 6). Fructose and glucose are mainly derived from the enzymatic cleavage of sucrose in situ. In actively metabolizing sink tissues, imported sucrose is rapidly metabolized or converted into other compounds, resulting in relatively low steady-state levels of free fructose monomers. Previous study of our group on the “source” organs of maize found that exogenous UCZ and ALA could increase the photosynthetic efficiency and soluble sugar content of maize leaves, and increase the size and weight of maize ear at harvest30. This study on the “sink” organs further demonstrates that exogenous foliar-treated with UCZ and ALA increased the contents of soluble sugar, sucrose and fructose of ear, contributing to improved the morphology of ear around silking.
Starch is the primary long-term carbohydrate reserve68. Post-silking, starch is decomposed into soluble sugar through starch and sucrose metabolism pathway69, which provides energy for the rapid growth of ear. Plant growth regulators can increase the accumulation of starch content and accelerate starch hydrolysis during the critical period of plant morphogenesis69,70. We previously reported that exogenous foliar-treated with UCZ elevated starch and sucrose in soybean leaves, pods and grains, and the contents of starch and sucrose in different organs were positively correlated71. ALA enhances amylase activity in radish main roots, promoting starch hydrolysis and conversion to glucose while raising total starch, increasing the fresh weight of aboveground and underground main roots70. Our results found that exogenous foliar-treated with UCZ and ALA increased the starch content of maize ear in varying degrees with ALA had stronger ability to hydrolyze starch70. Consistent with this hydrolytic capacity, the observed decrease in starch content at the R3 stage (Fig. 6) likely corresponds to the peak period of grain filling, during which enhanced starch hydrolysis into soluble sugars provides necessary substrates for ear morphogenesis. Thus, both UCZ and ALA increase the accumulation of non-structural carbohydrates in maize ear, providing more substrate sources for the morphogenesis of ear. It should be noted that although cob fresh and dry weights increased from R1 to R3 (Fig. 2), the contents of cellulose and hemicellulose decreased from R2 to R3 (Fig. 5). This can be explained by two factors. First, structural carbohydrate synthesis likely peaked during the rapid cell-wall construction phase (R1–R2), slowing thereafter relative to overall growth. Second, R3 marks the onset of rapid grain filling, during which the cob accumulates substantial non-structural carbohydrates (e.g., sucrose, starch) for kernel development. These compounds significantly contribute to cob biomass at R3, thereby diluting the proportional content of structural carbohydrates. Thus, the observed weight increase at R3 is largely driven by enhanced non-structural carbohydrate accumulation, consistent with the role of both regulators in strengthening sink activity during reproductive growth.
Limitations and Future Perspectives
This study has certain limitations. The findings are based on two widely used hybrids, and their applicability to a broader genetic range requires further validation. Moreover, the physiological insights presented here would benefit from future transcriptomic and metabolomic analyses to elucidate the underlying gene regulatory networks.
Conclusion
UCZ (delayed regulator) and ALA (promotive regulator) regulated maize ear morphogenesis differently due to their different mechanisms of action. UCZ lowered IAA and GA, restricting ear elongation, whereas ALA elevated these hormones and promoted vertical growth. UCZ up-regulated endogenous ZR, which may promoted cell lateral division and was the main factor leading to the increase of ear diameter. Moreover, UCZ also increased the content of structural carbohydratesin ear, and promoted the formation and diameter of cob; ALA had little effect on these polymers or cob diameter. Both regulators boosted non-structural carbohydrates, supplying metabolic substrates for ear morphogenesis. In summary, UCZ improves yield by producing shorter, thicker ears with increased row number, whereas ALA enhances yield through ear elongation accompanied by increased grain number per row. These distinct mechanisms provide a precise physiological basis for the targeted application of plant growth regulators in maize cultivation.
Data availability
The data presented in this study are available on request from the corresponding author.
References
Li, M. et al. Mapping QTLs for grain yield and yield components under high and low phosphorus treatments in maize (Zea mays L). Plant. Sci. 178, 454–462 (2010).
Tucker, S. L. et al. Evaluating maize phenotypic variance, heritability, and yield relationships at multiple biological scales across agronomically relevant environments. Plant. Cell. Environ. 43, 880–902 (2020).
Cantarero, M. G., Cirilo, A. G. & Andrade, F. H. Night temperature at silking affects kernel set in maize. Crop Sci. 39, 703–710 (1999).
Early, E. B., Mcilrath, W. O., Seif, R. D. & Hageman, R. Effects of shade applied at different stages of plant development on corn (Zea mays L.) production. Crop Sci. 7 (2), 151–156 (1967).
Pan, X. et al. Asymmetric transcriptomic signatures between the cob and florets in the maize ear under optimal- and low-nitrogen conditions at silking, and functional characterization of amino acid transporters ZmAAP4 and ZmVAAT3. J. Exp. Bot. 66, 6149–6166 (2015).
Laubengayer, R. A. The vascular anatomy of the eight-rowed ear and tassel of golden rantam sweet corn. Am. J. Bot. 36 (2), 236–244 (1949).
Seebauer, J. R., Moose, S. P., Fabbri, B. J., Crossland, L. D. & Below, F. E. Amino acid metabolism in maize earshoots. Implications for assimilate preconditioning and nitrogen signaling. Plant. Physiol. 136, 4326–4334 (2004).
Wolters, H. & Jurgens, G. Survival of the flexible: Hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 10, 305–317 (2009).
Bommineni, V. R. & Greyson, R. Effect of gibberellic acid and indole-3-acetic acid on growth and differentiation of cultured ear inflorescences of maize (Zea mays L). Plant Sci. 68 (2), 239–247 (1990).
Bentsink, L. & Koornneef, M. Seed dormancy and germination. Arabidopsis Book. 6, e119 (2008).
Ioio, R. D. et al. Genetic framework for the control of cell division and differentiation in the root meristem. Sci. (American Association Advancement Science). 322, 1380–1384 (2008).
Yee, D. A. & Tissue, D. T. Relationships between non structural carbohydrate concentration and flowering in a subtropical herb, Heliconia caribaea (Heliconiaceae). Caribb. J. Sci. 41 (2), 243–249 (2005).
Couée, I., Sulmon, C., Gouesbet, G. & El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Eep Bot. 57, 449–459 (2006).
Yuan, H. et al. Mass spectrometry-based method to investigate the natural selectivity of sucrose as the sugar transport form for plants. Carbohyd Res. 407, 5–9 (2015).
Zeeman, S. C., Kossmann, J. & Smith, A. M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant. Biol. 61, 209–234 (2010).
Silver, D. M., Kötting, O. & Moorhead, G. B. G. Phosphoglucan phosphatase function sheds light on starch degradation. Trends Plant. Sci. 19, 471–478 (2014).
Fletcher, R. A. & Hofstra, G. Improvement of uniconazole-induced protection in wheat seedlings. J. Plant. Growth Regul. 9, 207–212 (1990).
Wu, L. M. et al. Shading contributes to the reduction of stem mechanical strength by decreasing cell wall synthesis in Japonica rice (Oryza sativa L). Front. Plant Sci. 8, 881 (2017).
Ahmad, I. et al. Uniconazole application strategies to improve lignin biosynthesis, lodging resistance and production of maize in semiarid regions. Field Crop Res. 222, 66–77 (2018).
Ahmad, I. et al. Uniconazole and nitrogen fertilization trigger photosynthesis and chlorophyll fluorescence, and delay leaf senescence in maize at a high population density. Photosynthetica 59, 192–202 (2021).
Reinbothe, S. & Reinbothe, C. Regulation of chlorophyll biosynthesis in angiosperms. Plant Physiol. 111, 1–7 (1996).
Sasikala, C., Ramana, C. V. & Rao, P. R. 5-Aminolevulinic acid: A potential herbicide/insecticide from microorganisms. Biotechnol. Prog. 10 (5), 451–459 (1994).
Von, W. D., Gough, S. & Kannangara, C. G. Chlorophyll biosynthesis. Plant. Cell. 7, 1039–1057 (1995).
Hotta, Y., Tanaka, T., Takaoka, H., Takeuchi, Y. & Konnai, M. Promotive effects of 5-aminolevulinic acid on the yield of several crops. Plant. Growth Regul. 22, 109–114 (1997).
Al-Thabet, S. Promotive effect of 5-aminolevulinic acid on growth and yield of wheat grown under dry conditions. Agronomy 5 (1), 45–49 (2006).
Bindu, R. C. B. U. & Vivekanandan, M. Hormonal activities of 5-aminolevulinic acid in callus induction and micropropagation. Plant. Growth Regul. 26, 15–18 (1998).
Zhao, Y. Y. et al. Effects of exogenous 5-aminolevulinic acid on photosynthesis, stomatal conductance, transpiration rate, and PIP gene expression of tomato seedlings subject to salinity stress. Genet. Mol. Res. 14, 6401–6412 (2015).
Anwar, A., Yan, Y., Liu, Y., Li, Y. & Yu, X. 5-aminolevulinic acid improves nutrient uptake and endogenous hormone accumulation, enhancing low-temperature stress tolerance in cucumbers. Int. J. Mol. Sci. 19, 3379 (2018).
Naeem, M. S. et al. 5-Aminolevulinic acid ameliorates salinity-induced metabolic, water-related and biochemical changes in Brassica napus L. Actaphysiol Plant. 33, 517–528 (2011).
Xu, L. et al. Both uniconazole and 5-aminolevulinic acid increase maize (Zea mays L.) yield by changing its ear morphology and increasing photosynthetic efficiency and antioxidants in saline-alkali land. Photosynthetica 60, 408–419 (2022).
Yang, J., Zhang, J., Wang, Z., Zhu, Q. & Wang, W. Hormonal changes in the grains of rice subjected to water stress during grain filling. Plant. Physiol. 127, 315–323 (2001).
Hussain, S. et al. Effects of lignin, cellulose, hemicellulose, sucrose and monosaccharide carbohydrates on soybean physical stem strength and yield in intercropping. Photochem. Photobiol Sci. 19, 462–472 (2020).
Yemm, E. W. & Willis, A. J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 57, 508–514 (1954).
Wilson, R., Cataldo, A. & Andersen, C. P. Determination of total non-structural carbohydrates in tree species by highperformance anion-exchange chromatography with pulsed amperometric detection. Can. J. For. Res. 25, 2022–2028 (1995).
Hu, L. P. et al. Changes in carbohydrate levels and their metabolic enzymes in leaves, phloem sap and mesocarp during cucumber (Cucumis sativus L.) fruit development. Sci. Hort. 121 (2), 131–137 (2009).
Ren, B. et al. Effects of spraying exogenous hormone 6-benzyladenine (6-BA) after waterlogging on grain yield and growth of summer maize. Field Crop Res. 188, 96–104 (2016).
Zhang, W. et al. Plant growth regulator and its interactions with environment and genotype affect maize optimal plant density and yield. Eur. J. Agron. 91, 34–43 (2017).
Kaya, C., Tuna, A. L. & Okant, A. M. Effect of foliar applied kinetin and indole acetic acid on maize plants grown under saline conditions. Turk. J. Agric. For. 34, 529–538 (2010).
Kamran, M. et al. Paclobutrazol application favors yield improvement of maize under semiarid regions by delaying leaf senescence and regulating photosynthetic capacity and antioxidant system during grain-filling stage. Agron. (Basel). 10, 187 (2020).
Gong, L. et al. Improving maize grain yield by formulating plant growth regulator strategies in North China. J. Integr. Agr. 20, 622–632 (2021).
Wang, X. X. et al. The potential of EDAH in promoting kernel formation and grain yield in summer maize. Field Crops Res. 319, 109655 (2024).
Huo, D., Ning, Q., Shen, X., Liu, L. & Zhang, Z. QTL mapping of kernel number-related traits and validation of one major QTL for ear length in maize. Plos One. 11, e155506 (2016).
Ning, Q. et al. An ethylene biosynthesis enzyme controls quantitative variation in maize ear length and kernel yield. Nat. Commun. 12, 5832 (2021).
Ayoub, M., Mobasser, H. R. & Ganjali, H. R. Effect of times and foliar concentration of iron on grain yield, diameter of cob, plant height and dry weight of corn. Int. J. Biosci. 5 (2), 139–143 (2014).
Avat, S. & Yahya, E. Plant growth regulator (ethephon) alters maize (Zea mays L.) growth, water use and grain yield under water stress. J. Agron. 7 (1), 41–48 (2008).
Ren, B., Zhang, J., Dong, S., Liu, P. & Zhao, B. Exogenous 6-benzyladenine improves antioxidative system and carbon metabolism of summer maize waterlogged in the field. J. Agron. Crop Sci. 204, 175–184 (2018).
Khosravi, G. The Effect of Hormones and Chemical Growth Regulators on Ear Development and Grain Yield of Nonprolific Corn (Zea mays L) (Iowa State University, 1980).
Liu, Y. et al. The additive function of YIGE2 and YIGE1 in regulating maize ear length. Plant J. 119 (3), 1327–1335 (2024).
Zuo, Q. et al. Effects of uniconazole rate on agronomic traits and physiological indexes of rapeseed blanket seedling. Oil Crop Sci. 5, 198–204 (2020).
Cosgrove, D. J. Loosening of plant cell walls by expansins. Nature (London) 407, 321–326 (2000).
Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant. Biol. 59, 225–251 (2008).
Zhou, J. et al. CYP71D8L is a key regulator involved in growth and stress responses by mediating gibberellin homeostasis in rice. J. Exp. Bot. 71, 1160–1170 (2020).
Irshad, A. et al. Effects of plant growth regulators on seed filling, endogenous hormone contents and maize production in semiarid regions. J. Plant Growth Regul. 38, 1467–1480 (2018).
Xu, L. et al. 5-Aminolevulinic acid regulates the grain morphology, carbohydrate and hormone contents, increasing grain yield of maize in field land. Plant. Growth Regul. 102 (3), 633–646 (2023).
Li, Y. et al. Exogenous SA or 6-BA maintains photosynthetic activity in maize leaves under high temperature stress. Crop J. 11, 605–617 (2023).
Crizel, R. L. et al. Abscisic acid and stress induced by salt: Effect on the phenylpropanoid, L-ascorbic acid and abscisic acid metabolism of strawberry fruits. Plant. Physiol. Bioch. 152, 211–220 (2020).
Koentjoro, Y., Sukendah, Purwanto, E. & Purnomo, D. The role of silicon on content of proline, protein and abscisic acid on soybean under drought stress. IOP Conf. Ser. Earth Environ. Sci. 637, 12086 (2021).
Zeevaart, J. A. & Creelman, R. A. Metabolism and physiology of abscisic acid. Annu. Rev. Plant Biol. 39 (1), 439–473 (1998).
Han, Y. et al. Genome-wide transcriptome profiling reveals the mechanism of the effects of uniconazole on root development in Glycine max. J. Plant. Biol. 60, 387–403 (2017).
Dello, I. R. et al. A genetic framework for the control of cell division and differentiation in the root meristem. Science 322, 1380–1384 (2008).
Zhong, Y. et al. ZmCCD8 regulates sugar and amino acid accumulation in maize kernels via strigolactone signalling. Plant Biotechnol. J. 23 (2), 492–508 (2025).
Samanta, A. K. et al. Production and in vitro evaluation of xylooligosaccharides generated from corn cobs. Food Bioprod. Process. 90, 466–474 (2012).
Nurika, I., Rahmadhanti, A. & Suhartini, S. Bioconversion of lignin and methane production from Corn cobs (Zea mays L.) treated by lignin-degrading bacteria. IOP Conf. Ser. Earth Environ. Sci. 924, 12072 (2021).
Zhu, M. et al. Uniconazole enhances lodging resistance by increasing structural carbohydrate and sclerenchyma cell wall thickness of Japonica rice (Oryza sativa L.) under shading stress. Environ. Exp. Bot. 206, 1–14 (2023).
Behr, M., Lutts, S., Hausman, J. F. & Guerriero, G. Jasmonic acid to boost secondary growth in hemp hypocotyl. Planta 248, 1029–1036 (2018).
Polko, J. K. & Kieber, J. J. The regulation of cellulose biosynthesis in plants. Plant. Cell. 31, 282–296 (2019).
Ball, S. G. & Morell, M. K. From bacterial glycogen to starch: Understanding the biogenesis of the plant starch granule. Annu. Rev. Plant. Biol. 54, 207–233 (2003).
Maarel, J. E., Veen, B. V., Uitdehaag, J. C., Leemhuis, H. & Dijkhuizen, L. Properties and applications of starch-converting enzymes of the alpha-amylase family. J. Biotechnol. 94 (2), 137–155 (2002).
Jiang, Y. et al. Physiological and transcriptome analyses for assessing the effects of exogenous uniconazole on drought tolerance in hemp (Cannabis sativa L). Sci. Rep. 11, 14415–14476 (2021).
Hara, M. et al. Effects of 5-aminolevulinic acid on growth and amylase activity in the radish taproot. Plant. Growth Regul. 64, 287–291 (2011).
Liu, C. et al. Uniconazole and diethyl aminoethyl hexanoate increase soybean pod setting and yield by regulating sucrose and starch content. J. Sci. Food Agric. 99, 748–758 (2019).
Acknowledgements
The authors wish to thank the National Coarse Cereals Engineering Research Center of Heilongjiang Bayi Agricultural University for providing the research platform and technical support. We would like to express our gratitude to the anonymous reviewers for their valuable comments and suggestions, which greatly improved the quality of this manuscript.
Funding
This research was funded by Fujian Provincial Education Department Key Research Projects for Young and Mid-career Teachers (grant number JZ240089), Introduction of Talents and Scientific Research Project of Fujian Vocational College of Agriculture (grant number ZZXYJ11525002) and University-Enterprise Collaborative Development Project (grant number 1048250100301).
Author information
Authors and Affiliations
Contributions
Yue Zhao: Writing – review & editing, Writing – original draft, Formal analysis, Data curation, Conceptualization. Dianfeng Zheng: Writing – review & editing, Writing – original draft, Formal analysis, Data curation, Conceptualization. Lei Xu: Writing – review & editing, Writing – original draft, Formal analysis, Data curation, Conceptualization, Funding acquisition. Huiquan Wang: Investigation, Formal analysis, Data curation. Xueying Huang: Formal analysis, Data curation. Qinghua Ye: Formal analysis, Data curation. Tingping Chen: Formal analysis, Data curation. JEI Vincent: Writing – review & editing, Formal analysis, Data curation.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Xu, L., Wang, H., Huang, X. et al. Regulating female ear morphogenesis in maize by uniconazole and 5-aminolevulinic acid for yield improvement. Sci Rep 16, 13848 (2026). https://doi.org/10.1038/s41598-026-41685-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-41685-y
