
Abstract
Epidermal glands are well-known in adult beetles, but their presence and function are less studied in larvae. This study reveals, for the first time for the family Leiodidae, the morphology and ultrastructure of epidermal, paired glands of abdominal segments present in larvae. They are documented in all larval stages of the scavenger beetle Sciodrepoides watsoni watsoni (using a multimodal microscopic approach: light and scanning electron, and transmission electron microscopy). Overview of these structures is also documented for other representatives of Leiodidae and Agyrtidae, basal lineages of the megadiverse Staphylinoidea. Structures differ in their position relative to a transverse cuticle fold, in the size and shape of the perforate cuticular area, and in the position and form of their openings. Their ultrastructure in Sciodrepoides watsoni watsoni strongly indicates a secretory function. All abdominal segmental glands are formed by a complex of (1) regular class 1 gland cells type 1, (2) modified class 1 cells type 2, with an apical reservoir-like invagination, and (3) class 3 gland units, consisting of one basal secretory cell plus a canal cell. All these glands have their own openings to the cuticle surface. Their secretions do not mix before their transport to the surface.
Similar content being viewed by others
Introduction
Epidermal, exocrine glands are widespread among insects, and their secretions play various important functions, as reviewed by Betz1 and later by Foster and Casas2. Their classification according to their ultrastructure has been established for all insects3,4, and later updated1,5,6.
Epidermal glands are known in many adult beetles, such as Carabidae7, Cicindelidae8, Ciidae9, Chrysomelidae10, Coccinellidae11, Meloidae12, Staphylinidae13, and Tenebrionidae14. Significantly less is known about their presence in their immature stages15. Lawrence et al.16 suggested that exocrine abdominal glands are common in larvae but very often overlooked because of their small size. These structures were often described under various morpho-functional terms (like pores, campaniform sensilla, porous plates, openings, without histological studies), which complicated the situation17. However, exocrine abdominal glands are well-known in larvae of some families of beetles, e.g. Carabidae18, Chrysomelidae19, Coccinellidae20, Tenebrionidae21, and various Staphylinoidea.
The superfamily Staphylinoidea is one of the largest groups of Coleoptera, with more than 76,000 described species22. Currently, it is classified into seven families, with the following hypothesised relationships: Staphylinidae [incl. Silphinae] + ((Leiodidae + Agyrtidae) + (Colonidae + (Jacobsoniidae + (Ptiliidae + Hydraenidae))))23,24.
Larval glands, placed dorsally on the body segments, were mentioned in descriptions of larvae of two staphylinoid groups, the aquatic Hydraenidae25 and in primitive carrion beetles, Agyrtidae26. They were also briefly described in several lineages of megadiverse short-winged rove beetles, Staphylinidae17,27,28,29. Up to date, only a single currently published study provided ultrastructural and functional documentation30. In general, overall knowledge of staphylinids remains poor, given their enormous species diversity, with over 68,000 described species22. To date, there are no data about the presence of such glands in larval stages of carrion beetles (Silphinae), feather-winged beetles (Ptiliidae) or Jacobsoniidae, which was only recently transferred into Staphylinoidea23.
The family Agyrtidae23,24,31, including only 73 extant species22, shows a very high morphological and biological diversity as well as a relict distribution in the mountainous areas of the world31,32. There is only one detailed description with illustrations of the larval stage available for one European species33. The revision of this family32, although very valuable and including the characteristics and even a descriptive key to the larvae, unfortunately lacks any photographs or illustrations of the immature stages.
Its sister family Leiodidae belongs to one of the most interesting groups within the superfamily with a high ecological and biogeographical diversity31. However, for many subgroups, their biology is often virtually unknown31. Therefore, this family is particularly interesting in a phylogenetic and evolutionary context, as well as for searching for new morphological features which may bring new phylogenetic signals34.
Our knowledge of the glands on the segments in leiodid larvae is limited to a few illustrations, and descriptions of the presence of some pores which may be glandular35,36 and their photographs using light phase contrast and scanning electron microscopes37,38,39,40,41. The only scanning electron microscopic documentation of such glandular structures in larvae of Dissochaetus Reitter was provided by Gnaspini42. However, their functions have never been studied, although some sticky material on the surface of the cuticle was detected and documented (Figs. 69, 70 and 71 in37, Figs. 26 and 27 in26 and their glandular function was discussed37. Some character states of these structures were used in phylogenetic analyses of Leiodidae: Camiarinae26.
Here, we provide the first documentation of the ultrastructure and the formal reconstruction of abdominal glands in the family Leiodidae, based on the species Sciodrepoides watsoni watsoni (Spence). We also provide an overview of the morphological diversity of glandular abdominal structures in several Leiodidae and Agyrtidae, based on available larval material and a review of published larval descriptions and illustrations.
Results
Detailed description of abdominal glandular structures in larvae of Sciodrepoides watsoni watsoni.
External morphology (Figs. 1 and 2)
A paired complex of morphological structures is distributed on the latero-apical area of the dorsal side of each abdominal segment (I–X) of the larval body in all three instars, above the transverse cuticular fold (carina) (Fig. 1A–D).
Each complex consists of four cuticular openings and one large oval perforated structure of ca. 50–58 μm diameter and 20 μm high (n = 2) (Figs. 1C and D and 2A). One of openings, rosette-like, is located in the centre of the complex, and surrounded by cuticular folds (Figs. 1D and 2A, B and D; ro, cfa). Three cup-like openings lay close and slightly anterior the perforated oval-shaped region of the cuticle with ca. 300 pores. The fourth, additional, cup-like opening is lateral to the whole complex (Fig. 1D). Three types of cuticular openings are distinguished: a cup-like (Fig. 2A, 5 μm diameter; co), a rosette-like (Fig. 2A, B, D, ca. 5 μm diameter; ro), and fine pores (Fig. 2C). The fine pores are numerous (ca. 300) but present only on an oval-shaped region of the cuticle (cuticular perforated / pore plate), which is slightly raised (Fig. 2A; cpa). The rosette-like opening is surrounded by radially arranged folds of cuticle (Fig. 2A, B, D; ro).

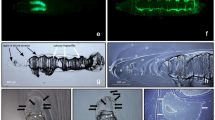
Habitus (dorso-lateral or dorsal aspect) of the third instar larva of Sciodrepoides watsoni watsoni (Spence) showing the locations of the paired glands, arrows (A, B). External morphology of the epidermal segmental glands in S. watsoni watsoni, tergite IV(C). Details of glandular complex, tergite IV, left side (D). Light microscopy. ca, carina; cpa, cuticular perforated area; co, cup-like opening; cs, campaniform sensillum; ro, rosette-like opening.

Details of external morphology of larval epidermal segmental glandular complex in Sciodrepoides watsoni watsoni (Spence). Glandular complex (A). Details of openings of cells class 1 and cell class 3 with cuticular fold area (B). Details of cuticular perforated area – openings of cells class 1 (C). Openings of lamellae cells and cell class 3 with secretion (D). Scanning Electron Microscopy. ca, carina; cfa, cuticular fold area; cpa, cuticular perforated area; co, cup-like opening; ro, rosette-like opening.
Morphology and ultrastructure
A series of longitudinal and transverse semithin sections through the larval abdomen have been used to demonstrate a complex of three types of modified glandular epidermal cells between the cuticle and muscles (Fig. 3A, supplementary Figs. S1, S2). The composition of each glandular complex is as follows: multiple cells of the C1 type 1, three cells of the C1 type 2, and one class C3 gland unit, composed of terminal cell and canal cell.
The first type of gland cells comprises C1 cells (named C1 type 1 cells). These cells are narrow and elongated, located in direct contact with the terminal cell of C3 gland unit (Fig. 3B, tc). Each cell forms a protrusion, that passes through the cuticle towards to the base of the perforated plate (Fig. 3B, asterisk, C). Apical part of these protrusions forms microvilli, which are located under the oval-shaped part of cuticle perforated by about 300 canals with a diameter of approximately 150 nm, and they are faced towards to the base of the perforated plate (Fig. 3C). The cytoplasm of C1 type 1 cells has a high electron density and is filled with a high number of microtubules, and vesicles (Fig. 3C, D).
The second type of gland cells also comprises C1 cells (in this case modified, named C1 type 2 cells) forming a pouch-like structure (Fig. 3A). These cells apically possess a deeply invaginated reservoir-like extracellular space bordered by microvilli. The highly invaginated apical membrane forms labyrinth of foldings (Fig. 4B; fl), which are modified into numerous microvilli that face toward a circular cup-like opening (diameter 5 μm). The cuticle surrounding the opening is deeply folded (Figs. 4B and 6) and forms irregular sieve-like canals by whom the secretion is transported out of the body. The basal part of the cuticle is invaginated, forming the shallow cupule faced towards microvilli (Figs. 4B and 6 and c). Secretion in extracellular space forms strands near the microvilli (Fig. 4B; arrows), and dense homogeneous mass near the cupule (Fig. 4B; sc). The external opening of the gland is plugged by electron dense secretion (Fig. 4D; s). C1 type 2 cells are characterised by a large, approximately 10 μm in diameter, roughly spherical nucleus, which is flattened in the basal part (Fig. 4A; n). A prominent nucleolus is present in the central part of the nucleus. Cytoplasm of the C1 type 2 cells have a high number of mitochondria, Golgi complexes and rough endoplasmic reticulum (Fig. 4A, C; m, Gc, rer).
The third type of gland can be classified as compound class 3 gland, consisting of the terminal secretory cell that distally connects to the canal cell (Figs. 5 and 6, S1). In the cytoplasm of the terminal cell, there is a very large nucleus of about 22 μm in diameter (Fig. 5A; n). Cytoplasm is densely filled with cisternae of rough endoplasmic reticulum, mitochondria, Golgi complexes and numerous secretory vesicles (Fig. 5B). Cisternae of the endoplasmic reticulum are very numerous and evenly distributed in the cytoplasm (Fig. 5A; rer), and the lumen of the cisternae is filled with medium electron density material (Fig. 5B). The terminal cell is characterized by an internalized extracellular space that is lined with numerous long microvilli (Fig. 5B; mv), which disappeared just near the canal cell (Fig. 5C). In the region of the end apparatus of the terminal cell, numerous highly electron-dense secretory granules, with an average diameter of 5 nm, are gathered near the microvilli, which surrounded the receiving canal (Fig. 5B). Near the cuticle the terminal cell is connected with the canal cell, which is filled with numerous microtubules (Fig. 5C; mt). The canal cell forms chitinous duct that ends in a rosette-shaped opening (Fig. 5D).

Histology and ultrastructure of the C1t1 cells. Structure and ultrastructure of the larval epidermal segmental glandular complex in Sciodrepoides watsoni watsoni (Spence). Cross section of larval abdomen with glandular complex (A). Longitudinal section of C1t1 gland cell (B). Section of the apical part of the C1t1 gland cell (C). Section of the C1t1 gland cells near the bottom part of the cuticle (D). Semi-thin section stained with methylene blue (A) and Transmission Electron micrographs (B–D). Asterisk, apical part of C1t1 gland cell; C1t1, C1t1 gland cell; C1t1p, processus of the C1t1 gland cell; C1t2, C1t2 gland cell; C3d, duct of the C3 canal cell; cpa, cuticular perforated area; cpac, canal of the cuticular perforated area; cu cuticle; ep, epidermal cells; m, mitochondrion; mt, microtubules; ms, muscle; mv, microvilli; tc, terminal cell; v, vesicle.

Ultrastructure of the C1t2 gland cells. Longitudinal section of basal part of the C1t2 gland cell (A). Longitudinal section of central and apical part of the C1t2 gland cell (B). Section of the central part of the C1t2 gland cell (C). Apical part of the cell with lamellae and gland opening (D). Transmission Electron Microscopy. Arrow, strands of secretion; bm, basement membrane; c, cupule; cu, cuticle; ecs, extracellular space; fl, folds of the central part of the C1t2 gland cell; Gc, Golgi complexes; sc, secretion gathered near cupule; m, mitochondria; mv, microvilli of the apical part C1t2 gland cell; n, nucleus; no, nucleolus.

Ultrastructure of aspects the C3 gland units. Cytoplasm of the terminal cell of the C3 gland unit (A). Microvilli (embracing the extracellular reservoir) of the terminal cell (B). Apical part of the terminal cell and the canal cell with the lumen of the duct (C). Canal cell with the chitinized duct (D). Transmission Electron Microscopy. cc, canal cell; ch, chitinized part of the duct; cu, cuticle; d, lumen of the glandular duct; Gc, Golgi complexes; m, mitochondrion; mt, microtubule; mv, microvilli; n, nucleus; rer, rough endoplasmic reticulum; s, secreted material in the lumen of the duct; sg, secretory granules; tc, terminal cell.
A generalised reconstruction of these structures is presented in Fig. 6 whereas differences between three types of glandular cells in the abdominal glandular complex are summarized in the Table 1.

Schematic reconstruction of the complex of the three types of glands found in abdominal segments of larvae of Sciodrepoides watsoni watsoni. c, cupule; co, cup-like gland opening; cpa, cuticular perforated area; cu, cuticle; d, duct; ep, epidermis; fl, filaments; l; ms, muscle fibres; mv, microvilli; n, nucleus; ro, rosette-like gland opening; s, external homogenous secretion; sc, secretion accumulated near cupule in extracellular space; C1t1, class 1 cell type 1; C1t1p, processus of the C1 cell; C1t2, class 1 cell type 2; C3cc, class3 canal cell; C3tc, class 3 terminal cell.
Overview of diversity of the external morphology of abdominal glandular structures in larvae of Leiodidae and Agyrtidae
Glandular structures on abdominal segments seem to be widespread in Leiodidae and Agyrtidae. Available information about the external morphology of abdominal glandular structures in larvae of 77 leiodid and four agyrtid species is summarized in Table S1 (the Supplementary File) and Figs. 7 - 8. New data, as well as data from a previous study26 and other papers (citations in the Supplementary File) are combined.We found a large morphological diversity consisting of different characters of perforated cuticular area and gland openings.The position of the perforated cuticular area (cpa) can be located either anterior to the cuticular transverse fold – carina (Fig. 7C, D, G, H) or posterior to it (Fig. 7A, B, E, F, see also Zearagytodes sp. and Dictydiella leleupi Jeannel in Figs. 116 and 122 in38). The shape of the perforated area of the cuticle (cpa) can be oval (Fig. 7A, B, E, e.g. in Necrophilus spp. in Agyrtidae, Fig. 7B; Neopelatops spp., Fig. 7G; Eublackburniella Jeannel; Fig. 17 in26; Leiodes cinnamomea (Panzer), Fig. 14D and E, 15 A, 16 C in41), bean-shaped (Fig. 7E, F and D. leleupi, Fig. 122 in38) or elongated like in Prionochaeta opaca (Say) (Figs. 2B and 7H in40) or in Dissochaetus vanini Gnaspini (Fig. 20 in42).
The position of the opening in the perforated area of the cuticle can be central (Agyrtidae, Fig. 7B, G; o), invaginated below this area (as in Eublackburniella, Fig. 17 in26) or invaginated above (Fig. 7F, H) as in D. leleupi (Fig. 122 in38). The presence of cuticular folds of the cuticle near the openings – they are well developed in species of Myrmicholeva Lea (Fig. 7D and C) and Neopelatops Jeannel (Fig. 27 in26) or slightly less developed (Figs. 23 and 24 in42).
Also, the number of openings of the gland cells can vary among leiodids, from one (e.g. Ragytodina tuberculosa Szymczakowski) up to even 20 pairs (e.g. Catops fuscus fuscus Panzer) (Table S1, Supplementary File).
The type of openings of glands can be rosette-like (Fig. 8A-D, ro) or cup-like (Fig. 8A, co) or another type (8D, o). One type may co-occur on the abdominal segment with others, as we have shown for S. watsoni watsoni (Figs. 2A and 6). Most probably, accordingly to our results, they belong to different classes of glands.
The inner side of the perforated area of cuticle has a different, rough texture than the rest of the cuticle, which is entirely smooth (Fig. 8E, F). A short cuticular duct is visible (Fig. 8E, F) on the rough part of the cuticle.

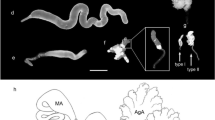
Diversity of external morphology of larval epidermal abdominal segmental glands in Necrophilus hydrophiloides Guérin-Méneville, tergites VI and VII (A, B); Myrmicholeva sp. 1, tergites II and III (C) and tergite IX (D); Cholevomorpha sp., tergites VII and VIII (E) and tergite IX (F); Neopelatops sp. 1, tergites IV and V (G); Prionochaeta opaca (Say), tergites VI and VII (H). Photographs A, C, E, G, H shows gland openings (black arrow). Photographs B, D, F shows details of external gland morphology. Light microscopy. ca, carina; cfa, cuticular fold area; co, cup-like opening; cpa, cuticular perforated area; o, gland opening.

Details of the external morphology of the larval epidermal abdominal glandular complex in other Leiodidae and Agyrtidae. Dictydiella sp. (Leiodidae, Camiarinae) with numerous cup-like openings (co) anterior to carina (ca.), tergite IV (A); details of rosette-like opening and cuticular fold and perforated areas, tergite IV (B); Myrmicholeva sp. 1 (Leiodidae, Camiarinae), tergite V (C); Myrmicholeva sp. 2 (Leiodidae, Camiarinae), tergite VII (D); Necrophilus pettiti Horn (Agyrtidae) (E); Leiodes cinnamomea (Panzer) (Leiodidae, Leiodinae) (F). Scanning Electron Microscopy. ca, carina; cd, cuticular duct; cfa, cuticular fold area; co, cup-like opening; cpa, cuticular perforated area; o, opening; ro, rosette-like opening.
Discussion
The ultrastructure of larval thoracic and abdominal morphology confirms our hypothesis about the glandular character of these structures in the family Leiodidae. The gland complex of each abdominal segment comprises:
-
Several class 1 cells type 1, secreting directly to the exterior through numerous (ca. 300) fine pores 150 nm diameter,
-
Three class 1 type 2 gland cells with infoldings of the apical cell membrane forming an extracellular space and secreting by cup-like openings,
-
One large class 3 cell unit (consisting of the terminal secretory cell and the canal cell), with the secretory duct, releasing the secretion by the rosette-like opening.
Via our light and scanning electron microscopic studies, a diversity of glandular structures could be revealed with respect to:
-
Their position according to the cuticular transverse fold (carina),
-
The shape and size of the perforated area of the cuticle,
-
The presence of the cuticular folds surrounding the openings,
-
The position of the gland cell class 3 opening according to the perforated area of cuticle,
-
The number of cup-like glandular openings.
Comparison of ultrastructure of abdominal glands in larvae of Sciodrepoides watsoni watsoni with other insect glands
The ultrastructure of larval abdominal segmental glands of Stenus Latreille30 shows differences between the glands of segments I–VII and the glands of the segments VIII and IX but all of them are compound glandular units consisting of a syncytium of secretory cells with a common reservoir and with a single duct and a single opening. On the contrary, all abdominal segmental glands of larvae of Sciodrepoides watsoni watsoni are a complex of two types of class 1 gland cells and a class 3 gland unit. All these secretory cells have their own openings to the surface of the cuticle. Their secretions do not mix before their transport to the surface. The ultrastructure of the C1 gland cells and the C3 gland unit are similar to those described by Quennedey5 and in other studies [e.g.30,47,58]. The presence of the rough endoplasmic reticulum (rer) and free ribosomes suggest the production of proteins.
The function of the second type of C1 gland cells remains to be elucidated. The strongly infolded apical membrane of these cells change their organizations and structures from irregular labyrinth near the nucleus to regular apically located microvilli; similar cells have not previously been described in beetle glands. However, the ultrastructure of intramandibular glands of ants shows that the microvilli of the C1 cells apparently originated from irregular folds[66, Fig. 4C].
It is noteworthy that our study reveals that each class 1 gland cell type 2 possesses an epicuticular modification, in the border of the cuticle and the gland cell, which forms a bowl-like barrier, near which the secretion accumulates and is transformed from linear strands to a homogenous mass. Quennedey et al.47 described a similar modification of class 1 gland cells for paired prototergal glands in the adult of the staphylinid Philonthus varians (Paykull, 1789) and called it as ‘cupule’.
The presence of a large number of mitochondria and relatively few, compared to C1 type 2 and C3 terminal cells, rough endoplasmic reticulum (rer) and free ribosomes suggests that these cells may be involved in the production of substances for organismal defence. Giglio et al.15 reported secretory cells containing numerous, large, elongated mitochondria arranged adjacent to electron-lucent vesicles, in the defensive abdominal glands of Carabus lefebvrei Dejean pupae. Similarly, Sánchez et al.43 described pheromone-producing glands in Rhynchophorus palmarum (Linnaeus) as featuring numerous mitochondria. However, a pheromone-producing function in S. watsoni watsoni is unlikely due to the larval stage of the specimens studied.
Morphological diversity and patterns of exocrine epidermal glands among Leiodidae and Agyrtidae
Our comparison of the external morphology of the glandular abdominal structures in larvae of Leiodidae and Agyrtidae indicates certain patterns. In all the studied larvae of Agyrtidae the glandular structures open posteriorly to the carina, whereas in Leiodidae their position differs. Openings posterior to the carina are present in Oritocatopini (Cholevinae), Agyrtodini (Camiarinae) and Eublackburniella (Camiarinae: Neopelatopini), whereas openings anterior to the carina are presented in most of the investigated Cholevinae (Cholevini, Anemadini, Leptodirini) and other Neopelatopini (Camiarinae). Some leiodid taxa lack a carina, so that their glandular structures are located in the apico-medial part of the segments (some Camiarinae, Cholevinae: Leptodirini, and Catopocerinae).
Different positions of glandular openings either anteriorly or posteriorly to tergal carina can support the evidence of31 that Camiarinae: Neopelatopini in the present concept may be paraphyletic.
Potential function of exocrine abdominal glands in larvae of Leiodidae
Beyond their role in cuticle formation, the role of exocrine epidermal glands and their secretions in insects can be generally divided into three categories, i.e. life history and physiology, communication and sociality, and protection from predators2. Our study has not addressed the chemistry of these glands, and it is also not yet possible to compare their function and chemistry with any other leiodid taxa, as none have yet been studied.However, a recent study of Staphylinidae: Steninae larval abdominal segmental glands30 provides good material for comparison. Zippel et al.30 described the retreat-spinning behaviour of the larvae of all three instars (for retreat building behaviours in other rove beetles see44,45), employing the abdominal glands of segments VIII and IX and discuss the adhesive role of glands on the remaining abdominal segments I–VII used in prey-capture. Also, the larvae of leiodid Agathidiini are known to form cocoons for pupation, but they probably do so in the way observed in Leiodes cinnamomea, where larvae chewed truffle masses to create cocoons (Figs. 1E and F and 2D in41).
In the here investigated larvae of S. watsoni, we could not observe any prey-capture or silk production behaviour. However, their body is certainly covered by substances, which are difficult for an investigator to clean off in any way (AK, JR, personal observation). Both larvae and adults live in wet habitats, feeding on rotten animal matter, with abundant pathogenic bacteria and fungi39,46. The third category of function (protection2) also includes protection against predators, parasitoids, and pathogens.
Quennedey47 provided a hypothesis about the prophylactic function of the prototergal glands of adult Philonthus rove beetles, which feed in decaying substrates, including dung and carrion, where abundant microbial activity by bacteria and fungi occurs. An antimicrobial function was recently documented for such glands in Carabidae48,49, Hydraephaga50, Staphylinidae51 and Staphylinidae: Silphinae, whose larvae use also their oral and abdominal secretions to contribute to resource defence and social immunity52.
On the other hand, fungi and slime moulds (Myxomycetes) show broad antimicrobial activity53,54. Larvae of Agathidiini, which are obligate myxomycetophages55,56, lack a perforated cuticular area (Figs. 18 and 19 in26, Table S1 in the Supplementary File). This may reflect their adaptation to specific habitats, where antimicrobial protection is provided by a food source. Currently, the knowledge about the prevalence and diversity of antimicrobial peptides (AMPs) produced by insect glands is still expanding57.
Many variable factors influence the interaction of glands in a single complex, and the chemical and physical properties of secretions, and determine their function2. For example, two-phase secretions can be formed – a lipoid and a proteinaceous fraction as in adults of Stenus rove beetles58. Moreover, the chemical composition of epidermal glandular secretions may vary even within one family59. There are some interesting hypothetical explanations for glandular complex interactions in adult free-living staphylinids60. First, secretion from additional individual glandular cells may improve the effectiveness of the main gland by sealing and enlarging the secretory area. A second hypothesis is that the secretory products of individual additional glandular cells enhance the effectiveness of the entire gland or activate other secretions. A third hypothesis is that individual glandular cells may protect the body surface around the glandular openings from their aggressive secretions.
The glandular complex of larvae of Sciodrepoides watsoni watsoni is formed from two classes (C1 and C3) of secretory cells. The class 1 gland cells type 2 possess an extracellular space, which allows storage of the secretion. Moreover, the other modification of these cells, the cupule, allow the secretion to transform before reaching the surface of the cuticle. Epidermal class 1 gland cells, which are important part of the glandular complex of S. watsoni watsoni, are the prevailing type of adhesive glandular cells in insects1. Epidermal class 3 gland units always co-occur with at least one canal cell and a duct, and can also perform adhesive functions1. Adhesive glands can play many roles: in locomotion, in defence, in water balance, capturing prey or building cocoons and others1– belonging to the first and the third category designated by Foster and Casas2, although they have never been described in Leiodidae61.
Our preliminary study of the ultrastructure of glands of S. watsoni watsoni, and an overview of their presence and diversity in the sister families Leiodidae and Agyrtidae indicate the need for further studies concerning their ultrastructure and function. These structures were visible under the optical light microscope in the larvae of many investigated species. However, this only provides indirect evidence. Only detailed histological and TEM studies could answer the question of what exactly these structures are. This is not possible unless the larvae are fixed directly in glutaraldehyde in proper conditions (see Material and methods). Conventionally stored larvae (in alcohol or Pampel’s fluid) are not suitable for such studies.
Although the thoracic exocrine glands were not subject to histological examination in our publication, we would like to note that they are present in S. watsoni watsoni, as well as in many of the species of Leiodidae. This could provide data for clarifying phylogenetic relationships and evolutionary scenarios within this basal lineage of the superfamily Staphylinoidea. Previous studies, in which character states of epidermal glands were used in phylogenetic analyses, both in adults8,13 and larvae62, encourage further investigation.
Materials and methods
This study is based on larvae of all stages of Sciodrepoides watsoni watsoni. Adults were collected in the Czech Republic (detail data below), bred in the laboratory on Petri dishes with wet soil and provided with chicken meat ad libitum at a constant temperature of 10 °C. Larvae were frequently collected from Petri dishes and preserved in 2.5% glutaraldehyde for TEM analysis or preserved in 75% ethanol for light microscopy slides and SEM analysis, and kept in 4 °C. Larval instars were identified based on head width of larvae according to67 and setation pattern39.
Material examined. Czech Republic, Bohemia bor., Lomnice nad Popelkou – Ploužnice, 540 m, 50°30.7’N 015°23.0’E, 4.VI. 2022, baited pitfall trap (fish meat), ecotone of old orchard and margin of deciduous forest, temp. 18–22 °C, Jan Růžička leg.; Czech Republic, Bohemia bor., Litoměřice env., Mentaurov, 420–440 m, 50.5711°N 014.1216–1225°E, 29.V.–1.VI. 2023, baited pitfall traps (chicken meat), margin of deciduous forest, temp. 20–22 °C, Pavel Jakubec leg.
Collecting data and deposition of additional larvae of Leiodidae and Agyrtidae used for comparative purposes (Table S1) are listed in the Supplementary File.
Light microscopy
Larvae were briefly macerated in warm 10% KOH, washed in distilled water, and mounted in glycerol-gelatin on semi-permanent slide mounts with cover slips. Photographs of habitus and details of larvae were made using Keyence® VHX6000 or VHX7000 digital microscope (University of Wrocław, Poland and University of Life Science Prague, Czech Republic). Additionally, some of the images were documented at magnifications up to 1,000× using Nikon® Eclipse 80i Phase Contrast microscope by a Nikon Coolpix® 4500 digital camera.
Scanning electron microscopy
Larvae were rinsed in distilled water and dehydrated in graded series of ethanol baths. To achieve the inner side of the cuticle, larvae of Leiodes cinnamomea and Zearagytodes sp. were cut longitudinally, treated in NaOH solution, and the fat body and gastrointestinal tract were removed. The integument was rinsed in water once again and dehydrated in graded series of ethanol baths. Then, the larvae and the prepared cuticle were dried using Hexamethyldisilazane, mounted on an SEM stub with carbon tab, coated with gold using Bal-Tec SCD 050 and examined at magnifications up to 20,000× with a Scanning Electron Microscope (JEOL® 6380 LV, Akishima, Tokyo, Japan) and magnifications up to 33,000× under HR SEM JEOL® IT800, Akishima, Tokyo, Japan (Viničná Microscopy Core Facility, VMCF of the Faculty of Science, Charles University, Prague, Czech Republic).
Histological analyses and transmission electron microscopy of abdominal segments
Larvae of S. watsoni watsoni were fixed in 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.4) for a few days at 4 °C. After fixation, the specimens were repeatedly rinsed with phosphate buffer. For better tissue preservation and contrast, the material was subsequently postfixed in a mixture containing 1% osmium tetroxide and 0.8% potassium ferrocyanide (according to63). After dehydration in an acetone series, larvae were embedded in Epon 812 (Serva, Heidelberg, Germany).
Next, the material was cut using an RMC PowerTome XL microtome (RMC Boeckeler) (Laboratory of Microscopic Techniques, University of Wrocław) for semi-thin sections. Semi-thin Sect. (0.6 μm) were stained with 1% methylene blue and examined with an Olympus® BHS light microscope equipped with ColorView IIIu CCD colour camera and Olympus® CellA imaging software (Olympus, Germany, Hamburg, RRID: SCR_014342). Whole mount observations were conducted using an Olympus® SZ61 microscope equipped with SC30 camera and Stream Start 1.6.1 software. Using this staining method, methylene blue stains nonspecifically lipid droplets green. Selected fragments of abdominal segments of larvae were cut into ultra-thin Sect. (70 nm) for examining their ultrastructure (RMC® PowerTome XL, Laboratory of Microscopic Techniques, University of Wrocław). Ultrathin sections were contrasted with uranyl acetate and lead citrate64 and examined under the transmission electron microscope Talos L120C (Thermo Fisher Scientific) (Laboratory of Microscopic Techniques, University of Wrocław).
Terminology
The classification of classes of epidermal gland cells are defined according to3,4,5. To distinguish between the two types of cells classified as class C1 according to Quennedey (1998)5, we use the terms C1 type 1 and C1 type 2. The term “rosette-like” structure was used for structures found in Archaeognatha and Thysanura, which are openings of epidermal glands class 3 epidermal gland units65 and in the description of the glandular complex described in the staphylinid Steninae30. Terminology of larval morphology follows16,27. Terms used in26,37,38 follow3,4: “pretergal and tergal glands” – secretory openings of glandular cells, “reservoirs” – perforated cuticular area.
Data availability
Data generated and/or analysed during the current study are available from the corresponding author upon reasonable request. Larvae of S. watsoni watsoni in alcohol are deposited in the collections of Aleksandra Kilian (Department of Biology, Evolution and Conservation of Invertebrates, Faculty of Biological Sciences, University of Wrocław, Poland) and Jan Růžička (Department of Ecology, Faculty of Environmental Sciences, Czech University of Life Sciences Prague, Czech Republic). Histological slides are deposited in the collection of Arnold Garbiec (Department of Animal Developmental Biology, Faculty of Biological Sciences, University of Wrocław).
References
Betz, O. Adhesive exocrine glands in insects: morphology, ultrastructure, and adhesive secretion in Biological adhesive systems. From nature to technical and medical application (eds von Byern, J. & Grunwald, I.) 111–152 (Springer, Berlin) (2010). https://doi.org/10.1007/978-3-7091-0286-2_8
Foster, S. P. & Casas, J. How insect exocrine glands work. Annu. Rev. Entomol. 70, 65–82. https://doi.org/10.1146/annurev-ento-011624-013339 (2025).
Noirot, C. & Quennedey, A. Fine structure of insect epidermal glands. Annu. Rev. Entomol. 19, 61–80. https://doi.org/10.1146/annurev.en.19.010174.000425 (1974).
Noirot, C. & Quennedey, A. Glands, gland cells, glandular units: some comment on terminology and classification. Ann. Soc. Entomol. Fr. Nouv Sér. 27, 123–128 (1991).
Quennedey, A. Insect epidermal gland cells: ultrastructure and morphogenesis in Microscopic anatomy of invertebrates, Volume 11, Insecta (ed Harrison, F. W.) 177–207 (Wiley-Liss, New York) (1998).
Quennedey, A. Perspectives on four decades of transmission-electron microscopy on insect exocrine glands. Le ghiandole esocrine degli insetti. Atti Accad. Naz. Ital. Entomol. 1, 85–116 (2000).
Giglio, A., Brandmayr, P., Talarico, F. & BrandmayT. Z. Current knowledge on exocrine glands in carabid beetles: structure, function and chemical compounds. ZooKeys 100, 193–201. https://doi.org/10.3897/zookeys.100.1527 (2011).
Pearson, D. L. et al. Historical perspective and the interpretation of ecological patterns: defensive compounds of tiger beetles (Coleoptera: Cincindelidae). Am. Nat. 132, 404–416 (1988).
Buder, G., Grossmann, C., Hundsdoerfer, A. & Klass, K. D. A contribution to the phylogeny of the Ciidae and its relationships with other cucujoid and tenebrionoid beetles (Coleoptera: Cucujiformia). Arthropod Syst. Phylog. 66, 165–190 (2008).
Pasteels, J. M., Rowell-Rahier, M., Braekman, J. C., Daloze, D. & Duffey, S. Evolution of exocrine chemical defense in leaf beetles (Coleoptera: Chrysomelidae). Experientia 45, 295–300 (1989).
Barbier, R., Ferran, A., Le Lannic, J. & Allo, M. R. Morphology and ultrastructure of integumentary glands of Semiadalia undecimnotata Schn. (Coleoptera: Coccinellidae). Intern. J. Insect Morph Embryol. 21, 223–234 (1992).
Berrioz-Ortiz, A. The presence of mesothoracic glands in Epicauta segmenta (Coleoptera: Meloidae) and other blister beetles. J. Can. Entomol. Soc. 58, 179–181 (1985).
Johannes, L., Steidle, M. & Dettner, K. Chemistry and morphology of the tergal gland of freeliving adult Aleocharinae (Coleoptera, Staphylinidae) and its phylogenetic significance. Syst. Entomol. 18, 149–168 (1993).
Tschinkel, W. R. A comparative study of the chemical defensive system of tenebrionid beetles. 3. Morphology of the glands. J. Morphol. 145, 355–370 (1975).
Giglio, A. et al. The defensive secretion of Carabus lefebvrei Dejean 1826 pupa (Coleoptera, Carabidae): gland ultrastructure and chemical identification. Microsc Res. Tech. 72, 351–361. https://doi.org/10.1002/jemt.20660 (2009).
Lawrence, J. F. in Order Coleoptera in Immature Insects. Vol. 2, 144–184 (eds Stehr, F. W.) (Kendall/Hunt Pub. Co., 1991).
Solodovnikov, A. & Yu Adult and larval descriptions of a new termitophilous genus of the tribe Staphylinini with two species from South America (Coleoptera: Staphylinidae). Proc. Russ. Entomol. Soc. 77, 274–283 (2006).
Erwin, T. L. A synopsis of the immature stages of Pseudomorphini (Coleoptera: Carabidae) with notes on tribal affinities and behaviour in relation to life with ants. Coleopterist Bull. 35, 53–68 (1981).
Bünninge, M. & Hilker, M. Do glanduliferous larvae of Galerucinae (Coleoptera: Chrysomelidae) possess defensive glands? A scanning electron microscopic study. Zoomorphol 124, 11–119. https://doi.org/10.1007/s00435-005-0116-0 (2005).
Pope, R. D. Wax production by coccinellid larvae (Coleoptera). Syst. Entomol. 4, 171–196 (1979).
Abhitha, P., Vinod, K. V. & Sabu, T. K. Defensive glands in the adult and larval stages of the darkling beetle, Luprops tristis. J. Insect Sci. 10, 7. https://doi.org/10.1673/031.010.0701 (2010).
Newton, A. F. Staphyliniformia world catalog database in Catalogue of Life Checklist (Aug 2022) (eds Bánki, O. et al.) (2025). https://doi.org/10.48580/d4sl-3gk
McKenna, D. D. et al. Phylogeny and evolution of Staphyliniformia and Scarabaeiformia: forest litter as a stepping stone for diversification of nonphytophagous beetles. Syst. Entomol. 40, 35–60. https://doi.org/10.1111/syen.12093 (2015).
Cai, C. et al. Integrated phylogenomics and fossil data illuminate the evolution of beetles. Royal Soc. Open. Sci. 9, 211771. https://doi.org/10.1098/rsos.211771 (2022).
Delgado, J. M. & Palma, R. L. Larval stages of Podaena latipalpis from New Zealand and phylogenetic relationships of the subfamily Orchymontiinae based on larval characters (Insecta: Coleoptera: Hydraenidae). New. Z. J. Zool. 31, 372–342 (2004).
Kilian, A. Phylogenetic analysis of Camiarinae (Coleoptera: Leiodidae) based on larval morphology. Ann. Zool. 62, 11–68. https://doi.org/10.3161/000345412X633667 (2012).
Ashe, J. S. & Watrous, L. E. Larval chaetotaxy of Aleocharinae (Staphylinidae) based on a description of Atheta coriaria Kraatz. Coleopterists Bull. 38, 165–179 (1984).
Thayer, M. K. & Staphylinidae Latreille pp. 394–442. In: Beutel, R. G. & R. A. B. Leschen (eds.). Coleoptera, Beetles. Vol. 1: Morphology and systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). 2nd edition. Handbook of Zoology; Arthropoda: Insecta (ed. R. G.Beutel & N. P. Kristensen); De Gruyter, Berlin/Boston. xvii, 684 pp. (2016). (1802).
Zaytsev, A. A. & Tokareva, A. S. Larval description of Nodynus leucofasciatus Lewis, 1879 (Coleoptera: Staphylinidae: Apateticinae) with discussion of Staphylinidae chaetotaxy. Russ Entomol. J. 30, 482–499. https://doi.org/10.15298/rusentj.30.4.10 (2021).
Zippel, A., Cao, Q. & Betz, O. Morphology of the abdominal segmental glands and spinning behaviour of Stenus larvae (Coleoptera, Staphylinidae). Arthropod Struct. Dev. 75, 101286. https://doi.org/10.1016/j.asd.2023.101286 (2023).
Newton, A. F. & Agyrtidae, L. in Coleoptera, Beetles. Volume 1. Morphology and systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). Handbook of Zoology; Arthropoda: Insecta, 2nd edition (eds Beutel, R. G. & Leschen, R. A. B.) 356–376 (De Gruyter, Berlin/Boston) (2016).
Newton, A. F. Review of Agyrtidae (Coleoptera), with a new genus and species from New Zealand. Ann. Zool. 47, 111–156 (1997).
Zwick, P. Die Jugendstadien des Käfer Necrophilus subterraneus (Col. Silph. Agyrtinae). Beitr. Naturkd Osthessens. 17, 133–140 (1981).
Antunes-Carvalho, C., Ribera, I., Beutel, R. G. & Gnaspini, P. Morphology-based phylogenetic reconstruction of Cholevinae (Coleoptera: Leiodidae): a new view on higher-level relationships. Cladistics 35, 1–41. https://doi.org/10.1111/cla.12230 (2017).
Deleurance-Glaçon, S. Recherches sur les Coléoptères troglobies de la sous-famille des Bathysciinae. Ann. Sci. Nat. Zool. 5, 1–172 (1963).
Wheeler, Q. D. Larval characters of a neotropical Scotocryptus (Coleoptera: Leiodidae). A nest associate of stingless bees (Hymenoptera: Apidae). J. N Y Entomol. Soc. 93, 1082–1088 (1985).
Kilian, A. Comparative morphology of larval Camiarinae (Coleoptera: Leiodidae). Genus Paragyrtodes Szymczakowski, Zootaxa 1640, 1–39 (1966). https://doi.org/10.11646/zootaxa.1640.1.1 (2007).
Kilian, A. & Kleina, A. Comparative morphology of larval Camiarinae (Coleoptera: Leiodidae). Part II. Agyrtodes, Dictydiella and Zearagytodes, with a review of characters of known larvae of Agyrtodini. Ann. Zool. 61, 381–420. https://doi.org/10.3161/000345411X584861 (2011).
Kilian, A. & Mądra, A. Comments on the biology of Sciodrepoides watsoni watsoni (Spence, 1813) with descriptions of larvae and pupa (Coleoptera: Leiodidae: Cholevinae). Zootaxa 3955, 45 – 64 (2015). https://doi.org/10.11646/zootaxa.3955.1.2
Kilian, A. & Newton, A. F. Unique larval morphology of Prionochaeta opaca (Say, 1825) (Coleoptera: Leiodidae: Cholevinae: Cholevini), the only Nearctic representative of Cholevina. Zool. Anz. 292, 111–125. https://doi.org/10.1016/j.jcz.2021.03.005 (2021).
Kilian, A., Kadej, M., Cooter, J. & Harvey, D. J. Larval morphological adaptations of Leiodes cinnamomea (Panzer, 1793) (Coleoptera: Leiodidae: Leiodinae)—obligatory feeder of Tuber species. Insects 13 (3), 249. https://doi.org/10.3390/insects13030249 (2022).
Gnaspini, P. Brazilian Cholevidae (Coleoptera), with emphasis on cavernicolous species. III. Dissochaetus larvae, with description of a new feature. Rev. Bras. Entomol. 37, 545–553 (1993).
Sánchez, P. A., Sánchez, F., Jaffé, K. & Caetano, F. H. Ultrastructure of the gland responsible for the synthesis of an aggregation pheromone in Rhynchophorus palmarum (L.) (Coleoptera: Curculionidae). Coleopterists Bull. 52, 297–305 (1998). http://www.jstor.org/stable/4009337
Frank, J. H. & Thomas, M. C. Cocoon-spinning and the defensive function of the median gland in larvae of Aleocharinae (Coleoptera: Staphylinidae): a review. Quest Entomol. 20, 7–24 (1984).
Leschen, R. A. B. Retreat-building by larval Scaphidiinae (Staphylinidae). Mola 4, 3–5 (1994).
Peck, S. B. & Cook, J. Systematics, distributions, and bionomics of the small carrion beetles (Coleoptera: Leiodidae: Cholevinae: Cholevini) of North America. Can. Entomol. 134, 723–787 (2002).
Quennedey, A., Drugmand, D. & Deligne, J. Morphology and ultrastructure of paired prototergal glands in the adult rove beetle Philonthus varians (Coleoptera, Staphylinidae). Arthropod Struct. Dev. 31, 173–183 (2002). (2002).
Gross, J. & Schmidtberg, H. Glands of leaf beetle larvae – protective structures against attacking predators and pathogens. In Research on Chrysomelidae, Vol. 2. (ed. Jolivet, P., Santiago-Blay, J. & Schmitt, M.) 177–190 (Brill, Leiden) (2009). https://doi.org/10.1163/ej.9789004152045.1-299.64
Vesović, N. et al. The chemical composition of the secretions, their antibacterial activity, and the pygidial gland morphology of selected European Carabini ground beetles (Coleoptera: Carabidae). Front. Ecol. Evol. Sec Chem. Ecol. 11, 1120006. https://doi.org/10.3389/fevo.2023.1120006 (2023).
Kovac, D. & Maschwitz, U. Secretion-grooming in aquatic beetles (Hydradephaga): a chemical protection against contamination of the hydrofuge respiratory region. Chemoecol 1, 131–138 (1990).
Lusebrink, I., Dettner, K. & Seifert, K. Stenusine, an antimicrobial agent in the rove beetle genus Stenus (Coleoptera, Staphylinidae). Naturwissenschaften 95, 751–755 (2008).
Arce, A. N., Smiseth, P. T. & Rozen, D. E. Antimicrobial secretions and social immunity in larval burying beetles, Nicrophorus vespilloides. Anim. Behav. 86, 741–745. https://doi.org/10.1016/j.anbehav.2013.07.008 (2013).
Tafakori, V. Slime molds as a valuable source of antimicrobial agents. AMB Express. 11, 92. https://doi.org/10.1186/s13568-021-01251-3 (2021).
Sułkowska-Ziaja, K. et al. Natural compounds of fungal origin with antimicrobial activity-potential cosmetics applications. Pharmaceuticals (Basel). 16, 1200. https://doi.org/10.3390/ph16091200 (2023).
Wheeler, Q. D. Evolution of slime mold feeding in leiodid beetles in Fungus-Insect Relationships (eds Wheeler, Q. & Blackwell, M.) 446–479 (Columbia University, New York) (1984).
Wheeler, Q. D. Associations of beetles with slime molds: ecological patterns in the Anisotomini (Leiodidae). Bull. Entomol. Soc. Am. 30, 14–18 (1984).
Zhou, L., Meng, G., Zhu, L., Ma, L. & Chen, K. Insect antimicrobial peptides as guardians of immunity and beyond: a review. Int. J. Mol. Sci. 25, 3835. https://doi.org/10.3390/ijms25073835 (2024).
Kölsch, G. The ultrastructure of glands and the production and function of the secretion in the adhesive capture apparatus of Stenus species (Coleoptera: Staphylinidae). Can. J. Zool. 78, 465–475. https://doi.org/10.1139/z99-213 (2000).
Will, K. W., Attygalle, A. B. & Herath, K. New defensive chemical data for ground beetles (Coleoptera: Carabidae): interpretations in a phylogenetic framework. Biol. J. Linn. Soc. 73, 167–168. https://doi.org/10.1006/bijl.2000.0456 (2001).
Steidle, J. L. M. & Dettner, K. Chemistry and morphology of the tergaI gIand of freeliving adult Aleocharinae (Coleoptera: Staphylinidae) and its phylogenetic significance. Syst. Entomol. 18, 149–168 (1993).
Betz, O. & Kölsch, G. The role of adhesion in prey-capture and predator defence in arthropods. Arthropod Struct. Dev. 33, 3–30. https://doi.org/10.1016/j.asd.2003.10.002 (2004).
Bünninge, M., Hilker, M. & Dobler, S. Convergent evolution of chemical defence in Galerucine larvae. Biol. J. Linn. Soc. 93, 165–175 (2008).
McDonald, K. Osmium ferricyanide fixation improves microfilament preservation and membrane visualization in a variety of animal cell types. J. Ultrastruct Res. 86, 107–118. https://doi.org/10.1016/s0022-5320(84)80051-9 (1984).
Reynolds, E. S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell. Biol. 17, 208–212. https://doi.org/10.1083/jcb.17.1.208 (1963).
Fröhlich, A. & Lu, Z. The rosette-like structures in the cuticle of Petrobius brevistylis are the openings of epidermal glands. Arthropod Struct. Dev. 42, 89–94. https://doi.org/10.1016/j.asd.2012.10.001 (2013).
Billen, J. & Wang, C. Intramandibular glands in ants. Myrmecol News. 35, 125–144. https://doi.org/10.25849/myrmecol.news_035:125 (2025).
Jakubec, P. Thermal summation model and instar determination of all developmental stages of necrophagous beetle, Sciodrepoides watsoni (Spence) (Coleoptera: Leiodidae: Cholevinae). PeerJ 4, e1944. https://doi.org/10.7717/peerj.1944 (2016).
Acknowledgements
We are grateful to Alfred Newton (Negaunee Integrative Research Center, Field Museum of Natural History in Chicago, USA) for lending large material from the collection of FMNH. To Anna Derdak (Microscopy Techniques Laboratory, University of Wrocław, Poland) for preparing serial sections. To Miroslav Hyliš and Jana Nebesářová for his support and assistance in this work at the Viničná Microscopy Core Facility (VMCF of the Faculty of Science, Charles University, Prague, Czech Republic), an institution supported by the MEYS CR (LM2023050 Czech-BioImaging). To Mateusz Mostowy (KEYENCE INTERNATIONAL (Belgium) NV/SA) for his support and assistance with microscopy. To Pavel Jakubec (Czech University of Life Sciences Prague, Czech Republic) for collecting adults of S. watsoni watsoni. Maxwell V.L. Barclay (Natural History Museum, London, UK) is thanked for comments and language corrections of a preliminary version.
Funding
This study was financially supported by the “Excellence Initiative – Research University” program at the University of Wrocław. Publication is financed by the program at the University of Wrocław “Excellence initiative–research university” (grant IDUB. 1. 2024; 10120 0320/2020/20 zad.7).
Author information
Authors and Affiliations
Contributions
All authors had full access to all the data in the study and take responsibility for the integrity of the data and the accuracy of data analysis. Study concept and design: AG, AK, JR. Providing materials: AK, JR. Acquisition, analysis and interpretation of data: AG, AK, JR, SN. Wrote the manuscript: AG, AK, JR. Preparing graphic images: AG, AK, JR. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kilian, A., Garbiec, A., Růžička, J. et al. Structure and diversity of abdominal exocrine glands in larvae of Leiodidae (Insecta: Coleoptera: Staphylinoidea). Sci Rep 16, 12426 (2026). https://doi.org/10.1038/s41598-026-41930-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-41930-4
