
Abstract
The demand for fishmeal is increasing, but its supply is stagnating or even declining. There is an urgent need to find an eco-friendly and cost-effective alternative protein source. This study evaluated poultry by-product and insect meal as alternatives to fishmeal for the health performance and bioactivity of Nile tilapia. A Nile tilapia fry was divided into four groups with three replicates (No = 168). The first group was fed a basal diet containing 20% fishmeal (TFM). The second, third, and fourth groups received a basal diet where the fishmeal was substituted with poultry by-product meal (TPM), insect meal from Hermetia illucens (TIM), and a mixture of poultry by-product and insect meal (TMIX), respectively. The overall growth performance data indicated that TFM and TIM significantly achieved the highest growth rates and feed utilization (P < 0.05). The TIM diet significantly exhibited the highest total phenolic content, and both TIM and TFM showed superior antioxidant activity in fish muscle. There were no abnormal hematological or serum biochemical parameters observed in Nile tilapia fed insect meal and/or poultry by-product. The fish fillet samples from all groups were microbiologically safe for human consumption. Fish fed TIM displayed the lowest levels of TNF-α and the highest levels of IL-10 (P < 0.05). All the groups exhibited normal architecture of the internal organs. The significantly highest recorded absorption surface area was found in both TFM and TIM diets (P < 0.05). The NF-κB immunostaining showed no significant changes among experimental groups (P > 0.05). Based on this study, it can be suggested that the insect meal can be a sustainable and cost-effective substitute for conventional fishmeal in aquaculture feed formulations.
Similar content being viewed by others
Introduction
According to a report from the Food and Agriculture Organization1 Egypt ranked first in Africa and sixth globally in aquaculture in 2018. One of the main fish species cultivated in Egypt is Nile tilapia (Oreochromis niloticus), which holds significant economic value2,3. Its popularity can be attributed to its ability to adapt to various environmental conditions, consume a wide range of diets, and exhibit rapid growth4. As global demand for fish continues to rise, driven by population growth and changing dietary preferences, the need for sustainable production of Nile tilapia has become increasingly important5.
For many years, fishmeal (FM) has been the main protein source in aquatic nutrition due to its palatable flavor, balanced amino acid profile, and ease of digestion, which are vital features for improving nutrient absorption and utilization6. Natural sources of fishmeal have remained stable over the past decade, but demand and prices are rising drastically7. This highlights the need for sustainable, inexpensive, low-trophic substitutes for aquaculture protein.
Plant-based protein sources, such as soybean meal, corn gluten, peanut meal, and rapeseed meal, are frequently utilized as alternatives to fishmeal. These substitutes are favored due to their widespread availability, competitive pricing, and consistent supply8. However, these ingredients often exhibit nutritional limitations, such as amino acid imbalances and anti-nutritive elements, which hinder nutrient digestion and absorption, resulting in low utilization for aquatic animals9,10.
Animal by-products such as poultry by-product meal (PBM), meat meal, and tankage offer major potential as cost-effective diet components in fish production11. These ingredients are excellent sources of high-quality protein, essential amino acids, and energy content, like FM12. It has considerable potential as a protein source to substitute for fishmeal in the fish feed. However, animal by-products may be deficient in one or more essential amino acids. The most limiting essential amino acids (EAA) in this by-product are typically lysine in PM. The deficiencies can be overcome in the diet by supplementing with synthetic lysine13. Koch et al.14 reported that the use of 30% PBM and 20% soybean meal (SBM) with supplemental lysine, methionine, and taurine achieved the best performance in the growth of juvenile Nile tilapia. These findings support the strategic incorporation of PBM in aquafeed formulations to balance nutritional performance, economic feasibility, and environmental sustainability.
Recently, more innovative raw materials, such as insect meal (IM), have been proven to be highly valuable as an unconventional protein source15,16. Several studies on the partial and total substitution of insect meal for FM in fish and shrimp culture exist17,18,19. Owing to its high nutritional content, affordable price, and accessibility, insect meal has been used in aquatic nutrition20. Additionally, there is an increasing global interest in insect protein, not just for animal feed but also for human food systems, backed by supportive regulatory frameworks and an increase in consumer acceptance21. A variety of insect species are used in aquatic feed, with the black soldier fly (Hermetia illucens) being particularly valued. This species effectively converts food waste into high-quality protein. Its larvae have a crude protein content of 42.1%, whereas defatted one has 56.9%, which is slightly less than fish meal and comparable to soybean meal. Moreover, the amino acid profile of larvae is superior, making them a favorable alternative to fish meal22. In Nile tilapia, 80 g/kg inclusion of black soldier meal (BSM) successfully substituted for 70 g/kg of fish meal (FM) and 350 g/kg of soybean meal (SBM) without adversely affecting growth rate or feed efficiency23. In modern aquaculture, fish farmers prioritize economic returns, as feed costs represent nearly half of their operating expenses. Reducing these costs is vital for profitability and sustainability. So, BSM offers a cost-effective protein alternative18.
When introducing new ingredients into aquafeeds, it is essential to ensure they do not adversely affect fish growth, health, or product quality. While poultry by-product meal has been increasingly explored, studies on the use of defatted black soldier fly larvae meal remain limited, particularly in combination with other alternative proteins. Therefore, Oreochromis niloticus (Nile tilapia) was selected as a model species to evaluate the effects of substituting fish meal with poultry by-product meal and/or Black soldier fly larvae meal (BSFLM) on growth performance, economic efficiency, chemical composition, carcass morphometric indices, hematology, serum biochemistry, liver cytokine expression, muscle microbial load, organ histomorphology, and NF-κB immunohistochemical responses.
Materials and methods
Ethical agreement
All experimental procedures involving fish were conducted following the guidelines and regulations of the Faculty of Veterinary Medicine, Suez Canal University, Egypt. The experimental protocols were reviewed and approved by the Institutional Animal Care and Use Committee of Faculty of Veterinary Medicine, Suez Canal University, Egypt (Approval No: SCU-VET-AREC-R-2025020). All methods are reported in accordance with the ARRIVE guidelines (https://arriveguidelines.org).
Experimental diet
Insect meal was sourced from EGY MAG® Biotechnology Company in Egypt. The larvae were cultivated using food residues, specifically organic matter and waste from fruits and vegetables. They were harvested 14 days before reaching the pupal stage and then oven-dried at 50 °C for 24 h. After drying, the larvae were processed into a uniform powder using a feed mill and stored at 4 °C until needed. The insect meal contains the following nutritional values: 5281.9 kcal/kg of gross energy, 55% crude protein, 2.5% calcium, 1% phosphorus, 2.1% lysine, and 0.9% methionine. Diet ingredients such as fish meal, poultry byproduct, soybean meal, corn gluten, yellow corn, wheat flour, and sunflower oil were acquired from a feed enterprise. The ingredients were ground into a fine powder, analyzed for proximate composition, and then processed into 2.5 mm pellets using a feed pelleting machine. The pelleted diets were dried at 25 °C for 12 h in a cool, ventilated space and then stored at − 20 °C until needed. The experimental diets were formulated to contain 33% crude protein (isonitrogenous) and 4588.91 gross energy Kcal/kg diet (isocaloric). Diets were subjected to chemical analysis and were determined according to AOAC24 as shown in Table 1.
Experimental design and feeding regime
A total of one hundred sixty-eight healthy Nile tilapia (Oreochromis niloticus) fish fry were collected from the Fisheries Research Institute at Suez Canal University (SCU) and transferred to the Farming and Technology Institute at Suez Canal University (SCU) for the experiment. Initially, the fish were acclimated for 2 weeks and fed a basal meal. After this adaptation period, they were randomly assigned to four groups (initial weight 11.99 ± 0.08 g). Each group consisted of 42 fish and was then divided into three replicates (14 fish per replicate). Each replicate was put and fed separately in a glass aquarium (90 × 50 × 40 cm), featuring 30% daily water changes using clean, dechlorinated water. The first control group (TFM) was fed a basal meal containing 20% fish meal. The second, third, and fourth groups were fed a basal meal where the fish meal was replaced with poultry by-product meal (TPM), insect meal (TIM) sourced from de-fatted black soldier fly larvae (Hermetia illucens), and a mixture (1:1) of poultry and insect meal (TMIX), respectively. The fish were fed at a rate of 3% of their body weight twice daily, at 8:00 a.m. and 2:00 p.m., for 10 weeks. Mortality was monitored daily, and the fish mass was measured every 2 weeks to adjust feeding amounts accordingly. The aquaria were equipped with automatic aerators, and daily monitoring of dissolved oxygen, pH, and temperature was conducted. Temperature, pH, and dissolved oxygen were measured during the experimental period, and it was recorded at 31.09 ± 0.51 °C, 7.8 ± 0.24, and 4.16 ± 1.36 mg/L respectively. Also, water conditions were maintained at ammonia levels below 0.1 mg/L, and nitrate concentrations under 1.7 mg/L25.
Sampling
The body weight of fish from all groups and replicates was measured every 2 weeks. After 10 weeks of being fed the experimental diets, the fish were subjected to various analyses. To minimize handling stress, the fish were fasted for 24 h before sampling, and three fish from each replicate were anesthetized with a clove oil solution (12.5 mg/L)26. Blood samples were collected from the caudal vein using a clean syringe and divided into two portions. One portion was placed in heparinized Eppendorf tubes for hematological assays, while the other portion was transferred to non-heparinized tubes. For biochemical analysis, serum from the non-heparinized blood was obtained through centrifugation at 3500×g for 15 min. Additionally, at the end of the experimental period, fish were humanely euthanized using an overdose of clove oil (200 mg/L) until complete cessation of opercular movement was observed, after which immediate dissection and tissue collection were performed. Then, three other random fish samples from each replicate were taken to assess carcass indices. Also, three fish from each replicate were used to determine the microbial quality of the fish. Another three random fish samples were stored at − 20 °C for proximate analysis. Frozen muscle and liver samples were preserved in labeled Eppendorf tubes at − 20 °C to evaluate total phenolic content, antioxidant activity, Malondialdehyde (MDA) content, and liver cytokine assays. For histopathological examination, tissues from the intestine, liver, kidney, and spleen were removed and immediately fixed in 10% formalin.
Growth performance
Every 2 weeks, all fish from each replicate were weighed to determine the following growth indicators as follows:
Economic evaluation
The feed cost to produce one kilogram (kg) of body weight at the end of the study period was analyzed to evaluate the economic parameters of the control diet vs the test diet27. Detailed economic evaluation data are provided in Table S1 (Supplementary Materials).
Chemical composition and bioactivity profile of experimental diets and carcasses of Nile tilapia
Proximate composition of experimental diets and whole carcasses
Experimental diets, and whole fish carcasses were analyzed for moisture, crude protein, crude fat, and ash following the methods outlined by24. Moisture content was determined by drying the samples at 105 °C until a constant weight was achieved. Crude protein was assessed using the Kjeldahl method (Kjeldahl-ATN-300 BonninTech, China), with nitrogen content multiplied by 6.25 to calculate the protein content. Ash content was analyzed by incinerating the samples at 550 °C for 12 h. Crude fat was quantified using the Soxhlet method with extraction by petroleum ether. All analyses were conducted in triplicate.
Total phenolic content (TPC) in experimental diets and Nile tilapia muscle
The Folin–Ciocalteu technique was used to determine the total phenolic content28 with slight modifications. First, extraction was carried out by adding 50 mL of methanol to 1 g of the sample, which was then homogenized for 4 h at 25 °C and filtered. Next, 900 μL of Folin–Ciocalteu reagent was mixed thoroughly with 100 μL of the extract and allowed to stand for 5 min. After that, 0.75 mL of a 7% sodium carbonate solution was added to the mixture, which was vortexed for 30 s and then left to settle in the dark for 60 min. The absorbance was measured using a PG spectrophotometer (PG Instruments Ltd., Felsted, Dunmow, UK) at a wavelength of 725 nm. The phenolic content was calculated using gallic acid as a standard and expressed as mg/100 g on dry basis.
Antioxidant activity determination in experimental diets and Nile tilapia muscle through DPPH assay
According to Tamsen et al.29, 2,2-diphenyl-1-picrylhydrazyl (DPPH) was utilized as a free radical to assess antioxidant activity. One gram of the sample was mixed with 50 mL of methanol and shaken for 3 h at room temperature. Afterward, the mixture was centrifuged for 20 min at 3000 rpm. Next, 3.9 mL of the DPPH methanol solution was combined with 100 μL of the methanolic extract (supernatant) of the sample. This mixture was then incubated at room temperature for 30 min in the dark. Finally, the absorbance was measured at 517 nm using a PG spectrophotometer. To determine the % DPPH radical scavenging activity, the following formula was used:
Determination of MDA in Nile tilapia muscle
The malondialdehyde (MDA) content of fish muscle was measured using fish MDA ELISA kits (Cat. No. EK750261) from AFG Bioscience LLC, Northbrook, Illinois, USA, following the method of Botsoglou et al.30.
Carcass morphometric indices
The carcass morphometric indices were determined as follows18:
Hematological parameters
Hemoglobin (Hb), packed cell volume (PCV), mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), and platelet values were measured using a blood cell analyzer (Tecom 5000, 2017, China). The red blood cell (RBC) and white blood cell (WBC) counts were determined according to the method described by31. Differential counts of lymphocytes, neutrophils, monocytes, eosinophils, and basophils were identified by smears stained with Wright Giemsa.
Serum biochemical parameters
The aspartate aminotransferase (AST), alanine aminotransferase (ALT), and alkaline phosphatase (ALP) were detected colorimetrically by a semi-auto chemistry analyzer (Mindray, India) using chemical kits provided by Bio Med® Diagnostic Co., Egypt, following the manufacturer’s instructions. Serum total proteins and albumins were measured according to32, while globulin and A/G ratio were calculated mathematically. Creatinine and urea were measured calorimetrically using available kits from SPECTRUM® Co., Egypt, according to standard protocols33. Serum cholesterol, triglycerides, high-density lipoprotein (HDL), and low-density lipoprotein (LDL) were determined by atomic absorption spectrophotometry using the commercial kits provided by Bio Med ® Diagnostic Co., Egypt, while very low-density lipoprotein (VLDL) was calculated mathematically.
Liver cytokines assay
Liver tissues were collected and homogenized in a glass homogenizer at a ratio of 1:10 in phosphate-buffered saline (PBS) with a pH of 7.4. After homogenization, the mixture was centrifuged at 5000×g for 5 min at a temperature between 6 and to 8 °C. The supernatant was then carefully removed for cytokine analysis.
For measuring fish interleukin-10 (IL-10) levels in the liver, fish IL-10 ELISA kits (Cat. No. QS0059FI SL0043Ch) from Sun Long Biotech Co., LTD, China, were utilized following the manufacturer’s instructions. Additionally, fish tumor necrosis factor-α (TNF-α) levels were quantitatively determined using fish TNF-α ELISA kits (Cat. No. SL0055FI) from Sun Long Biotech Co., LTD, as per the manufacturer’s protocol.
Microbial load examination
Three fish samples were collected from each replicate to assess the microbial load. Aseptically, 10 g of the blended sample were removed from the Petri dish, and 90 mL of sterile buffered peptone water was added. After 2 min, the samples were homogenized. The pour plate method (Merck, Darmstadt, Germany) was employed to determine aerobic plate counts (APC). Violet red bile (VRB) agar was used to measure the total coliform count, while Escherichia coli (E. coli) colonies were grown on eosin methylene blue (EMB) agar plates to confirm the presence of typical purple colonies. On Slant Agar, presumed colonies that appeared blue–black with dark centers and a green metallic sheen were streaked. The results are reported as log CFU/g of sample. Molds and yeasts were identified using plate count agar that contained 100 µg/mL of cidostane34.
Organ histomorphology
Liver, kidney, spleen, and the initial segment of the small intestine samples were gathered from each fish (three fish/group). Organs measuring about 0.5 mm were preserved for 24 h in a 10% buffered neutral formalin solution, dehydrated using a sequence of increasing ethanol concentrations (from 70 to 100%), cleaned in xylene, and then embedded in paraffin wax. Paraffin sections were cut with a microtome to a thickness of 5–7 μm (Leica RM 2155, England). Routine staining procedures were conducted using Harris’ Hematoxylin and Eosin (H&E) stain35. Photomicrographs were captured using an Olympus BX-41 research microscope, equipped with a digital AMT camera and its image-capturing software (AMT V600.259). 50 well-aligned villi were inspected from each section of all intestinal segments to measure the intestinal villi length, width, and absorption surface area. The intestinal villi length was assessed from their tip to the base, and the width was assessed at the half-height point. These parameters were analyzed using Image J software (version 1.33–1.34; National Institutes of Health, Bethesda, MD, USA). Absorption surface area was calculated as follows: ASA (mm2) = villus height × villus width36.
Immunohistochemical investigation
Liver samples were preserved in 4% paraformaldehyde at pH 7.4 for 48 h. The fixed tissue was processed on positively charged slides for NF-κB immunostaining, deparaffinized in xylene, and rehydrated through decreasing alcohol concentrations. Sections were treated with an endogenous peroxidase blocking solution (DAKO reagent, Cat. No S2001)35. The primary antibody used was anti-mouse polyclonal NF-κB (1:100 dilution, catalog # sc-8008, Santa Cruz Biotechnology, Heidelberg, Germany). Slides were rinsed three times in 0.1 M PBS (pH 7.4) with 0.5% Triton X-100 for 5 min each and then incubated for 4 h at room temperature with biotinylated goat anti-mouse IgG (1:600, catalog # 31800, Invitrogen, Waltham, MA, USA). For detection, slides were treated with 3,3′-diaminobenzidine (DAB) for 30 min and counterstained with Mayer̛ s hematoxylin. The slides were then examined under a microscope for target protein expression35. The percentage of immunoreactivity intensity was determined using Image J software (version 1.33-1.34; National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
In this study, statistical analysis using the SPSS program (SPSS Statistics 20 for Windows) we was conducted to compare the means and standard errors of various groups. It was performed using a one-way ANOVA, followed by a Duncan test as a post hoc analysis to identify differences among the groups37. For each parameter, we reported the mean ± standard error (SE), along with P values. The significant differences were considered at P < 0.05.
Results
Growth performance
Table 2 displays the overall growth data of Nile tilapia. The final BW, weight gain, WG%, and SGR% of fish in TIM were not differ significantly from those of fish in TFM. Besides, TFM and TIM significantly achieved the best FCR. The cumulative feed intake and protein intake didn’t reveal significant changes among groups. TFM had the greatest PER than TPM and TMIX. PER of TIM was not significantly different from TFM, TPM, or TMIX.
There was no significant difference between survival rates of the TFM, TIM, and T PM groups. While TMIX had the lowest survival rates (Table 2).
Chemical composition profile of experimental diets and carcasses of Nile tilapia
Table 3 presents the results of the chemical composition assay. The whole-body composition analysis revealed that TFM and TIM had the highest moisture content, while TPM and TMIX had slightly lower moisture levels. Significant differences were noted in the protein content, with TMIX exhibiting the highest protein value, whereas TIM had the lowest at 20.88%. Additionally, TMIX had the significantly highest fat content, while TFM recorded the lowest fat value (P < 0.001). TFM also had the highest ash content, followed by TIM, while TPM had the lowest ash content.
The total phenolic content of the experimental meals showed significant variation among the groups (P < 0.001). The highest phenolic content was found in the TIM group, followed by TFM and TMIX, while TPM displayed the lowest level. Additionally, the total phenolic content in the fish muscle reflected the trends observed in the meals, with a significant increase noted among treatments (from TPM to TMIX/TFM to TIM). Similarly, the antioxidant activity (%) in the meals also differed significantly between treatments (P < 0.001). The TFM group exhibited the highest antioxidant activity, whereas TPM had the lowest. In terms of muscle antioxidant activity, significant variation was present, with TFM and TIM recording the highest values, while TPM again had the lowest. Notable differences were also seen in the MDA content of the muscle among the groups, with TPM registering the highest value and TIM showing the lowest.
Carcass morphometric indices
TFM and TIM showed significantly higher live body weights and dressing weights. TFM and TIM had significantly heavier livers than TPM and TMIX. There were no significant changes detected in HIS, visceral weight, and visceral index among treatments (P > 0.05), Table 4.
Hematology (complete blood picture, CBC)
The means of red blood cells, PCV, Hb, MCV, MCH, and MCHC were not statistically different among groups. White blood cells, differential leukocyte counts (lymphocytes, neutrophils, monocytes, eosinophils, and basophils), and platelets had minimal changes among groups with no significant effect detected (P > 0.05). The hematological indices varied from the normal values for healthy fish (Table 5).
Serum biochemical parameters
Regarding the data related to the liver function test. For the ALT level, TPM had the lowest significant value compared to TMIX. No significant differences were detected among groups in AST, ALP, total protein, albumin, globulin, and A/G ratio. For the creatinine level, there was a significant difference observed (P = 0.003). TPM was significantly higher than the other groups. For urea, no significant differences among groups were found. Concerning the data related to the lipid profile, TMIX showed higher cholesterol levels, either significantly compared to TPM or numerically compared to TFM and TIM. Other parameters, including triglycerides, HDL, LDL, and VLDL, did not differ significantly among groups (P > 0.05), Table 6.
Microbial load of muscle fillets
Table 7 presents the microbial load of fish muscle fillets. Fish samples from the TFM and TMIX groups exhibited statistically similar Aerobic Plate Count (APC) values, which were significantly higher than those of the TPM and TIM groups (P = 0.001). Fish from the TIM group had a significantly lower total coliform count compared to the other groups (P < 0.001). Additionally, fish from the TFM and TPM groups did not differ statistically from each other, while the TMIX group had the highest coliform count (TIM < TFM/TPM < TMIX). No colonies of E. coli, yeast, or mold were detected in any of the fish fillet samples from all groups.
Liver cytokines assay
TIM significantly showed the lowest liver TNF-α level compared to TFM, TPM, and TMIX (P < 0.05). For IL-10 level, significant differences were shown among groups (P < 0.05), with the TIM group exhibiting higher levels followed by TFM, TMIX, and then TPM (Fig. 1).

Liver TNF-α and IL-10 (pg/mL) level among treatment groups at 10 weeks. Means having separate letters are significantly different from each other, P < 0.05.
Organ histomorphology
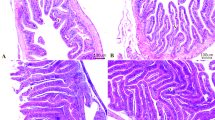
All the experimental groups exhibited a normal architecture of the intestine. There were no noticeable signs of inflammation or damage (Fig. 2A–D). Morphometric analysis of intestinal sections by ImageJ software is presented in Table 8. TFM had the maximum villous width compared to the TIM and TMIX groups. The significantly highest recorded absorption surface area (ASA) was detected in the TFM and TIM groups compared to TMIX.

Photomicrograph of H&E-stained sections from the intestine of N. tilapia (Scale bar 100 μm) showing: normal architectures of simple columnar enterocytes lining mucosal villi (V), submucosal layer and muscular layer (M) (A–D). Enhanced absorption surface area in the TFM group (A) and TIM group (C), followed by the TPM group (B). The lowest values of intestinal parameters at the TMIX group (D).
All experimental groups displayed normal liver structure, including lipid droplets in the cytoplasm of the hepatocytes and central hepatocyte nuclei. Also, normal exocrine pancreatic acini were observed. There were no indications of capillary hyperemia, vacuolar degeneration, vasodilatation, or hepatocyte ballooning in the different groups (Fig. 3A–D).

Photomicrograph of H&E-stained sections from the liver (Scale bar 20 μm) showing: normally arranged vacuolated hepatic cells (H), exocrine pancreatic acini (PA), and portal vein (PV) in all examined groups (TFM, TPM, TIM, and TMIX) (A–D).
All experimental groups showed normal architecture of renal parenchyma with well-defined renal glomeruli and tubules (Fig. 4A–D).

Photomicrograph of H&E-stained sections from Kidney (Scale bar 20 μm) showing: normal morphology of renal tubules (arrows), glomerular corpuscles (arrowheads), and other stromal structures in all examined groups (TFM, TPM, TIM, and TMIX) (A–D).
The histological analysis of the spleen showed a normal structure of white pulp around ellipsoidal arterioles in all examined groups. Moreover, areas of melanomacrophage centers (MMCs) beside normal red pulp were noticed (Fig. 5A–D). The size of white pulp lymphoid follicles increased in TFM, TPM, and TIM groups (Fig. 5A–C) in comparison with the TMIX group (Fig. 5D). An Abundant area of MMCs within white pulp was observed in the TMIX group (Fig. 5D).

Photomicrograph of H&E-stained sections from spleen (Scale bar 20 μm) showing: (A) normal histological structures of white pulps (WP) around ellipsoids arterioles (arrowheads) with areas of melanomacrophages centers beside normal red pulps (RP) in all examined groups (A–D). Increased size of white pulp lymphoid follicles at group TFM, TIM, and TPM in comparison with group TMIX. Abundant area of melanomacropahges centers (MMCs) within white pulp areas at group TMIX (D).
Liver NF-κB immunohistochemistry
Immunostaining for NF-κB marker revealed negative expression in all dietary experimental groups (Fig. 6I). The mean percentage area of NF-κB immunostaining intensity showed no significant changes among experimental groups (Fig. 6II).

(I) Immunohistochemical detection of NF-κB expression in hepatic tissue of N. tilapia. photomicrographs showing predominantly negative NF-κB immunoreactivity (blue arrow) in hepatocytes and hepatopancreatic cells among all experimental groups: A (control, TFM), B (poultry meal, TPM), C (insect meal, TIM), and F (insect meal + poultry meal, TMIX). Rare, weakly positive cells showing non-specific staining (red arrows). scale bar = 100 μm. (II) The percentages of expression of NF-κB in the hepatic tissue of all experimental groups.
Discussion
The overall growth performance indicated that the insect meal group (TIM) achieved the greatest growth data and nutrient utilization, as did the fish meal group (TFM). These results were in harmony with Devic et al.38 and Nairuti et al.39, who demonstrated that incorporating various levels of BSFLM as an alternative to fishmeal did not negatively affect growth indices. Further, Tippayadara et al.6 showed that growth performance was unaffected by the addition of BSFLM up to 100% in N. tilapia diets. Muin et al.40 noted that Tilapia could ideally consume BSFLM at a maximum inclusion level of 50%. However, the specific growth rates (SGR) and feed conversion ratios (FCR) were not adversely impacted by replacing fish meal with BSFLM up to 100%. Wachira et al.41 found that Nile Tilapia fed a diet supplemented with up to 67% BSFLM did not show any compromise in growth quality measures.
The improved growth indices observed in the TIM group, similar to those in the fish meal group, can be attributed to the inclusion of black soldier fly larvae in their diets. BSFLM is rich in lauric acid, chitin, and antimicrobial peptides, which may enhance fish welfare, reduce the prevalence of aquatic diseases, and increase resistance to bacterial and parasitic infections. Additionally, it has been reported that adding BSFLM to diets increases the biodiversity of intestinal bacterial composition, which is often associated with the host’s health in various fish species42. Furthermore, BSFLM is recognized as a valuable source of omega-3, omega-6, and omega-9 fatty acids43,44, a structure that could enhance the host’s growth performance. Lastly, dietary insect meal may increase the mucosal surface area in fish, potentially explaining the improvements in feed conversion ratio and feed utilization45.
In contrast, Dietz and Liebert46 reported that N. Tilapia experienced negative effects when soy protein concentrate was completely replaced with 100% partially defatted black soldier fly (BSF) meal. Fayed et al.47 found that the growth rate of N. tilapia decreased when their diet included 30% fish meal (FM) replacement with BSF larvae meal. These variations in growth performance may stem from differences in the composition of the insect meal and the experimental conditions used. In a related study, Guerreiro et al.48 also showed that switching from 17%, 35%, and 52% FM to BSFLM resulted in a decrease in the growth of meagre. These negative effects could be attributed to the presence of chitin, which may hinder growth performance and feed utilization in these species49. As an omnivorous species, Nile tilapia has a high capacity for consuming plankton and possesses certain advantages when it comes to breaking down chitin. The digestion of chitin relies on chitinolytic enzymes, which are vital to the digestive physiology of Nile tilapia50. Furthermore, incorporating chitin into their diet may help combat bacterial infections and enhance the diversity of the gut microbiota51.
Despite variations in final weight, feed intake per fish remains consistent across different treatments. This suggests that factors such as feed quality or composition may be more important for weight gain than the quantity of feed provided. This is in accordance Limbu et al.52, who noticed that N. tilapia fed diets supplemented with BSFLM up to 75% did not show any noticeable changes in feed intake. On the other hand, earlier research revealed that feeding N. tilapia up to 100% BSFLM reduced their feed intake53. This was correlated to the decreased palatability of the feed. These differences may arise from the methods used in insect meal preparation (full-fat or defatted meal), additional dietary components, and the duration of the experiment.
TPM and TMIX demonstrated improved growth parameters, showing no significant difference from TIM. Poultry by-product meal is a popular alternative to fish meal (FM) in aquaculture feed formulations due to its wide availability, high protein content, and excellent source of phospholipids and cholesterol54. Consequently, numerous studies have explored various fish and crustacean species fed diets containing different amounts of poultry by-product meal. However, the findings of these studies have varied considerably, as poultry meals can differ in digestibility, processing methods, nutrient composition, and proportions of their components (bone, meat, blood, etc.). Nevertheless, when high-quality poultry meal was used, many species were able to accept up to 100% substitution levels55.
Survival rate is a vital parameter in determining the production efficiency of Nile tilapia. Fish physiological activities have a chief role in their survival rates; thus, appropriate feeding schedules and accommodation of fish to their habitats are critical. TFM had a perfect survival rate; also, the TIM group and TPM group showed normal survival rates. This suggests excellent conditions conducive to the health and growth of experimental diets. In the same trend, Devic et al.38 showed that tilapia fish consumed a diet including BSFLM at 80% had the highest survival rate (90%), while the group fed 30% BSFLM had the lowest survival rate (81%). Also, Tippayadara et al.6 declared that BSFLM up to 100% did not adversely impact the survival rate in tilapia fish. The survival rate of the fry was unaffected by substituting BSFLM diets at all levels for FM41,52, similar data were recorded in European sea bass (Dicentrarchus labrax)56. Moreover, Ushakova et al.57 found that the survival rate of Mozambique tilapia fed BSFLM pre-pupae at a rate of 0.5 g/kg feed did not differ significantly. TMIX had the lowest survival rate but did not differ significantly from TPM. This drop in the survival rate of TMIX may indicate some underlying issues, such as minor stressors. Most mortalities were noted a day after weekly weighing or sampling, which could be due to sustained stress.
In our study, the proximate composition of the whole body revealed significant differences among the experimental groups. Proximate analysis plays a crucial role in the food industry, particularly for food product development and quality control58. A rise in moisture content was directly linked to higher protein levels in animal body tissues, attributed to the superior water retention capacity of proteins59. Our results revealed that the moisture and crude protein content of TIM did not significantly differ from the body composition of the TFM group. The crude fat content was significantly elevated in all treatments compared to TFM. Ash content varies, suggesting alterations in mineral composition among groups.
The current findings on the effects of diet composition on meat and protein content were in line with other researchers. Muin et al.40 showed that BSFLM addition in the diet had a variable degree of influence on the crude fat content of the fish body, where increased crude fat levels were found in O. niloticus. The current findings support Mahmoud et al.60, who concluded that body lipid was increased with higher FM substitution with PM in N. tilapia diets. It can be due to the high fat content of the poultry byproducts, viscera, and skin61.
In contrast, other studies observed that the diet composition had no significant effect on protein and fat content in fish flesh62,63. Also, Devic et al.38 revealed comparable outcomes when examining the proximate composition of N. tilapia fed different amounts of BSFLM.
It was stated that when 100% poultry by-product meal was added, the tilapia carcass proximate composition showed no change in moisture, lipid, protein, or ash content64. The alterations may be due to the varied quality of PM, which was significantly influenced by their processing methods.
The total phenolic content in the fish muscle mirrored the trends observed in the diets. It increased significantly among treatments (TPM → TMIX&TFM → TIM). Besides, the muscle antioxidant activity recorded the highest values in TFM and TIM. BSFLM had a higher level of phenolic compounds and antioxidant activity, which are influenced by their rearing substrate and processing methods. Their antioxidant properties derive from phenolics, peptides, chitin, and tocopherols in larvae44. Additionally, feeding BSFLM with polyphenol-rich agricultural by-products can significantly enhance their bioactive profile, making BSFLM a promising functional feed ingredient for improving oxidative stability65. From a practical perspective, the increase in phenolic content in fish muscle has important implications for product quality and shelf life. Enhanced antioxidant levels in fish tissue can reduce lipid oxidation, improve sensory attributes, and potentially offer added nutritional benefits to consumers. Furthermore, these findings support the strategic inclusion of phenolic-rich ingredients in aquafeeds as a functional approach to improving fish health and resilience. It was also reported that TIM had the lowest MDA levels in fish muscle, a marker of lipid peroxidation, suggesting superior oxidative stability and potential anti-inflammatory effects of the insect-based meal.
Our findings indicated that TFM and TIM significantly enhanced live body weight and dressing weights in N. tilapia. This suggested that both meals provided excellent nutrition, supporting growth and carcass yield. The increased body weight was likely due to the high-quality protein and favorable amino acid profiles in fish meal and BSFLM diets, which meet the needs of rapidly growing fish66. Moreover, the significantly heavier liver weights observed in fish fed fishmeal and BSFLM could indicate higher metabolic activity or nutrient storage capacities. In fish, liver size can reflect both growth rate and metabolic processing of nutrients67, suggesting that TFM and TIM diets promoted not only somatic growth but also internal organ development with better feed utilization efficiency. These results are consistent with previous studies that have reported the effectiveness of black soldier fly larvae27,52. The visceral index was used as an indicator of gut health since the viscera impact digestion, secretion of enzymes, and nutrient absorption. They are frequently used to evaluate the biological states and nutritional attributes of fish18. Our study showed that TIM does not affect visceral index and gut health in Nile tilapia. This agreed with research by Tippayadara et al.6, who mentioned that the level of BSFLM up to 100% in Tilapia diets did not have harmful effects on somatic indexes. Also, Renna et al.68 proved the same findings in yellow catfish.
Hematological indices of fish were considered important components for estimating the overall health condition and biological stress responses of fish fed formulated diets69. Our study verified that N. tilapia fed on insect meal and poultry by-product meal or a mixture of both did not have an abnormal effect on hematological parameters, and the values were considered within the normal range for healthy fish69. This result was in agreement with studies, which reported that substitution of fish meal with insect meal had no adverse effect on hematological values in European sea bass, hybrid tilapia, and N. tilapia fish56,70,71. Also, it was observed that poultry by-product meal did not change hematology data in gilthead seabream72. Conversely, another study implied that there was an increase in the hemoglobin level of Mozambique tilapia that received a diet supplemented with black soldier fly pre-pupae57. The differences among these results may have been related to protein source quality and processing, fish species and size, experimental period, and culture systems.
Biochemical parameters were used to inspect the effects of feed additives, detect stress, and assess the possible negative impact of immunostimulants on the immune system of the fish73. The liver markers did not display significant variation among treatments. All detected values remained in the normal range for liver enzymes (28.3–121 U/L)74. This consistency among groups indicated that these alternatives did not affect overall liver integrity and protein metabolism and offered hepatoprotective effects similar to those of fishmeal. In terms of kidney function, all creatinine levels showed consistent values with the reference limits (0–0.8 mg/dL)75. In addition, urea levels did not significantly differ among groups, indicating a stable nitrogen metabolism and excretion rate. Plasma urea content in aquatic animals is the second most important nitrogen excretion product after ammonia, whose changes were used to evaluate the digestion of amino acids, proteins, and kidney function76. The lipid profile results demonstrated that TMIX had significantly higher cholesterol levels compared to TPM, and a numerical increase compared to TFM and TIM. The elevation in cholesterol under TMIX treatment may reflect alterations in lipid metabolism or absorption; however, since no significant alterations were detected in triglycerides, HDL, LDL, and VLDL levels, the overall lipid metabolic status appeared to remain stable among groups. Overall, most liver, kidney, and lipid parameters remained unaffected by the dietary treatments. These findings suggest that the tested diets are generally safe for tilapia health.
Our findings came in harmony with Oliveira et al.77 who found that blood parameters in N. Tilapia (creatinine, total serum protein, HDL, LDL, AST, and ALT) showed no differences between treatments containing 0, 33, 66, and 100% BSFM as a substitute for FM. Also, FM with BSFM replacement up to 140 g/kg BSFM has no effects on total protein, albumin, globulin, AST, and ALT in carp78. Dietary BSFM does not affect plasma metabolites, such as total protein, albumin, globulin, and total lipids in snakehead juveniles62.
In contrast, total cholesterol and circulating triglycerides were lower in the animals fed 100% of BSFLM in their diet. Also, for Jian carp, with a drop in cholesterol levels when fed diets containing 2.6–10.6% BSFM (lowering FM from 7.5 to 0%)79. Fish fed 100% BSFLM replacing FM had lower albumin values. High values for albumins could be associated with an impaired immune system in tilapia or protein synthesis in tilapia liver tissues80. Besides, Abdullahi et al.81 showed that serum albumin and plasma urea levels in the diet containing 50% and 100% PM were augmented compared to the FM group. They reported that the triglyceride levels significantly reduced compared to those in the control group. Lin and Luo82 revealed that the amount of liver enzymes of AST, ALP, and ALT increased significantly with the replacement of 100% PBM with fishmeal. These differences in biochemical parameters can vary depending on various issues such as season and environmental circumstances, and stressors, even within the same species77.
The microbial quality of fish fillets can be directly impacted by feed if it is microbiologically deficient, and indirectly by inadequate breeding conditions and management, which can alter water parameters83. It was observed that the APC counts decreased in TPM and TIM, due to their synergistic influence. Furthermore, all detectable values in the different groups were below the maximum allowable limit of 7 log cfu/g, as specified by the International Commission on Microbiological Specifications for Foods84 for fresh fish. Therefore, these values in all diets did not pose a significant risk to public health. The findings indicate that including insect meal in diets did not have a significant impact on the microbiological profile of the fish. Stenberg et al.85 noted that insect meals contain high levels of antibacterial agents and bioactive components that enhance the overall health of fish. Also, chitin and antimicrobial peptides present in larvae can be utilized to create new antimicrobial products, possibly decreasing the need for antimicrobial medications in aquaculture86. The existence of coliform bacteria in fish indicated environmental contamination, as coliforms were not part of the normal bacterial flora in fish. The standard limits of total coliforms and fecal coliforms for fresh water were 100 MPN/g87. Our findings revealed that the TIM sample was within the acceptable limits due to the antimicrobial activity of insect diets. Rimoldi et al.88 mentioned that high-fat content and carbohydrates in insect diets could modify microbial populations. Notably, all samples tested negative for Escherichia coli, yeast, and mold, indicating effective inhibition of pathogenic and spoilage organisms among groups. This suggests that the tested fish groups were microbiologically safe for human consumption.
Within our results, the TIM group had the lowest liver TNF-α and the highest IL-10 levels compared to other groups. It suggested that Tilapia fish fed BSFLM were in a healthy state without being exposed to toxic environments or being infected by pathogens. These findings highlighted the crucial role of cytokines in regulating the inflammatory process, which is vital for modulating immune response in both health and disease. TNF-α, recognized as the initial proinflammatory cytokine released in response to pathogens, amplifies the acute phase of the immune response by promoting vascular permeability and drawing in inflammatory cells. IL-10, an anti-inflammatory cytokine, moderates inflammation by suppressing macrophage activation and the production of anti-inflammatory cytokines such as IL-1β89. The balance between pro-inflammatory and anti-inflammatory cytokines is crucial for an effective immune response against pathogens while protecting healthy tissues from damage90. Zhang et al.42 indicated that the cytokines (IL-10, IL-1β, TNF-α, and IL-8) were upregulated significantly (P < 0.05) in rainbow trout fish that received a diet containing BSFLM meal with increasing fishmeal substitution levels of 25, 50, 75, and 100%. These results may be attributed to insect-based diets that primarily contain chitin, a molecule that has a valuable modulatory impact on the innate immunity of various fish species. For instance, the inclusion of chitin in diets based on black soldier fly larvae might stimulate the innate immune response and enhance resistance to bacterial infections42. A prolonged subclinical inflammatory response in fish resulted in consistently reduced performance and lower feed intake, as energy was diverted towards cellular defense mechanisms instead of being utilized for production. Our results indicated that TIM achieved the highest level of IL-10. Consequently, the energy and nutrients that would typically be used for inflammatory reactions could instead be allocated for productive purposes.
Optimal diets for aquaculture fish require various analytical methods to assess their health effects. Histomorphology studies serve as reliable biomarkers for assessing fish health status91. The relationship between nutritional absorption and assimilation is linked to immune function and the structural characteristics of the intestine, especially the diameter and arrangement of the microvilli92. The health of the intestinal lining cells is essential for nutrient absorption and overall fish well-being. Gut damage can result in decreased disease resistance, immune problems, loss of appetite, and stunted growth93. Histological analyses showed that BSFLM and PM were well accepted by N. tilapia. Similarly, replacing fishmeal with insect and PM meals could improve gut histomorphology in European Seabass93,94.
The liver is a vital indicator of health due to its roles in energy storage, metabolism, detoxification, and immune protection94,95. The histological findings indicated positive liver health in all fish fed various experimental diets, consistent with prior research showing that replacing fishmeal with insect meal and PM did not affect the liver histomorphology of European Seabass94. Additionally, recent research has found that incorporating BSFLM and PM into diets devoid of FM led to enhanced gut and liver health in both gilthead seabream and rainbow trout96,97,98. Also, the liver of tilapia fish remained unchanged when the protein from fishmeal was entirely substituted with the protein from poultry meal60. In our study, the histological examination of the kidney in different experimental dietary groups revealed no signs of acute or chronic inflammation in the kidney. These obtained result agreed with those reported by another study, which showed that there was no difference in the photomicrographs of rainbow trout68 fish fed insect meal-based diets. Investigators observed that the inclusion of the incorporation of Musca domestica larvae meal into the diets of tilapia did not induce any metabolic stress, as it seems to be devoid of any compounds that could generate reactive oxygen species, leading to oxidative stress99. The spleen histological structure showed no significant changes among groups. The same result was observed by Elia et al.100 who stated that the architecture of liver, spleen, and gut histological characteristics were not significantly changed by substituting 20% and 40% of BSFLM meal with 25% and 50% of FM, indicating no detrimental effects on the digestive ability of rainbow trout.
The immunostaining results for NF-κB revealed negative expression among groups, with no significant differences in staining intensity, indicating that none of the diets induced an inflammatory response. NF-κB has a central role in immune and inflammatory signaling regulation, and its activation is often associated with tissue stress or immune challenge101. Also, NF-κB is a key upstream transcription factor that regulates a broad range of immune-related genes, including both pro-inflammatory (e.g., IL-1β, IL-6, TNF-α, IL-8) and anti-inflammatory (e.g., IL-10) cytokines. NF-κB serves as a central regulator of the immune response. Its activation reflects a broader, system-level immune activation, making it a more comprehensive and integrative marker in evaluating inflammatory responses in fish102.
Therefore, the absence of detectable NF-κB activation suggests that the experimental diets, including BSFLM and PM or both, were well-tolerated and did not provoke pro-inflammatory signaling in the target tissues. This supports the immunological safety of these alternatives as replacements for traditional fishmeal in formulated diets. Such findings were aligned with previous studies reporting that BSFLM and PM do not adversely affect immune parameters when included at appropriate levels and may even support mucosal integrity and immune homeostasis103,104.
Conclusion
Black soldier fly larvae meal (TIM) has shown general potential in improving growth parameters, carcass traits, tissue quality, and immune response in Nile tilapia. This alternative not only offers comparable nutritional benefits to fishmeal but also provides functional advantages, particularly in terms of antioxidant protection, lipid stability, and product safety. These qualities support the use of BSFLM in cost-effective and sustainable aqua feed formulations. Further research focus on the long-term performance of Nile Tilapia in commercial farming conditions using BSFLM is highly recommended.
Data availability
All data are provided within the article.
References
FAO. The state of world fisheries and aquaculture. Rev. Fish. Sci. Aquac. 26, 2007 (2020).
Abozaid, H. et al. Effect of replacing dietary soybean meal with Galleria mellonella larvae powder on growth performance of the Nile tilapia (Oreochromis niloticus). Egypt. J. Aquat. Biol Fish. 28, 123–148. https://doi.org/10.21608/ejabf.2024.377638 (2024).
Khader, M. et al. Effect of replacement of fish meal by corn by product meal on growth performance for Nile tilapia (Oreochromis niloticus). Egypt. J. Vet. Sci. 56, 321–334. https://doi.org/10.21608/ejvs.2024.267728.1825 (2025).
Munguti, J. M. et al. Nile tilapia (Oreochromis niloticus Linnaeus, 1758) culture in Kenya: Emerging production technologies and socio‐economic impacts on local livelihoods. Aquac. Fish. Fish. 2, 265–276. https://doi.org/10.1002/aff2.58 (2022).
Tran, N. et al. Prospects of fish supply-demand and its implications for food and nutrition security in Egypt. Mar. Policy 146, 105333. https://doi.org/10.31235/osf.io/pbdkg (2022).
Tippayadara, N. et al. Replacement of fish meal by black soldier fly (Hermetia illucens) larvae meal: Effects on growth, haematology, and skin mucus immunity of Nile tilapia, Oreochromis niloticus. Animals 11, 193. https://doi.org/10.3390/ani11010193 (2021).
Islam, S. M. M. et al. Insect meal in aquafeeds: A sustainable path to enhanced mucosal immunity in fish. Fish Shellfish Immunol. 150, 109625. https://doi.org/10.1016/j.fsi.2024.109625 (2024).
Hussain, S. M. et al. Substitution of fishmeal: Highlights of potential plant protein sources for aquaculture sustainability. Heliyon 10, e26573. https://doi.org/10.1016/j.heliyon.2024.e26573 (2024).
Colombo, S. M. et al. Towards achieving circularity and sustainability in feeds for farmed blue foods. Rev. Aquac. 15, 1115–1141. https://doi.org/10.1111/raq.12766 (2023).
Lin, S. M. et al. Intestinal morphology, immunity and microbiota response to dietary fibers in largemouth bass, Micropterus salmoide. Fish Shellfish Immunol. 103, 135–142. https://doi.org/10.1016/j.fsi.2020.04.070 (2020).
Shapawi, R. Waste Biorefineries 179–203 (Apple Academic Press, 2024).
Luthada-Raswiswi, R., Mukaratirwa, S. & O’brien, G. Animal protein sources as a substitute for fishmeal in aquaculture diets: A systematic review and meta-analysis. Appl. Sci. 11, 3854. https://doi.org/10.3390/app11093854 (2021).
Rossi, W. Jr. & Davis, D. A. Replacement of fishmeal with poultry by-product meal in the diet of Florida pompano Trachinotus carolinus L. Aquaculture 338, 160–166 (2012).
Koch, J. F. et al. Optimizing fish meal-free commercial diets for Nile tilapia, Oreochromis niloticus. Aquaculture 452, 357–366 (2016).
Basto, A., Matos, E. & Valente, L. M. Nutritional value of different insect larvae meals as protein sources for European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 521, 735085. https://doi.org/10.1016/j.aquaculture.2020.735085 (2020).
Xiong, H. & Xu, H. Vol. 9 259 (MDPI, 2024).
Hua, K. A meta-analysis of the effects of replacing fish meals with insect meals on growth performance of fish. Aquaculture 530, 735732. https://doi.org/10.1016/j.aquaculture.2020.735732 (2021).
Kariuki, M. W. et al. Partial replacement of fishmeal with black soldier fly larvae meal in Nile tilapia diets improves performance and profitability in earthen pond. Sci. Afr. 24, e02222. https://doi.org/10.1016/j.sciaf.2024.e02222 (2024).
Mousavi, S., Zahedinezhad, S. & Loh, J. Y. A review on insect meals in aquaculture: The immunomodulatory and physiological effects. Int. Aquat. Res. 12, 100–115. https://doi.org/10.22034/iar(20).2020.1897402.1033 (2020).
Ferrer Llagostera, P., Kallas, Z., Reig, L. & de Amores Gea, D. The use of insect meal as a sustainable feeding alternative in aquaculture: Current situation, Spanish consumers’ perceptions and willingness to pay. J. Clean. Prod. 229, 10–21. https://doi.org/10.1016/j.jclepro.2019.05.012 (2019).
Sogari, G., Amato, M., Biasato, I., Chiesa, S. & Gasco, L. The potential role of insects as feed: A multi-perspective review. Animals 9, 119 (2019).
Sangsawang, A. et al. Impacts of substituting fish meal with full-fat or defatted black soldier fly (Hermetia illucens) larvae on growth, quality, and health of Nile tilapia (Oreochromis niloticus) fingerlings. Aquac. Rep. 38, 102348. https://doi.org/10.1016/j.aqrep.2024.102348 (2024).
Dietz, C. & Liebert, F. Does graded substitution of soy protein concentrate by an insect meal respond on growth and N-utilization in Nile tilapia (Oreochromis niloticus)?. Aquac. Rep. 12, 43–48. https://doi.org/10.1016/j.aqrep.2018.09.001 (2018).
AOAC. (AOAC, Washington, DC, 2002).
APHA. Standard Methods for the Examination of Water and Wastewater Vol. 6 (American Public Health Association, 1926).
Javahery, S., Nekoubin, H. & Moradlu, A. H. Effect of anaesthesia with clove oil in fish (review). Fish Physiol. Biochem. 38, 1545–1552. https://doi.org/10.1007/s10695-012-9682-5 (2012).
Ouko, K. O. et al. Effect of replacing fish meal with black soldier fly larvae meal on growth performance and economic efficiency of Nile tilapia. Fundam. Appl. Agric. 9, 1–9. https://doi.org/10.5455/faa.154509 (2024).
Osorio-Esquivel, O., Álvarez, V. B., Dorantes-Álvarez, L. & Giusti, M. M. Phenolics, betacyanins and antioxidant activity in Opuntia joconostle fruits. Food Res. Int. 44, 2160–2168. https://doi.org/10.1016/j.foodres.2011.02.011 (2011).
Tamsen, M., Shekarchizadeh, H. & Soltanizadeh, N. Evaluation of wheat flour substitution with amaranth flour on chicken nugget properties. LWT 91, 580–587. https://doi.org/10.1016/j.lwt.2018.02.001 (2018).
Botsoglou, N. A. et al. Rapid, sensitive, and specific thiobarbituric acid method for measuring lipid peroxidation in animal tissue, food, and feedstuff samples. J. Agric. Food Chem. 42, 1931–1937. https://doi.org/10.1021/jf00045a019 (1994).
Brown, B. Routine hematology procedures. In Hematology: Principles and Procedures (1988).
Doumas, B. T. Standards for total serum protein assays—A collaborative study. Clin. Chem. 21, 1159–1166. https://doi.org/10.1093/clinchem/21.8.1159 (1975).
Heinegård, D. & Tiderström, G. Determination of serum creatinine by a direct colorimetric method. Clin. Chim. Acta 43, 305–310. https://doi.org/10.1016/0009-8981(73)90466-x (1973).
Aref, S., Habiba, R., Morsy, N., Abdel-Daim, M. & Zayet, F. Improvement of the shelf life of grey mullet (Mugil cephalus) fish steaks using edible coatings containing chitosan, nanochitosan, and clove oil during refrigerated storage. Food Prod. Process. Nutr. 4, 27. https://doi.org/10.1186/s43014-022-00106-z (2022).
Suvarna, K. S., Layton, C. & Bancroft, J. D. Bancroft’s Theory and Practice of Histological Techniques (Elsevier Health Sciences, 2018).
Sallam, E. A. et al. Replacing fish meal with rapeseed meal: Potential impact on the growth performance, profitability measures, serum biomarkers, antioxidant status, intestinal morphometric analysis, and water quality of Oreochromis niloticus and Sarotherodon galilaeus fingerlings. Vet. Res. Commun. 45, 223–241. https://doi.org/10.1007/s11259-021-09803-5 (2021).
Duncan, D. B. Multiple range and multiple F tests. Biometrics 11, 1. https://doi.org/10.2307/3001478 (1955).
Devic, E., Leschen, W., Murray, F. & Little, D. C. Growth performance, feed utilization and body composition of advanced nursing Nile tilapia (Oreochromis niloticus) fed diets containing black soldier fly (Hermetia illucens) larvae meal. Aquac. Nutr. 24, 416–423. https://doi.org/10.1111/anu.12573 (2018).
Nairuti, R. N., Munguti, J. M., Waidbacher, H. & Zollitsch, W. Growth performance and survival rates of Nile tilapia (L.) reared on diets containing black soldier fly (L.) larvae meal. Die Bodenkultur J. Land Manag. Food Environ. 72, 9–19. https://doi.org/10.2478/boku-2021-0002 (2021).
Muin, H., Taufek, N., Kamarudin, M. & Razak, S. Growth performance, feed utilization and body composition of Nile tilapia, Oreochromis niloticus (Linnaeus, 1758) fed with different levels of black soldier fly, Hermetia illucens (Linnaeus, 1758) maggot meal diet. Iran. J. Fish. Sci. 16, 567–577 (2017).
Wachira, M. N. et al. Efficiency and improved profitability of insect-based aquafeeds for farming Nile tilapia fish (Oreochromis niloticus L.). Animals 11, 2599. https://doi.org/10.3390/ani11092599 (2021).
Zhang, Z. et al. Effect of full-fat black soldier fly (Hermetia illucens L.) larvae on growth performance, immunological parameters, and gene expressions in zebrafish (Danio rerio). Int. Aquat. Res. 16, 55–69. https://doi.org/10.22034/iar.2024.2003652.1570 (2024).
Liland, N. S. et al. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 12, e0183188. https://doi.org/10.1371/journal.pone.0183188 (2017).
Shumo, M. et al. The nutritive value of black soldier fly larvae reared on common organic waste streams in Kenya. Sci. Rep. 9, 10110. https://doi.org/10.1038/s41598-019-46603-z (2019).
Priyadarshana, M. K. C., Walpita, C. N., Ruwandeepika, H. A. D. & Magamage, M. P. S. Effects of black soldier fly, Hermetia illucens (Linnaeus, 1758), larvae incorporated feed on histomorphology, gut microbiota and blood chemistry of cultured fishes: A review. Int. J. Fish. Aquac. 35, 269–281. https://doi.org/10.33997/j.afs.2022.35.3.005 (2022).
Dietz, C. & Liebert, F. Does graded substitution of soy protein concentrate by an insect meal respond on growth and N-utilization in Nile tilapia (Oreochromis niloticus)?. Aquac. Rep. 12, 43–48 (2018).
Fayed, W. M. et al. Water quality change, growth performance, health status in response to dietary inclusion of black soldier fly larvae meal in the diet of Nile tilapia, Oreochromis niloticus. Ann. Anim. Sci. 24, 533–544. https://doi.org/10.2478/aoas-2023-0088 (2024).
Guerreiro, I. et al. Oxidative stress response of meagre to dietary black soldier fly meal. Animals (Basel) 12, 3232. https://doi.org/10.3390/ani12233232 (2022).
Giannetto, A. et al. Hermetia illucens (Diptera: Stratiomydae) larvae and prepupae: Biomass production, fatty acid profile and expression of key genes involved in lipid metabolism. J. Biotechnol. 307, 44–54. https://doi.org/10.1016/j.jbiotec.2019.10.015 (2020).
Fontes, T. V. et al. Digestibility of insect meals for Nile tilapia fingerlings. Animals (Basel) 9, 181. https://doi.org/10.3390/ani9040181 (2019).
Nawaz, A., Irshad, S., Hoseinifar, S. H. & Xiong, H. The functionality of prebiotics as immunostimulant: Evidences from trials on terrestrial and aquatic animals. Fish Shellfish Immunol. 76, 272–278. https://doi.org/10.1016/j.fsi.2018.03.004 (2018).
Limbu, S. M. et al. Black soldier fly (Hermetia illucens L.) larvae meal improves growth performance, feed efficiency and economic returns of Nile tilapia (Oreochromis niloticus L.) fry. Aquaculture 2, 167–178. https://doi.org/10.3153/ar22023 (2022).
Rana, K. S., Salam, M., Hashem, S. & Islam, M. A. Development of black soldier fly larvae production technique as an alternate fish feed. Int. J. Fish. Aquac. 5, 41–47 (2015).
Bhatt, D. & Pandey, A. Potential novel feed ingredients in aquaculture for future feed: A REVIEW. J. Exp. Zool. India https://doi.org/10.51470/jez.2024.27.1.63 (2024).
Galkanda‐Arachchige, H. S., Wilson, A. E. & Davis, D. A. Success of fishmeal replacement through poultry by‐product meal in aquaculture feed formulations: A meta‐analysis. Rev. Aquac. 12, 1624–1636. https://doi.org/10.1111/raq.12401 (2020).
Abdel-Tawwab, M. et al. Effects of black soldier fly (Hermetia illucens L.) larvae meal on growth performance, organs-somatic indices, body composition, and hemato-biochemical variables of European sea bass, Dicentrarchus labrax. Aquaculture 522, 735136. https://doi.org/10.1016/j.aquaculture.2020.735136 (2020).
Ushakova, N. et al. Biological efficiency of the prepupae Hermetia illucens in the diet of the young Mozambique Tilapia Oreochromis mossambicus. Biol. Bull. 45, 382–387. https://doi.org/10.1134/s1062359018040143 (2018).
Kari, N., Ahmad, F. & Ayub, M. Proximate composition, amino acid composition and food product application of anchovy: A review. Food Res. 6, 16–29. https://doi.org/10.26656/fr.2017.6(4).419 (2022).
Venugopal, V. & Shahidi, F. Structure and composition of fish muscle. Food Rev. Int. 12, 175–197. https://doi.org/10.1080/87559129609541074 (1996).
Mahmoud, R. E., Gadallah, H. & Orma, O. A. Effects of replacing protein of fishmeal with protein of poultry by-product meal on growth performance, body composition, liver histological changes and selected serum parameters of Nile tilapia. J. Adv. Vet. Res. 13, 871–876 (2023).
Peña-Saldarriaga, L. M., Fernández-López, J. & Pérez-Alvarez, J. A. Quality of chicken fat by-products: Lipid profile and colour properties. Foods 9, 1046. https://doi.org/10.3390/foods9081046 (2020).
Siddaiah, G. et al. Dietary fishmeal replacement with Hermetia illucens (Black soldier fly, BSF) larvae meal affected production performance, whole body composition, antioxidant status, and health of snakehead (Channa striata) juveniles. Anim. Feed Sci. Technol. 297, 115597. https://doi.org/10.1016/j.anifeedsci.2023.115597 (2023).
Teye-Gaga, C. Evaluation of Larval Meal Diet of Black Soldier Fly (Hermetia illucens: L. 175) On Fingerlings Culture of Nile Tilapia (Oreochromis Niloticus: L.) (2017).
Hernández, C. et al. Complete replacement of fish meal by porcine and poultry by-product meals in practical diets for fingerling Nile tilapia Oreochromis niloticus: Digestibility and growth performance. Aquac. Nutr. 16, 44–53. https://doi.org/10.1111/j.1365-2095.2008.00639.x (2010).
Huang, J., Yu, T., Yuan, B., Xiao, J. & Huang, D. The addition of Hermetia illucens to feed: Influence on nutritional composition, protein digestion characteristics, and antioxidant activity of Acheta domesticus. Foods 14, 1140. https://doi.org/10.3390/foods14071140 (2025).
Mohan, K. et al. Use of black soldier fly (Hermetia illucens L.) larvae meal in aquafeeds for a sustainable aquaculture industry: A review of past and future needs. Aquaculture 553, 738095. https://doi.org/10.1016/j.aquaculture.2022.738095 (2022).
Danfær, A. Nutrient metabolism and utilization in the liver. Livest. Prod. Sci. 39, 115–127. https://doi.org/10.1016/0301-6226(94)90163-5 (1994).
Renna, M. et al. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens L.) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss Walbaum) diets. J. Anim. Sci. Biotechnol. 8, 1–13. https://doi.org/10.1186/s40104-017-0191-3 (2017).
Dawood, M. A., Eweedah, N. M., Khalafalla, M. M. & Khalid, A. Evaluation of fermented date palm seed meal with Aspergillus oryzae on the growth, digestion capacity and immune response of Nile tilapia (Oreochromis niloticus). Aquac. Nutr. 26, 828–841. https://doi.org/10.1111/anu.13042 (2020).
Yildirim-Aksoy, M., Eljack, R., Schrimsher, C. & Beck, B. H. Use of dietary frass from black soldier fly larvae, Hermetia illucens, in hybrid tilapia (Nile x Mozambique, Oreocromis niloticus x O. mozambique) diets improves growth and resistance to bacterial diseases. Aquac. Rep. 17, 100373. https://doi.org/10.1016/j.aqrep.2020.100373 (2020).
Amer, A. A., El‐Nabawy, E. S. M., Gouda, A. H. & Dawood, M. A. The addition of insect meal from Spodoptera littoralis in the diets of Nile tilapia and its effect on growth rates, digestive enzyme activity and health status. Aquac. Res. 52, 5585–5594. https://doi.org/10.1111/are.15434 (2021).
Karapanagiotidis, I. T., Psofakis, P., Mente, E., Malandrakis, E. & Golomazou, E. Effect of fishmeal replacement by poultry by‐product meal on growth performance, proximate composition, digestive enzyme activity, haematological parameters and gene expression of gilthead seabream (Sparus aurata). Aquac. Nutr. 25, 3–14. https://doi.org/10.1111/anu.12824 (2019).
Docan, A., Grecu, I. & Dediu, L. Use of hematological parameters as assessment tools in fish health status. J. Agroaliment. Process. Technol. 24, 317–324 (2018).
Mauel, M. J., Miller, D. L. & Merrill, A. L. Hematologic and plasma biochemical values of healthy hybrid tilapia (Oreochromis aureus× Oreochromis nilotica) maintained in a recirculating system. J. Zoo Wildl. Med. 38, 420–424. https://doi.org/10.1638/06-025.1 (2007).
Chotolli, A. P. et al. Dietary fruit by-products improve the physiological status of Nile tilapias (Oreochromis niloticus) and the quality of their meat. Antioxidants 12, 1607. https://doi.org/10.3390/antiox12081607 (2023).
Liang, H. et al. Effects of dietary calcium levels on growth performance, blood biochemistry and whole body composition in juvenile bighead carp (Aristichthys nobilis). Turk. J. Fish. Aquat. Sci. 18, 623–631. https://doi.org/10.46989/001c.21646 (2018).
Oliveira, CGd. et al. Impact of replacing fish meal with black soldier fly (Hermetia illucens) meal on diet acceptability in juvenile Nile tilapia: Palatability and nutritional and health considerations for dietary preference. Aquac. Res. 2024, 3409955. https://doi.org/10.1155/2024/3409955 (2024).
Zhou, J., Liu, S., Ji, H. & Yu, H. Effect of replacing dietary fish meal with black soldier fly larvae meal on growth and fatty acid composition of Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 24, 424–433. https://doi.org/10.1111/anu.12574 (2018).
Li, S., Ji, H., Zhang, B., Zhou, J. & Yu, H. Defatted black soldier fly (Hermetia illucens) larvae meal in diets for juvenile Jian carp (Cyprinus carpio var. Jian): Growth performance, antioxidant enzyme activities, digestive enzyme activities, intestine and hepatopancreas histological structure. Aquaculture 477, 62–70. https://doi.org/10.1016/j.aquaculture.2017.04.015 (2017).
Elbialy, Z. I. et al. Yucca schidigera extract mediated the growth performance, hepato‐renal function, antioxidative status and histopathological alterations in Nile tilapia (Oreochromis niloticus) exposed to hypoxia stress. Aquac. Res. 52, 1965–1976. https://doi.org/10.1111/are.15045 (2021).
Abdullahi, N. A., Basir, Z., Peyghan, R. & Fatemi-Tabatabaei, S. R. Effect of fishmeal replacement with poultry by-product meal on serum parameters and histomorphology of liver and kidney in Nile tilapia ((Oreochromis niloticus), Linnaeus, 1758). Iran. Vet. J. 20, 5–19. https://doi.org/10.22055/ivj.2024.408390.2599 (2024).
Lin, S. & Luo, L. Effects of different levels of soybean meal inclusion in replacement for fish meal on growth, digestive enzymes and transaminase activities in practical diets for juvenile tilapia, Oreochromis niloticus× O. aureus. Anim. Feed Sci. Technol. 168, 80–87. https://doi.org/10.1016/j.anifeedsci.2011.03.012 (2011).
Rurangwa, E. & Verdegem, M. C. Microorganisms in recirculating aquaculture systems and their management. Rev. Aquac. 7, 117–130. https://doi.org/10.1111/raq.12057 (2015).
ICMSF. International Commission on Microbiological Specifications for Foods Vol. 6 (Springer, 2006).
Stenberg, O. K. et al. Effect of dietary replacement of fish meal with insect meal on in vitro bacterial and viral induced gene response in Atlantic salmon (Salmo salar) head kidney leukocytes. Fish Shellfish Immunol. 91, 223–232. https://doi.org/10.1016/j.fsi.2019.05.042 (2019).
Choi, W. H., Yun, J. H., Chu, J. P. & Chu, K. B. Antibacterial effect of extracts of Hermetia illucens (Diptera: Stratiomyidae) larvae against Gram‐negative bacteria. Entomol. Res. 42, 219–226. https://doi.org/10.1111/j.1748-5967.2012.00465.x (2012).
Sanjee, S. A. & Karim, M. E. Microbiological quality assessment of frozen fish and fish processing materials from Bangladesh. Int. J. Food Sci. 2016, 8605689. https://doi.org/10.1155/2016/8605689 (2016).
Rimoldi, S., Antonini, M., Gasco, L., Moroni, F. & Terova, G. Intestinal microbial communities of rainbow trout (Oncorhynchus mykiss) may be improved by feeding a Hermetia illucens meal/low-fishmeal diet. Fish Physiol. Biochem. 47, 365–380. https://doi.org/10.1007/s10695-020-00918-1 (2021).
Arango Duque, G. & Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 5, 491. https://doi.org/10.3389/fimmu.2014.00491 (2014).
Al-Qahtani, A. A., Alhamlan, F. S. & Al-Qahtani, A. A. Pro-inflammatory and anti-inflammatory interleukins in infectious diseases: A comprehensive review. Trop. Med. Infect. Dis. 9, 13. https://doi.org/10.3390/tropicalmed9010013 (2024).
De Marco, G., Cappello, T. & Maisano, M. Histomorphological changes in fish gut in response to prebiotics and probiotics treatment to improve their health status: A review. Animals 13, 2860. https://doi.org/10.3390/ani13182860 (2023).
Kord, M. I. et al. Impacts of water additives on water quality, production efficiency, intestinal morphology, gut microbiota, and immunological responses of Nile tilapia fingerlings under a zero-water-exchange system. Aquaculture 547, 737503. https://doi.org/10.1016/j.aquaculture.2021.737503 (2022).
Pleić, I. L. et al. A plant-based diet supplemented with Hermetia illucens alone or in combination with poultry by-product meal: One step closer to sustainable aquafeeds for European seabass. J. Anim. Sci. Biotechnol. 13, 77. https://doi.org/10.1186/s40104-022-00725-z (2022).
Rimoldi, S. et al. The replacement of fish meal with poultry by-product meal and insect exuviae: Effects on growth performance, gut health and microbiota of the European seabass, Dicentrarchus labrax. Microorganisms 12, 744. https://doi.org/10.3390/microorganisms12040744 (2024).
Bruslé, J. & Anadon, G. G. Fish Morph 77–93 (Routledge, 2017).
Randazzo, B. et al. Physiological response of rainbow trout (Oncorhynchus mykiss) to graded levels of Hermetia illucens or poultry by-product meals as single or combined substitute ingredients to dietary plant proteins. Aquaculture 538, 736550. https://doi.org/10.1016/j.aquaculture.2021.736550 (2021).
Randazzo, B. et al. (s Note: MDPI stays neutral with regard to jurisdictional claims in published …, 2021).
Donadelli, V. et al. Effects of dietary plant protein replacement with insect and poultry by-product meals on the liver health and serum metabolites of sea bream (Sparus aurata) and sea bass (Dicentrarchus labrax). Animals 14, 241. https://doi.org/10.3390/ani14020241 (2024).
Ogunji, J. O., Nimptsch, J., Wiegand, C. & Schulz, C. Evaluation of the influence of housefly maggot meal (magmeal) diets on catalase, glutathione S-transferase and glycogen concentration in the liver of Oreochromis niloticus fingerling. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 147, 942–947. https://doi.org/10.1016/j.cbpa.2007.02.028 (2007).
Elia, A. C. et al. Influence of Hermetia illucens meal dietary inclusion on the histological traits, gut mucin composition and the oxidative stress biomarkers in rainbow trout (Oncorhynchus mykiss). Aquaculture 496, 50–57. https://doi.org/10.1016/j.aquaculture.2018.07.009 (2018).
Li, Q. & Verma, I. M. NF-κB regulation in the immune system. Nat. Rev. Immunol. 2, 725–734. https://doi.org/10.1038/nri910 (2002).
Chu, Q. & Xu, T. MicroRNA regulation of Toll-like receptor, RIG-I-like receptor and Nod-like receptor pathways in teleost fish. Rev. Aquac. 12, 2177–2193 (2020).
Sayramoğlu, H. et al. Effects of black soldier fly meal feeding on rainbow trout gut microbiota, immune-related gene expression, and Lactococcus petauri resistance. J. Insects Food Feed. https://doi.org/10.1163/23524588-20230057 (2023).
Yones, A. & Metwalli, A. Effects of fish meal substitution with poultry by-product meal on growth performance, nutrients utilization and blood contents of juvenile Nile tilapia (Oreochromis niloticus). J. Aquac. Res. Dev. 7, 1000389. https://doi.org/10.4172/2155-9546.1000389 (2015).
Acknowledgements
The authors would like to acknowledge the Deanship of Graduate Studies and Scientific Research, Taif University, Kingdom of Saudi Arabia for funding this work. This research was funded by the Hungarian National Research, Development, and Innovation Office, grant number TKP2021-NVA-22. This work was also supported by the Flagship Research Groups Programme of the Hungarian University of Agriculture and Life Sciences.
Funding
Open access funding provided by Hungarian University of Agriculture and Life Sciences. The work was funded by the Deanship of Graduate Studies and Scientific Research, Taif University, Kingdom of Saudi Arabia.
Author information
Authors and Affiliations
Contributions
All authors contributed to the conception and design of the research. Heba Alian led the preparation of the initial draft of the manuscript and provided methodological support for the experimental procedures. Samar Aref, Fatma Khodary, András Székács, Omar Saeed, Mohamed Hamdy Eid, Abdallah Elshawadfy Elwakeel, M. Alhumedi, Atef Fathy Ahmed, Tamer Moussa Ayoub, and Mohamed Salem were responsible for the collection and analysis of data, along with statistical assessments, with assistance from Heba Alian. Each author carefully reviewed and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Aref, S.M., Alian, H.A., Khodary, F.M. et al. Fish meal replacement with poultry byproduct and black soldier fly larvae proteins: effects on growth, flesh quality, bioactivity, and physiological responses of Nile tilapia. Sci Rep 16, 9536 (2026). https://doi.org/10.1038/s41598-026-43600-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-43600-x
