
Abstract
Tomatoes cultivated in facility agriculture often encounter low-light stress in autumn, winter, and early spring. This study aimed to investigate the effects of supplementary lighting with different spectral compositions dominated by red-blue (RB) light on plant growth, fruit development, and quality formation of early spring facility-grown tomatoes. Six supplementary lighting treatments were set up: red-blue (RB), red-blue + red (RB + R), red-blue + blue (RB + B), red-blue + green (RB + G), red-blue + far-red (RB + FR), and red-blue + ultraviolet A (RB + UVA). The treatment with no supplementary lighting served as the control (CK), and supplementary lighting was provided from 06:00 to 08:30 and from 18:30 to 22:00 daily. The results showed that, compared with CK, supplementary lighting with different spectral compositions significantly promoted plant growth, accelerated fruit ripening, and improved yield and quality (P < 0.05). Compared with the RB treatment, the RB + R treatment significantly increased the number of large fruits and yield per plant by 25.79% and 6.94%, respectively, demonstrating a superior effect on promoting yield formation over other treatments; the RB + B treatment was beneficial for increasing stem diameter and leaf SPAD value, and the contents of vitamin C and soluble protein were significantly higher than those in the RB treatment during the late stage of fruit development (fruit color-breaking stage and mature stage); the RB + G treatment had the highest average fruit weight, which was significantly increased by 8.29% compared with the RB treatment; during the late stage of fruit development, the contents of lycopene, soluble protein, amino acids, and total phenols in the RB + G treatment were significantly higher than those in the RB treatment, and the soluble sugar content and sugar-acid ratio was the highest at the fruit mature stage; the RB + FR treatment significantly increased the contents of soluble protein, soluble sugar, amino acids, and the sugar-acid ratio during the late stage of fruit development, and the fruit color parameters (a* and b* values) were significantly higher than those of the RB treatment; the RB + UVA treatment stimulated plant height growth in the short term and significantly increased the contents of soluble protein, soluble sugar, and organic acids during the late stage of fruit development. The results of this study provide a theoretical basis and technical reference for the regulation of the light environment in facility agriculture and the efficient production of facility-grown tomatoes.
Similar content being viewed by others

Introduction
Facility agriculture is an agricultural production method characterized by high yield, efficiency, and added value. When adverse conditions such as low temperature and weak light occur in the open field environment, facility agriculture can maintain normal crop growth by artificially regulating environmental factors such as light, temperature, water, air, and fertilizer. However, due to the enclosed structure inside the facilities, the lighting time is often short, the light intensity is weak, and the light distribution is uneven. Such a light environment is often unsuitable for the growth and development of sun-loving crops such as tomatoes. In Guizhou Province, overcast and rainy weather is frequent during the winter and spring seasons, with low sunshine duration. According to statistics from the Guizhou Provincial Meteorological Bureau, the national average annual sunshine duration is about 2107 h, and the average sunshine duration in winter and spring is about 428 h and 558 h, respectively. From 2020 to 2025, the average annual sunshine duration in Guizhou Province was 1377.4 h, and the average sunshine duration in winter and spring was 185.5 h and 337.4 h, respectively, which is far lower than the national average. Weak light and short sunshine duration have become key factors limiting the high-quality and efficient production of local facility agriculture. Artificial light supplementation can ameliorate the problem of insufficient light inside the facilities under short sunshine duration, which is of great significance for enhancing the productivity and quality of facility agriculture.
Red light and blue light are the two primary spectra involved in plant photosynthesis. Red light can promote carbohydrate accumulation, induce the biosynthesis of ethylene and abscisic acid, and accelerate tomato fruit ripening1. Red light has a significant effect on plant morphology and biomass accumulation in tomatoes2. Supplementary red or blue light at night can significantly increase the content of lycopene, β-carotene, fructose, and glucose in tomatoes and accelerate fruit ripening3. Properly increasing the proportion of blue light (R: B = 1:0.8) can induce the upregulation of genes related to photosynthesis and carotenoid synthesis in tomatoes, thereby improving light energy capture efficiency and carotenoid content, and promoting fruit development4. Long-term exposure to monochromatic red light can decrease the photosynthetic capacity of plants (red light syndrome), and the addition of blue light can eliminate this adverse effect5. Blue light can increase chlorophyll content, promote the development of chloroplasts, palisade tissue, and spongy tissue, induce stomatal opening, and protect the photosynthetic system, thereby facilitating the improvement of plant photosynthetic capacity6,7. However, an excessively low intensity of blue light can trigger a shade avoidance response8. Green light can reverse the stomatal opening induced by blue light, trigger the shade avoidance response in plants, and maintain circadian rhythms9,10,11. Adding green light to red-blue light can promote biomass accumulation and yield formation in vegetable crops such as tomatoes and peppers. Due to plants’ low absorption of green light, most green light radiation is reflected or transmitted through the leaves, increasing the light intensity within the plant canopy, thereby improving the photosynthetic capacity at the bottom of the canopy and regulating carbohydrate distribution, promoting biomass accumulation12,13. A low red/far-red ratio can trigger a shade avoidance response, manifested as increased petiole/leaf length, and this effect is further enhanced by green light14. Research has shown that the addition of far-red light promotes biomass accumulation and internode elongation in tomato stems and leaves, thereby increasing yield15. Far-red light and ultraviolet A (UVA) have significant effects on strengthening cucumber and tomato seedlings16. Studies have indicated that UVA can promote the development of photosynthetic organs and exhibit functions similar to blue light in plant photosynthesis. Adding a low dose of UVA to red light or red-blue light can stimulate the growth of tomatoes and lettuce, promote the elongation of tomato stems and the expansion of leaf area, increase leaf light interception, and facilitate the accumulation of biomass, anthocyanins, vitamin C (VC), and soluble sugars17,18,19.
To explore the effects of supplementary lighting with different spectral compositions on the growth, yield, quality, and fruit ripening of facility-grown tomato plants under short-day conditions, this study, based on red-blue (1R:1B) light, added red, blue, green, far-red, and ultraviolet A light with different light intensities. It systematically analyzed the effects of the different spectral supplementary lighting treatments on tomato growth, yield components, fruit ripening dynamics, and quality formation. The aim is to provide a theoretical basis for the regulation of the facility light environment and efficient tomato production.
Materials and methods
Test materials and plant growth conditions
The tomato cultivar used in this experiment was ‘Provence’. The supplementary lighting fixtures used were LED (light-emitting diode) tubes (105.0 × 5.2 cm), capable of emitting red light (R, 660 nm), blue light (B, 450 nm), green light (G, 525 nm), far-red light (FR, 730 nm), and ultraviolet A light (UVA, 395 nm), with adjustable light quality and intensity. The fixtures were provided by Qiming Ditong Technology (Beijing) Co., Ltd.
The experiment was conducted in a plastic greenhouse at the Guizhou Academy of Agricultural Sciences from March to June 2025. Tomato seeds were soaked in warm water at 55 °C for 15 min, germinated at room temperature for 24 h, and then sown in 72-hole seedling trays. When the seedlings had developed 5 leaves and 1 bud (March 3rd), uniform seedlings were transplanted into pots (31 cm diameter, 21 cm height) filled with a substrate (peat: vermiculite: perlite = 3:1:1, v/v/v). Each pot was supplemented with 1 kg of organic fertilizer made from cow and sheep manure. Plants were irrigated with water before flowering and with a tomato soilless cultivation AB water-soluble fertilizer after flowering. Side buds and lower old leaves were promptly removed, and the plants were topped when they reached 5 fruit clusters. Except for the supplementary lighting treatment, all other cultivation management measures were identical across treatments. Each treatment included 3 biological replicates, arranged in a completely randomized design.
From plant transplanting to the end of the experiment, the maximum day/night temperature in the greenhouse was monitored as 43.0/27.5 °C, the average day/night temperature was 22.0/15.9 °C, and the minimum day/night temperature was 3.6/3.4 °C. The maximum day/night humidity was 99.0/99.0%, with an average day/night humidity of 72.1/87.6%, and the minimum day/night humidity was 15.0/27.0%. The highest, average, and lowest light intensities during the day (photosynthetic photon flux density, PPFD / photon flux density, PFD) were 1052.1/1546.8, 196.7/288.0, and 0.2/0.5 µmol·m−2·s−1, respectively. From fruit setting to sampling, the maximum day/night temperature in the greenhouse was 36.5/27.5 °C, the average day/night temperature was 22.6/17.7 °C, and the minimum day/night temperature was 8.8/8.4 °C (monitored by the Xiaomengyu device provided by Xiaoyu (Beijing) Technology Co., Ltd.). PPFD refers to the total number of photons with wavelengths between 400 and 700 nm incident on a unit area per unit time, expressed in µmol·m− 2·s− 1. PFD refers to the total number of photons within a specified spectral range incident on a unit area per unit time; the range most commonly used in botany is 350–800 nm, expressed in µmol·m− 2·s− 1.
Light treatments
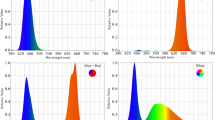
The experiment included a control with no supplementary lighting (CK) and six supplementary lighting treatments with different spectral compositions(Table 1): RB (100 µmol·m− 2·s− 1 R and 100 µmol·m− 2·s− 1 B), RB + R (RB + 100 µmol·m− 2·s− 1 R), RB + B (RB + 100 µmol·m− 2·s− 1 B), RB + G (RB + 50 µmol·m− 2·s− 1 G), RB + FR (RB + 30 µmol·m− 2·s− 1 FR), and RB + UVA (RB + 10 µmol·m− 2·s− 1 UVA). Supplementary lighting began at the bud expansion stage. To maintain a 16 h light / 8 h dark photoperiod, lights were turned on from 06:00 to 08:30 and from 18:30 to 22:00 daily, providing a total of 6 h of supplementary lighting per day. The LED light sources were positioned approximately 25 cm above the plant growing points. The distance was adjusted timely according to plant growth (when the distance reached 25–30 cm, it was adjusted back to 20–25 cm) to ensure that the distance from the plant growing points to the light source was consistently maintained at 20–30 cm. The spectrum measured 25 cm below the light source (at the height of the plant growing points) using a PG200N handheld spectroradiometer (Upoptics Co., Ltd.) is shown in Fig. 1, and a photograph of the lighting setup is shown in Fig. 2.

Spectra of LED (light-emitting diode) measured 25 cm below the light source for the six supplementary lighting treatments: RB, RB + R, RB + B, RB + G, RB + FR, and RB + UVA. PFD-R, PFD-B, PFD-G, PFD-FR, PFD-UA, and PFD-Y denote the photon flux densityfor red (600–700 nm), blue (400–500 nm), green (500–600 nm), far-red (700–800 nm), ultraviolet A (350–400 nm), and the total photon flux density, respectively.

Photographs of tomato plants under different supplementary lighting treatments: CK (no supplementary lighting), RB, RB + R, RB + B, RB + G, RB + FR, and RB + UVA.
Plant morphology and SPAD measurement
After 14 and 42 days of supplementary lighting (before topping), 12 tomato plants were randomly selected (4 plants per replicate, 3 biological replicates) from each treatment for morphological measurements. Plant height was determined by measuring the distance from the cotyledon node to the growing point with a tape measure. Stem diameter was measured at the cotyledon node with a digital caliper. Leaf length and leaf width of the fourth fully expanded leaf were measured using a ruler. SPAD values of the fourth fully expanded leaf were measured using a handheld chlorophyll meter.
Recording of fruit ripening dates
Starting from the onset of fruit color-breaking, the following stages were recorded for each treatment: First fruit color-breaking, first fruit mature, half plants color-breaking, half plants mature, all plants color-breaking, and all plants mature.
Yield determination
Eighteen tomato plants (6 plants per replicate, 3 biological replicates) per treatment were harvested for color-breaking and mature fruits. The total fruit number and total fresh weight were recorded to calculate the average fruit weight and yield per plant. During the fruit expansion stage, color-breaking stage, and mature stage, the number of large fruits (transverse diameter > 70.0 mm), number of medium fruits (50.0 mm < transverse diameter ≤ 70.0 mm), and number of small fruits (transverse diameter ≤ 50.0 mm) per plant were counted.
Determination of nutrient contents
Biochemical reagent kits provided by Suzhou Greathness Biotechnology Co., Ltd. were used to determine nutrient contents. The specific operating procedures are detailed in the kit manuals. During the fruit expansion stage, color-breaking stage, and mature stage, six fruits were sampled (two per replicate) from each treatment for the determination of lycopene, vitamin C, soluble protein, soluble sugar, amino acids, and total phenols. The determination methods employed were petroleum ether extraction for lycopene, the phenanthroline method for vitamin C, the coomassie brilliant blue method for soluble protein, the anthrone colorimetric method for soluble sugar, the ninhydrin colorimetric method for amino acids, and the folin-ciocalteu method for total phenols. Organic acids were determined using acid-base titration. The sugar-acid ratio was calculated as soluble sugar content divided by organic acid content.
Determination of appearance quality
During the fruit mature stage, nine fruits were collected (3 fruits per replicate, 3 biological replicates) per treatment. Color parameters (L, a*, b*) and fruit firmness were measured at three equidistant points on the equatorial part of each tomato fruit using an NF555 colorimeter and a GY-4 fruit firmness tester, respectively. The measurements were averaged for each fruit.
Statistical analysis
All data are presented as mean ± standard error. Data were organized using excel software, and SPSS 26 software was used for significance testing (P < 0.05). Post hoc multiple comparisons were performed using the LSD test (assuming equal variances) and Tamhane’s T2 test (not assuming equal variances). Origin 2022 software was used for plotting.
Results and analysis
Effects of supplementary lighting with different spectral compositions on tomato plant growth
Supplementary lighting significantly promoted plant growth (Table 2). After 14 days of lighting, the RB treatment significantly increased plant stem diameter and leaf SPAD values (P < 0.05), with no significant differences in other morphological traits compared with CK. After 42 days of lighting, plant height and leaf count increased more rapidly. Plant height, stem diameter, leaf count, and SPAD values in the RB treatment were significantly greater than those in CK. Additionally, leaf count in the RB treatment was significantly higher than that in the RB + G, RB + FR, and RB + UVA treatments, reaching the maximum value among all treatments. The RB + R treatment resulted in significantly greater stem diameter and SPAD values than CK after 14 days of lighting, a trend similar to that of the RB treatment. After 42 days of lighting, plant height, leaf count, leaf length, and leaf length × leaf width in the RB + R treatment increased significantly faster than in CK, with leaf count and leaf length significantly greater than those in the RB + FR and RB + UVA treatments. The RB + B treatment significantly increased stem diameter after 14 days of lighting. After 42 days of lighting, plant height and leaf SPAD values increased significantly, with SPAD values significantly higher than those of other treatments. The growth trend of plants in the RB + B treatment was similar to that in the RB + G treatment. The RB + FR treatment resulted in significantly greater stem diameter and SPAD values than CK after 14 days of lighting, similar to the RB and RB + R treatments. After 42 days of lighting, the leaf length growth rate accelerated, becoming significantly higher than that of CK. The RB + UVA treatment resulted in significantly higher plant height, stem diameter, and leaf count than CK after 14 days of lighting. After 42 days of lighting, SPAD values increased significantly, but leaf count growth decreased.
Effects of supplementary lighting with different spectral compositions on tomato fruit ripening
Supplementary lighting significantly promoted fruit ripening (Fig. 3A). After 60 days of supplementary lighting, color-breaking fruits appeared in the RB + B treatment. After 65 days, all supplementary lighting treatments had at least two fruits at the color-breaking or mature stage, with two mature fruits in the RB + B and RB + FR treatments, respectively. After 69 days, two mature fruits appeared in the CK and RB + UVA treatments, respectively, while all three labeled fruits in the other treatments were fully ripe.

Fruit ripening process recorded photographically at 45, 60, 65, and 69 days after the initiation of supplementary lighting. For each treatment, three fruits at the same position on three different plants were selected and marked during the fruit expansion stage. “1” indicates color-breaking fruits, and “2” indicates mature fruits.
As shown in Table 3, compared with CK, the RB, RB + R, RB + B, RB + G, RB + FR, and RB + UVA treatments advanced fruit color-breaking by 1–3 d, 0–2 d, 1–4 d, 2–3 d, 2–3 d, and 2–6 d, respectively, and advanced fruit ripening by 0–3 d, 0–4 d, 1–4 d, 1–3 d, 2–4 d, and 2–5 d, respectively.
Effects of supplementary lighting with different spectral compositions on tomato fruit expansion and yield formation
Supplementary lighting promoted tomato fruit expansion and increased yield (Fig. 4A). The yield per plant under each supplementary lighting treatment was significantly higher than that of CK, ranging from 8.22% to 16.10% higher (P < 0.05). The RB + R and RB + G treatments significantly increased yield by 6.94% and 6.31%, respectively, compared with the RB treatment. The average fruit weight in the RB + G treatment was the highest (Fig. 4B), significantly increasing by 13.05% and 8.29% compared with CK and the RB treatment, respectively. The next highest were the RB + FR and RB + R treatments, which significantly increased average fruit weight by 11.24% and 7.58% compared with CK, respectively.

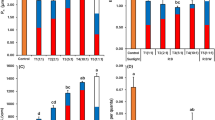
Yield per plant (A) and average fruit weight (B) of tomato under different supplementary lighting treatments. The average total fruit number per plant (C), number of large fruits per plant (D), number of medium fruits per plant (E), and number of small fruits per plant (F) were recorded at the fruit expansion, color-breaking, and mature stages. Large fruits: transverse diameter > 70.0 mm; medium fruits: 50.0 mm < transverse diameter ≤ 70.0 mm; small fruits: transverse diameter ≤ 50.0 mm. No large fruits were observed in the RB + FR and RB + UVA treatments at the fruit expansion stage. Data are presented as mean ± standard error (n = 3 biological replicates). Different letters indicate significant differences at P < 0.05.
The period from the fruit expansion stage to the color-breaking stage is critical for yield formation. As shown in Fig. 4C-E, from the fruit expansion stage to the mature stage, the increment in the number of large fruits was largest in the RB + R treatment, increasing from 0.06 to 5.17 per plant, followed by the RB + G treatment, increasing from 0.33 to 4.78 per plant. The number of small fruits in the CK treatment decreased from 11.89 to 3.17 per plant, a greater reduction than in the supplementary lighting treatments. During the expansion stage (Fig. 4C-E), except for the RB + FR and RB + UVA treatments, large fruits appeared in CK and the other four supplementary lighting treatments. The number of medium fruits in all supplementary lighting treatments was significantly higher than that in CK, and the RB + G treatment had a significantly higher number of medium fruits than the RB treatment. There were no significant differences in the number of small fruits among the various supplementary lighting treatments, but supplementary lighting tended to reduce the number of small fruits. During the fruit color-breaking stage (Fig. 4C-E), the total fruit number in the RB + B treatment was significantly higher than that in CK, while the number of large fruits in the RB + G treatment was significantly higher than that in CK. The number of medium fruits in the RB + G treatment was significantly lower than that in the RB + R and RB + B treatments. During the fruit mature stage (Fig. 4C-E), the total fruit number in the RB + R and RB + B treatments was significantly higher than that in CK. The number of large fruits in all supplementary lighting treatments was higher than that in CK, and the differences were significant for all supplementary lighting treatments except RB + UVA. Notably, the number of large fruits in the RB + R treatment was significantly increased by 63.09% and 25.79% compared with CK and the RB treatment, respectively. During the fruit mature stage, supplementary lighting treatments had no significant effect on the number of medium fruits, but the RB + B treatment had the highest number of medium fruits, significantly higher than the RB + G treatment. The RB + UVA treatment had the highest number of small fruits, significantly higher than the RB + R treatment.
Effects of supplementary lighting with different spectral compositions on nutrient contents in tomato fruits at different growth stages
Supplementary lighting significantly upregulated lycopene accumulation (Fig. 5A). From the fruit color-breaking stage to the mature stage, the increment in lycopene content was largest in the RB + G treatment, increasing from 0.90 to 39.08 g·kg− 1, while the increment in CK was the smallest, increasing from 0.73 to 13.66 g·kg− 1. During the fruit color-breaking stage, the lycopene content was highest in the RB + R treatment, with significant differences compared with other treatments (P < 0.05). Except for the RB treatment, all supplementary lighting treatments had significantly higher lycopene content than CK. From the color-breaking stage to the mature stage, lycopene accumulated rapidly, and all supplementary lighting treatments had significantly higher lycopene content than CK. At fruit maturity, the lycopene content in the RB + G treatment was the highest, followed by the RB + FR treatment. The difference between these two treatments was significant, and both were significantly higher than the other treatments.

Contents of nutritional substances in tomato fruits at the expansion, color-breaking, and mature stages under supplementary lighting with different spectral compositions. Lycopene (A), Vitamin C (B), Soluble protein (C), Soluble sugar (D), Organic acid (E), Sugar-acid ratio (F), Amino acid (G), Total phenols (H). Lycopene was not detected at the fruit expansion stage. Data are presented as mean ± standard error (n = 3 biological replicates). Different letters indicate significant differences at P < 0.05.
Vitamin C content gradually increased during fruit development, and its accumulation was influenced by light exposure (Fig. 5B). From the fruit expansion stage to the mature stage, the increment in vitamin C content was largest in the RB + B treatment, increasing from 3.07 to 11.40 g·100 g− 1, while the increment in CK was the smallest, increasing from 3.47 to 9.65 g·100 g− 1. During the fruit expansion stage, supplementary lighting decreased vitamin C content. Except for the RB + R treatment, all supplementary lighting treatments had significantly lower vitamin C content than CK. By the fruit color-breaking stage, the RB + R, RB + B, and RB + G treatments significantly promoted vitamin C accumulation, while the RB treatment remained significantly lower than CK and the other supplementary lighting treatments. At fruit maturity, the RB + B, RB + G, RB + FR, and RB + UVA treatments significantly promoted vitamin C accumulation, with the RB + B treatment having the highest content, significantly higher than other treatments except RB + G.
Soluble protein content exhibited a trend of initially decreasing and then increasing during tomato fruit development (Fig. 5C). During the fruit expansion stage, the RB, RB + R, and RB + B treatments significantly promoted soluble protein accumulation. By the fruit color-breaking stage, the RB + G treatment notably inhibited the degradation of soluble proteins, with accumulation significantly higher than in other treatments. After fruit ripening, soluble protein content in the RB + B and RB + UVA treatments was significantly higher than that in the CK, RB, and RB + FR treatments. During fruit development, the RB treatment exhibited an effect of initially promoting and subsequently inhibiting soluble protein synthesis.
Sugars and acids are two crucial indicators affecting the taste of tomatoes, and their contents and ratio are regulated by different spectral compositions (Fig. 5D-F). Supplementary lighting had no significant effect on sugar and acid contents during the early stage of tomato fruit development. As the duration of supplementary lighting gradually increased, by the fruit color-breaking stage, the RB + FR and RB + UVA treatments significantly promoted soluble sugar accumulation, while the RB + G and RB + UVA treatments significantly promoted organic acid accumulation. When the duration of supplementary lighting was further extended to fruit maturity, the RB + R and RB + G treatments significantly promoted soluble sugar accumulation, with the soluble sugar content in the RB + G treatment reaching the highest value. Soluble sugar contents in the RB + R, RB + G, RB + FR, and RB + UVA treatments were significantly higher than those in CK and the RB treatment. The RB + R and RB + UVA treatments significantly promoted organic acid accumulation during the mature stage, whereas the RB, RB + G, and RB + FR treatments showed the opposite effect. The sugar-acid ratio in the RB + G treatment was the highest at fruit maturity, followed by the RB + FR treatment. The difference between these two treatments was significant, and both were significantly higher than other treatments.
The trend of amino acid content changes was similar to that of soluble protein, and its content was also influenced by different spectral compositions (Fig. 5G). During the fruit expansion stage, the RB, RB + B, and RB + FR treatments significantly promoted amino acid synthesis. During the fruit color-breaking stage, in addition to the RB treatment, the other supplementary lighting treatments also significantly promoted amino acid synthesis, and amino acid contents were significantly higher than in the RB treatment. After fruit ripening, amino acid contents in the RB and RB + UVA treatments decreased significantly. The effect of the RB treatment on amino acid synthesis was similar to its effect on soluble protein, exhibiting an initial promotion followed by inhibition.
Total phenol content in tomato fruits at all growth stages was influenced by different spectral compositions (Fig. 5H). Specifically, during the fruit expansion stage, the RB + R, RB + G, and RB + FR treatments significantly promoted total phenol accumulation. During the fruit color-breaking stage, the RB and RB + G treatments significantly promoted total phenol accumulation. At fruit maturity, total phenol content in the RB + R, RB + B, RB + G, and RB + UVA treatments was significantly higher than that in CK, RB, and RB + FR treatments.
Effects of supplementary lighting with different spectral compositions on tomato fruit firmness and color
Fruit firmness and color are important appearance quality traits and serve as significant indicators for assessing fruit storability and maturity. Supplementary lighting had no significant effect on fruit firmness or the L value of tomato fruits (Fig. 6). Except for the RB + G treatment, which had no impact on the a* value, all other supplementary lighting treatments significantly increased the a* value of the fruits (P < 0.05). The RB + FR treatment exhibited the highest a* and b* values, significantly surpassing those of all other treatments.

Effects of supplementary lighting with different spectral compositions on tomato fruit firmness and color. Fruit firmness (A), L (B) value (lightness), a* (C) value (green-red: a* < 0 indicates green, a* > 0 indicates red), b* (D) value (blue-yellow: b* < 0 indicates blue, b* > 0 indicates yellow). Data are presented as mean ± standard error (n = 3 biological replicates). Different letters indicate significant differences at P < 0.05.
Discussion
The light environment (including light quality, light intensity, and photoperiod) plays a significant regulatory role in plant morphogenesis. Previous studies have shown that supplementary red-blue light can significantly increase tomato plant height, stem diameter, leaf area, and biomass2. Red, blue, and green light can significantly increase leaf area20,21. This study found that in the later stage of supplementary lighting (42 days), the RB and RB + R treatments significantly (P < 0.05) increased leaf count and leaf length × leaf width. This may be because the increased light intensity induced leaf growth and leaf area expansion, and red light (with a red light ratio ≥ 50%) may play a major role in these processes. However, there was no significant difference in leaf count and leaf length × leaf width between the RB + B, RB + G, RB + FR, RB + UVA treatments and CK. This might be due to the relatively low proportion of red light in the total light intensity (red light ratio < 50%). Blue light or mixed red-blue light is beneficial for chlorophyll accumulation22. In this study, the leaf SPAD value in the RB + B treatment was significantly higher than that in other treatments in the later stage of supplementary lighting. This could be because blue light facilitates chlorophyll accumulation, or because the higher light intensity of the RB + B treatment promoted chlorophyll accumulation. Some research results indicate that adding UVA significantly promotes stem growth in tomato seedlings17, while another study reported opposite results19. This discrepancy arises because different species have different responses to light of the same wavelength. In this study, adding UVA promoted plant height growth in the early stage of supplementary lighting (14 days), but there was no significant difference compared with other treatments in the later stage (42 days). It is speculated that short-term UVA exposure can stimulate tomato plant growth.
Light quality and light intensity can influence the ripening process of tomato fruits. Red light accelerates fruit ripening by inducing ethylene synthesis1 and promotes early fruit coloring by enhancing the accumulation of lycopene and β-carotene23. Blue light also exhibits similar effects24. Studies have shown that red-blue light induce the expression of related genes by activating the phototransduction factors HY5 and PIFs, thereby regulating lycopene, carotenoid, and sugar metabolism to promote fruit ripening3,25. Post-harvest tomato fruit ripening can also be promoted by a combination of red and ultraviolet light26. The results of this study indicate that the different spectral supplementary lighting treatments dominated by red-blue light promoted lycopene accumulation and accelerated fruit ripening, which is consistent with previous research findings. Currently, research on the regulatory mechanisms of green, far-red, and ultraviolet light on fruit ripening is relatively scarce. Future studies are needed to deeply explore the roles of these light qualities and light intensities in fruit development to expand the understanding of light-regulated ripening mechanisms.
These key light environmental factors (light quality, light intensity, and photoperiod) are also critical determinants of tomato yield formation. Under low-light conditions, artificial supplementary lighting, especially the supplementation of red-blue light, can significantly increase tomato yield. The mechanism involves, on one hand, optimizing the light quality ratio and increasing light intensity to enhance the crop photosynthetic rate, and on the other hand, extending the photoperiod to prolong the duration of crop photosynthesis. Photosynthesis is the foundation of yield formation. Both increasing the photosynthetic rate and extending its duration are conducive to biomass accumulation27. The results of this study indicate that, compared with no supplementary lighting (CK), supplementing with composite light dominated by red-blue light promoted fruit expansion by optimizing the light quality combination, increasing light intensity, and prolonging illumination time, thereby increasing yield per plant. Notably, the yield per plant in the RB + R and RB + G treatments was significantly higher than that in the RB treatment. This may result from the combined effect of increased light intensity and the photobiological characteristics of red light and green light. Specifically, red light may be more effective in promoting plant photosynthesis than other light qualities, while green light has the potential to regulate carbohydrate distribution, improve the crop canopy light environment, and drive photosynthesis12,13,28. The addition of red light and green light increased light intensity and enhanced plant photosynthetic capacity, thus increasing tomato yield per plant. In lettuce, replacing part of the blue light with far-red light can promote biomass accumulation29, and in tomatoes, adding far-red light to red light can increase fruit dry and fresh weight30. This study found that the order of tomato yield per plant from high to low was RB + R, RB + G, RB + B, RB + FR, RB, RB + UVA, CK. The yield per plant of the RB + B and RB + FR treatments was higher than that of the RB treatment, but the difference was not significant. Therefore, it is speculated that this may be related to the magnitude of the increase in light intensity from blue and far-red light. Subsequent studies could conduct relevant research based on this hypothesis, exploring different light intensities.
Light quality, light intensity, and photoperiod all have significant effects on tomato quality formation. Studies have found that red light can positively regulate the accumulation of lycopene, soluble sugars, and phenolic compounds. Under conditions of combined red and ultraviolet light or a high red/far-red ratio, lycopene accumulation is significantly increased26,31,32. Blue light is conducive to the accumulation of vitamin C, soluble proteins, amino acids, total phenols, lycopene, and carotenoids33,34. In this study, compared with no supplementary lighting, the increased light intensity and illumination duration from supplementary lighting provided more light energy for the synthesis of various nutrients, promoting the overall accumulation of lycopene, vitamin C, soluble sugars, and other nutrients. However, this study also observed that some supplementary lighting treatments inhibited the accumulation of certain nutrients. For example, the RB treatment reduced the contents of vitamin C, soluble protein, soluble sugar, and amino acids in the later stage of supplementary lighting (fruit color-breaking stage and mature stage), and the RB, RB + G, and RB + FR treatments reduced organic acid content at the fruit mature stage. These results indicate that tomato plants exhibit significantly different responses to different light qualities and intensities at different growth stages. Previous studies have shown that far-red light positively regulates soluble sugar accumulation in tomatoes35. UVA can stimulate lettuce growth and promote the accumulation of vitamin C, soluble sugars, soluble protein, and phenolic compounds18,36. Replacing a small amount of red light with green light can significantly promote sucrose accumulation and regulate nitrogen metabolism37. Compared with the RB treatment, the differences in nutrient contents in the other supplementary lighting treatments were mainly manifested during the late stage of fruit development (color-breaking stage and mature stage). Specifically, compared with the RB treatment, supplemental red light and blue light significantly increased vitamin C and soluble protein during the late stage of fruit development; supplemental green light significantly increased lycopene, soluble protein, amino acid, and total phenol contents; supplemental far-red light significantly increased soluble protein, soluble sugar, and amino acid contents, as well as the sugar-acid ratio; supplemental UVA significantly increased soluble protein, soluble sugar, and organic acid contents. This indicates that both increased light intensity and altered light quality can regulate nutrient metabolism in tomato fruits, and tomato fruits may have enhanced light sensitivity during the late stage of development. The color of tomato fruits is related to the accumulation of pigments such as lycopene, carotenoids, and anthocyanins. During fruit ripening, chlorophyll is degraded while pigments like lycopene are synthesized. Supplementary lighting accelerates chlorophyll degradation and promotes pigment accumulation38, thereby increasing the fruit redness value.
Due to the varying requirements of plants for different spectral compositions, this study designed different spectral supplementary lighting treatments based on conclusions from previous research. Consequently, there were differences in both light quality and light intensity among the treatments. Therefore, it remains unclear whether the results of this experiment are driven solely by light quality, solely by light intensity, or by the interaction of light quality and light intensity. Subsequent research can be conducted to address this limitation. Additionally, this study only selected the ‘Provence’ cultivar for investigation and did not measure photosynthetic parameters. These aspects can be explored in future research.
Conclusion
This study systematically compared the effects of supplementary lighting with different spectral compositions dominated by red-blue light on plant growth, fruit ripening indicators, yield components, and quality formation in facility-grown tomatoes under low-light and short-day conditions. The results showed that, compared with no supplementary lighting, supplementary lighting with different spectral compositions all promoted plant growth and had significant effects on improving quality, increasing yield, and accelerating fruit ripening (P < 0.05). Compared with the RB treatment, the RB + R treatment significantly increased yield per plant and the number of large fruits by 6.94% and 25.79%, respectively (P < 0.05), demonstrating the best yield-increasing effect among all supplementary lighting treatments; the RB + B treatment was beneficial for increasing stem diameter and leaf SPAD values, and the contents of vitamin C and soluble protein were significantly higher than those in the RB treatment during the late stages of fruit development (fruit color-breaking stage and mature stage); the RB + G treatment had the highest average fruit weight, which was significantly higher (by 8.29%) than that of the RB treatment; during the late stages of fruit development, the RB + G treatment had significantly higher contents of lycopene, soluble protein, amino acids, and total phenols than the RB treatment; the RB + FR treatment significantly increased soluble protein, soluble sugar, and amino acid contents, as well as the sugar-acid ratio, during the late stages of fruit development, and significantly elevated fruit color parameters (a* and b* values) compared with the RB treatment; the RB + UVA treatment stimulated plant height growth in the short term and significantly increased soluble protein, soluble sugar, and organic acid contents during the late stages of fruit development.
Implications and future research
This study addresses the practical production problem of low-light stress in facility-cultivated tomatoes caused by insufficient sunshine in winter and spring in Guizhou Province, and clarifies the regulatory effects of supplementary lighting with different spectral compositions on tomato growth, yield, and quality. Growers can select the corresponding supplementary lighting schemes according to different production objectives, such as increasing yield, improving nutritional quality, and promoting early fruit ripening, which provides a practical reference for supplementary lighting cultivation of facility tomatoes in the local area. In subsequent research, the independent regulatory effects of light quality and light intensity can be further explored, adaptability studies of supplementary lighting can be carried out for more main cultivated tomato varieties, and the technical parameters of supplementary lighting can be optimized in combination with actual field production conditions.
Data availability
The datasets generated during or analysed during the current study are not publicly available due to the data that has been used is confidential but are available from the corresponding author on reasonable request.
References
Zhang, J. Y. et al. Supplementary red light results in the earlier ripening of tomato fruit depending on ethylene production. Environ. Exp. Bot. 175, 104044 (2020a).
Wang, S. Y. et al. Red and blue LED light supplementation in the morning pre-activates the photosynthetic system of tomato (Solanum lycopersicum L.) leaves and promotes plant growth. Agronomy 12, 897 (2022).
Wang, J. C. et al. Manipulating the light systemic signal HY5 greatly improve fruit quality in tomato. Adv Sci. 12, 2500110 (2025).
Zhang, Y. et al. Manipulation of artificial light environment improves plant biomass and fruit nutritional quality in tomato. J. Adv. Res. 75, 79–93 (2024).
Trouwborst, G., Hogewoning, S. W., Kooten, O. V., Harbinson, J. & Ieperen, W. V. Plasticity of photosynthesis after the ‘red light syndrome’ in cucumber. Environ. Exp. Bot. 121, 75–82 (2016).
Yang, X. L. et al. Response of photosynthetic capacity of tomato leaves to different LED light wavelength. Environ. Exp. Bot. 150, 161–171 (2018).
Li, Y. et al. Effects of red and blue light on leaf anatomy, CO2 assimilation and the photosynthetic electron transport capacity of sweet pepper (Capsicum annuum L.) seedlings. BMC Plant. Biol. 20, 318 (2020).
Kong, Y., Schiestel, K. & Zheng, Y. B. Pure blue light effects on growth and morphology are slightly changed by adding low-level UVA or far-red light: A comparison with red light in four microgreen species. Environ. Exp. Bot. 157, 58–68 (2019).
Frechilla, S., Talbott, L. D., Bogomolni, R. A. & Zeiger, E. Reversal of blue light-stimulated stomatal opening by green light. Plant. Cell. Physiol. 41, 171–176 (2000).
Zhang, T. T., Maruhnich, S. A. & Folta, K. M. Green light induces shade avoidance symptoms. Plant. Physiol. 157, 1528–1536 (2011).
Battle, M. W. & Jones, M. A. Cryptochromes integrate green light signals into the circadian system. Plant. Cell. Environ. 43, 16–27 (2020a).
Kaisera, E., Weerheim, K., Schipper, R. & Dieleman, J. A. Partial replacement of red and blue by green light increases biomass and yield in tomato. Sci. Hortic. 249, 271–279 (2019).
Lanoue, J., Little, C., Hawley, D. & Hao, X. M. Addition of green light improves fruit weight and dry matter content in sweet pepper due to greater light penetration within the canopy. Sci. Hortic. 304, 111350 (2022).
Wang, Y. H., Zhang, T. T. & Folta, K. M. Green light augments far-red-light-induced shade response. Plant. Growth Regul. 77, 147–155 (2015).
Zhang, Y. T., Zhang, Y. Q., Yang, Q. C. & Li, T. Overhead supplemental far-red light stimulates tomato growth under intra-canopy lighting with LEDs. J. Integr. Agric. 18, 62–69 (2019).
Liu, X. J. et al. Growth of tomato and cucumber seedlings under different light environments and their development after transplanting. Front. Plant. Sci. 14, 1164768 (2023).
Kang, S. et al. Ultraviolet-A radiation stimulates growth of indoor cultivated tomato (Solanum lycopersicum) seedlings. HortScience 53, 1429–1433 (2018).
Chen, Y. C. et al. UVA radiation is beneficial for yield and quality of indoor cultivated lettuce. Front. Plant. Sci. 10, 1563 (2019).
Zhang, Y. T. et al. UVA radiation promotes tomato growth through morphological adaptation leading to increased light interception. Environ. Exp. Bot. 176, 104073 (2020b).
Claypool, N. B. & Lieth, J. H. Physiological responses of pepper seedlings to various ratios of blue, green, and red light using LED lamps. Sci. Hortic. 268, 109371 (2020).
Yavari, N. et al. The effect of light quality on plant physiology, photosynthetic, and stress response in Arabidopsis thaliana leaves. PLoS One. 16, e0247380 (2021).
Li, Y., Liu, Z. L., Shi, Q. H., Yang, F. J. & Wei, M. Mixed red and blue light promotes tomato seedlings growth by influencing leaf anatomy, photosynthesis, CO2 assimilation and endogenous hormones. Sci. Hortic. 290, 110500 (2021).
Panjai, L. et al. Effect of postharvest irradiation with red light on epidermal color and carotenoid concentration in different parts of tomatoes. J. Food Meas. Charact. 15, 1737–1746 (2021).
Wang, W. et al. Effects of supplemental lighting on potassium transport and fruit coloring of tomatoes grown in hydroponics. Int. J. Mol. Sci. 22, 2687 (2021).
Xie, B. X. et al. Supplemental blue and red light promote lycopene synthesis in tomato fruits. J. Integr. Agric. 18, 590–598 (2019).
Panjai, L., Noga, G., Fiebig, A. & Hunsche, M. Effects of continuous red light and short daily UV exposure during postharvest on carotenoid concentration and antioxidant capacity in stored tomatoes. Sci. Hortic. 226, 97–103 (2017).
Yu, H. J. et al. The effects of different durations of night-time supplementary lighting on the growth, yield, quality and economic returns of tomato. Plants 13, 1516 (2024).
Battle, M. W., Vegliani, F. & Jones, M. A. Shades of green: untying the knots of green photoperception. J. Exp. Bot. 71, 5764–5770 (2020b).
Hu, Y. Z. et al. Effects of substituting B with FR and UVA at different growth stages on the growth and quality of lettuce. Agronomy 13, 2547 (2023).
Kim, H. J. et al. Supplemental intracanopy far-red radiation to red LED light improves fruit quality attributes of greenhouse tomatoes. Sci. Hortic. 261, 108985 (2020).
Li, Q. & Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 67, 59–64 (2009).
Nájera, C., Guil-Guerrero, J. L., Enríquez, L. J., Álvaro, J. E. & Urrestarazu, M. LED-enhanced dietary and organoleptic qualities in postharvest tomato fruit. Postharvest Biol. Technol. 145, 151–156 (2018).
He, R. et al. Supplemental blue light frequencies improve ripening and nutritional qualities of tomato fruits. Front. Plant. Sci. 13, 888976 (2022).
Xiao, L., Shibuya, T., Watanabe, T., Kato, K. & Kanayama, Y. Effect of light quality on metabolomic, ionomic, and transcriptomic profiles in tomato fruit. Int. J. Mol. Sci. 23, 13288 (2022).
Fanwoua, J. et al. Supplemental LED lighting affects the dynamics of tomato fruit growth and composition. Sci. Hortic. 256, 108571 (2019).
Loconsole, D. & Santamaria, P. UV lighting in horticulture: A sustainable tool for improving production quality and food safety. Horticulturae 7, 9 (2021).
Chen, Y. Q. & Liu, W. K. Substituting green light for partial red light promoted the growth and quality, and regulated the nitrogen metabolism of Medicago sativa grown under red-blue LEDs. Environ. Exp. Bot. 220, 105623 (2024).
Xu, Y. N. et al. Red and blue light promote tomato fruit coloration through modulation of hormone homeostasis and pigment accumulation. Postharvest Biol. Technol. 207, 112588 (2024).
Funding
This work was supported by the Guizhou Provincial Science and Technology Project [Qian Ke He Platform Talent-YQK(2023)025], Key Core Technology Research Project for Mountainous Agriculture in Guizhou Province[GZNYGJHX-2024005] and the Guizhou Provincial Basic Research Program (Qian Ke He Ji Chu MS [2026]402)
Author information
Authors and Affiliations
Contributions
Dachun Sun: Writing-original draft, Writing-review & editing, Conceptualization, Formal analysis, Methodology, Validation, Data curation. Chao Ma: Writing-review & editing, Conceptualization, Funding acquisition, Project administration, Supervision, Validation. Xiaocui Liu: Conceptualization, Project administration, Validation, Methodology. Tiantian Ban: Conceptualization, Project administration, Resources, Methodology. Zhuo Chen: Project administration, Resources.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sun, D., Ma, C., Liu, X. et al. Effects of supplementary lighting with different spectral compositions on plant growth, fruit development, and quality formation of facility-grown tomatoes. Sci Rep 16, 10737 (2026). https://doi.org/10.1038/s41598-026-44021-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-44021-6
