
Abstract
The drought resistance of Cinnamomum migao seedlings inoculated with endophytic fungi was explored under different drought stress conditions. The seedlings of C. migao were inoculated with Trichoderma harzianum and Trichoderma longibrachiatum in the rhizosphere, with non-inoculated seedlings serving as the control. Then, the seedlings of C. migao were exposed to different drought stress gradients: normal water supply (80 ± 5% moisture, control), mild (60 ± 5% moisture, T1), moderate (45 ± 5% moisture, T2), and severe (25 ± 5% moisture, T3) drought stress. After 30 days of each drought stress treatment, samples were collected, and growth parameters, osmotic substance content, antioxidant enzyme activity, chlorophyll content, and endogenous hormone concentrations were measured. Compared with the control group, inoculation with T. harzianum and T. longibrachiatum promoted the growth of C. migao seedlings to varying extents. This inoculation also enhanced their root absorption capacity, increased the relative water content, chlorophyll, and abscisic acid (ABA) levels in leaf tissues (p < 0.05), and facilitated the accumulation of osmotic substances while boosting antioxidant enzyme activity (p < 0.05). Furthermore, it reduced water potential and decreased the contents of malonaldehyde (MDA), indoleacetic acid (IAA), gibberellin (GA₃), and ribosylzeatin (ZR) (p < 0.05). T. harzianum and T. longibrachiatum significantly improved the drought resistance of C. migao seedlings under stress conditions. These findings provide a theoretical basis for further research and development of microbial inoculants.
Similar content being viewed by others

Introduction
Cinnamomum migao is an evergreen tall tree belonging to the Lauraceae family and the Cinnamomum genus. It is a unique medicinal plant that is endemic to the dry-hot valley regions at the intersection of Yunnan, Guizhou, and Guangxi provinces in China1. The dried mature fruit of C. migao is a traditional medicine commonly used by the Miao ethnic group in Guizhou. It was first documented in the Supplement to the Compendium of Materia Medica (Bencao Gangmu Shiyi)2 and is known for its therapeutic effects, including warming the middle jiao to dispel cold, regulating Qi to alleviate pain, and treating conditions like stomachache, abdominal pain, chest pain, rheumatoid arthritis, vomiting, asthma, chest tightness, and motion sickness-induced nausea3,4,5.
In recent years, Chinese patent medicines that use the fruit of C. migao as the primary raw material—such as Liqi Huoxue Dropping Pills, Jin Houjian Spray, Xinwei Zhitong Capsules, and Xinnaoning Capsules—have generated annual output values reaching billions of yuan. Among them, four ethnic Chinese patent medicines developed by enterprises in Guizhou Province, using the fruit of C. migao as the raw material have achieved production values in the billions of RMB, indicating broad prospects for development and utilization6. However, due to the extremely low wild reserves of the fruit of C. migao and the decline of its natural populations, the supply–demand imbalance has become increasingly severe, significantly restricting the development of related pharmaceutical industries. Although our research team has initiated artificial cultivation, the slow growth rate of C. migao and drought stress remain major limiting factors for its optimal growth. When drought stress causes water deficit, it disrupts the water supply to meristem cells and reduces their turgor pressure. This inhibits cell division and growth, leading to impaired cell development. Consequently, plants exhibit reduced leaf area and smaller leaves, which diminishes the total photosynthetic area. This hinders vegetative growth, resulting in stunted plants and a prolonged growth period from seedling to fruit maturation, ultimately affecting both crop yield and quality. Under normal environmental conditions, the production and scavenging of reactive oxygen species (ROS) in plant cells maintain a dynamic equilibrium. However, under drought stress, this balance is disrupted, leading to excessive ROS accumulation. ROS-mediated oxidative stress can trigger various harmful cytological effects, including membrane lipid peroxidation, nuclear damage, impaired photosynthesis, and abnormal respiration7,8.
Related studies have found that inoculating plants with endophytic fungi can significantly enhance their drought resistance and mitigate the adverse effects of drought stress9. Endophytes, which include both bacteria and fungi, inhabit healthy plant tissues and contribute to promoting plant growth10. Ongoing research on endophytic fungi has revealed that mycorrhizal fungi play a crucial role in improving water metabolism and drought tolerance in trees. These fungi absorb nutrients such as nitrogen and phosphorus from the soil and transfer them to their host plants to support growth. In exchange, the plants provide carbohydrates in the form of lipids or sugars, which are essential for fungal growth11,12. Additionally, mycorrhizal fungi mediate interactions between plants and the microbiome in the mycorrhizosphere, further promoting plant growth, nutrient uptake, and tolerance to biotic and abiotic stresses13. Furthermore, studies have shown that Trichoderma harzianum and Trichoderma longibrachiatum can effectively reduce ROS accumulation, enhance the synthesis of osmoregulatory substances, and improve cellular water absorption capacity9,14. The detrimental effects of drought extend beyond impaired plant physiology, critically compromising fruit yield and overall agricultural productivity. Water deficit during key growth stages disrupts flowering, fruit set, and development, leading to significant reductions in both the quality and quantity of the harvest, which poses a substantial threat to food security and the economic viability of crops like C. migao.
Currently, limited research exists regarding the role of endophytic fungi in enhancing the drought resistance of C. migao's. Therefore, in this study, six-month-old C. migao seedlings, which had been inoculated with T. harzianum, T. longibrachiatum, or subjected to a non-inoculation control treatment, were exposed to different water gradients for 90 days. We measured the growth indices, biomass, physiological parameters, and endogenous signaling molecules to analyze the effects of these endophytic fungi on the drought resistance of C. migao seedlings under varying water conditions.
Materials and methods
Source of endophytic fungi
The endophytic fungi T. harzianum (accession number: PX916279) and T. longibrachiatum (accession number: PX916280) were isolated by our research group from the root systems of three healthy C. migao trees collected in Qiannan Prefecture, Guizhou Province.
Source of C. migao seedlings
Seeds of C. migao were collected from the Pingtang County plantation base in Qiannan Prefecture, Guizhou Province. In October 2020, the seeds were surface-sterilized by immersion in a 0.2% potassium permanganate (KMnO₄) solution for 30 min, followed by thorough rinsing with distilled water. The sterilized seeds were sown in a sterilized germination substrate composed of river sand and wood chips (sterilized at 105℃ for 2 h) within seedling trays. The trays were maintained in a constant temperature incubator at 25℃ for germination. After 20 d, the seedlings were transferred from the incubator for a hardening-off period to acclimate them to the eventual planting environment. During this period, the seedlings were irrigated with an appropriate amount of distilled water. In early March 2021, the seedlings were transplanted into plastic pots (upper outer diameter: 20.5 cm; bottom outer diameter: 14.5 cm; height: 17 cm) for subsequent cultivation.
Methods
This experiment comprised three groups: one inoculated with T. harzianum, one inoculated with T. longibrachiatum, and a non-inoculated control group. Inoculation was performed via a rhizosphere injection method. Using a 10 mL pipette (with the fine tip appropriately cut off), 15 mL of the pre-prepared fungal suspension was administered to each relevant pot weekly. This was repeated three times, resulting in a total inoculation volume of 45 mL per pot. 90 d after inoculation, drought stress was simulated using the pot weighing method. 4 soil moisture gradients were established: 80 ± 5% of field capacity (FC) as the control (CK), 60 ± 5% FC (mild drought, T1), 45 ± 5% FC (moderate drought, T2), and 25 ± 5% FC (severe drought, T3). Each drought stress treatment lasted for 30 d before proceeding to the next gradient. The experiment included three treatments, with 20 pots per treatment and three replicates, resulting in a total of 180 pots (one seedling per pot). After 30 days of each moisture treatment, C. migao seedlings were sampled (5 plants per sampling) for measurements of growth parameters, water physiology, biomass, physiological indicators, and endogenous signaling substances (Figure S1).
Water physiology measurements
The measurements were initiated 10 months after sowing. For each treatment, 5 C. migao seedlings were randomly selected. The number of leaves and branches were recorded. The seedling height (vertical distance from the substrate surface to the highest natural growth point), branch length, and ground diameter (at a position 5 cm above the substrate) were measured using a measuring tape and a vernier caliper. The leaves were labeled, immediately placed in a foam icebox with ice packs, and transported to the laboratory for prompt analysis. Surface impurities were gently wiped off, the relative water content (RWC, %) was calculated using the following formula: RWC (%) = (W1 − W3)/(W2 − W3) × 10015. Additionally, leaf water potential (Ψ) was measured using a portable plant water potential pressure chamber. And the fresh weight (W1) was measured using a 0.001 g precision electronic balance. The weighed leaves were then fully submerged in distilled water in a beaker, sealed with plastic wrap, and kept in darkness for 24 h to achieve full saturation. After saturation, excess water was removed, and the saturated fresh weight (W2) was recorded. The leaves were then oven-dried at 105 °C for 30 min to deactivate enzymes, followed by drying at 60 °C to a constant weight to obtain the dry weight (W3).
Growth parameters and biomass measurement
5 C. migao plants were randomly selected from each treatment group. The number of leaves and branches were recorded. Plant height (vertical height from the substrate surface to the highest natural growth point), branch length, and ground diameter (measured at 5 cm above the substrate) were measured using a tape measure and vernier calipers. Leaves from the middle-upper part of the seedlings were collected, laid flat in the center of the leaf area meter’s scanning area without overlapping or curling, and scanned according to the instrument’s manual to determine leaf area. After the above measurements, the seedling pots were carefully cut open to extract intact root systems. The roots were rinsed with clean water, and the taproot length was measured. A root scanner was used to obtain the average root diameter.
The stems of the seedlings were cut at ground level, and the separated leaves and stems were placed in pre-labeled envelope bags. The roots were cleaned of adhering soil, rinsed under running water, and blotted dry with a towel before being placed in labeled envelopes. All samples were then oven-dried at 105 °C for 30 min to deactivate enzymes, followed by drying at 60 °C to a constant weight. The dry weights of the seedlings’ organs (leaves, stems, and roots) were recorded to determine biomass allocation.
Physiological index measurement
Soluble protein content was determined using the Coomassie Brilliant Blue G-250 staining method16,17. Soluble sugar content was measured using the anthrone colorimetric method18. SOD activity was determined using the xanthine oxidase method, as previously described. Briefly, fresh leaves of C. migao were rinsed with distilled water, blotted dry, and cut into pieces (midribs removed). The leaf segments were homogenized, and 0.1 g of the tissue (with 5 biological replicates per treatment) was weighed into an ice-cooled mortar. The tissue was ground to a homogenate with pre-chilled phosphate buffer (pH 7.8). The homogenate was transferred to a centrifuge tube and centrifuged at 10,000 × g for 15–20 min at 4 °C. The resulting supernatant was collected for the assay. The reaction mixture, prepared in sequence, consisted of 30 μL of phosphate buffer, 50 μL of substrate mix, 20 μL of the crude enzyme extract, and 20 μL of xanthine oxidase solution. The mixture was incubated at 37 °C for 30 min. Subsequently, 20 μL of color-developing agent (a mixture of sulfanilic acid and N-1-naphthylethylenediamine dihydrochloride) was added, mixed thoroughly, and allowed to stand for 20 min. The absorbance was measured at 550 nm using a spectrophotometer19. Free proline content, catalase (CAT), and peroxidase (POD) activities were measured using visible spectrophotometry with commercial assay kits. Fresh leaf samples were processed as described above. For proline extraction, 0.1–0.2 g of leaf tissue (5 replicates per treatment) was homogenized in 2 mL of 3% sulfosalicylic acid on ice. The homogenate was transferred to a centrifuge tube, incubated in a boiling water bath for 10 min, and then cooled under running water. The extract was centrifuged at 4,000 × g for 10 min at 4 °C, and the supernatant was collected. Subsequent steps followed the manufacturer’s instructions for the respective kits, and absorbance was recorded at 520 nm for proline quantification. CAT and POD activities were determined according to their respective kit protocols using the same spectrophotometric approach. MDA content was measured using the thiobarbituric acid (TBA) method with a commercial assay kit. Fresh leaf tissue (0.1–0.3 g, 5 replicates per treatment) was homogenized in 2 mL of 10% trichloroacetic acid (TCA) and a small amount of quartz sand in an ice-cooled mortar. The homogenate was centrifuged at 4,000 × g for 10 min at 4 °C, and the supernatant was collected. The assay was performed according to the kit manufacturer’s instructions, and absorbance readings were taken at 532 nm and 600 nm to calculate MDA concentration based on the specific absorbance difference. All assay kits used for the above measurements were purchased from Guizhou BioTec Trading Co., Ltd., sourced from the Nanjing Jiancheng Bioengineering Institute, China.
The pieces were mixed evenly and 0.2 g of the cut fresh sample were weighed, with five replicates for each treatment. The weighed samples were immersed in 25 ml of a mixture of 80% acetone and absolute ethanol (acetone: absolute ethanol = 1.5:1). The samples were soaked in the dark for 48 h until the tissue turns completely white. The solution was filtered into a 25 ml volumetric flask using filter paper, then dilute to the mark with 80% acetone, mix well, and the chlorophyll extract was obtained. Using the mixture of 80% acetone and absolute ethanol as a blank control, the absorbance (OD) values were measured at wavelengths of 470 nm, 645 nm, and 663 nm. Chlorophyll a, chlorophyll b, and chlorophyll (a + b) were calculated using the following formulas20:
\(Chlorophyll{\text{ }}a{\text{ }} = {\text{ }}\left( {12.72{\text{ }} \times {\text{ }}OD_{{663}} - {\text{ }}2.59{\text{ }} \times {\text{ }}OD_{{645}} } \right){\text{ }} \times {\text{ }}V{\text{ }}/{\text{ }}\left( {1000{\text{ }} \times {\text{ }}W} \right)\)
\(Chlorophyll{\text{ }}b{\text{ }} = {\text{ }}\left( {22.88{\text{ }} \times {\text{ }}OD_{{645}} - {\text{ }}4.67{\text{ }} \times {\text{ }}OD_{{663}} } \right){\text{ }} \times {\text{ }}V{\text{ }}/{\text{ }}\left( {1000{\text{ }} \times {\text{ }}W} \right)\)
\(Chlorophyll{\text{ }}\left( {a + b} \right){\text{ }} = {\text{ }}\left( {20.29{\text{ }} \times {\text{ }}OD_{{645}} + {\text{ }}8.04{\text{ }} \times {\text{ }}OD_{{663}} } \right){\text{ }} \times {\text{ }}V{\text{ }}/{\text{ }}\left( {1000{\text{ }} \times {\text{ }}W} \right)\)
Where: V is the volume of the extraction solution (ml), and W is the fresh weight of each sample (g).
Determination of endogenous hormone content
The contents of indole-3-acetic acid (IAA), abscisic acid (ABA), gibberellic acid (GA₃), and zeatin riboside (ZR) in C. migao seedlings from the three inoculation groups (EF, T. harzianum, and T. longibrachiatum) under different drought levels were determined using enzyme-linked immunosorbent assay (ELISA). Fresh leaf samples (0.1 g) were homogenized in liquid nitrogen, and 1 mL of phosphate-buffered saline (PBS), pH 7.4 was added. The homogenates were flash-frozen in liquid nitrogen and stored for 12 h. After thawing at 2–8 °C, the samples were thoroughly homogenized manually or with a homogenizer, followed by centrifugation at 2000–3000 × g for 20 min. The supernatant was carefully collected, aliquoted, and stored at –20 °C for subsequent analysis.
All reagent preparations and assay procedures were strictly performed according to the manufacturer’s instructions. Absorbance was measured at 450 nm using a microplate reader (Rayto RT-6100), and hormone concentrations for each treatment group were calculated based on standard curves.
Data processing and analysis
Data were processed using Microsoft Excel 2019 and SPSS 26.0. Comparisons betw-een different treatments were conducted using one-way ANOVA, and the results were expressed as the mean ± standard deviation (n = 5). Graphs were generated using Origin 2024 and Adobe Photoshop 2024. Bioinformatic analysis was performed usi-ng the OmicStudio tools at https://www.omicstudio.cn/tool.
Results
Effects of endophytic fungi on physiological growth of C. migao seedlings under drought stress
The plant height of C. migao seedlings in all three inoculated groups exhibited a gradual increase. However, significant differences were observed among seedlings subjected to the three inoculation treatments. Under varying drought treatments, the ranking of plant height was as follows: T. harzianum group > T. longibrachiatum group > Control group, with the inoculated groups showing significantly greater plant height than the Control group (Fig. 1a). In the T. harzianum and T. longibrachiatum groups, growth slowed under the T2 drought treatment, with no significant difference in plant height compared to the T1 treatment. In the Control group, stem thickness growth slowed under the T3 drought treatment, showing no significant difference from the T2 treatment. The ground diameter of the inoculated groups was significantly greater than that of the Control group, though no notable difference was observed between the two inoculated groups (Fig. 1b). In the Control group, ground diameter increased significantly under the T1 drought treatment. However, both the T. harzianum group and the Control group showed no significant growth under the T3 treatment, whereas the T. longibrachiatum group exhibited notable growth under the same conditions. These results indicate that inoculation with T. harzianum and T. longibrachiatum is more conducive to the growth of plant height and ground diameter.

Effects of endogenous fungi inoculation on plant height and ground diameter of C. migao seedlings under different drought stress. 80 ± 5% (CK), 60 ± 5% (T1), 45 ± 5% (T2), 25 ± 5% (T3), the same below.
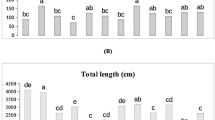
The decreasing water availability, the leaf number of C. migao seedlings in all three inoculated groups reached their maximum values under T1 water treatment (30.4, 38, and 41.8 for Control, T. longibrachiatum, and T. harzianum groups, respectively) (Fig. 2b). Both T. harzianum and Control groups achieved their maximum leaf areas under T1 treatment (4075.26 cm2 and 3964.64 cm2, respectively; Fig. 2b), while the T. longibrachiatum group peaked under CK conditions. The leaf length of T. harzianum and T. longibrachiatum groups showed a positive correlation with water availability, whereas the Control group exhibited an initial increase followed by a decrease, reaching maximum values under T1 treatment (Fig. 2c). Under T1 conditions, both leaf area and length were significantly greater in T. harzianum and Control groups compared to T. longibrachiatum. Under T2 treatment, T. harzianum group demonstrated significantly larger leaf area and length than both T. longibrachiatum and Control groups. These results suggest that inoculation with T. harzianum is more beneficial for C. migao seedling growth under drought conditions when considering leaf area and length parameters. Leaf width followed similar trends to leaf number across water treatments (Fig. 2d). Under T2 and T3 conditions, both T. harzianum and T. longibrachiatum groups showed significantly greater leaf width than the Control group. However, under CK and T1 treatments, the T. longibrachiatum group exhibited lower leaf width compared to the Control group.

Effects of endophytic fungi inoculation on leaf growth of C.migao seedlings under different drought stress.
The under T1 drought treatment, the ranking of branch length was: T. longibrachiatum group > T. harzianum group > Control group. The branch diameter of inoculated C. migao seedlings showed an initial increase followed by a decrease with varying drought treatments. The T. harzianum group reached its maximum value (2.792 mm) under T2 treatment, while the T. longibrachiatum group peaked (2.77 mm) under T1 treatment. No significant differences in branch diameter were observed among the three inoculated groups under CK conditions. Under T1 and T3 treatments, the ranking of branch diameter followed the same pattern as branch length under T1 treatment. However, under T2 treatment, the T. harzianum group exhibited significantly greater branch diameter compared to both the T. longibrachiatum and Control groups (Fig. 3).

Effects of endophytic fungi inoculation on the branching of C. migao seedlings under different drought stress.
The root diameter exhibited distinct responses to different drought treatments. The T. longibrachiatum group showed an initial decrease followed by an increase, reaching its minimum value under T1 treatment. In contrast, the Control group displayed an initial increase followed by a decrease, peaking at 1.868 mm under T2 treatment Fig. 4. The T. harzianum group demonstrated a significant reduction in root diameter under T2 but rebounded sharply under T3. For root length (Fig. 5b), both T. harzianum and Control groups exhibited an initial increase followed by a decrease with declining soil moisture. The T. harzianum group reached its maximum (42.92 mm) under T2, while the Control group peaked at 45.14 mm under T1. The T. longibrachiatum group showed an opposite trend (decrease-increase), with its minimum occurring at T2 (Fig. 4).

Effects of endophytic fungi inoculation on the roots of C. migao seedlings under different drought stress.
The T. harzianum-inoculated and Control groups showed similar trends in leaf relative water content (RWC) under drought stress, exhibiting an initial increase followed by a decrease. The maximum RWC values (89.08% for T. harzianum and 80.49% for Control) were observed under T1 treatment. All three groups displayed consistent water potential (Ψ) dynamics—decreasing initially, then recovering with progressive drought. The T. harzianum and T. longibrachiatum groups reached their Ψ minima under T2 treatment, while the Control group showed lowest Ψ at T1 (Fig. 5).

Effects of endophytic fungi inoculation on tissue water content of C. migao seedlings under different drought stress.
There was no significant difference between the T. Harzianum group and the T. Longibrachiatum group under the CK and T2 drought treatments. Under the T1 and T3 drought treatments, the leaf biomass of the T. Longibrachiatum group was slightly higher than that of the T. Harzianum group (Fig. 6a). The leaf biomass of the Control group increased significantly under the T1 drought treatment. Additionally, the leaf biomass of the C. migao seedlings inoculated with T. Harzianum and T. Longibrachiatum was generally higher than that of the Control group. The root and stem biomass of the inoculated groups was also generally higher than that of the Control group (Fig. 6b,c). Furthermore, the root and stem biomass of the T. Longibrachiatum group was higher than that of the T. Harzianum group under all drought treatments except CK. In terms of total biomass, the total biomass of the T. Harzianum group and the T. Longibrachiatum group was generally higher than that of the Control group (Fig. 6d). The difference in total biomass between the T. Longibrachiatum group and the Control group was greatest under the T2 drought treatment, while the Control group showed a significant increase under the T3 drought treatment.

Effects of endophytic fungi inoculation on biomass of C. migao seedlings under different drought stress.
The chlorophyll a content in both the T. longibrachiatum group and the Control group gradually decreased, while the T. harzianum group exhibited a trend of initial decrease, followed by an increase, and then another decrease. Under drought stress conditions, the chlorophyll a content in both inoculated groups (T. harzianum and T. longibrachiatum) was generally higher than that in the Control group. In the CK and T1 drought treatments, the chlorophyll a content in the T. longibrachiatum group was slightly higher than that in the T. harzianum group, whereas under the T3 treatment, it was lower than that in the T. harzianum group. Notably, under the T2 treatment, the chlorophyll a content in the T. harzianum group was 127.88% higher than that in the T. longibrachiatum group. The trend of chlorophyll b content in the T. longibrachiatum and Control groups was similar to that of chlorophyll a. However, in the T. harzianum group, chlorophyll b displayed an increase–decrease-increase pattern, which was opposite to chlorophyll a, reaching its peak (1.47 mg/g) under the T1 treatment. Under the CK treatment, the chlorophyll b content in the Control, T. harzianum, and T. longibrachiatum groups was nearly identical (1.09 mg/g, 1.09 mg/g, and 1.07 mg/g, respectively). Similarly, under the T2 treatment, the values for T. harzianum and T. longibrachiatum were almost equal (0.76 mg/g and 0.75 mg/g, respectively). In contrast, under the T1 and T3 treatments, the chlorophyll b content followed the order: T. harzianum group > T. longibrachiatum group > Control group*. Except for CK, the chlorophyll b content in both inoculated groups was significantly higher than that in the Control group. For chlorophyll (a + b), under all water treatments except CK, the values ranked as: T. harzianum group > T. longibrachiatum group > Control group*. Overall, the inoculation of endophytic fungi helped mitigate the reduction in photosynthetic pigments, benefiting the growth and development of C. migao seedlings—with T. harzianum demonstrating a more pronounced effect (Fig. 7).

Effects of endophytic fungi inoculation on osmolytes content of C. migao seedlings under different drought stress.
Effects of endophytic fungi on osmotic substance content in C. migao seedlings under drought stress
The soluble sugar content in the three inoculation groups—T. harzianum, T. longibrachiatum, and Control—reached their maximum values under the T2 drought treatment, measuring 232.4242 (μg/g FW), 312.8006 (μg/g FW), and 155.705 (μg/g FW), respectively. Additionally, under all drought treatments, the soluble sugar content in the T. longibrachiatum group was higher than that in the T. harzianum group (Fig. 8a). The trends in soluble protein content for the T. harzianum and T. longibrachiatum groups were consistent with those of soluble sugar content (Fig. 8b). The free proline content in the T. harzianum group peaked under the T2 drought treatment at 82.3334 (μg/g FW), while the largest difference between the T. longibrachiatum and Control groups occurred under the T3 drought treatment, with free proline contents of 81.3336 (μg/g FW) and 58.500 (μg/g FW), respectively (Fig. 8c). As these substances increase, the cytoplasmic osmotic pressure rises, thereby improving cellular water absorption capacity and osmotic regulation. Consequently, this enhances the drought resistance of C. migao seedlings.

Effects of endophytic fungi inoculation on antioxidant enzyme activities of C. migao seedlings under different drought stress.
Effects of endophytic fungi on antioxidant enzyme activities of C. migao seedlings under drought stress
The CAT activity in the leaves of C. migao seedlings in the three inoculated groups exhibited an initial increase followed by a decline. The Control group reached its maximum value of 60.753 U/g under the T1 drought treatment. The T. harzianum group and T. longibrachiatum group both peaked under the T2 drought treatment, with values of 112.803 U/g and 92.698 U/g, respectively. The CAT activity in the T. longibrachiatum group was slightly lower than that in the T. harzianum group under the CK and T1 drought treatments, whereas the T. harzianum group showed significantly higher CAT activity than the T. longibrachiatum group under the T2 and T3 drought treatments. The trend of POD activity in the leaves of C. migao seedlings was similar to that of CAT (Fig. 9a,b). The SOD activity in the leaves of C. migao seedlings in the T. harzianum, T. longibrachiatum, and Control groups followed the same trend, peaking under the T2 drought treatment at 186.918 U/g, 261.422 U/g, and 153.768 U/g, respectively. Overall, the SOD activity in the T. longibrachiatum-inoculated seedlings was consistently higher than that in the T. harzianum and Control groups (T. longibrachiatum > T. harzianum > Control) (Fig. 9c). These results indicate that inoculation with T. harzianum and T. longibrachiatum can enhance antioxidant enzyme activity, maintain intracellular redox balance, and improve the seedlings’ ability to cope with drought-induced damage, with T. longibrachiatum exhibiting a more pronounced effect. In the T. longibrachiatum group, the MDA content reached its maximum value of 67.819 nmol/g under the T2 drought treatment, which was higher than that in the T. harzianum group. However, under the T3 drought treatment, it was lower than in the T. harzianum group. Additionally, across all water treatments, the MDA content in the inoculated groups was significantly lower than that in the Control group (Fig. 9d).

Effects of endophytic fungi inoculation on photosynthetic pigments in leaves of C. migao under different drought stress.
Effects of endophytic fungi on endogenous hormones in C. migao seedlings under drought stress
The IAA content in C. migao seedlings varied differently among inoculation groups as drought stress intensified. The Control group exhibited an “increase–decrease-increase” trend, reaching a maximum value of 253.082 ng/g under the T1 drought treatment. The T. harzianum group showed an initial decrease followed by an increase, while the T. longibrachiatum group displayed a “decrease-increase–decrease” trend. Both T. harzianum and T. longibrachiatum groups reached their highest IAA levels under the CK treatment (241.0887 ng/g and 252.8910 ng/g, respectively). However, the T. longibrachiatum group experienced a sharp decline in IAA content under T3 stress (98.8491 ng/g). Notably, the T. harzianum group had lower IAA levels than the Control group in all treatments except CK (Fig. 10). For ABA content, the T. harzianum group first decreased and then increased with intensifying drought, whereas the T. longibrachiatum group showed a continuous decline. The Control group exhibited a steady increase. Both fungal-inoculated groups peaked under CK conditions (T. harzianum: 610.2788 ng/g; T. longibrachiatum: 510.3348 ng/g), while the Control group reached its maximum (467.3644 ng/g) under T3. Importantly, the T. harzianum group maintained higher ABA levels than both T. longibrachiatum and Control groups across all treatments (Fig. 10b). In terms of GA₃ content, the Control group peaked at 304.3620 ng/g under T3—significantly higher than the inoculated groups (T. harzianum: 220.1840 ng/g; T. longibrachiatum: 163.9831 ng/g). By contrast, both fungal groups achieved their highest GA₃ levels under CK (T. harzianum: 258.7352 ng/g; T. longibrachiatum: 263.1539 ng/g), surpassing the Control group (Fig. 9d). The ZR content in the T. longibrachiatum group correlated positively with water availability. Except under T2, the Control group consistently showed higher ZR levels than both inoculated groups. The T. harzianum group outperformed T. longibrachiatum under T1 and T2 but showed no significant difference under T3 (Fig. 10d).

Effects of endophytic fungi inoculation on endogenous hormones in C. migao seedlings under different drought stress.
Correlation analysis between endophytic fungi and the growth physiology of C. migao seedlings under drought conditions, and their relationship with enzyme and endogenous hormone content changes
The correlation analysis and Redundancy Analysis (RDA) were conducted on the growth indicators (plant height, ground diameter, leaf weight, stem weight, root weight, and whole plant weight) of inoculated and control groups of C. migao seedlings under different drought treatments, along with related enzymes, hormones, and photosynthetic pigment content. The results revealed significant positive correlations between Height and ZR, Leaf weight and SOD, Culm weight and SOD, Root dry weight and POD, Root dry weight and SOD, Whole plant weight and POD, Whole plant weight and SOD, and Whole plant weight and ZR (Fig. 11a). This suggests that appropriately increasing the content of these enzymes, hormones, and photosynthetic pigments can promote the growth and development of C. migao, thereby enhancing biomass accumulation.

RDA analysis and correlation analysis of physiological and biochemical indexes of C. migao seedlings. AS: soluble sugar content, AP: soluble protein content, Apro: free proline content, CAT: catalase activity, POD: peroxidase activity, SOD: superoxide dismutase activity, MDA: malondialdehyde content, GA3: gibberellin content, ZR: zeatin nucleoside content, RWC: relative water content.
It can be observed that the plant height, ground diameter, leaf weight, stem weight, and root weight of C. migao seedlings are strongly correlated with the inoculated group under moderate and severe drought stress. Meanwhile, the content of chlorophyll a, chlorophyll b, chlorophyll (a + b), and RWC shows a stronger correlation with the inoculated group under mild drought and control conditions. This analysis further indicates that inoculation has a significant impact on C. migao seedlings under different drought stress conditions (Fig. 11b).
Discussion
Upon exposure to drought stress, plants activate a suite of coordinated responses spanning morphological, physiological, and molecular levels to ensure survival. This study demonstrates that inoculation with T. harzianum and T. longibrachiatum significantly enhances the host plant’s resilience to drought stress. This enhanced tolerance originates from the systemic regulation exerted by Trichoderma on root system architecture, osmotic adjustment, antioxidant defense, photosynthetic apparatus, and the endogenous phytohormone network.
Trichoderma enhances water foraging capacity by remodeling root system architecture
Under drought stress, plants often allocate more photoassimilates to roots to optimize root architecture for accessing deeper soil moisture. In this study, both Trichoderma strains promoted root elongation and increased the proportion of coarse roots, representing a classic adaptive strategy to water deficit. The underlying mechanism likely involves the secretion of bioactive compounds such as indole-3-acetic acid (IAA) and ACC deaminase by Trichoderma, which directly stimulate cell division in the root apical meristem and cell expansion in the elongation zone21,22. T. harzianum may preferentially induce lateral root formation to expand the absorptive surface area, whereas T. longibrachiatum might be more effective in promoting primary root penetration. By employing distinct strategies to optimize the spatial distribution of roots (root system architecture), both fungi significantly enhance the plant’s “water foraging” capacity under drought conditions, providing fundamental support for maintaining aboveground physiological activities.
Trichoderma maintains cellular homeostasis by reinforcing osmotic adjustment and antioxidant systems
Under mild to moderate drought stress, Trichoderma-inoculated plants accumulated higher levels of proline, soluble sugars, and soluble proteins. This is not merely a passive response but an active physiological process driven by the Trichoderma-mediated activation of key osmotic adjustment gene pathways, such as P5CS, a critical gene in proline biosynthesis23,24. The coordinated accumulation of these osmolytes effectively lowers cellular osmotic potential, facilitates water uptake, and serves as compatible solutes to protect macromolecular structures.
Concurrently, oxidative stress induced by drought is a primary cause of cellular damage. Our results confirm that both Trichoderma species significantly enhance the activities of key antioxidant enzymes—superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT)—forming an efficient enzymatic scavenging system against reactive oxygen species (ROS). T. harzianum might more potently induce SOD activity, accelerating the dismutation of superoxide anion to hydrogen peroxide (H₂O₂), while T. longibrachiatum could be more focused on enhancing the detoxification capacity of POD and CAT towards H₂O₂. This synergistic interaction collectively mitigates membrane lipid peroxidation and maintains the structural integrity of cell membranes, which is crucial for the sustained physiological activity observed in Trichoderma-treated plants under drought25,26.
However, under severe drought (T3 treatment), the decline in osmolyte content indicates that the plant’s physiological threshold has been exceeded. At this stage, severe cellular dehydration and oxidative damage likely impair mitochondrial function, leading to disrupted ATP synthesis27,28. Consequently, energy-dependent biosynthetic processes for osmolyte accumulation fail. This suggests a limit to the mitigating effects of Trichoderma, although the more robust root system and stabilized cellular environment established under mild to moderate stress undoubtedly delay the onset of this collapse.
Trichoderma sustains carbon assimilation via multi-faceted protection of the photosynthetic apparatus
Chlorophyll stability is a prerequisite for photosynthetic performance. In this study, Trichoderma inoculation alleviated drought-induced chlorophyll degradation, which can be explained by several mechanisms. Firstly, the enhanced antioxidant system protects chloroplast membranes from ROS attack, reducing photo-oxidative degradation of chlorophyll. Secondly, evidence suggests Trichoderma can upregulate the expression of photosynthesis-related genes and may secrete hormones like cytokinins (CK) to suppress the activity of chlorophyll-degrading enzymes, directly delaying chlorophyll breakdown29. Finally, improved root function and subsequent better plant water status provide the most fundamental support for photosynthesis. Thus, Trichoderma ensures the maintenance of photosynthetic carbon assimilation under drought through a synergistic "Protection (antioxidation)—Maintenance (degradation inhibition)—Support (water supply)" model.
Trichoderma integrates stress responses by modulating hormonal homeostasis
The regulation of drought tolerance in plants is a highly integrated process, wherein endogenous phytohormones act as central regulators. Our preliminary findings indicate that Trichoderma inoculation alters the balance between auxin (IAA) and abscisic acid (ABA). We hypothesize that Trichoderma, on one hand, promotes root growth under drought by producing IAA or modulating host IAA metabolism30. On the other hand, it may fine-tune stomatal aperture by regulating ABA biosynthesis or metabolism, thereby achieving an optimal trade-off between reducing water loss (water conservation) and maintaining CO₂ uptake (carbon assimilation)31. This precise modulation of hormonal signaling is central to the systemic enhancement of drought tolerance by Trichoderma, effectively linking root absorption capacity, leaf stomatal behavior, and cellular stress defense mechanisms.
Conclusion
This study systematically evaluated the physiological mechanisms by which two endophytic fungi, T. harzianum and T. longibrachiatum, enhance drought tolerance in C. migao seedlings under varying water gradients. Our results demonstrate that both Trichoderma species significantly improved seedling drought resistance through multiple interconnected physiological pathways. Specifically, fungal inoculation promoted plant growth and root absorptive capacity. It lowered leaf water potential, thereby increasing the soil–plant water potential gradient, which enhanced leaf relative water content and overall plant water status. Physiologically, inoculation stimulated the accumulation of osmolytes, aiding in turgor maintenance. Concurrently, it significantly boosted the activity of key antioxidant enzymes (SOD, POD, CAT), facilitating reactive oxygen species (ROS) scavenging and reducing malondialdehyde (MDA) content, thereby preserving membrane integrity. Furthermore, Trichoderma inoculation modulated endogenous hormone homeostasis by promoting abscisic acid (ABA) accumulation to initiate drought stress signaling, while reducing indole-3-acetic acid (IAA), gibberellin (GA₃), and zeatin riboside (ZR) levels. This hormonal reprogramming likely optimizes resource allocation under stress by curbing growth and reducing water consumption. In summary, T. harzianum and T. longibrachiatum enhance the adaptability of C. migao seedlings to drought through synergistic "morpho-physio-biochemical" modifications. This research provides a theoretical foundation and practical prospects for utilizing these endophytic fungi as bio-inoculants to improve afforestation success in arid and semi-arid regions.
Data availability
Trichoderma harzianum (Accession Number: PX-916279) and Trichoderma longibrachiatum (Accession Number: PX-916280) sequences have been deposited in the GenBank database of the National Center for Biotechnology Information (NCBI), under the BioProject accession number SUB15966526.
References
Chen, J. et al. Effects of rhizosphere fungi on the chemical composition of fruits of the medicinal plant Cinnamomum migao endemic to Southwestern China. BMC Microbiol. 21(1), 206. https://doi.org/10.1186/s12866-021-02216-z (2021).
Wu, W. et al. Ethanol extracts of Cinnamomum migao H.W. Li attenuates neuroinflammation in cerebral ischemia-reperfusion injury via regulating TLR4-PI3K-Akt-NF-κB pathways. J. Ethnopharmacol. 339, 119150. https://doi.org/10.1016/j.jep.2024.119150 (2025).
Chen, J. Z. et al. Seed dormancy release and germination requirements of Cinnamomum migao, an endangered and rare woody plant in Southwest China. Front. Plant Sci. 13, 770940. https://doi.org/10.3389/fpls.2022.770940 (2022).
Muhammad, I. et al. An extensive pharmacological evaluation of novel anti-nociceptive and IL-6 targeted anti-inflammatory guaiane-type sesquiterpenoids from Cinnamomum migao H. W. Li through in-depth in-vitro, ADMET, and molecular docking studies. Biomed. Pharmacother. 164, 114946. https://doi.org/10.1016/j.biopha.2023.114946 (2023).
Sun, Q. W., Chen, J. Z., Liao, X. F., Huang, X. L. & Liu, J. M. Identification of keystone taxa in rhizosphere microbial communities using different methods and their effects on compounds of the host Cinnamomum migao. Sci. Total Environ. 926, 171952. https://doi.org/10.1016/j.scitotenv.2024.171952 (2024).
Xiao, X. et al. Different arbuscular mycorrhizal fungi established by two inoculation methods improve growth and drought resistance of Cinnamomum migao seedlings differently. Biology https://doi.org/10.3390/biology11020220 (2022).
Huang, X. et al. Transcriptome analysis of Cinnamomum migao seed germination in medicinal plants of Southwest China. BMC Plant Biol. 21(1), 270. https://doi.org/10.1186/s12870-021-03020-7 (2021).
Tiwari, R. K. et al. Mechanistic insights on melatonin-mediated drought stress mitigation in plants. Physiol. Plant. 172(2), 1212–1226. https://doi.org/10.1111/ppl.13307 (2021).
Yu, C., Jiang, X., Xu, H. & Ding, G. Trichoderma longibrachiatum inoculation improves drought resistance and growth of Pinus massoniana seedlings through regulating physiological responses and soil microbial community. J. Fungi https://doi.org/10.3390/jof9070694 (2023).
Lata, R., Chowdhury, S., Gond, S. K. & White, J. F. Jr. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 66(4), 268–276. https://doi.org/10.1111/lam.12855 (2018).
Almario, J., Fabiańska, I., Saridis, G. & Bucher, M. Unearthing the plant-microbe quid pro quo in root associations with beneficial fungi. New Phytol. 234(6), 1967–1976. https://doi.org/10.1111/nph.18061 (2022).
Zhang, X. et al. Sustainable remediation of soil and water utilizing arbuscular mycorrhizal fungi: A review. Microorganisms https://doi.org/10.3390/microorganisms12071255 (2024).
Shi, J., Wang, X. & Wang, E. Mycorrhizal symbiosis in plant growth and stress adaptation: From genes to ecosystems. Annu. Rev. Plant Biol. 74, 569–607. https://doi.org/10.1146/annurev-arplant-061722-090342 (2023).
Bashyal, B. M., Parmar, P., Zaidi, N. W. & Aggarwal, R. Molecular programming of drought-challenged Trichoderma harzianum-bioprimed rice (Oryza sativa L.). Front. Microbiol. 12, 655165. https://doi.org/10.3389/fmicb.2021.655165 (2021).
Zhou, Q. et al. Effects of different drought degrees on physiological characteristics and endogenous hormones of soybean. Plants https://doi.org/10.3390/plants11172282 (2022).
Karimi, F. et al. An applicable method for extraction of whole seeds protein and its determination through Bradford’s method. Food Chem. Toxicol. 164, 113053. https://doi.org/10.1016/j.fct.2022.113053 (2022).
Krauspe, R. & Scheer, A. Coomassie brilliant blue G-250 dye-binding technique for determination of autolytic protein breakdown in Euglena gracilis and comparison to other methods of autolysis measurement. Anal. Biochem. 153(2), 242–250. https://doi.org/10.1016/0003-2697(86)90088-6 (1986).
Chen, S. B. et al. High-performance thin-layer chromatographic fingerprints of isoflavonoids for distinguishing between Radix Puerariae Lobate and Radix Puerariae Thomsonii. J. Chromatogr. A 1121(1), 114–119. https://doi.org/10.1016/j.chroma.2006.04.082 (2006).
Zhou, L. N. et al. [Protective effect of different solvent extracts from platycladi cacumen carbonisatum on LPS-induced human umbilical vein endothelial cells damage]. Zhongguo Zhong Yao Za Zhi 38(22), 3933–3938 (2013).
Jakl, M., Ćavar Zeljković, S., Kovač, I., Bělonožníková, K. & Jaklová Dytrtová, J. Side effects of triazoles on treated crops. Chemosphere 277, 130242. https://doi.org/10.1016/j.chemosphere.2021.130242 (2021).
Brotman, Y. et al. Trichoderma-plant root colonization: Escaping early plant defense responses and activation of the antioxidant machinery for saline stress tolerance. PLoS Pathog. 9(3), e1003221. https://doi.org/10.1371/journal.ppat.1003221 (2013).
Contreras-Cornejo, H. A., Macías-Rodríguez, L., Beltrán-Peña, E., Herrera-Estrella, A. & López-Bucio, J. Trichoderma-induced plant immunity likely involves both hormonal- and camalexin-dependent mechanisms in Arabidopsis thaliana and confers resistance against necrotrophic fungi Botrytis cinerea. Plant Signal. Behav. 6(10), 1554–1563. https://doi.org/10.4161/psb.6.10.17443 (2011).
Mastouri, F., Björkman, T. & Harman, G. E. Seed treatment with *Trichoderma harzianum* alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 100(11), 1213–1221. https://doi.org/10.1094/phyto-03-10-0091 (2010).
Shukla, N., Awasthi, R. P., Rawat, L. & Kumar, J. Biochemical and physiological responses of rice (*Oryza sativa* L.) as influenced by *Trichoderma harzianum* under drought stress. Plant Physiol. Biochem. 54, 78–88. https://doi.org/10.1016/j.plaphy.2012.02.001 (2012).
Harman, G. E., Howell, C. R., Viterbo, A., Chet, I. & Lorito, M. Trichoderma species–Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2(1), 43–56. https://doi.org/10.1038/nrmicro797 (2004).
Mastouri, F., Björkman, T. & Harman, G. E. Trichoderma harzianum enhances antioxidant defense of tomato seedlings and resistance to water deficit. Mol. Plant-Microbe Interact. 25(9), 1264–1271. https://doi.org/10.1094/mpmi-09-11-0240 (2012).
Møller, I. M., Jensen, P. E. & Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 58, 459–481. https://doi.org/10.1146/annurev.arplant.58.032806.103946 (2007).
Wang, X. et al. Metabolic adaptation of wheat grains contributes to a stable filling rate under heat stress. J. Exp. Bot. 69(22), 5531–5545. https://doi.org/10.1093/jxb/ery303 (2018).
Contreras-Cornejo, H. A., Macías-Rodríguez, L., Alfaro-Cuevas, R. & López-Bucio, J. Trichoderma spp. improve growth of Arabidopsis seedlings under salt stress through enhanced root development, osmolite production, and Na⁺ elimination through root exudates. Mol. Plant-Microbe Interact. 27(6), 503–514. https://doi.org/10.1094/mpmi-09-13-0265-r (2014).
Contreras-Cornejo, H. A., Macías-Rodríguez, L., Cortés-Penagos, C. & López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 149(3), 1579–1592. https://doi.org/10.1104/pp.108.130369 (2009).
Zhang, S., Gan, Y. & Xu, B. Mechanisms of the IAA and ACC-deaminase producing strain of Trichoderma longibrachiatum T6 in enhancing wheat seedling tolerance to NaCl stress. BMC Plant Biol. 19(1), 22. https://doi.org/10.1186/s12870-018-1618-5 (2019).
Funding
This research was funded by the Guizhou Provincial Basic Research Programs (Natural Science) (QKH-ZK [2022276), Guizhou Provincial forestry Research Program (QLKH [202503).
Author information
Authors and Affiliations
Contributions
Tao Fu: Conceptualization, Methodology, Software. Lan-yue Zhang: Data curation, Writing—original draft. Shuang-yan Hu: Visualization, Investigation. Jing-zhong Chen: Supervision.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, Ly., Fu, T., Chen, Jz. et al. Two Trichoderma endophytic fungi alleviates drought stress and improves plant growth in Cinnamomum migao seedlings. Sci Rep 16, 14261 (2026). https://doi.org/10.1038/s41598-026-44455-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-44455-y
