
Abstract
Fukui Cave, located in the southwestern Japanese Archipelago, is a cave site containing cultural layers dating from the Late Pleistocene to the early Holocene. This study aimed to identify the animal taxa of burned bone fragments excavated from Layer IV of the cave, dated to approximately 16,000 years ago, using a non-destructive histomorphological approach. Because faunal remains are extremely scarce at Palaeolithic sites in the Japanese Archipelago, these specimens provide important evidence for understanding the relationship between humans and animals during the Late Pleistocene. However, all excavated bones were burned (calcined) fragments less than 1 cm in length, making macroscopic taxonomic identification difficult. To address this, synchrotron radiation X-ray computed tomography (voxel size: 2.74 μm) was performed at SPring-8 to analyze the internal bone microstructure. As a result, secondary osteons were identified in three of the seven burned fragments, and plexiform bone in one. The cross-sectional areas of osteons and Haversian canals were measured and compared statistically with reference data from various mammalian taxa, taking into account possible shrinkage due to burning. The results indicated that these fragments all fall within the range of medium-sized artiodactyls, such as deer or wild boar. In contrast, derivation from large mammals such as Naumann’s elephant or Yabe’s giant deer, which were extinct in the Late Pleistocene, can be excluded. These findings demonstrate the effectiveness of non-destructive histomorphological identification using high-resolution CT for burned bone fragments and provide new insights into animal exploitation by Late Pleistocene humans in the Japanese Archipelago.
Similar content being viewed by others

Introduction
Burned bone fragments from the Late Pleistocene at Fukui Cave
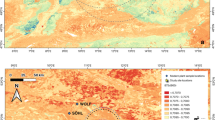
Fukui Cave (33°17′21″ N, 129°42′36″ E) is located in Sasebo City, Nagasaki Prefecture, in the southwestern part of the Japanese Archipelago (Fig. 1), and has long been recognized as an archaeological site spanning the Late Pleistocene to the earliest Holocene, culturally from the Paleolithic to the emergence of Jomon pottery1,2,3,4. In recognition of its high archaeological significance, the cave was designated a Special Historic Site by the Agency for Cultural Affairs of the Government of Japan in 20245.

Fukui Cave and Paleolithic sites in the Japanese Archipelago with recovered animal bones.
Excavations conducted between 2011 and 2015 uncovered a number of burned bone fragments in Layer 4 (Fig. 2), a terminal Paleolithic deposit (ca. 16 ka BP)6. The excavated area of Layer 4 measured 1.12 m², and the burned bones were concentrated in a small portion of this space. The curator from the Sasebo City Board of Education who authored the excavation report suggested that a hearth may have existed near the find spot of the burned bones. Based on their size and bone tissue characteristics, the fragments were thought to represent compact bone from terrestrial medium and/or large-sized mammals larger than small mammals such as rodents.

Burned bone fragments from Fukui Cave (left to right: FK-B-001, FK-B-002 … FK-B-007). Scale bar = 5 mm.
The burned bone fragments exhibited a gray coloration due to burning, with some showed a white coloration. Considering the observed whitening and the inferred exposure to high temperatures (see the Materials section), the materials examined in this study are generally regarded as calcined bone, which is exposed to temperatures above approximately 550°C7,8,9,10. Only these calcined bone fragments were recovered from this stratigraphic layer, and no unburned bones were found. In general, bone remains were scarce in the excavation of Fukui cave6, suggesting that the depositional environment was not favorable for bone preservation. However, exposure to high temperatures likely resulted in the complete combustion of organic components such as collagen and increased the crystallinity of bioapatite7,8,11,12,13, thereby making the bone more resistant to microbial decomposition and chemical dissolution.
It is generally accepted that human groups began to inhabit the Japanese Archipelago in the Late Pleistocene, around 40–35 ka BP14,15. The subsequent Paleolithic period, which lasted until approximately 15 ka BP, yielded more than 14,000 sites16. However, many of these sites are situated in acidic geological environments derived from volcanic deposits, and, combined with the warm and humid climate that promotes the decomposition of faunal remains, animal bones are only rarely preserved15,17,18,19. In the Japanese Archipelago, only six Paleolithic sites yielded animal remains: the Kashiwadai 1 site (ca. 26–27 ka BP, medium-sized cervids)18; Shitsukari-Abe Cave (ca. 20–39 ka BP, hare, Japanese serow, moose, brown bear)20,21; the Hanaizumi site (ca. 26 ka BP, bison, Yabe’s giant deer, moose, Naumann’s elephant)22,23; and the C-Area of the Yoshioka site group (ca. 25 ka BP, wild boar)24, Gyoja-ana cave site (ca. 20 ka BP and 27–28 ka BP, medium-sized mammals)25,26, and the Nojiriko-Tategahana lake site (ca. 54–38 ka BP, Naumann’s elephant, Yabe’s giant deer, moose)27,28.
Consequently, the actual nature of animal exploitation by Late Pleistocene humans in the Japanese Archipelago remains poorly understood. By the end of the Late Pleistocene, several large mammalian species, including elephants and Yabe’s giant deer, had gone extinct in the archipelago29,30,31,32,33. Whether these extinction events were related to human activity during the Paleolithic period remains unresolved15,19.
In this context, the burned bone fragments from the terminal Paleolithic of Fukui Cave are of considerable importance for understanding human–animal relationships in the Late Pleistocene of the Japanese Archipelago. Nevertheless, the fragments are heavily degraded, and all measure less than 1 cm in maximum length, making taxonomic identification through macroscopic examination extremely difficult.
Histomorphological taxonomic identification of small bone fragments
Taxonomic identification of animal bones from archaeological sites represents one of the most fundamental approaches for investigating past human use of animals. Standard methods of taxonomic identification generally rely on macroscopic morphological observation of specimens34. However, for highly fragmented bone pieces with little or no remaining macroscopic anatomical features, histomorphological approaches have proven effective35,36,37,38,39,40.
Mammalian bones are fundamentally composed of two types of bone tissue, namely spongy (cancellous) bone and compact (cortical) bone. Compact bone typically contains numerous vascular canals. In many mammals, particularly in the diaphyses of long bones, the compact bone is predominantly composed of osteons (secondary osteons, Haversian systems), which consist of longitudinally oriented canals (Haversian canals) surrounded by concentric lamellae, except in very young individuals. The size of osteons and Haversian canals are influenced by multiple factors, including body mass, lifestyle, and locomotor ability41,42, but interspecific differences have long been recognized43,44. In non-human animals, osteon banding, in which osteons appear in linear arrangements, is sometimes observed40,45. In addition, in certain mammals—particularly artiodactyls such as wild boars, deer, and cattle, perissodactyls such as horses, and carnivores such as dogs and bears—the compact bone of the limbs often exhibits a prominent development of plexiform bone, a distinctive structure in which vascular channels are arranged in a brick-like pattern and surrounded by lamellar bone46,47,48,49,50,51,52,53. Although plexiform bone can appear in human infants and children54,55,56, it has not been reported in adult human bone.
Based on accumulated comparative histomorphological knowledge, the presence or absence of osteon banding or plexiform bone, as well as the size of osteons and Haversian canals, provides an effective means of identifying animal taxa. Although the method has inherent limitations in accuracy and applicability, histomorphological analysis has proven reliable for taxonomic identification40,43,55,57,58,59,60,61,62. In the case of highly fragmented burned bones, such as those from Fukui Cave, the application of biochemical identification methods, including DNA analysis or Zooarchaeology by Mass Spectrometry (ZooMS), is generally challenging due to high-temperature burning, which degrades DNA and collagen and prevents molecular identification63,64. Under these circumstances, histomorphological approaches currently represent the most practical method for taxonomic identification.
Conventional histomorphological analysis typically requires thin-sectioning of bone fragments and preparation of microscope slides, which entails partial destruction of the specimens. When materials are scarce or when preservation of cultural heritage precludes destructive analysis, non-destructive histomorphological approaches are required. Computed tomography (CT) enables the non-destructive observation of bone microstructure. While medical and industrial CT scanners typically achieve voxel resolutions of several hundred micrometers, osteons in most mammals, including humans, are generally less than 500 μm in diameter, and Haversian canals are even smaller40,65,66,67,68,69. Therefore, high-resolution X-ray CT is necessary for the detailed investigation of bone microstructure70.
Objective of this study: non-destructive histomorphological taxonomic identification using high-resolution X-ray CT
In this study, we attempted the histomorphological identification of taxonomic group from Late Pleistocene burned bone fragments from Fukui Cave by non-destructively imaging their bone microstructure using synchrotron radiation X-ray CT (SR X-ray CT) at SPring-8, a large synchrotron radiation facility. SPring-8, located in Sayo Town, Hyogo Prefecture, Japan, is capable of generating world-leading synchrotron radiation and operates as a joint-use facility open to researchers from academia and industry both domestically and internationally. Using the CT at this facility, we obtained high-resolution images of the bone microstructure of the Fukui Cave fragments at a voxel size of 2.74 μm.
As prior studies utilizing CT images for histomorphological taxonomic identification of archaeological bones, Bradfield71 identified the raw materials of worked bone artifacts from the Middle Stone Age in South Africa, and in forensic science, Andronowski et al.72 examined methods for histomorphological differentiation between human and bear metacarpals. However, such studies remain scarce, and no research has yet applied this approach to tiny fragments of burned bone, such as those from Fukui Cave. Establishing this method as a practical and reliable tool requires the accumulation of empirical results. This paper demonstrates that high-resolution X-ray CT is an effective approach for non-destructive taxonomic identification of highly fragmented burned bone. Furthermore, based on the identification results obtained in this study, we aim to provide new insights into the exploitation of animals by Late Pleistocene humans in the Japanese Archipelago.
Results
CT observations of the burned bone fragments from Fukui Cave
Among the seven burned bone fragments (Specimen IDs: FK-B-001 to FK-B-007) examined by CT, specimens FK-B-002, FK-B-003, and FK-B-006 showed osteons (secondary osteons) distributed across the observable compact bone, constituting its predominant component (Fig. 3). Haversian canals were clearly visible at the centers of osteons, but Haversian lamellae and lacunae could not be observed because they were beyond the resolution limits of the CT. FK-B-007 exhibited plexiform bone throughout the observable area (Fig. 3). No pathological features, such as abnormally enlarged canals indicative of osteoporosis, were observed in FK-B-002, FK-B-003, FK-B-006, or FK-B-007, suggesting that all retained normal histological characteristics. FK-B-001, FK-B-004, and FK-B-005 displayed structures resembling Haversian canals, but the outlines of osteons were indistinct, making it difficult to clearly identify bone microstructures.

CT images of burned bones from Fukui Cave. Left: osteons in specimen FK-B-003; right: plexiform bone structure in specimen FK-B-007. Arrows point to osteons. Scale bar = 200 μm.
Histomorphometric values
Among the calcined bone fragments from Fukui Cave, the cross-sectional area of complete osteons (On.Ar) and the cross-sectional area of the Haversian canal within each osteon (H.Ar) were measured for FK-B-002, FK-B-003, and FK-B-006, in which osteons were identified. Taking into account the estimated burning temperatures of 500–700 °C, Table 1 presents the values under two scenarios: (1) no shrinkage at 500–600 °C (corresponding to the lower bound in the Methods section), and (2) shrinkage at 700 °C (corresponding to the upper bound).
The On.Ar ranges (lower bound–upper bound) were as follows: FK-B-002, 13,743.9 (± 2,766.0) to 18,325.2 (± 3,687.9) µm²; FK-B-003, 18,174.1 (± 5,226.5) to 24,232.1 (± 6,968.7) µm²; and FK-B-006, 15,936.1 (± 3,689.1) to 21,248.2 (± 4,918.8) µm². The H.Ar ranges were: FK-B-002, 490.5 (± 190.6) to 654.0 (± 254.1) µm²; FK-B-003, 478.7 (± 186.2) to 638.3 (± 248.3) µm²; and FK-B-006, 500.5 (± 370.7) to 667.3 (± 494.2) µm² (values represent mean ± SD). The actual On.Ar and H.Ar values are assumed to lie within these ranges.
The mean values and standard errors of On.Ar and H.Ar for the comparative faunal specimens are presented in Table 2. Since animals belonging to the same order and of similar body size tended to exhibit comparable On.Ar and H.Ar values, the comparative specimens were grouped according to order-level taxonomy and body size: proboscideans (Naumann’s elephant, fossil elephant, Asian elephant), human, medium-sized primates (macaque), lagomorphs (leporid), medium-sized artiodactyls (wild boar, sika deer, ancient sika deer, reindeer, Japanese serow), large-sized artiodactyls (Yabe’s giant deer, cattle, bison), medium-sized carnivores (Japanese marten, dog, raccoon dog, red fox, leopard), and large-sized carnivores (brown bear). A bivariate plot was then generated with On.Ar on the X-axis and H.Ar on the Y-axis, in which the ranges of each animal group were represented by rectangles defined by the minimum and maximum mean values of On.Ar and H.Ar calculated across individual specimens within each group (Fig. 4).

Ranges of mean On.Ar and H.Ar values for the Fukui Cave specimens and comparative mammalian groups, based on histomorphometric data in Tables 1 and 2. The Fukui Cave specimens are, from left to right, FK-B-002, FK-B-006, and FK-B-003. The lower-left corners of the Fukui Cave range indicate values estimated for burning temperatures of 500–600 °C, and the upper-right corners indicate those estimated for 700 °C.
When the ranges from the lower to upper bounds of On.Ar and H.Ar for FK-B-002, FK-B-003, and FK-B-006 from Fukui Cave were indicated on the graph as rectangles, all three specimens fell within the range of medium-sized artiodactyls, and were next close to the ranges of carnivores. In contrast, the values for humans and proboscideans were clearly distant from those of the Fukui Cave burned bones. Among the large artiodactyls, the values for bovids (cattle and bison) were relatively close to the Fukui Cave range, whereas Yabe’s giant deer, which became extinct in the Pleistocene, lay farther away (Table 2; Fig. 4).
Statistical results
Assuming that no shrinkage of the bone tissue structure occurred at a burning temperature of 500–600 °C, significant differences (p < 0.05), determined by Games–Howell tests, in either or both On.Ar and H.Ar were detected between FK-B-002, FK-B-003, and FK-B-006, and the following comparative specimens: Naumann’s elephant, fossil elephant, Asian elephant, human, macaque, sika deer (one of seven specimens), Yabe’s giant deer, cattle (three of five specimens), red fox, and brown bear (one of four specimens) (Table 3). Additional significant differences were also observed between each of FK-B-003 and FK-B-006 and Japanese serow and brown bear (one of the remaining three specimens), as well as between FK-B-003 and leporid, wild boar (one of four specimens), sika deer (one of the remaining six specimens), bison, Japanese marten, dog (one of two specimens), and brown bear (one of the remaining two specimens) (Table 3).
Assuming that shrinkage of the bone tissue structure occurred at a burning temperature of 700 °C, significant differences (p < 0.05) in either or both On.Ar and H.Ar were detected between FK-B-002, FK-B-003, and FK-B-006, and the following comparative specimens: Naumann’s elephant, fossil elephant, Asian elephant, human (five of seven specimens), Yabe’s giant deer, and cattle (one of five specimens) (Table 4). Additional significant differences were also observed between each of FK-B-002 and FK-B-003 and human (one of the remaining two specimens), as well as between FK-B-003 and human (one of the remaining specimens), leporid, sika deer (two of seven specimens), ancient sika deer (one of two specimens), reindeer, Japanese marten, dog, red fox, and brown bear (one of four specimens) (Table 4).
Discussion
Histomorphological features of the burned bone fragments observed in CT images and identification of animal taxa
The fact that osteons, Haversian canals, and plexiform bone structures could be observed in CT images of the burned bone fragments from Fukui Cave provides evidence that high-resolution X-ray CT allows for histomorphological identification of animal taxa without destructive sampling, even in small Pleistocene calcined bone fragments. However, in three of the seven specimens, the histological structures could not be clearly observed. In general, bones recovered from archaeological sites are likely to have been subjected to various biological, chemical, and physical damage during long-term deposition in the sediment34,73,74; such taphonomic alterations were probably imposed on the present specimens as well.
The On.Ar and H.Ar values of the burned bone fragments fell within the range observed in medium-sized artiodactyls such as wild boar and sika deer (Fig. 4). Therefore, these burned bone fragments were most plausibly derived from a medium-sized artiodactyl. The presence of plexiform bone identified in FK-B-007, a structure characteristic of artiodactyls, also supports this inference. However, the On.Ar and H.Ar values of wild boar, sika deer, and Japanese serow largely overlap (Table 2), making it impossible to identify the species within the medium-sized artiodactyl group based on this histomorphological method alone. Moreover, since the values are also close to those of some carnivores, the possibility that the specimens belong to these taxa cannot be completely ruled out.
Both On.Ar and H.Ar are known to vary within a single bone depending on the sampled region67,75, and a fragment does not always reflect the overall histomorphological characteristics of the entire bone76,77. Therefore, taxonomic identification based solely on histomorphometric observations requires caution. Even so, the measured values of the burned bone fragments differed markedly from those of proboscideans, Yabe’s giant deer, and humans (Tables 3 and 4), suggesting that the Fukui Cave specimens can be effectively excluded from these taxa.
Taken together, the present analysis indicates that (1) the burned bone fragments from Fukui Cave were most likely derived from a medium-sized artiodactyl, although the possibility of their origin from a carnivore cannot be ruled out; and (2) the possibility that they originated from proboscideans, Yabe’s giant deer, or humans can be considered highly unlikely.
Utilization of animal resources in the Japanese Archipelago during the terminal Palaeolithic
The mammalian fauna of the Japanese archipelago during the Late Pleistocene included species that still inhabit the archipelago today—such as medium-sized artiodactyls (wild boar, sika deer, and serow) and carnivores (bears, raccoon dogs, and foxes)—as well as extinct taxa including proboscideans (Naumann’s elephant and mammoths) and large artiodactyls (Yabe’s giant deer and bison). Because Palaeolithic sites yielding animal bones were long considered extremely rare in Japan, the limited number of finds led to the widespread image that humans of the time hunted large mammals such as Naumann’s elephant and Yabe’s giant deer28,78. This “big-game hunting” view has been disseminated through school textbooks and has become part of the general public’s understanding of the Palaeolithic period.
In recent years, several Palaeolithic sites have yielded faunal remains predominantly consisting of medium- to small-sized mammals such as cervids and hares18,21. In addition, analyses of lithic assemblages and studies on pitfall hunting have suggested that medium- and small-sized mammals were among the main hunting targets during the Palaeolithic period79,80, and thus a reconsideration of the traditional view that emphasizes the hunting of large game has been proposed19,81.
Regarding faunal transitions in the Late Pleistocene of the Japanese Archipelago, Naumann’s elephant and Yabe’s giant deer have traditionally been thought to have become extinct between ca. 20 and 10 ka BP30,32. However, Takahashi82 and Iwase et al.29, who compiled reliable AMS radiocarbon dates for large mammal fossils, demonstrated that no definitive ages later than ca. 25 ka BP are available for Naumann’s elephant and Yabe’s giant deer, and suggested that environmental changes associated with the onset of the Last Glacial Maximum (LGM) around 30 ka BP may have led to their extinction by approximately 25 ka BP. Furthermore, Sawada and Yoshinaga83, who reviewed major sites yielding faunal remains from the Incipient Jomon period (ca. 15–12 ka BP), confirmed that the taxonomic composition of the faunal assemblages from this period was broadly similar to that of the succeeding Jomon period (ca. 12–3 ka BP), indicating that large mammals had already disappeared by that time.
However, between ca. 25 and 15 ka BP, reliable AMS radiocarbon dates are extremely scarce not only for Naumann’s elephant and Yabe’s giant deer but also for medium- and small-sized mammals. In other words, the faunal composition of this period itself remains poorly understood, and to examine the faunal transition—including the extinction of large mammals—it is essential to accumulate additional faunal remains from this timespan.
Burned bone fragments from Fukui Cave, dated to approximately 16 ka BP, are important in filling this chronological gap. Although detailed taxonomic identification was not achieved in this study, the specimens were shown to possess morphological characteristics consistent with medium-sized artiodactyls and not with proboscideans or Yabe’s giant deer. This finding provides valuable clues for understanding both the extinction process of large mammals in the Japanese Archipelago and the nature of animal exploitation during the terminal phase of the Palaeolithic period.
Conclusions
The extinction of large mammals worldwide during the Late Pleistocene has generally been attributed to environmental changes associated with climatic fluctuations and human hunting activities. The relative contribution of these factors differed among regions84,85; hence, elucidating local processes in detail is crucial for reconstructing Quaternary natural history and understanding how humans first became involved in animal extinctions. While considerable progress has been made in studies addressing this issue in Europe, Siberia, Australia, and the Americas86,87,88,89,90,91, investigations of the relationship between human activities and faunal changes during the Late Pleistocene have lagged in the Japanese Archipelago, located at the easternmost end of Eurasia.
In this study, burned bone fragments from the terminal Palaeolithic layers of Fukui Cave were analyzed using high-resolution X-ray CT to investigate their histomorphological characteristics. The results indicated that these remains did not derive from large mammals that became extinct during the Late Pleistocene. Although the sample size was limited and the specimens may not represent the overall pattern of animal exploitation at the site, the findings contribute to understanding the extinction processes of large mammals in the Japanese Archipelago. The accumulation of such data is particularly significant in Japan, where faunal remains from Pleistocene human sites are extremely scarce.
Bone histomorphological identification still involves several methodological challenges, such as limited taxonomic resolution and insufficient comparative data by species and skeletal element. Nevertheless, in regions such as the Japanese Archipelago—where bone preservation at Pleistocene archaeological sites is generally poor—methods capable of extracting information from highly fragmented specimens are especially valuable. Non-destructive taxonomic identification based on histomorphological observation using high-resolution X-ray CT is promising, as it allows for the preservation and future utilization of materials and is expected to play an important role in subsequent research.
Materials and methods
Burned bone fragments from Fukui Cave
Seven small burned bone fragments (Specimen IDs: FK-B-001 to FK-B-007), presumed to be mammalian cortical bone, recovered from Layer 4 of Fukui Cave (ca. 16 ka BP), were subjected to CT scanning (Fig. 2; Table 5). These specimens are curated by the Board of Education of Sasebo City, where Fukui Cave is located, and the present study was conducted with the Board’s permission.
These fragments exhibited gray to white coloration due to burning. The color of bone changes with burning: low temperature burning results in a brown coloration, which turns black and then bluish or gray as the temperature rises, and eventually becomes white calcined bone7,8,92,93,94,95. The relationship between bone color and burning temperature varies with factors such as oxygen availability, combustion duration, and cortical bone thickness; therefore, when estimating burning temperature from color, it is necessary to consider these factors and infer an approximate temperature range. Absolonová et al.96 summarized many previous studies on the relationship between burning temperature and bone coloration, reporting that bones become gray at burning temperatures of 500–700 °C. Accordingly, the burned bone fragments recovered from Fukui Cave are considered to have been burned within this temperature range.
Additional analytical methods such as Fourier transform infrared spectroscopy (FTIR), Raman spectroscopy, and X-ray diffraction (XRD) could provide further information on burning temperatures7,10,11. However, these analyses were not performed because the specimens are extremely small and fragile, and destructive damage was a concern.
Synchrotron radiation X-ray CT imaging
SR X-ray CT imaging of the burned bone fragments excavated from Fukui Cave was performed at beamline BL20B2 of the large synchrotron radiation facility SPring-8. The imaging was conducted using monochromatic synchrotron radiation, with the X-ray energy set to 35 keV. Each specimen was mounted on a rotation stage with a custom holder and rotated 180° during image acquisition. The exposure time was 120 msec per projection, and the specimen -to-detector distance was set to 150 mm. A total of 1800 projections were acquired and reconstructed using the filtered back-projection (FBP) method. The resulting three-dimensional dataset had an isotropic voxel size of 2.74 μm. This resolution is considered sufficient for measuring the areas of osteons (approximately 10,000–30,000 μm²) and Haversian canals (approximately 200–1,200 μm²) in the burned bone fragments from Fukui Cave (see the Results section). The reconstructed images were converted to TIFF format and subsequently analyzed using ImageJ.
Observation and measurement of bone microstructure
For the seven burned bone fragments from Fukui Cave, osteons (secondary osteons) and plexiform bone were examined in the CT images. For fragments in which osteons were identifiable, the cross-sectional area of complete osteons (On.Ar) and the cross-sectional area of the Haversian canal within each osteon (H.Ar) were measured. Measurements were performed on CT slices obtained at the midpoint along the length of each fragment in the direction where the osteon cross-section could be observed. Osteon and Haversian canal contours were traced, and areas were calculated using ImageJ (National Institutes of Health, USA). All histomorphometrical data of the Fukui Cave specimens were collected by one of the authors (J.S.).
Comparative mammal bones
For comparative reference, histological sections of compact bone were taken from the midshaft of limb bones (humerus, radius, femur, and tibia) or the mandibular body of the following taxa: Naumann’s elephant (Palaeoloxodon cf. naumanni), unidentified Pleistocene elephant fossil (Elephantidae), Asian elephant (Elephas maximus), human (Homo sapiens), macaque (Macaca fuscata), leporid (Leporidae), wild boar (Sus scrofa), sika deer (Cervus nippon), ancient sika deer (Cervus praenipponicus), reindeer (Rangifer tarandus), Japanese serow (Capricornis crispus), Yabe’s giant deer (Sinomegaceros yabei), cattle (Bos taurus), bison (Bison bison), Japanese marten (Martes melampus), dog (Canis lupus), raccoon dog (Nyctereutes procyonoides), red fox (Vulpes vulpes), leopard (Panthera pardus), and brown bear (Ursus arctos). These animals represent terrestrial mammals that inhabited the Japanese Archipelago during the Late Pleistocene or Holocene31,32,33,97. Detailed information on specimen IDs, sample numbers, and attributes of the comparative materials is provided in Table 6.
Although no Late Pleistocene mammalian remains have been reported from the vicinity of Fukui Cave, fossils of Naumann’s elephant, macaque, ancient sika deer, and Yabe’s giant deer have been recovered from presumed Late Pleistocene deposits in limestone caves in the Hiraodai area, approximately 120 km from the site98. In addition, wild boar remains have been identified from the earliest Holocene layers of Fukui Cave99. As noted in the Introduction section, the excavated bone fragments were considered to originate from terrestrial mammals larger than small animals; therefore, small mammals such as rodents, moles, bats, and weasels were excluded from comparison.
For the comparative animal specimens, histological sections of compact bone were examined under transmitted and polarized light microscopy to record microstructural features. Micrographs were captured using a light microscope (Axio Imager.A1, Zeiss) equipped with a CMOS camera (AdvanCam-CTR20, AdvanVision). The cross-sectional areas of complete osteons and their Haversian canals were measured with ImageJ, following the procedures of Pfeiffer67 and Sawada et al.18,69. For some comparative animals, osteon and Haversian canal area data reported by Habill100, Martiniaková et al.65, Pfeiffer et al.68, and Sawada et al.18,101 were also used (Table 5).
Optical microscopy was used to observe histological structures in the comparative animal specimens instead of the CT imaging applied to the burned bones from Fukui Cave. Ideally, the same CT imaging should also have been applied to the comparative animal specimens; however, synchrotron-based CT image analysis requires substantial resources and limited access, making such validation difficult. Although CT and optical microscopy differ in image contrast and spatial resolution, the structures targeted in bone histomorphometric analysis (osteons and Haversian canals) are morphological features that can be observed using both methods. A previous study comparing histomorphometric measurements, including Haversian canal diameter, between µCT images of compact bone and microscopic images of histological sections reported no significant differences between the two methods102, supporting the validity of using optical microscopic images for comparison in the present study.
Histomorphological shrinkage of bone specimens due to burning
Calcination causes the loss of organic components, an increase in the crystallinity of bioapatite, and a reorganization of bone structure7,10,13,103,104. Therefore, the effects of these physicochemical changes must be taken into account when performing bone morphometric analysis.
It is well known that bones shrink when exposed to high temperatures105,106,107, although the degree of shrinkage varies depending on the burning temperature, duration, and the state of the bone during burning55,108. As described in the Materials section, the burned bone fragments from Fukui Cave exhibited a gray color, suggesting they were burned at approximately 500–700 °C.
Regarding the shrinkage of osteons and Haversian canals due to burning, Absolonová et al.96 measured the cross-sectional areas of osteons and Haversian canals in both unburned and burned bones, providing detailed data. Their results indicate that bones burned at 500 °C and 600 °C showed no shrinkage of osteons or Haversian canals. In bones burned at 700 °C, however, the osteon area decreased to 76.6% of the pre-burning value (mean osteon area before burning: 28,089.85 μm²; mean osteon area after burning at 700 °C: 21,504.71 μm²), and the Haversian canal area decreased to 76.4% (mean Haversian canal area before burning: 2,172.26 μm²; mean after burning: 1,659.02 μm²).
Based on this experimental results, our study assumed that the On.Ar and H.Ar of the burned bone fragments from Fukui Cave fell within a range defined by: (1) no change due to burning, i.e., the measured values equal the original values (corresponding to 500–600 °C), and (2) shrinkage to 75% of the original values (corresponding to approximately 700 °C). The upper bound of the burning temperature was set at 700 °C, which seems reasonable considering that Hanson and Cain8 noted the histological structure becomes indistinct in bones fired at higher temperatures, and Squires et al.95 reported that microstructural identification becomes difficult in bones exposed to temperatures above 900 °C.
The lower and upper bounds of On.Ar and H.Ar for the estimated burning temperature range were calculated using the following formulas:
(1) Lower bounds (assuming a burning temperature of 500–600 °C): no shrinkage, so the measured value equals the original value.
On.Ar (or H.Ar) before burning = actual measured value of On.Ar (or H.Ar).
(2) Upper bounds (assuming a burning temperature of 700 °C ): shrinkage due to burning, so the measured value equals 75% of the original value.
On.Ar (or H.Ar) before burning = actual measured value of On.Ar (or H.Ar) / 0.75.
Statistical analysis
For both On.Ar and H.Ar, differences in mean values were tested between the burned bone fragments from Fukui Cave and the comparative animal specimens. For the comparative specimens, only the data measured in the present study and those reported by Sawada et al.18 were used. Considering the shrinkage caused by burning in the Fukui Cave fragments, two sets of comparisons were performed: one assuming no shrinkage and the other assuming the maximum estimated shrinkage due to burning. As the assumption of equal variances was not met for either On.Ar or H.Ar, the Games–Howell test was applied. The significance level was set at 0.05, and all statistical analyses were conducted using SPSS Statistics (IBM).
Ethics statement
This study did not involve living humans or animals. The primary materials analyzed were archaeological faunal remains dating to approximately 16,000 years ago. Additionally, the comparative specimens used in this study were obtained from the permanent osteological collections of the Institute of Physical Anthropology, Niigata University of Health and Welfare. All specimens were used in accordance with the institutional guidelines for the use of archival biological materials. No animals were specifically sacrificed for this research.
Data availability
All data supporting the findings of this study are available at Zenodo: https://zenodo.org/records/17688116.
References
Hayashi, K. The Fukui microblade technology and its relationships in Northeast Asia and North America. Arct. Anthropol. 5, 128–190 (1968).
Japanese Archaeological Association Ed. Nihon no Doketsu Iseki (Heibonsya, 1967).
Kobayashi, T., Kaner, S. & Nakamura, O. Jomon Reflections: Forager Life and Culture in the Prehistoric Japanese Archipelago (Oxbow Books, 2004). http://books.casematepublishers.com/Jomon_Reflections.pdf
Takahashi, R., Toizumi, T. & Kojo, Y. Archaeological studies of Japan: current studies of the Jomon archaeology. Nihon Kokogaku. 5, 47–72 (1998). https://doi.org/10.11215/nihonkokogaku1994.5.47
Agency for Cultural Affairs, Government of Japan. Bunka Shingikai no Toshin (2024). https://www.bunka.go.jp/koho_hodo_oshirase/hodohappyo/pdf/94070501_01.pdf
Sasebo City Board of Education Ed. The Fukui Cave: Excavation of Paleolithic and Jomon Cultural Layers in Kyushu, Western Japan (Primex, 2016).
Gallo, G. et al. Characterization of structural changes in modern and archaeological burnt bone: Implications for differential preservation bias. PLoS ONE. 16, e0254529 (2021). https://doi.org/10.1371/journal. pone.0254529
Hanson, M. & Cain, C. R. Examining histology to identify burned bone. J. Archaeol. Sci. 34, 1902–1913 (2007). https://doi.org/10.1016/j.jas.2007.01.009
Stiner, M. C., Kuhn, S. L., Weiner, S. & Bar-Yosef, O. Differential burning, recrystallization, and fragmentation of archaeological bone. J. Archaeol. Sci. 22, 223–237 (1995). https://doi.org/10.1006/jasc.1995.0024
van Hoesel, A., Reidsma, F. H., van Os, B. J. H., Megens, L. & Braadbaart, F. Combusted bone: Physical and chemical changes of bone during laboratory simulated heating under oxidising conditions and their relevance for the study of ancient fire use. J. Archaeol. Sci. Rep. 28, 102033 (2019). https://doi.org/10.1016/j.jasrep.2019.102033
Gallo, G., Ushakov, S. V., Navrotsky, A. & Stahlschmidt, M. C. Impact of prolonged heating on the color and crystallinity of bone. Archaeol. Anthropol. Sci. 15, 143 (2023). https://doi.org/10.1007/s12520-023-01842-0
Reidsma, F. H. Laboratory-based experimental research into the effect of diagenesis on heated bone: implications and improved tools for the characterisation of ancient fire. Sci. Rep. 12, 17544 (2022). https://doi.org/10.1038/s41598-022-21622-5
Reidsma, F. H., van Hoesel, A., van Os, B. J. H., Megens, L. & Braadbaart, F. Charred bone: Physical and chemical changes during laboratory simulated heating under reducing conditions and its relevance for the study of fire use in archaeology. J. Archaeol. Sci. Rep. 10, 282–292 (2016). https://doi.org/10.1016/j.jasrep.2016.10.001
Ono, A., Sato, H., Tsutsumi, T. & Kudo, Y. Radiocarbon dates and archaeology of the Late Pleistocene in the Japanese islands. Radiocarbon 44, 477–494 (2002). https://doi.org/10.1017/S0033822200031854
Tsutsumi, T. Archaeology of the Japanese Archipelago: Retto no Kokogaku: Kyusekki-jidai (Kawade Shobo Shinsha, 2011).
Japanese Palaeolithic Research Association. Palaeolithic Sites of the Japanese Archipelago Database (2024). https://jpra-db.github.io/Japanese_Palaeolithic_Sites_DB_English/
Inada, T. Zetsumetsu dobutsu to Nihon-retto no kyusekki-jidai jin. Kagaku 68, 345–352 (1998).
Sawada, J., Nara, T., Fukui, J., Hirata, K. & Dodo, Y. Histomorphological species identification of tiny bone fragments from a Paleolithic site in the Northern Japanese Archipelago. J. Archaeol. Sci. 46, 270–280 (2014). https://doi.org/10.1016/j.jas.2014.03.025
Sawada, J. Kyusekki-jidai no dobutsu-kokogaku wo meguru syomondai. In: Tohoku Nihon no Kyusekki-jidai (ed The Society for the Study of Paleolithic Culture in Northeastern Japan) 35–46 (Rokuichi Syobo, 2018).
Nara, T., Watanabe, T., Sawada, J., Sawaura, R. & Sato, T. (eds) Shitsukari Abe Cave Vol. 1 (Rokuichi Shobo, 2015).
Sawaura, R., Sawada, J., Sato, T., Suzuki, T. & Sasaki, K. Late Pleistocene hares of the Japanese archipelago: Paleobiogeographic implication at the Last Glacial Maximum. Int. J. Osteoarchaeol. 28, 179–187 (2018). https://doi.org/10.1002/oa.2645
Kato, S. Iwateken Hanaizumi kaseki-sho syutsudo no jinrui ihin. Gekkan Bunkazai 138, 12–21 (1975).
Matsumoto, H., Mori, H., Marui, K. & Ozaki, H. On the discovery of the Upper Pliocene fossiliferous and culture-bearing bed at Kanamori, Hanaizumi Town, Province of Rikuchu. Bull. Nat. Sci. Mus. (Tokyo). 4, 287–324, plates 25–48 (1959).
Kanagawa Archaeology Foundation. Yoshioka Iseki-gun IV: Kyusekki-jidai 2, Jomon-jidai 1 (Kanagawa Archaeology Foundation Research Report, 1997).
Yamaoka, T. Shizuoka-ken Hamamatsu-shi Kita-ku Gyoja-ana cave site dai 2 ji chosa. In: Kokogaku kenkyu-shitsu Chosa Kenkyu Shuho 2022 3–8 (Midori Bijutsu, 2023).
Yamaoka, T., Niimi, M., Fukuchi, A., Harumura, A. & Toyama, M. Shizuoka-ken Hamamatsu-shi Hamana-ku Gyoja-ana cave site dai 3 ji chosa. In: Kokogaku kenkyu-shitsu Chosa Kenkyu Shuho 2024 3–10 (Midori Bijutsu, 2025).
Nojiri-ko Excavation Research Group. The Late Quaternary environment around Lake Nojiri in central Japan. In: Proceedings of the 29th International Geological Congress Part B 269–277 (1994).
Nojiri-ko Excavation Research Group. Saishu Hyoki no Shizen to Jinrui (Kyoritsu Shuppan, 1997).
Iwase, A., Hashizume, J., Izuho, M., Takahashi, K. & Sato, H. Timing of megafaunal extinction in the late Late Pleistocene on the Japanese Archipelago. Quat. Int. 255, 114–124 (2012). https://doi.org/10.1016/j.quaint.2011.03.029
Kamei, T., Taruno, H. & Kawamura, Y. Implication of mammal fauna for the Quaternary geohistory of the Japanese Islands. Daiyonki Kenkyu 26, 293–303 (1988). https://doi.org/10.4116/jaqua.26.3_293
Kawamura, Y. Quaternary mammalian faunas in the Japanese Islands. Daiyonki Kenkyu 30, 213–220 (1991). https://doi.org/10.4116/jaqua.30.213
Kawamura, Y. Last glacial and Holocene land mammals of the Japanese Islands: their fauna, extinction and immigration. Daiyonki Kenkyu 46, 171–177 (2007). https://doi.org/10.4116/jaqua.46.171
Kawamura, Y. & Nakagawa, R. Terrestrial mammal faunas in the Japanese Islands during OIS 3 and OIS 2. In: Environmental Changes and Human Occupation in East Asia during OIS3 and OIS2, BAR International Series 2352 (eds Ono, A. & Izuho, M) 33–86 (Archaeopress, 2012).
Lyman, R. L. Vertebrate Taphonomy (Cambridge University Press, 1994).
Cattaneo, C. et al. Determining the human origin of fragments of burnt bone: a comparative study of histological, immunological and DNA techniques. Forensic Sci. Int. 102, 181–191 (1999). https://doi.org/10.1016/S0379-0738(99)00059-6
Cattaneo, C., Porta, D., Gibelli, D. & Gamba, C. Histological determination of the human origin of bone fragments. Forensic Sci. Int. 54, 531–533 (2009). https://doi.org/10.1111/j.1556-4029.2009.01000.x
Crescimanno, A. & Stout, S. D. Differentiating fragmented human and nonhuman long bone using osteon circularity. J. Forensic Sci. 57, 287–294 (2012). https://doi.org/10.1111/j.1556-4029.2011.01973.x
Greenlee, D. M. & Dunnell, R. C. Identification of fragmentary bone from the Pacific. J. Archaeol Sci. 37, 957–970 (2010). https://doi.org/10.1016/j.jas.2009.11.029
Morales, J. P., Roa, H. I., Zavando, D. & Suazo, G. I. Determination of the species from skeletal remains through histomorphometric evaluation and discriminant analysis. Int. J. Morphol. 30, 1035–1041 (2012). https://doi.org/10.4067/S0717-95022012000300044
Mulhern, D. M. & Ubelaker, D. H. Differentiating human from nonhuman bone microstructure. In: Bone Histology: An Anthropological Perspective (eds Crowder, C. M. & Stout, S. D.) 109–134 (CRC Press, 2012).
Felder, A. A. et al. Secondary osteons scale allometrically in mammalian humerus and femur. R Soc. Open. Sci. 4, 170431 (2017). https://doi.org/10.1098/rsos.170431
Zedda, M. & Babosova, R. Does the osteon morphology depend on the body mass? A scaling study on macroscopic and histomorphometric differences between cow (Bos taurus) and sheep (Ovis aries). Zoomorphology 140, 169–181 (2021). https://doi.org/10.1007/s00435-021-00516-6
Harsányi, L. Differential diagnosis of human and animal bone. In: Histology of Ancient Human Bone: Methods and Diagnosis (eds Grupe, G. & Garland, A. N.) 79–94 (Springer-Verlag, 1993).
Jowsey, J. Studies of Haversian systems in man and some animals. J. Anat. 100, 857–864 (1966).
Mulhern, D. M. & Ubelaker, D. H. Differences in osteon banding between human and nonhuman bone. J. Forensic Sci. 46, 220–222 (2001). https://doi.org/10.1520/JFS14952J
Brits, D., Steyn, M. & ́Abbé, L. A histomorphological analysis of human and non-human femora. Int. J. Legal Med. 128, 369–377 (2014). https://doi.org/10.1007/s00414-013-0854-3
Currey, J. D. Differences in the blood-supply of bone of different histological types. Quart. J. Microsc Sci. 101, 351–370 (1960). https://doi.org/10.1242/jcs.s3-101.55.351
Enlow, D. H. & Brown, S. O. A comparative histological study of fossil and recent bone tissues, Part II. Tex. J. Sci. 9, 186–214 (1957).
Enlow, D. H. & Brown, S. O. A comparative histological study of fossil and recent bone tissues, Part III. Tex. J. Sci. 10, 187–230 (1958).
Locke, M. Structure of long bones in mammals. J. Morphol. 262, 546–565 (2004). https://doi.org/10.1002/jmor.10282
Mori, R. et al. Comparative histology of the laminar bone between young calves and foals. Cells Tissues Organs. 175, 43–50 (2013). https://doi.org/10.1159/000073436
Ricqlès, A. Recherches paléohistologiques sur les os longs des tétrapodes VII, Sur la classification, la signification fonctionnelle et l’histoire des tissus osseux des tétrapodes. Première partie Ann. Paléontol. 61, 51–129 (1975).
Veitschegger, K., Kolb, C., Amson, E., Scheyer, T. M. & Sánchez-Villagra, M. R. Palaeohistology and life history evolution in cave bears, Ursus spelaeus sensu lato. PLOS one. 13, e0206791 (2018). https://doi.org/10.1371/journal.pone.0206791
Caccia, G. et al. Histological determination of the human origin from dry bone: a cautionary note for subadults. Int. J. Legal Med. 130, 299–307 (2016). https://doi.org/10.1007/s00414-015-1271-6
Correia, P. M., Horocholyn, K. & Pointer, K. The use of histology to distinguish animal from human burnt bone with reference to some limitations. In: Burnt Human Remains: Recovery, Analysis, and Interpretation (eds Ellingham S., Adserias-Garriga, J., Zapico, S.C. & Ubelaker, D.H.) 241–271 (John Wiley & Sons Ltd., 2023).
Cuijpers, S. .M. Distinguishing between the bone fragments of medium-sized mammals and children. A histological identification method for archaeology. Anthropol. Anz. 67, 181–203 (2009). https://doi.org/10.1127/0003-5548/2009/0021
Cuijpers, A. G. F. M. Histological identification of bone fragments in archaeology: telling humans apart from horses and cattle. Int. J. Osteoarchaeol. 16, 465–480 (2006). https://doi.org/10.1002/oa.848
Dittmann, K., Grupe, G., Manhart, H., Peters, J. & Strott, N. Histomorphometry of mammalian and avian compact bone. In: Microscopic Examinations of Bioarchaeological Remains: Keeping a Close Eye on Ancient Tissues, Documenta Archaeobiologiae, vol. 4 (eds Grupe, G. & Peters, J.) 49–101 (Verlag Marie Leidorf, 2006).
Dominguez, V. M. & Crowder, C. M. The utility of osteon shape and circularity for differentiating human and non-human Haversian bone. Am. J. Phys. Anthropol. 149, 84–91 (2012). https://doi.org/10.1002/ajpa.22097
Hillier, M. & Bell, L. S. Differentiating human bone from animal bone: a review of histological method. J. Forensic Sci. 52, 249–263 (2007). https://doi.org/10.1111/j.1556-4029.2006.00368.x
Larentis, O. et al. Transformed by fire: A ritual practice dating back to the Early Neolithic in Italy. Interdisciplinary analysis of burnt bone remains in Lugo di Grezzana (Veneto), 5000 – 4850 cal BCE. Archaeol. Anthropol. Sci. 17, 206 (2025). https://doi.org/10.1007/s12520-025-02326-z
Robbins, G. & Gray, K. What remains: species identification and bone histology. In: An Archaeology of Desperation: Exploring the Donner Party’s Alder Creek Camp (eds Dixon, K. J., Schablitsky, J. M. & Novak, S. A.) 163–184 (University of Oklahoma Press, 2011).
Imaizumi, K., Taniguchi, K. & Ogawa, Y. DNA survival and physical and histological properties of heat-induced alterations in burnt bones. Int. J. Legal Med. 128, 439–446 (2014). https://doi.org/10.1007/s00414-014-0988-y
McGrath, K. et al. Identifying archaeological bone via non-destructive ZooMS and the materiality of symbolic expression: Examples from Iroquoian bone points. Sci. Rep. 9, 11027 (2019). https://doi.org/10.1038/s41598-019-47299-x
Martiniaková, M. et al. Histological study of compact bone tissue in some mammals: a method for species determination. Int. J. Osteoarchaeol. 17, 82–90 (2007). https://doi.org/10.1002/oa.856
Martiniaková, M., Grosskopf, B., Omelka, R., Vondráková, M. & Bauerová, M. Differences among species in compact bone tissue microstructure of mammalian skeleton: use of a discriminant function analysis for species identification. J. Forensic Sci. 51, 1235–1239 (2006). https://doi.org/10.1111/j.1556-4029.2006.00260.x
Pfeiffer, S. Variability osteon size in recent human populations. Am. J. Phys. Anthropol. 106, 219–227 (1998).
Pfeiffer, S., Crowder, C., Harrington, L. & Brown, M. Secondary osteon and Haversian canal dimensions as behavioral indicators. Am. J. Phys. Anthropol. 131, 460–468 (2006).
Sawada, J., Kondo, O., Nara, T., Dodo, Y. & Akazawa, T. Bone histomorphology of Dederiyeh Neanderthal child. Anthropol. Sci. 112, 247–256 (2004). https://doi.org/10.1537/ase.00094
Watanabe, S., Miyamoto, N., Sakurada, M., Sawada, J. & Ueno, Y. Forensic discrimination between burned human and animal bone fragments using SR X-ray CT. SPring-8/SACLA Res. Rep. 12, 275–279 (2024). https://doi.org/10.18957/rr.12.5.275
Bradfield, J. Identifying animal taxa used to manufacture bone tools during the Middle Stone Age at Sibudu, South Africa: results of a CT-rendered histological analysis. PLOS one. 13, e0208319 (2018). https://doi.org/10.1371/journal. pone.0208319
Andronowski, J. M., Davis, R. A. & Stephan, H. E. Inferring bone attribution to species through micro-Computed Tomography: A comparison of third metapodials from Homo sapiens and Ursus americanus. J. Forensic Radiol. Imaging 18, 11–17 (2019). https://doi.org/10.1016/j.jofri.2019.08.001
O’Connor, T. The Archaeology of Animal Bones (Texas A & M University Press, 2000).
Reitz, E. J. & Wing, E. S. Zooarchaeology, Second Edition (Cambridge University Press, 2008).
Nguyen, J. T. & Barak, M. M. Secondary osteon structural heterogeneity between the cranial and caudal cortices of the proximal humerus in white-tailed deer. J. Exp. Biol. 223, jeb225482 (2020). https://doi.org/10.1242/jeb.225482
Lagacé, F., Schmitt, A., Martrille, L., Benassi, J. & Adalian, P. Using histomorphometry for human and nonhuman distinction: a test of four methods on fresh and archaeological fragmented bones. Forensic Sci. Int. 313, 110369 (2020). https://doi.org/10.1016/j.forsciint.2020.110369
Ubelaker, D. H. & Wu, Y. Fragment analysis in forensic anthropology. Forensic Sci. Res. 5, 260–265 (2020). https://doi.org/10.1080/20961790.2020.1811513
Ohtaishi, N. Kyusekki iseki no ichi to shuryo-ju no kisetsu ido ruto ni kansuru kosatsu. Daiyonki Kenkyu 29, 287–289 (1990).
Inada, T. Sentoki bunka no shutsugen to kyusekki teki sekki seisaku no kaitai. Kokogaku Kenkyu 15, 3–18 (1969).
Sano, T. Evidence for the use of the bow-and-arrow technology by the first modern humans in the Japanese islands. J. Archaeol. Sci. Rep. 10, 130–141 (2016). https://doi.org/10.1016/j.jasrep.2016.09.007
Tsutsumi, T. Nihon-retto kyusekki-jidai no sutereotaipu na ogata-ju shuryo kasetsu heno gimon. Archa Kenkyu Ronsyu 6, 16–24 (2024).
Takahashi, K. The formative history of the terrestrial mammalian fauna of the Japanese Islands during the PlioPleistocene. Kyusekki Kenkyu 3, 5–14 (2007).
Sawada, J. & Yoshinaga, A. Kamikuroiwa iwakage to Jomon Sosoki no dobutsu riyo. Bessatsu Kikan Kokogaku 34, 32–35 (2020).
Barnosky, A. D., Koch, P. L., Feranec, R. S., Wing, S. L. & Shabel, A. B. Assessing the Causes of Late Pleistocene Extinctions on the Continents. Science 306, 70–75 (2004). https://doi.org/10.1126/science.1101476
Lorenzen, E. et al. Species-specific responses of Late Quaternary megafauna to climate and humans. Nature 479, 359–364 https://doi.org/10.1038/nature10574 (2011).
Dehasque, M. et al. Combining Bayesian age models and genetics to investigate population dynamics and extinction of the last mammoths in northern Siberia. Quat. Sci. Rev. 259, 106913 (2021). https://doi.org/10.1016/j.quascirev.2021.106913
Haynes, G. (ed) American Megafaunal Extinctions at the End of the Pleistocene (Springer, 2009).
O’Keefe, F. R. et al. Pre–Younger Dryas megafaunal extirpation at Rancho La Brea linked to fire-driven state shift. Science 381, 746 (2023). https://doi.org/10.1126/science.abo3594
Stuart, A. J., Kosintsev, P. A., Higham, T. F. G. & Lister, A. M. Pleistocene to Holocene extinction dynamics in giant deer and woolly mammoth. Nature 431, 684–689 (2004). https://doi.org/10.1038/nature02890
Surovell, T. A., Pelton, S. R., Anderson-Sprecher, R. & Myers, A. D. Test of Martin’s overkill hypothesis using radiocarbon dates on extinct megafauna. PNAS 113, 886–891 (2016). https://doi.org/10.1073/pnas.1504020112
Wroe, S. et al. Climate change frames debate over the extinction of megafauna in Sahul (Pleistocene Australia-New Guinea). PNAS 110, 8777–8781 (2013). https://doi.org/10.1073/pnas.1302698110
Carroll, E. L. & Smith, M. Burning questions: Investigations using field experimentation of different patterns of change to bone in accidental vs deliberate burning scenarios. J. Archaeol. Sci. Rep. 20, 952–963 (2018). https://doi.org/10.1016/j.jasrep.2018.02.001
Nicholson, R. A. A morphological investigation of burnt animal bone and an evaluation of its utility in archaeology. J. Archaeol. Sci. 20, 411–428 (1993). https://doi.org/10.1006/jasc.1993.1025
Shipman, P., Foster, G. & Schoeninger, M. Burnt bones and teeth: An experimental study of colour, morphology, crystal structure and shrinkage. J. Archaeol. Sci. 11, 307–325 (1984). https://doi.org/10.1016/0305-4403(84)90013-X
Squires, K. E., Thompson, T. J. U., Islam, M. & Chamberlain, A. The application of histomorphometry and Fourier Transform Infrared Spectroscopy to the analysis of early Anglo-Saxon burned bone. J. Archaeol. Sci. 38, 2399–2409 (2011). https://doi.org/10.1016/j.jas.2011.04.025
Absolonová, K., Dobisíková, M., Beran, M., Zocová, J. & Velemínský, P. The temperature of cremation and its effect on the micro- structure of the human rib compact bone. Anthropol. Anz. 69, 439–460 (2012). https://doi.org/10.1127/0003-5548/2012/0213
Ohdachi, S. D., Ishibashi, Y., Iwasa, M. A. & Saitoh, T. The Wild Mammals of Japan (Shoukadoh, 2009).
Kawamura, Y. & Sotsuka, T. Preliminary report on the Quaternary mammalian remains from several caves on the Hiraodai Plateau, Fukuoka Prefecture, northern Kyushu, Japan. Bull. Kitakyushu Mus. Nat. Hist. 5, 163–188 (1984). https://doi.org/10.34522/bkmnh.5.0_163
Uzawa, K.,Toizumi, T. & Kurozumi, T. Report on the analysis of animal remains from the Fukui Cave excavation. In:The FukuiCave: Excavation of Paleolithic and Jomon Cultural Layers in Kyushu, Western Japan(ed. Sasebo City Board ofEducation) 275–282 (Primex, 2016).
Havill, L. M. Osteon remodeling dynamics in Macaca mulatta: normal variation with regard to age, sex, and skeletal maturity. Calcif. Tissue Int. 74, 95–102 (2004). https://doi.org/10.1007/s00223-003-9038-3
Sawada, J. et al. Histomorphological discrimination between human and nonhuman bones of fragmentary osteal remains: Analyses of burnt bones from the ancient Heian site in the northern Tohoku district, Japan. Anthropol. Sci. (Jpn Ser). 118, 23–36 (2010). https://doi.org/10.1537/asj.118.23
Particelli, F. et al. A comparison between micro-CT and histology for the evaluation of cortical bone: effect of polymethylmethacrylate embedding on structural parameters. J. Microsc. 245, 302–310 (2012). https://doi.org/10.1111/j.1365-2818.2011.03573.x
Gallo, G., Aldeias, V. & Stahlschmidt, M. Revisiting the thermal alteration of buried bone. J. Archaeol. Sci. Rep. 63, 105080 (2025). https://doi.org/10.1016/j.jasrep.2025.105080
Marques, M. P. M. et al. Profiling of human burned bones: oxidising versus reducing conditions. Sci. Rep. 11, 1361 (2021). https://doi.org/10.1038/s41598-020-80462-3
Fairgrieve, S. I. Forensic Cremation Recovery and Analysis (CRC Press, 2008).
Nelson, R. A microscopic comparison of fresh and burned bone. J. Forensic Sci. 37, 1055–1060 (1992). https://doi.org/10.1520/JFS13291J
Thompson, T. J. U. Heat-induced dimensional changes in bone and their consequences for forensic anthropology. J. Forensic Sci. 50, 1008–1015 (2005). https://doi.org/10.1520/JFS2004297
Ellingham, S. & Sandholzer, M. A. Determining volumetric shrinkage trends of burnt bone using micro-CT. J. Forensic Sci. 65, 196–199 (2020). https://doi.org/10.1111/1556-4029.14150
Acknowledgements
We are grateful to Prof. Takuya Yamaoka of Shizuoka University and Prof. Takashi Nara of Niigata University of Health and Welfare for their advice and help writing this manuscript.
Funding
This work was supported by JSPS KAKENHI Grant Numbers 18K18533 and 24K00160.
Author information
Authors and Affiliations
Contributions
JS conceived and designed the study. YY collected the materials. JS, MY, KU (Uzawa), and TT organized the materials. JS, KU (Uesugi), MH, SW, and NM performed SR X-ray CT imaging of the materials at SPring-8. JS, RA, and FS conducted histomorphological analyses. MY carried out the dating analysis. JS drafted the initial manuscript. All authors contributed to manuscript preparation and approved the final version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sawada, J., Yoneda, M., Uesugi, K. et al. Non-destructive histomorphological identification of Late Pleistocene burned bone fragments using synchrotron radiation X-ray CT at SPring-8. Sci Rep 16, 13908 (2026). https://doi.org/10.1038/s41598-026-50208-8
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-50208-8
